
Abstract
Nonalcoholic fatty liver disease (NAFLD) is considered the hepatic manifestation of metabolic syndrome. The more clinically concerning form of the disease, nonalcoholic steatohepatitis (NASH), is characterized by steatosis, lobular inflammation, and ballooning degeneration. Here we describe a naturally occurring syndrome in the common marmoset that recapitulates the pathologic findings associated with NAFLD/NASH in humans. Hepatomegaly determined to result from NAFLD was observed in 33 of 183 marmosets. A comprehensive histopathologic assessment performed in 31 marmosets demonstrated that NAFLD was characterized by variably sized, Oil Red O staining cytoplasmic vacuoles and observed primarily in animals with evidence of obesity and insulin resistance. A subset of marmosets (16 of 31) also demonstrated evidence of NASH characterized by multifocal inflammation combined with ballooning hepatocellular degeneration. Marmosets with NASH demonstrated an increase in immunostaining with an antibody targeted against the human leukocyte antigens (HLA)–DP, HLA-DQ, and HLA-DR compared with marmosets without NASH (38.89 cells/10× field vs 12.05 cells/10× field, P = .05). In addition, marmosets with NASH demonstrated increased Ki-67 immunopositive cellular proliferation compared with those without (5.95 cells/10× field vs 1.53 cells/10× field, P = .0002). Finally, animals with NASH demonstrated significantly increased mean circulating serum iron levels (160.47 μg/dl, P = .008) and an increase in numbers of Prussian blue–positive Kupffer cells (9.28 cells/40× field, P = .005) relative to marmosets without NASH (97.75 μg/dl and 1.87 cells/40×, respectively). This study further characterizes the histopathology of NAFLD/NASH and suggests that the marmoset may be a valuable animal model with which to investigate the host and environmental factors contributing to the progression of NAFLD/ NASH.
Keywords
Nonalcoholic fatty liver disease (NAFLD) is considered the hepatic manifestation of the metabolic syndrome and is characterized by a progressive accumulation of lipid within hepatocytes. This is a significant public health concern that affects up to 30% of the US population and is the leading cause of chronic liver disease. 23 Obesity and type 2 diabetes are both major risk factors, and NAFLD prevalence rates of 75% to 100% are observed in obese patients. 11 The pathogenesis of hepatic lipid accumulation remains unresolved but is likely influenced by a number of factors, including increased caloric intake, increased liver fat synthesis, leptin resistance, and insulin resistance. 17 The interaction between genetic and environmental factors is thought to play a critical role.
While NAFLD is a relatively benign condition, progression to nonalcoholic steatohepatitis (NASH), observed in 20% to 30% of patients with NAFLD, is the more aggressive form of the disease and results in significant morbidity and mortality. 31 NASH is characterized by the presence of inflammation, fibrosis, and hepatocellular necrosis. In the most severe state, NASH is associated with cirrhosis and an increased risk of hepatocellular carcinoma. There is a critical need to determine host or environmental factors involved in the progression from NAFLD to NASH, and animal models are useful in these endeavors. This transition from NAFLD to the more clinically concerning NASH likely involves disruption of multiple compensatory pathways within the liver. 31 One hypothesis suggests that excess levels of hepatic iron result in oxidative stress and subsequent hepatocellular damage, inflammation, and fibrosis. 10 An alternate and more widely accepted hypothesis suggests that aberrant cytokine expression and an excessive inflammatory response serve as significant contributing factors to the progression of NAFLD to NASH.30,31
Unfortunately, many rodent models fail to recapitulate one or more features of the human disease.17,18,20 A nonhuman primate model would have benefits over rodent models, including a closer genetic relationship to humans and more analogous physiology. During preventative health assessments of common marmosets housed at the New England Primate Research Center (NEPRC), profound hepatic enlargement and histologic lesions of NAFLD were observed in a number of animals. Furthermore, in captive housed callitrichids, hepatic siderosis as a consequence of iron overload is one of the most common postmortem findings. 28 We hypothesized that exposure to excess dietary iron may be one environmental factor contributing to NAFLD/NASH observed in common marmosets. To further investigate these findings and to examine the contribution of dietary iron to liver disease, we describe a retrospective study of 31 marmosets to evaluate the association between increased hepatic mass, NAFLD, and NASH.
Methods
Study Design and Population
The study was based on marmosets housed at the NEPRC in accordance with all applicable regulations and in a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International. Animals were maintained under an animal holding and breeding protocol or experimental protocols approved by Harvard Medical School’s Standing Committee on Animals.
To determine the prevalence of palpable hepatomegaly in the NEPRC marmoset colony, a total of 183 animals (n = 102 females, 81 males) that ranged in age from 1.0 to 13.8 years were assessed during routine preventive health care. Breeding females were not assessed to exclude potential confounding due to pregnancy. All animals had morphometric data collected (described below) as part of routine colony management. Radiographs were performed on a subset of animals, and 4 animals were subjected to liver biopsies (see supplemental materials) to determine the cause of hepatomegaly.
A retrospective analysis of necropsy records was performed to identify a study population of 31 subjects. All recruited subjects were humanely euthanized within the past 6 years for study-related end points or nonhepatic clinical conditions. Only those animals with sufficient clinical data and hepatic tissue collected at necropsy were included. Older archived samples were not considered for inclusion. Animals were assigned to groups based on hepatic mass and NAFLD Activity Score. Animals were characterized as having increased hepatic mass if the hepatic weight collected at necropsy exceeded the median for the cohort (21.3 g) or normal hepatic mass if the terminal hepatic weight was below this cut point. Terminal hepatic weight was not recorded for 2 animals. The second independent variable examined was the likelihood of NASH as determined by the NAFLD Activity Score (described below). Animals were considered as having NASH if the Activity Score was ≥4, based on the system used in humans. 25
Fifteen animals had been fed standard commercial chow (Harlan Teklad New World Primate Chow 8791, Teklad Diets, Madison, WI and Zupreme Canned Marmoset Diet, Zupreme, Shawnee, KS). Eight animals had been fed a commercially available high-fat diet containing 20% added saturated fat (Laboratory Diets, Richmond, IN). A third group of 8 animals had been fed a commercially available diet enriched in glucose (Mazuri Callitrichid Diet 5MI5; PMI Nutrition, Henderson, CO). The specific composition of these diets has been published previously. 36 Dietary supplements such as fruits, vegetables, seeds, and hard-boiled eggs were provided to all subjects.
Morphometrics and Clinical Chemistry
Total body mass was obtained by weighing animals on an electronic scale. Each animal was assigned a body condition score of 1 to 5 based on palpation of body contour and adipose deposits as follows: 1, emaciated; 2, thin; 3, optimum body composition; 4, heavy; and 5, obese. Fat and lean mass were quantified with an EchoMRI quantitative magnetic resonance analyzer (QMR; Echo Medical Systems, Houston, TX).
Serum chemistry data for the retrospective cohort were mined from the sample database maintained by the contracted veterinary diagnostic laboratory (IDEXX Laboratories, North Grafton, MA). Parameters examined include aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), γ-glutamyltransferase (GGT), serum triglycerides, serum cholesterol, and blood glucose. Frozen plasma collected within 6 months of euthanasia was available from 23 of 31 animals, allowing measure of insulin levels, serum iron, and total iron binding capacity (TIBC). Hemoglobin A1c (HgbA1c) was measured in whole blood using the DCA 2000+ HbA1c analyzer (Siemens Diagnostics, Tarrytown, NY). Data most proximate to the time of death were used for this measure and were available for 19 of 31 subjects (see supplemental materials for more information).
Necropsy and Tissue Collection
A complete necropsy examination was performed on all subjects in the retrospective cohort. Representative sections of all major organs were collected, fixed in 10% neutral buffered formalin, and processed routinely. Fresh samples of liver were both flash frozen for chemical analysis or collected in Tissue-Tek O.C.T. (Sakura Finetek USA, Inc., Torrance, CA) compound and flash frozen at –80°C for histologic analysis.
Histologic Evaluation of Liver
Liver sections stained with hematoxylin and eosin were reviewed in a blinded fashion by one pathologist (J.A.K.) unaware of the clinical data associated with each subject. Sections were evaluated for the degree of steatosis (scale of 0–3), lobular inflammation (scale of 0–3), and hepatocellular ballooning (scale of 0–2) as described by Kleiner et al. 16 NAFLD Activity Score was calculated as the unweighted sum of the recorded scores (scale of 0–8). 16 The degree of fibrosis was examined in Mallory trichrome-stained slides. Prussian blue–stained slides were evaluated to investigate hepatic iron load, and slides were scored semi-quantitatively using the method described by Rowe et al 24 (see supplemental materials).
Immunohistochemistry
Immunohistochemistry for CD3 (rabbit polyclonal; Dako North America, Carpinteria, CA), Ki-67 (mouse monoclonal, clone MIB-1; Dako North America), IBA1 (rabbit polyclonal; Wako Chemicals, USA, Richmond, VA), and human leukocyte antigen (HLA)–DR, HLA-DP, and HLA-DQ (mouse monoclonal, clone CR3/43; Dako North America) was carried out using an ABC immunostain technique on formalin-fixed, paraffin-embedded tissues as previously described. 9 Additional information is supplied in the supplemental materials.
Quantitative Image Analysis
Full details are available in the supplemental methods. Briefly, 3 representative photomicrographs were obtained from each immunohistochemically stained slide and imported into commercially available quantitative image analysis software (SigmaScan Pro; Systat Software, San Jose, CA), and the number of positively stained cells was counted using threshold and intensity gating. The average number of cells in each field was then averaged over the 3 sections and results reported as total number of cells/10× field. Prussian blue–stained hepatic macrophages were enumerated in 10 representative sections photographed at 40× magnification. Images were imported into ImageJ (National Institutes of Health [NIH], Bethesda, MD), and manual counts were performed in a blinded fashion. 26
Frozen samples were stained with Oil Red O (ORO). To quantify the amount of fat stained with ORO, two 40× images per slide were analyzed with SigmaScan Pro. A color threshold was used to highlight areas of stained lipid, and the total ORO-stained area was then calculated. This was divided by the overall area of the image being analyzed to determine a percent of total area occupied by lipid. The ORO staining process did not always highlight an entire lipid droplet due to processing artifact. For example, the center of a large droplet may have remained unstained. Because of this, the reported percentages are likely underestimations of the total area represented.
Quantification of Hepatic Lipid and Glycogen
Hepatic triglycerides were measured from 100 mg of fresh-frozen liver tissue in a subset of subjects (n = 16 of 31) using a Triglycerides Reagent Set (cat. T7532; Pointe Scientific, Inc., Canton, MI). Hepatic glycogen was measured from 10 mg of fresh-frozen liver tissue in the same subset of animals using a Glycogen Assay Kit (cat. K646; BioVision, Inc., Milpitas, CA). Further detail is available in the supplemental material.
Quantification of Hepatic Trace Elements
Fresh-frozen hepatic tissue samples (n = 24 of 31; weight range, 60–210 g) were assayed for trace element concentration by the Animal Health Diagnostic Center at the Cornell University College of Veterinary Medicine (Ithaca, NY) using inductively coupled plasma atomic emissions spectroscopy. Concentration was reported as parts per million (ppm). Elements evaluated included hepatic iron, copper, manganese, magnesium, zinc, and molybdenum.
Statistical Analysis
Statistical analysis was performed using commercially available statistical software (Stata Software; StataCorp LP, College Station, TX). Dependent variables examined included demographic, morphometric, biochemical, and histologic parameters as well as select trace elements measured in serum and hepatic tissue. Sex and diet were considered as possible confounders and were examined for relationships with independent and dependent variables. Because the study was retrospective, data on all parameters were not collected for all subjects. The Shapiro-Wilk test was used to determine normal or nonnormal distribution of data. Statistical comparisons between cases and controls were performed with the 2-tailed t test for normally distributed continuous parameters, Mann-Whitney test for nonnormally distributed continuous parameters, and Fisher exact test for categorical parameters. Correlations were calculated using the nonparametric Spearman correlation. P values of less than .05 were considered significant.
Results
Clinical Detection of Hepatomegaly
During routine health assessments, an increased prevalence of organomegaly was observed in colony-housed marmosets fed a standard diet. Organomegaly was detected by palpation of the right upper abdominal quadrant and confirmed as hepatic enlargement by radiography in a subset of animals (Fig. 1).
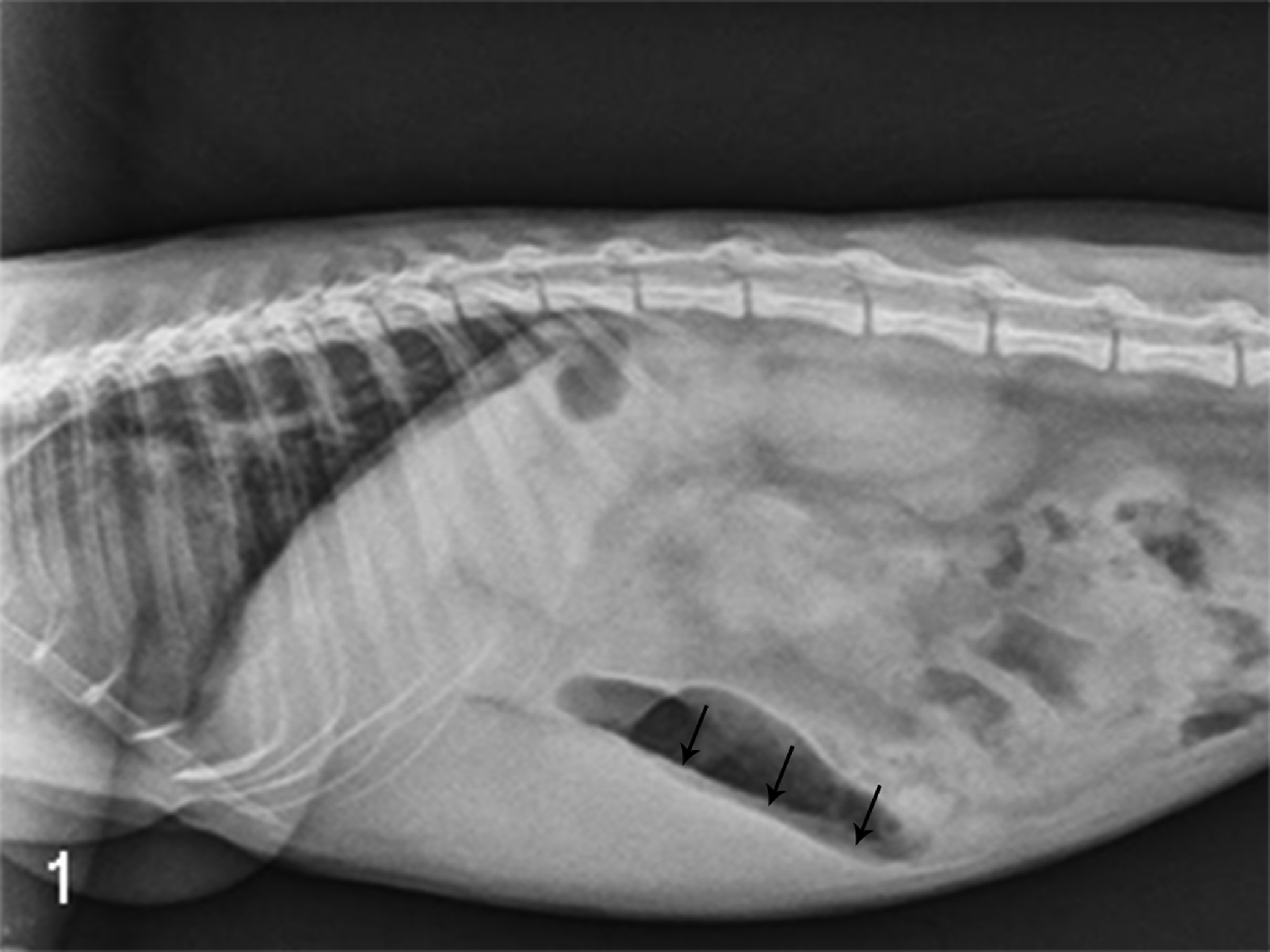
Radiograph from a common marmoset with palpable hepatomegaly. Radiograph demonstrates a liver margin (arrows) that extends beyond the ribs.
Palpable hepatomegaly was detected in 33 of 183 (18%) animals. Histologically, liver biopsies from 2 of 4 animals had macrovesicular vacuolar hepatopathy consistent with hepatic steatosis. The lesions were characterized by variable, multifocal, random hepatocellular enlargement due to accumulation of large, clear cytoplasmic vacuoles resulting in nuclear margination, affecting 25% and 80% of hepatocytes in these 2 animals. Biopsies from the other 2 marmosets demonstrated diffuse, microvesicular vacuolar hepatopathy characterized by enlargement of hepatocytes by numerous small cytoplasmic vacuoles that contributed to a reticular appearance of the cytoplasm, consistent with glycogen or microvesicular lipid deposition.
Morphometric measures were compared between marmosets with palpable hepatomegaly and those without. Marmosets in which hepatomegaly was detected by palpation had higher body condition scores (median score, 4.5; range, 3.5–5) compared with those without (median score, 3.5; range, 2.5–4.5; Mann-Whitney, P < .00001). Correspondingly, marmosets with hepatomegaly had an increased total body mass (491.36 g [SD 57.18]) compared with marmosets without hepatomegaly (404.75 g [SD 47.95], t test, P < .00001). A trend toward increased prevalence of hepatomegaly was observed in females relative to males (69.7% and 30.3% of marmosets with hepatomegaly, respectively), although this difference was not statistically significant (Fisher exact, P = .06). Mean age did not differ between animals with and without hepatomegaly (t test, P = .68).
Association of Hepatic Mass With Morphometry and Serum Chemistry Parameters
Because of the association between clinically palpable hepatomegaly and obesity, we performed a retrospective survey to further characterize this phenotype. A group of 31 subjects, comprising 13 males and 18 females ranging in age from 2 to 12 years (mean age, 5.68 years [SD 3.20]), was selected. Fourteen animals demonstrated an increased hepatic mass, and 15 animals had normal hepatic mass. Terminal liver weights were not available from 2 animals. Select biochemical parameters and measures of body composition were compared between animals with increased and normal hepatic mass (Table 1). Diet (Fisher exact, P = .26) and sex (Fisher exact, P = .71) were excluded as confounders as there was no association between these parameters and the presence or absence of increased hepatic mass. As expected, animals with increased hepatic mass demonstrated an obese phenotype with increased mean body mass (t test, P = .003) and increased mean fat mass (Mann-Whitney, P = .02). Significant differences in lean mass were not observed.
Demographic, Morphometric, and Biochemical Measures Associated With the Presence or Absence of Increased Hepatic Mass.
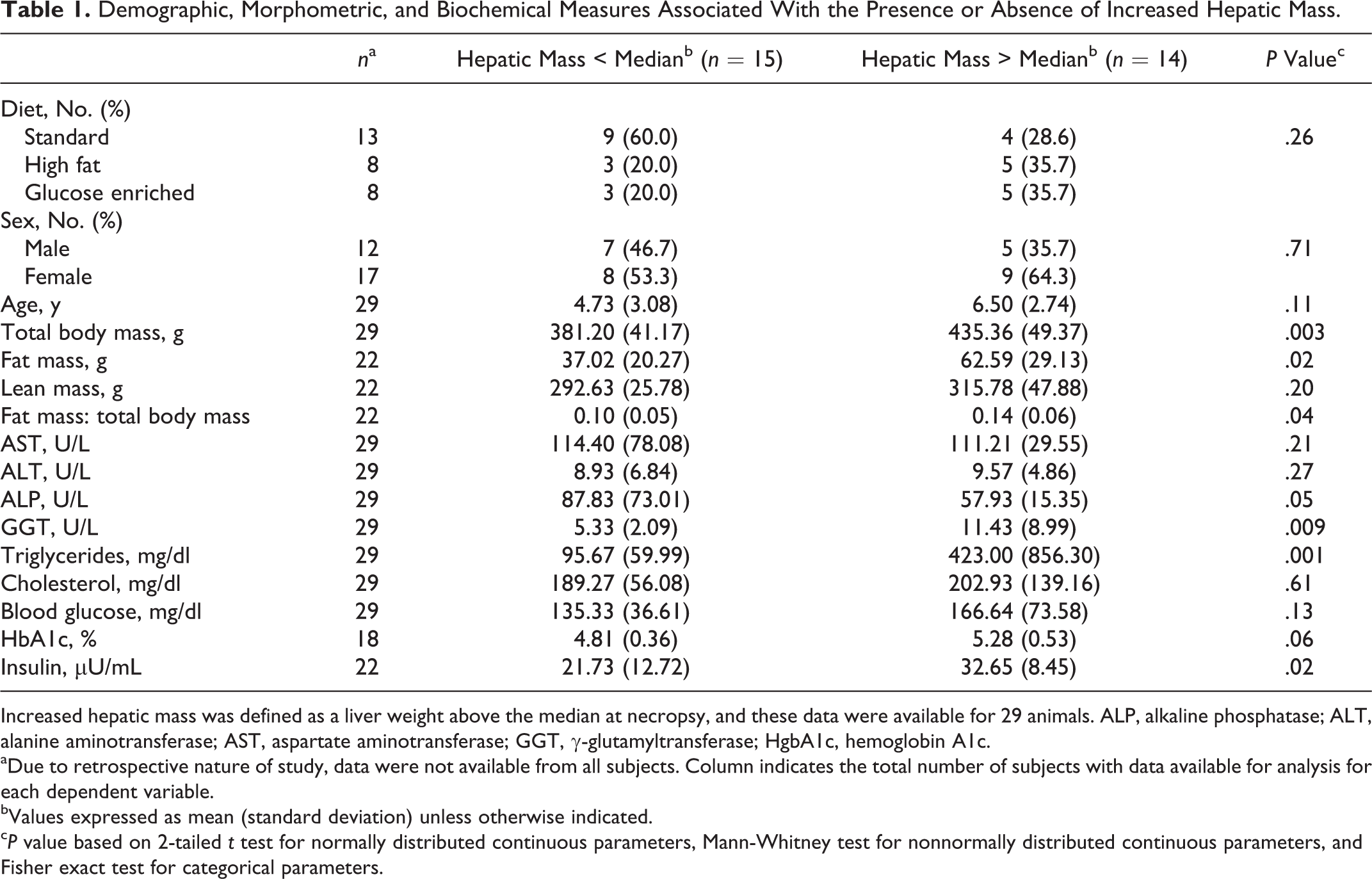
Increased hepatic mass was defined as a liver weight above the median at necropsy, and these data were available for 29 animals. ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; GGT, γ-glutamyltransferase; HgbA1c, hemoglobin A1c.
aDue to retrospective nature of study, data were not available from all subjects. Column indicates the total number of subjects with data available for analysis for each dependent variable.
bValues expressed as mean (standard deviation) unless otherwise indicated.
c P value based on 2-tailed t test for normally distributed continuous parameters, Mann-Whitney test for nonnormally distributed continuous parameters, and Fisher exact test for categorical parameters.
Animals with increased hepatic mass had increased mean GGT (Mann-Whitney, P = .009) and reduced mean ALP (Mann-Whitney, P = .05) compared with those with lower hepatic mass but no differences in ALT and AST. Animals with increased hepatic mass also had increased mean serum triglycerides (Mann-Whitney, P = .001), increased mean plasma insulin levels (t test, P = .02), but comparable blood glucose and HgbA1c.
Association of Hepatic Mass With Histologic Alterations
Semi-quantitative evaluation of histologic sections using the Kleiner scoring system 16 revealed some degree of steatosis (greater than 5% hepatic parenchymal involvement) in 23 of 31 animals (74.2%), with 11 of 31 (35.5%) demonstrating steatosis that affected greater than 66% of the hepatic parenchyma. Of the 8 animals with no observed steatosis, 1 had increased hepatic mass and 7 did not. Of the 23 animals with steatosis, 13 had increased hepatic mass and 8 did not (liver weight was not available from 2 animals). There was a significant association between steatosis and increased hepatic mass (Fisher exact, P = .04), but lipid deposition occasionally occurred in the absence of gross hepatic enlargement. Animals with lipid deposition and no evidence of hepatic enlargement tended to have a lesser degree of steatosis (2 animals with <33% parenchymal involvement and 4 animals with 33%–66% involvement).
Steatosis was characterized by variably sized, well-defined cytoplasmic vacuoles that compressed and peripheralized hepatocellular nuclei. Steatosis was most prominent in centrilobular hepatocytes and faded closer to portal tracts (Fig. 2). In most animals, the lipid was macrovesicular, but microvesicular steatosis was also seen. Vacuoles were confirmed to be lipid using ORO staining of frozen sections of liver (Fig. 3). Glycogen was also seen with periodic acid–Schiff (PAS) stain but did not appear to be the primary contributor to the vacuolar hepatopathy. Overall, microscopic findings were consistent with a diagnosis of NAFLD.3–7,19
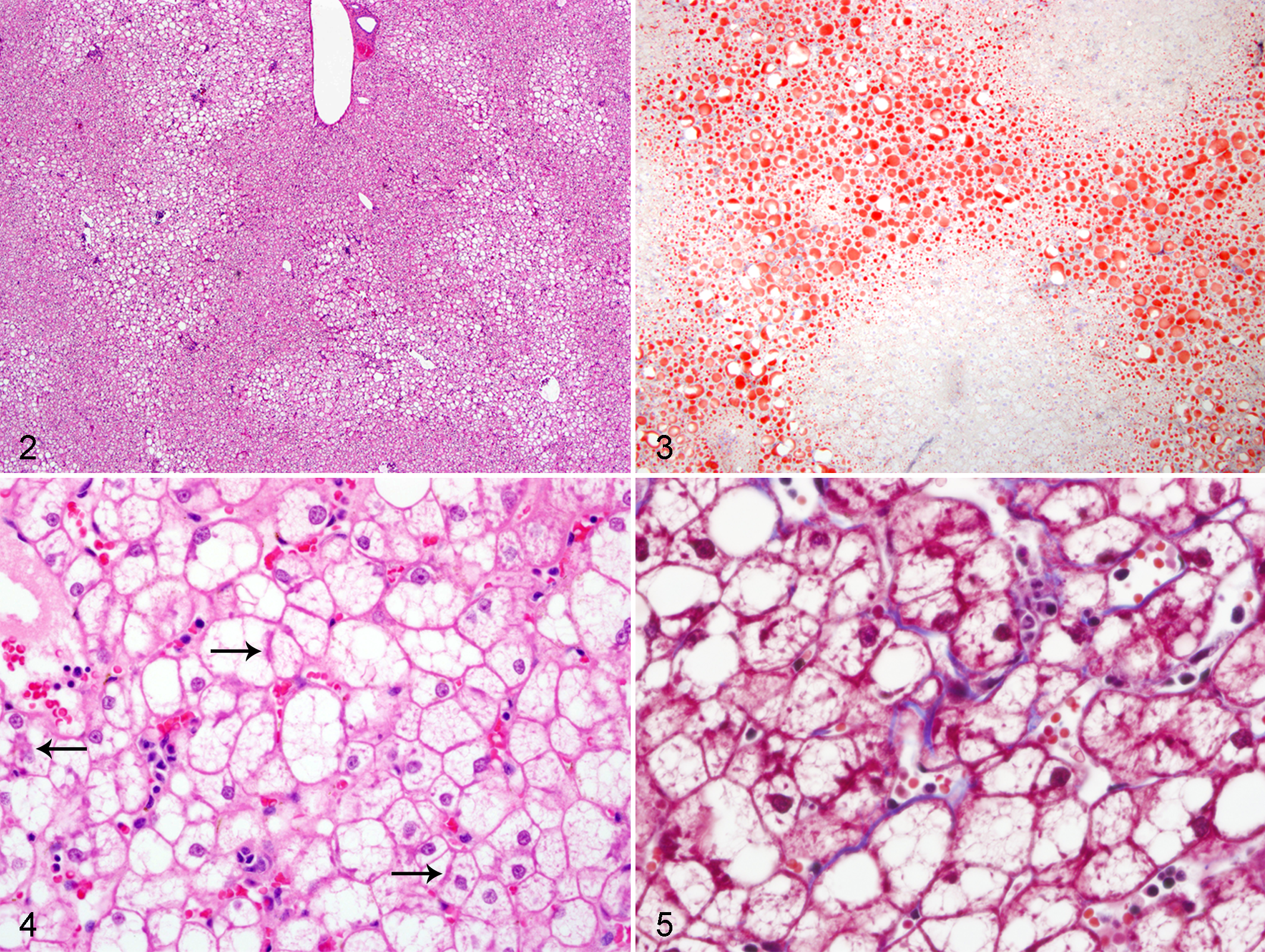
Steatosis was variably associated with a mixed inflammatory cell infiltrate. There was an absence of inflammatory foci in 7 of 31 (22.6%) animals, none of which had increased hepatic mass. Inflammation was minimal in 13 of 31 animals (41.9%), moderate in 8 of 31 (25.8%), and marked in 3 of 31 (9.7%). Infiltrates were predominantly composed of lymphocytes and macrophages, with fewer neutrophils. Inflammation ranged from pinpoint foci to locally extensive areas. Several animals had ballooning hepatocellular degeneration and Mallory-Denk bodies associated with steatosis and inflammation (Fig. 4). Pericellular fibrosis was rarely observed with trichrome staining but was seen in 2 animals with the most severe steatosis (Fig. 5).
Association of NASH With Hepatic Lipid and Serum Measures of Hepatic Injury
Based on an NAFLD Activity Score of ≥4, 16 of 31 animals were diagnosed with NASH. Diet (Fisher exact, P = .73) and sex (Fisher exact, P = .28) were excluded as confounders as there was no association between these parameters and Activity Score. Animals with NASH demonstrated a significant increase in mean GGT (Mann-Whitney, P = .006) (Table 2), ALT (Mann-Whitney, P = .01), and AST (Mann-Whitney, P = .04) but no difference in ALP.
Biochemical and Hepatic Histologic Findings Stratified by Diagnosis of Nonalcoholic Steatohepatitis (NASH) as Determined by the Nonalcoholic Fatty Liver Disease Activity Score (NAS).
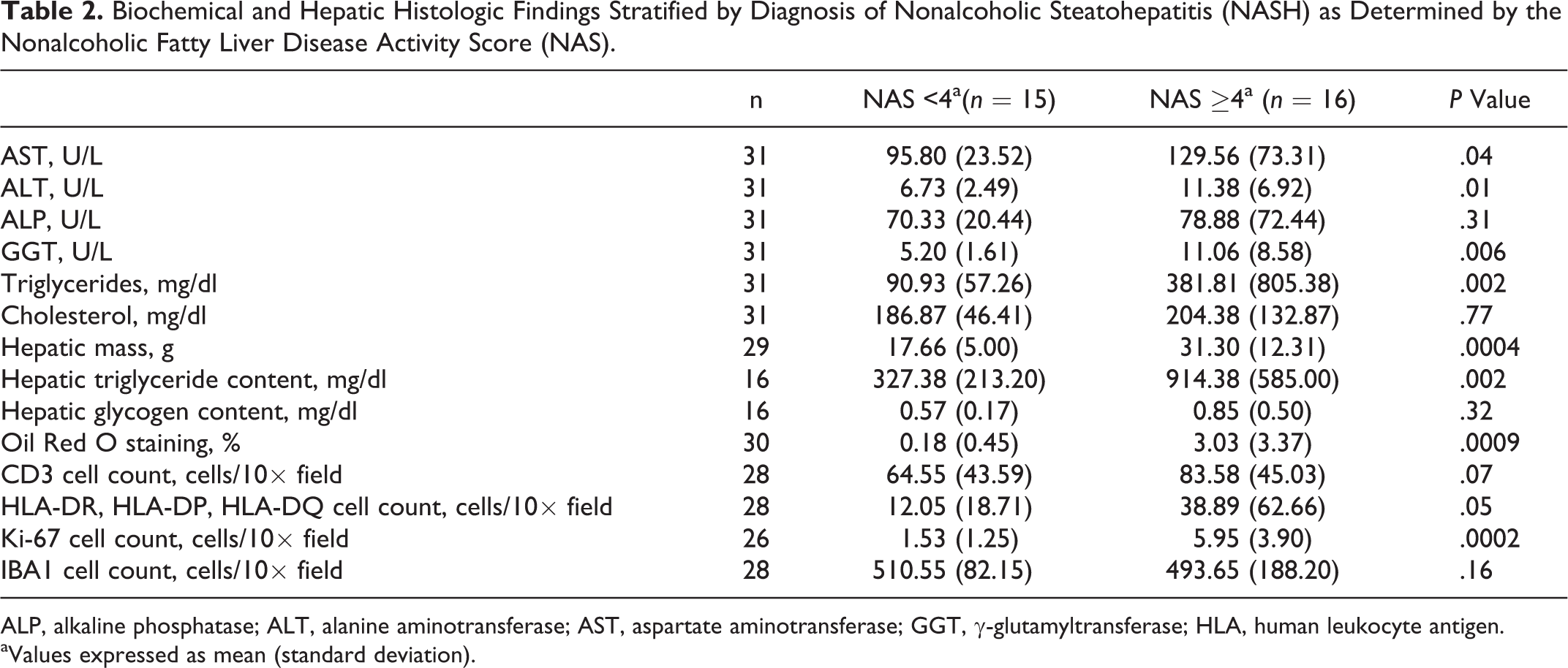
ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; GGT, γ-glutamyltransferase; HLA, human leukocyte antigen.
aValues expressed as mean (standard deviation).
Marmosets with NASH had a greater mean hepatic triglyceride content (Mann-Whitney, P = .002, n = 16; Table 2) and lipid deposition (based on percent ORO staining; Mann-Whitney, P = .0009, n = 30) relative to those without NASH. There was also excellent agreement between hepatic triglyceride measures and percent ORO staining (r = 0.89, P < .00001). Hepatic glycogen content was minimal and not significantly different between animals with and without NASH.
Phenotyping of Hepatic Leukocytes
Phenotyping of macrophages and lymphocytes in marmoset livers was performed using 4 markers: CD3 (T cells); HLA-DR, HLA-DP, and HLA-DQ (B cells, macrophages, and activated T lymphocytes); IBA1 (macrophages); and Ki-67 (cellular proliferation) (Table 2). Marmosets with NASH had a nonsignificant increase in CD3-positive cells (Mann-Whitney, P = .07) (Suppl. Figs. S1, S2); a marginally significant increase in immunostaining for HLA-DR, HLA-DP, and HLA-DQ (Mann-Whitney, P = .05) (Suppl. Figs. S3, S4); and no significant difference in IBA1-positive cells (Suppl. Figs. S7, S8). Ki-67 immunostaining, found predominantly in cells morphologically consistent with lymphocytes, was increased in animals with NASH (Mann-Whitney, P = .0002) (Suppl. Figs. S5, S6).
Hepatic and Serum Micronutrient Measures
Serum and tissue iron levels are represented in Table 3. Animals with NASH demonstrated a significantly increased mean circulating serum iron level relative to animals without NASH (Mann-Whitney, P = .008) but no difference in TIBC or hepatic iron load. Hepatic copper levels were lower in marmosets with NASH than in those without (t test, P = .03).
Select Trace Elements in Serum and Hepatic Tissue Stratified by Nonalcoholic Fatty Liver Disease Activity Score (NAS).
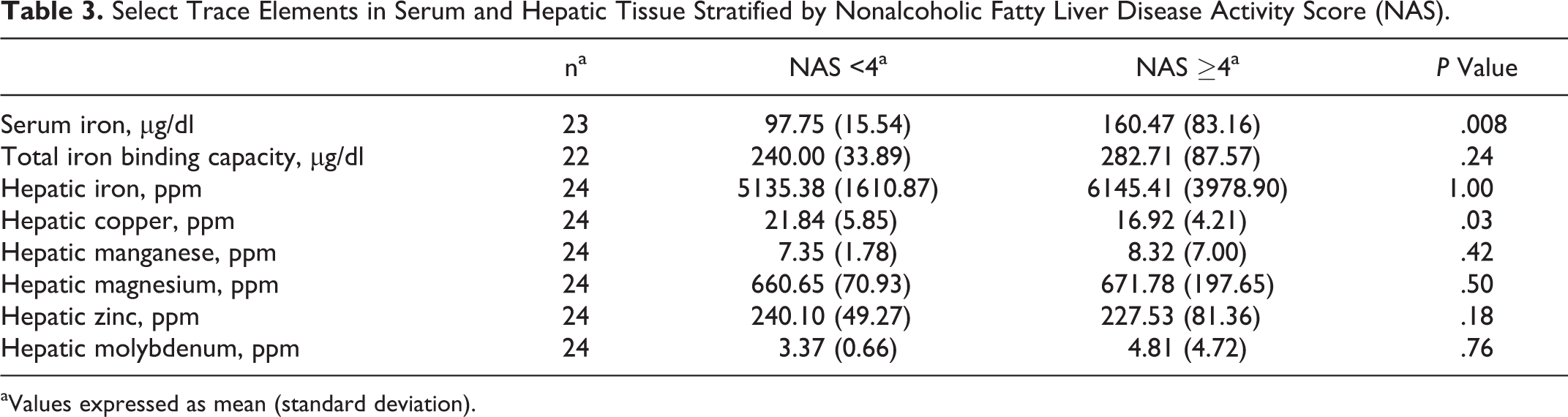
aValues expressed as mean (standard deviation).
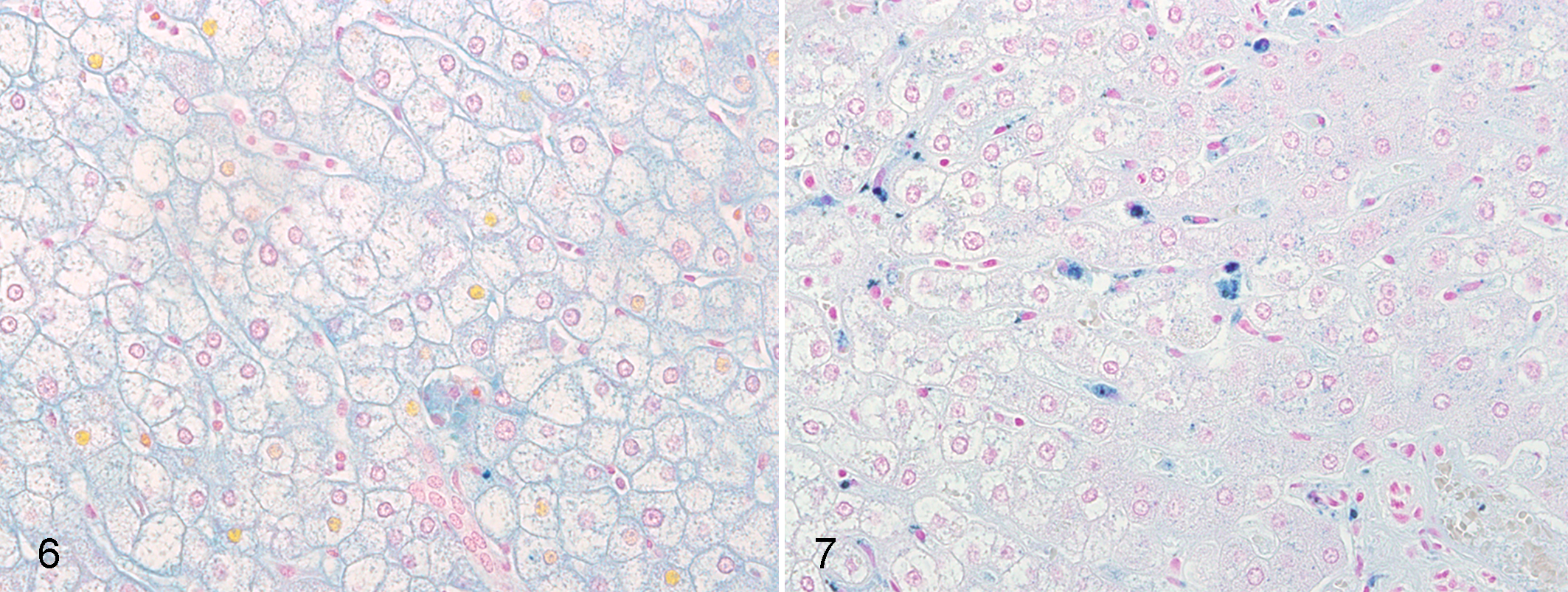
Rowe scoring of Prussian blue–stained hepatic sections was performed to assess for parenchymal iron load. Five of 16 marmosets (31.3%) with NASH had a Rowe score of 3. In contrast, none of the animals without NASH had a Rowe score greater than 2. Only 5 of 16 (31.3%) marmosets with NASH had a normal Rowe score of 1 or less, in contrast to 10 of 15 (66.7%) marmosets without NASH. However, these differences were not significant (Fisher exact, P = .06).
Kupffer cells were the primary cell type demonstrating positive staining. The number of Prussian blue–positive Kupffer cells for animals with NASH averaged 9.28 cells/40× field (SD 12.32) in contrast to 1.87 cells/40× field (SD 3.23) for those without NASH (Mann-Whitney, P = .005; Figs. 6, 7).
Discussion
This study demonstrated that marmosets develop hepatic steatosis that recapitulates NAFLD as reported in humans. In the marmoset, this condition is associated with hepatic enlargement and, like people, is more commonly reported in obese animals. The frequency of clinically palpable hepatomegaly in the NEPRC marmoset colony suggests a prevalence rate of at least 18%. However, nearly one-third of animals with histologically demonstrated hepatic lipid deposition did not have clinical evidence of increased hepatic mass, suggesting that the colony prevalence rate of NAFLD is likely far greater.
Approximately 50% of the marmosets demonstrated a progression to NASH characterized by the presence of steatosis, lobular inflammation, and ballooning degeneration. This is slightly higher than the 20% to 30% prevalence rate of NASH observed in humans with NAFLD but suggests that this natural disease state in marmosets may be used to elucidate mechanisms contributing to the development of NASH from NAFLD.
As in humans, both NAFLD and NASH were associated with alterations in serum biochemical markers. In marmosets with increased hepatic mass and those diagnosed with NASH, there was evidence of biliary stasis suggested by a 2-fold increase in serum GGT levels. Not only was this increase statistically significant, but it was outside of the reported reference range for this species. This finding is similar to alterations in GGT reported in humans with NAFLD and NASH. 13 ALP levels were not different in marmosets with and without NASH. The discrepancy in the measures of GGT and ALP may be related to species differences in the sensitivity and specificity of these 2 enzymes for the detection of cholestasis. Both groups had significant and marked increases in serum triglycerides well above the normal reference range. 37 Serum GGT and triglycerides may therefore be useful biochemical markers for NAFLD and NASH and serve as a screening tool in addition to thorough abdominal palpation. Animals diagnosed with NASH also had significant increases in the hepatic leakage enzymes AST and ALT, suggesting a degree of direct hepatocellular injury occurring in this group that was not seen in animals with increased hepatic mass but without NASH. While these alterations were statistically significant, ALT and AST remained within the reference range for this species. 37
In humans, NAFLD is associated with insulin resistance. 32 Serum blood glucose and glycosylated hemoglobin were insignificantly elevated in animals with increased hepatic mass compared with those without. There was, however, a significant increase in plasma insulin. This suggests that, although marmosets with increased hepatic mass did not have evidence of overt diabetes, animals were producing more insulin perhaps as a compensatory mechanism for end-organ insulin resistance.
This study suggests that NASH was associated with increased hepatic lipid deposition and increased inflammation. Both absolute hepatic triglyceride measurement and quantitative assessment of the percent ORO staining confirmed the finding that lipid deposition was significantly greater in animals with a diagnosis of NASH relative to those without. These results also suggested that quantitative microscopy is a valid surrogate technique to measure hepatic lipid in this model. Animals with NASH had increased immunoreactivity for HLA-DP–, HLA-DQ–, and HLA-DR–positive cells. This is consistent with inflammation, perhaps due to hepatocellular damage secondary to steatosis, and suggests that additional B-cell and activated T-cell markers should be examined. Ki-67 immunoreactivity was also increased in animals with NASH. While these findings are less specific, Ki-67 immunoreactivity has been shown to increase in both mouse models of steatohepatitis and in humans with the disease, 35 consistent with hepatocellular remodeling and increased inflammation.
Hepatic hemosiderosis as a consequence of iron overload is one of the most common postmortem findings in captive-housed callitrichids. 28 Despite refinements to commercial monkey chow, the levels of dietary iron provided remain high (approximately 18 mg/d for a 400-g marmoset compared with the recommended 8–18 mg/d for an adult human). The situation may be exacerbated by ascorbic acid supplementation that is thought to enhance iron absorption and the lack of access to natural tannins, which serve to inhibit iron absorption. 29 These tannins are obtained from leaves, barks, and fruits present within natural environments. To date, provision of these tannins has not been feasible in captive settings.
Several mechanisms are suggested to explain the role for iron in the pathogenesis of NASH. Excess iron catalyzes the production of reactive oxygen species, facilitates lipid peroxidation, and results in direct hepatocellular damage. Lipid peroxidation contributes to the generation of products such as malondialdehyde and 4-hydroxynonenal that activate hepatic stellate cells and induce fibrogenesis. 10 Finally, excess iron within hepatocytes activates Kupffer cells, allowing for inflammatory cell recruitment. 38 Epidemiologic studies in human populations have demonstrated associations among hepatic iron load, hemochromatosis gene (HFE) mutations, and the severity of hepatic fibrosis of NASH.2,15,33 Iron reduction therapy via phlebotomy has proven clinically useful for reducing fasting blood glucose, circulating insulin levels, and liver function tests in the presence of NAFLD.12,34 Despite this evidence, the relationship between iron metabolism and NASH remains controversial. Independent studies have failed to confirm the association between hepatic iron level, HFE mutation, and severity of liver damage.8,39 Because of these conflicting results, it is unknown if increased iron load associated with progressive NAFLD/NASH is a primary driver of disease or a secondary feature that nonetheless exacerbates the syndrome.
Our data demonstrated that serum iron load and number of iron-laden Kupffer cells are significantly increased in marmosets with NASH. As in humans, there was a corresponding reduction in hepatic copper levels as well. When considered with results showing similar numbers of IBA1 immunoreactive macrophages, our results suggest that hepatic iron accumulation may be limited to parenchymal macrophage populations and not due to increased histiocytic inflammation. There is clearly an association between iron levels and steatohepatitis. Future studies should include prospective examinations using diets with different iron levels to confirm that dietary iron load contributes directly to hepatic injury and studies to elucidate the coordination between copper and iron bioavailability in this model.
In addition to the findings, it is important to mention the limitations of this study. Due to the retrospective nature of this project, data were not collected for all subjects, and the resulting reduced sample size may limit the ability to detect differences between groups. A second limitation is related to the classification of study groups based on the median terminal hepatic weight. Eight animals had liver weights below the median despite histologic evidence of steatosis. It is unknown if steatosis without hepatic enlargement results in similar alterations in glucose and lipid metabolism as those observed in marmosets with steatosis and increased hepatic mass. Misclassification of these 8 animals may have resulted in an underestimation of the differences between study groups, potentially masking alterations in glucose and diminishing the magnitude of changes in insulin, triglycerides, and GGT.
The common marmoset model has inherent advantages compared with other nonhuman primate models and rodent models to investigate the pathogenesis of NAFLD/NASH. Marmosets are small nonhuman primates that are easily bred and maintained in captivity. They have a relatively short life span compared with other nonhuman primates (10–15 years) and reach sexual maturity by 18 months of age. 1 Interestingly, marmosets give birth to twins, which are bone marrow chimeras of one another. This can be leveraged when designing experiments to limit variability between control and experimental groups. They carry fewer zoonotic diseases than old-world nonhuman primates.21,22 Compared with rodents, marmosets’ cytochrome p450 enzyme structure more closely mimics that of humans. 27 The marmoset genome has recently been sequenced, allowing for analysis of possible genetic predispositions for NAFLD/NASH. 14 However, there are also disadvantages to experimental work using this animal model. While antibodies, assays, and other experimental resources are readily available for mice, rats, and rhesus macaques, there are fewer commercially available options for marmosets, although this is improving.
Factors resulting in the transition from NAFLD to NASH remain elusive, and animal models of the disease are necessary to determine the exact pathogenesis. The natural onset of this syndrome and epidemiology comparable to that observed in human populations suggest that the marmoset can serve as a valuable model with which to perform prospective and interventional studies. Future work should focus on further characterizing the disease and on dietary or environmental mechanisms that cause a more rapid progression to NASH.
Footnotes
Acknowledgement
We thank Audra Hachey, Heather Knight, and Chris Pearson for assistance with various immunohistochemical stains.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Insulin was measured by the Wisconsin National Primate Research Center Assay Services Division, University of Wisconsin–Madison, with partial support by grant P51 RR000167 from the NIH. Work performed at the NEPRC was supported in part by grant P51 OD011103 from the NIH.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.