
Abstract
Objectives
Chlorogenic acid (CGA) is an antioxidant dietary factor. We investigated the effects of CGA on endothelial cell dysfunction in diabetic mice and the mechanistic role of nuclear factor erythroid-related factor 2 (Nrf2) in the antioxidant effect of CGA.
Methods
Diabetic (db/db) mice were fed normal chow or chow containing 0.02% CGA for 12 weeks. Human umbilical vein endothelial cells (HUVECs) and mouse aortas were treated with normal or high glucose.
Results
CGA treatment induced upregulation of Nrf2 in HUVECs in a dose-dependent manner. CGA pretreatment prevented reactive oxygen species generation and preserved nitric oxide bioavailability in HUVECs and aortas from wild-type but not Nrf2−/− mice. CGA improved endothelium-dependent relaxation in high glucose-treated aortas from wild-type and db/db mice, but not Nrf2−/− mice. Dietary CGA improved endothelium-dependent relaxation in db/db mice.
Conclusions
CGA ameliorates endothelial dysfunction in diabetic mice through activation of the Nrf2 anti-oxidative pathway.
Keywords
Introduction
Diabetes is a major risk factor for cardiovascular disease. 1 Between 70% and 80% of diabetic patients will eventually die from cardiovascular complications, imposing a heavy financial burden on society. 1 Glucose toxicity, low-grade inflammation, and oxidative stress induce insulin resistance, damage pancreatic β cells, and cause endothelial dysfunction, ultimately resulting in micro- and macro-vascular complications.2,3 Unfortunately, there is no specific treatment for diabetic endothelial dysfunction.
Oxidative stress is a condition in which reactive oxygen species (ROS) production exceeds that of antioxidants. 4 Oxidative stress is a critical contributor to diabetic vascular injury. 5 Increased intracellular ROS levels can induce apoptosis of endothelial cells. 6 As a redox-sensitive transcription factor, nuclear factor erythroid 2-related factor 2 (Nrf2) exerts antioxidant actions by inducing expression of antioxidant enzymes. 7 Under physiologic conditions, Nrf2 is found in the cytoplasm at low concentration. 8 Under stressful conditions, Nrf2 translocates into the nucleus and acts as a transcription factor to maintain redox homeostasis. 9 Recent studies have revealed that Nrf2 plays an important role in diabetic vascular injury and that targeting Nrf2 may be a promising strategy for treatment of diabetes-related cardiovascular complications.
Chlorogenic acid (CGA) is a polyphenolic compound that is abundant in coffee, strawberries, and sunflowers. 10 CGA possesses many biological properties, including antioxidant and anti-carcinogenic activities. 11 Recently, the roles of CGA in glucose and lipid metabolic disorders have also been investigated. 12 Dietary administration of CGA attenuated isoproterenol-induced oxidative stress in the rat myocardium. 13 CGA directly induces endothelium-dependent vasodilation in isolated rat vessels. 14 Although CGA has consistently been shown to inhibit oxidative stress and exert cardioprotective effects, its role in diabetes-induced endothelial dysfunction remains unknown. The present study aimed to test the hypothesis that CGA attenuates diabetes- and high-glucose-induced endothelial dysfunction via Nrf2-mediated antioxidant effects.
Methods
Animals
All animal experiments were approved by the Institutional Animal Care and Use Committee of Chengdu Medical College (2018013). Male Nrf2 knockout mice (Nrf2−/−, 12 weeks old) were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). Male leprdb diabetic (db/db) mice and age-matched wild-type control mice (C57BL/KsJ) were purchased from the Model Animal Research Center (Nanjing, Jiangsu, China). Mice were housed under a 12-hour/12-hour light/dark cycle with free access to food and water. Mice were fed normal chow (control group) or normal chow containing 0.02% CGA (Sigma-Aldrich, St. Louis, MO, USA) (CGA group) for 12 weeks.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from Schenkel Research Laboratories (San Diego, CA, USA). HUVECs were grown in serum- and supplement-free Dulbecco’s Modified Eagle’s Medium (DMEM, HyClone, Logan, UT, USA) containing normal glucose (NG, 5 mM) for 24 hours. Then, cells were either maintained in serum-free NG DMEM (containing 25 mM mannitol as an osmotic control) or switched to serum-free high glucose (HG, 30 mM) DMEM with or without CGA (10 μM, diluted in dimethyl sulfoxide). The nuclear fractions were prepared using NE-PER™ Nuclear Extraction Reagents (Thermo Scientific, Waltham, MA, USA). 15
Delivery of small interfering RNA (siRNA)
HUVECs were cultured in medium containing 20 µg of Nrf2 siRNA or scrambled siRNA (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 6 hours per day on 2 consecutive days according to the manufacturer’s instructions.
Measurement of vascular tone
Aortic rings from WT and Nrf2−/− mice were dissected in sterile phosphate-buffered saline and incubated in DMEM supplemented with 10% fetal bovine serum (Gibco, Carlsbad, CA, USA), 100 IU/mL penicillin, and 100 μg/mL streptomycin. After a 24-hour incubation period, a wire myograph was used to assess vascular reactivity of the aortic rings. 16 An optimized baseline tension was applied to the arterial rings, and arterial segments were balanced for 1 hour. The rings were pre-contracted with phenylephrine (1 µM), then endothelium-dependent relaxation in response to acetylcholine (ACh, 10−9 M – 10−5 M) and endothelium-independent relaxation in response to nitroglycerine (NTG, 10−9 M – 10−5 M) were recorded.
Determination of ROS
Dihydroethidium (DHE, Sigma-Aldrich) was used to detect superoxide as described previously. 17 In situ formation of ROS was measured by DHE staining. DHE staining was visualized using a fluorescence microscope (TE2000, Nikon, Toyoko, Japan). Images were analyzed using Image-Pro Plus 6.0 (Media Cybernetics, Rockville, MD, USA).
Evaluation of nitric oxide (NO) levels
HUVECs and aortic segments were washed three times in Krebs solution and stained with diaminofluorescein-2 diacetate (DAF-2DA, a NO-sensitive fluorescent dye; Sigma-Aldrich). After incubation for 45 minutes at 37°C, DAF-2 DA fluorescence intensity was analyzed using NIS-Elements 3.0 software (Nikon). 18
Measurement of protein expression
Membranes were incubated with primary antibodies against Nrf2 (Abcam, Cambridge, MA, USA), nitrotyrosine (Santa Cruz Biotechnology), quinone oxidoreductase 1 (NQO1, Santa Cruz), heme oxygenase-1 (HO-1, Santa Cruz), P22phox (Santa Cruz), P47phox (Santa Cruz), P67phox (Santa Cruz), glutathione peroxidase 1 (GPx-1, Santa Cruz), glyceraldehyde 3-phosphate dehydrogenase (GAPDH, Santa Cruz), and Histone H1 (Abcam). Blotted membranes were washed and incubated with secondary antibodies (ZSGB-Bio, Beijing, China) for 2 hours at room temperature. Protein levels were normalized to levels of GAPDH or Histone H1. Blots were visualized using a Gel Doc 2000 Imager (Bio-Rad, Hercules, CA, USA).
Statistical analysis
All data were presented as means ± standard errors of the means (SEMs). Maximum response (Emax) was calculated based on concentration-response curves using GraphPad Prism software version 6.0 (GraphPad Software Inc., La Jolla, CA, USA). Differences in the means of three or more groups were assessed using one-way analysis of variance with post-hoc Tukey’s honestly significant difference test. Differences between the means of two groups were assessed using the Student’s t-test. Values of P<0.05 were considered statistically significant.
Results
CGA upregulated Nrf2 and downregulated nicotinamide adenine dinucleotide phosphate (NADPH) oxidase in HUVECs
Previous studies reported a Nrf2-mediated cytoprotective effect of CGA, 19 but the role of CGA in endothelial cell function remains unknown. We first investigated the effect of CGA on Nrf2 expression in HUVECs. Treatment with CGA increased nuclear Nrf2 expression in HUVECs in a dose-dependent manner. The effects of 10 µM and 100 µM CGA doses were similar (Figure 1a). As shown by western blotting, CGA (10 µM) also upregulated expression of the Nrf2 downstream targets HO-1, NQO1, and GPx1 (Figure 1b–1e). CGA (10 µM) pretreatment decreased the expression of the NADPH oxidase submits P22phox, P47phox, and P67phox, and reduced the generation of nitrotyrosine, a marker of ONOO−-mediated nitration that plays a critical role in vascular injury under hyperglycemic conditions. CGA also upregulated Nrf2 expression in HG-treated HUVECs (Figure 2a). Furthermore, CGA attenuated HG-induced upregulation of P22phox, P47phox, and P67phox, as well as HG-induced generation of nitrotyrosine (Figure 2b–2f). The beneficial effects of CGA were abrogated by Nrf2 silencing using siRNAs.

Effects of chlorogenic acid (CGA) on expression of Nrf2 and its downstream targets in human umbilical vein endothelial cells (HUVECs). (a) Representative data showing the effects of different doses of CGA (0–100 μM) on Nrf2 expression in HUVECs. Histone H1 expression was used as a reference. CGA increased Nrf2 expression in a dose-dependent manner. *P < 0.05, **P < 0.01 compared with control group (Cont). (b–e) CGA increased the expression of the Nrf2 downstream target proteins heme oxygenase-1 (HO-1), quinone oxidoreductase 1 (NQO1), and glutathione peroxidase 1 (GPx-1) in HUVECs. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression was used as a reference. **P < 0.01 compared with control group (Cont). Data are presented as means ± SEMs (n = 6).

Effects of chlorogenic acid (CGA) on P22phox, P47phox, P67phox and nitrotyrosine levels in human umbilical vein endothelial cells (HUVECs) cultured under high glucose (HG) conditions. Histone H1 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression were used as a reference. (a) Exposure to HG (30 mM) increased the expression of Nrf2, and Nrf2 expression was further elevated by CGA treatment. (b–f) HG (30 mM) increased P22phox, P47phox, P67phox and nitrotyrosine levels, while CGA pretreatment decreased P22phox, P47phox, P67phox and nitrotyrosine levels under HG conditions. These effects were blocked by siRNA silencing of Nrf2. **P < 0.01 compared with the normal glucose (NG, 5 mM) group; ##P < 0.01 compared with the HG group; △△P < 0.01 compared with the HG+CGA group (n = 6).
CGA decreased superoxide levels and preserved NO levels in HUVECs
NO plays a pivotal role in protection of endothelial cells. Loss of endothelium-derived NO is a hallmark of endothelial dysfunction and is implicated in the pathogenesis of cardiovascular disease. 20 We next asked whether the effects of CGA on Nrf2 and its downstream targets impacted NO levels in HUVECs cultured under HG conditions. CGA pretreatment prevented superoxide anion generation (Figure 3a and 3c) and preserved NO levels in HG-treated HUVECs (Figure 3b and 3d), as shown by DHE and DAF-2DA staining, respectively. The beneficial effects of CGA were abrogated by Nrf2 silencing using siRNAs (Figure 3). These results suggested that CGA prevents HG-induced ROS generation and preserves NO bioavailability in HUVECs through an Nrf2-dependent pathway.
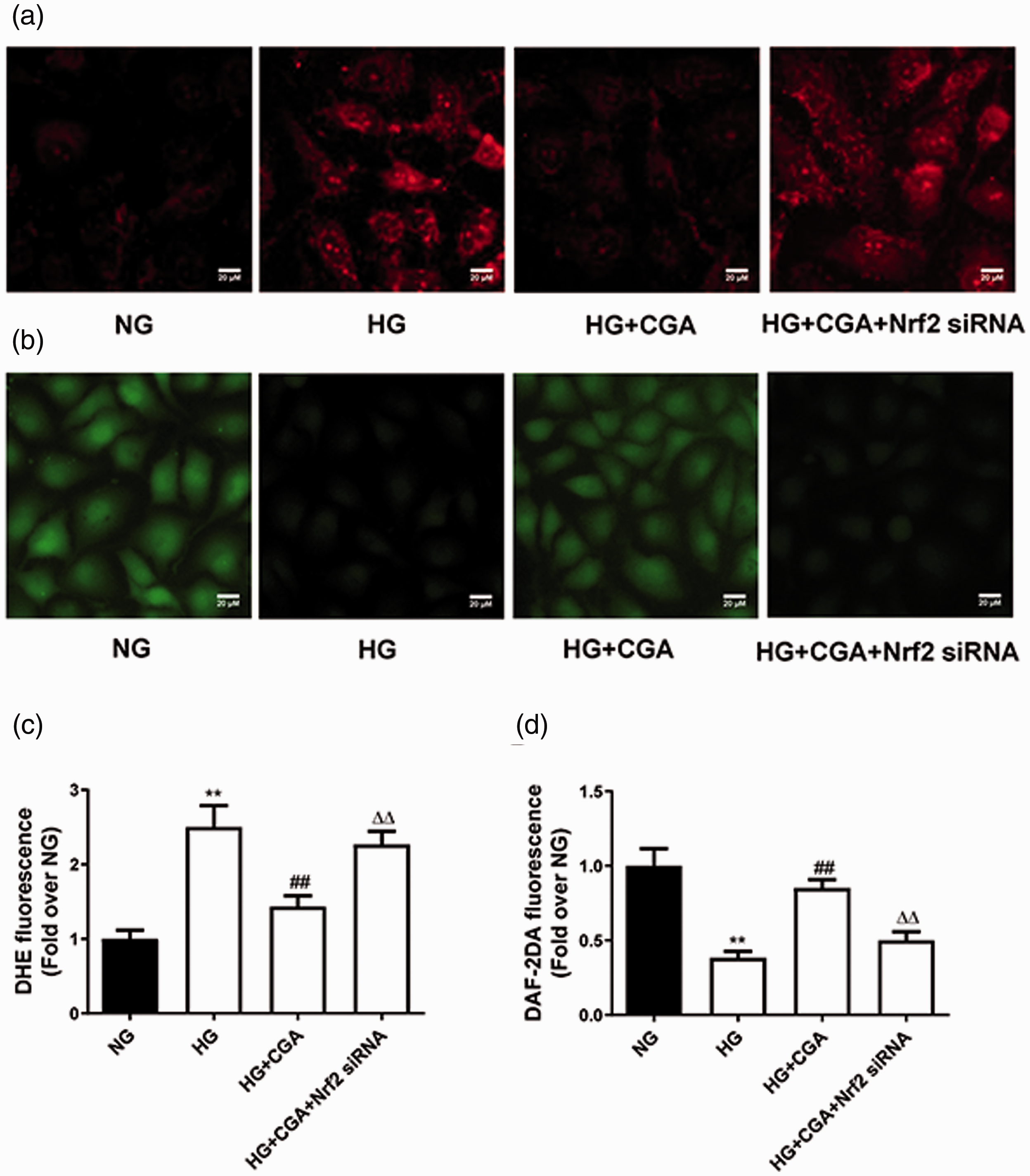
Effects of chlorogenic acid (CGA) on reactive oxygen species production and nitric oxide (NO) levels in high glucose (HG)-treated human umbilical vein endothelial cells (HUVECs). (a, c) Representative images and data obtained from dihydroethidium (DHE)-stained HUVECs. HG exposure increased the levels of superoxide anions in HUVECs, while CGA administration (10 µM) decreased superoxide anion levels under HG conditions. These effects were blocked by siRNA silencing of Nrf2. (b, d) Representative images of diaminofluorescein-2 diacetate (DAF-2DA)-stained HUVECs. High glucose exposure decreased NO levels in HUVECs, while CGA treatment increased NO levels under HG conditions. These effects were blocked by siRNA silencing of Nrf2. **P < 0.01 compared with the normal glucose (NG, 5 mM) group; ##P < 0.01 compared with the HG group; △△P < 0.01 compared with the HG+CGA group. Data are presented as means ± SEMs (n = 6). Scale bar, 20 µm.
CGA preserved the endothelial function of mouse aortas via Nrf2
Aortic rings were isolated from wild-type and Nrf2−/− mouse aortas and cultured in NG, HG, or HG medium containing CGA. CGA (10 µM) reversed HG-induced increases in ROS production and decreases in NO levels in aortas from wild-type but not Nrf2−/− mice (Figure 4). Moreover, we found that CGA ameliorated impairment of ACh-induced endothelium-dependent relaxation under HG conditions in cultured aortic rings from wild-type (Figure 5a) but not Nrf2−/− mice (Figure 5b). We observed no effect of CGA on NTG-induced endothelium-independent relaxation in either wild-type or Nrf2−/− mice (Figure 5c and 5d). These results suggested that CGA preserves endothelial function under HG conditions in an Nrf2-dependent manner.

Effects of chlorogenic acid (CGA) on reactive oxygen species (ROS) production and nitric oxide (NO) levels in mouse aortas treated with high glucose (HG) ex vivo. (a, b, e) ROS production was measured by dihydroethidium (DHE) staining in the en face endothelium of aortas from Nrf2 knockout (Nrf2−/−) and matched wild type (WT) mice after treatment with normal glucose (NG), HG or HG+CGA (10 µM). (c, d, f) NO production in the en face endothelium of aortas was measured by diaminofluorescein-2 diacetate (DAF-2DA) staining. Red: DHE fluorescence. Green: DAF-2DA fluorescence. **P < 0.01 compared with the NG group; ##P < 0.01 compared with the HG group. Data are presented as means ± SEMs (n = 6). Scale bar, 20 µm.

Effects of chlorogenic acid (CGA) on high glucose (HG)-induced endothelial dysfunction. (a, b) HG exposure (30 mM, 12 hours) impaired the endothelium-dependent relaxation of mouse aortas in vitro. CGA (10 µM, 12 hours) ameliorated endothelial dysfunction in wild type mouse aortas but not in Nrf2−/− mouse aortas cultured under HG conditions. **P < 0.01 compared with the normal glucose (NG) group; ##P < 0.01 compared with the HG group; △△P < 0.01 compared with the HG+CGA group. (c–d) Endothelium-independent relaxation is shown for all groups tested. Relaxation is shown as a percentage compared with phenylephrine (Phe)-induced contraction. Data are presented as means ± SEMs (n = 6).
Chronic CGA administration improved endothelium function in diabetic mice via Nrf2
Diabetic (db/db) mice and age-matched wild-type mice were used to explore the effects of CGA on endothelial dysfunction in vivo. After 12 weeks of CGA (0.02%) feeding, db/db mice had decreased body weights (Figure 6a), fasting blood glucose (Figure 6b), and hemoglobin A1c levels (Figure 6c) compared with control mice. Dietary CGA attenuated ROS production and increased NO levels in the endothelium of db/db mice (Figure 6d–6g). A wire myograph was used to assess relaxation of mouse aortas. Twelve weeks of CGA feeding improved ACh-induced aortic relaxation in db/db mice

Effects of dietary chlorogenic acid (CGA) on body weights, fasting blood glucose levels, endothelial oxidative stress, and nitric oxide (NO) levels in diabetic mice. (a) Effect of 12 weeks of CGA administration on the body weights of db/db mice. (b) Representative data show that 12 weeks of dietary CGA administration decreases fasting blood glucose level in db/db mice. (c) Effect of 12 weeks of CGA administration on serum hemoglobin (HbA1c) levels in db/db mice. (d–e) Superoxide anion production assessed via dihydroethidium (DHE) staining in aortas from lean littermate control C57BL/KsJ mice and db/db mice fed either a normal diet (db/db Cont) or a normal diet containing 0.02% CGA (db/db CGA). (f–g) NO production assessed via diaminofluorescein-2 diacetate (DAF-2DA) staining in aortas from C57BL/KsJ and db/db Cont mice fed a normal diet or a normal diet containing 0.02% CGA for 12 weeks. **P < 0.01 compared with the C57BL/KsJ group; ##P < 0.01 compared with the db/db Cont group (n = 6). Scale bar, 20 µm.
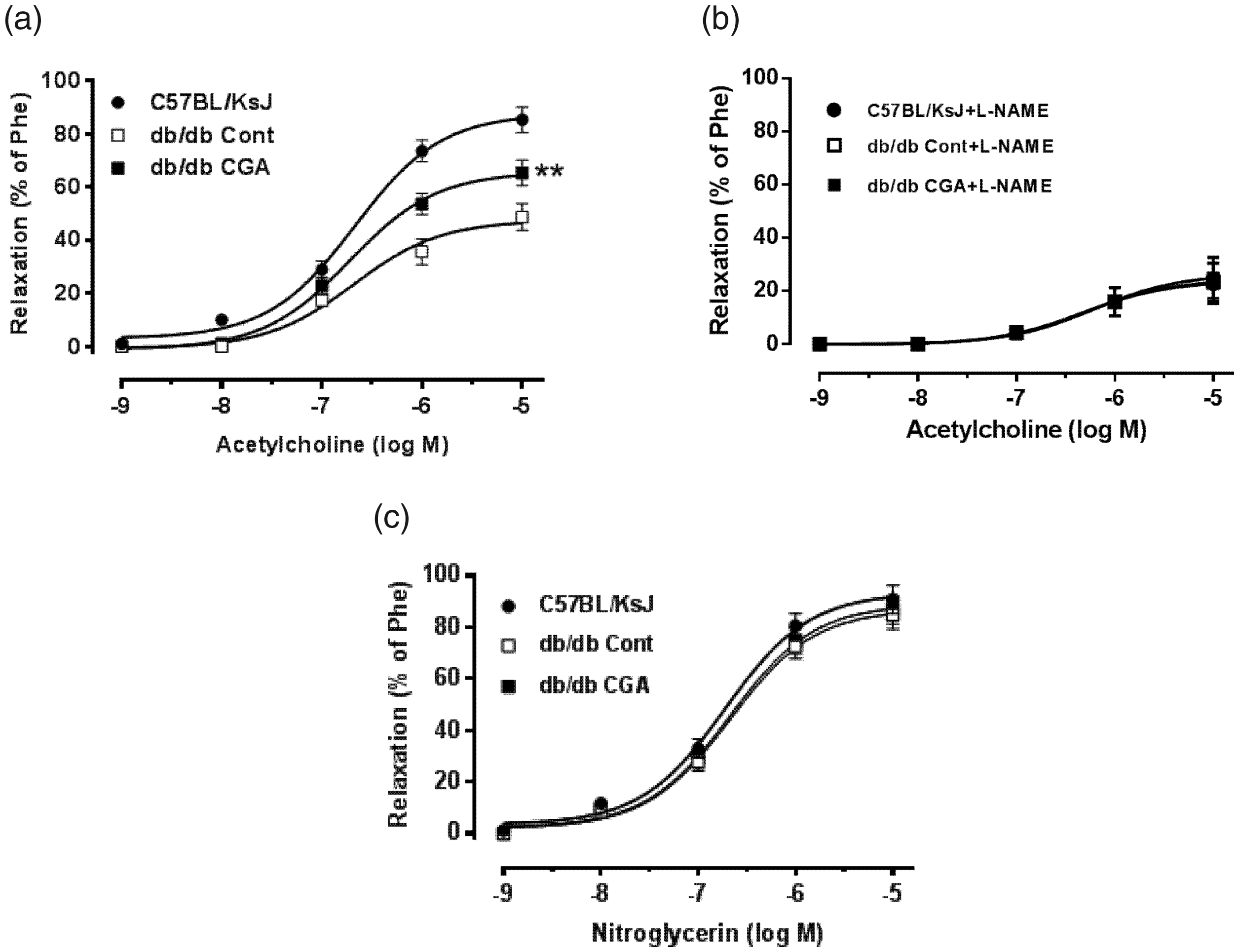
Effects of dietary chlorogenic acid (CGA) on endothelium-dependent relaxation in db/db mice. (a) Quantitation of endothelium-dependent relaxation induced by acetylcholine treatment (10−9 M – 10−5 M) in aortas from db/db mice fed a normal diet or a normal diet containing CGA for 12 weeks. (b) The inhibitory effect of L-NAME (100 µM, 30 minutes) on acetylcholine-induced maximal relaxation in aortas from db/db mice fed a normal diet or a normal diet containing CGA for 12 weeks. (c) Representative data showing endothelium-independent relaxation of aortas from db/db mice fed a normal diet or a normal diet containing CGA for 12 weeks. Data are presented as the means ± SEMs of six rings (n = 6 mice per group). Relaxation is shown as a percentage compared with phenylephrine (Phe)-induced contraction. *P < 0.05 and **P < 0.01 compared with db/db mice fed a normal diet (db/db Cont).

Effects of dietary chlorogenic acid (CGA) on expression of Nrf2, P22phox, P47phox, P67phox, and levels of nitrotyrosine in the vasculature of diabetic mice. Histone H1 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression were used as a reference. (a, b) Representative western blotting data showing Nrf2 (a), P22phox, P47phox, P67phox and nitrotyrosine (b) levels in db/db mice fed a normal diet (db/db Cont) or a normal diet containing 0.02% CGA (db/db CGA) and in lean littermate control C57BL/KsJ mice. (c–f) Effects of dietary CGA on levels of P22phox (c), P47phox (d), P67phox (e) and nitrotyrosine (f) in the vasculature of db/db mice. Data are presented as means ± SEMs (n = 6). **P < 0.01 compared with C57BL/KsJ group; #P < 0.05 compared with the db/db mice fed a normal diet (db/db Cont).
Discussion
The present study showed that under HG conditions, CGA upregulated Nrf2 and its downstream targets, downregulated P22phox, P47phox, and P67phox, prevented ROS and nitrotyrosine production, and preserved NO levels in HUVECs. Similar effects were observed in cultured aortic rings from wild-type mice but not Nrf2−/− mice. Furthermore, an in vivo study demonstrated that dietary CGA increased the expression of antioxidant enzymes (HO-1, GPx-1, and NQO1) and improved endothelial function. These findings suggest that CGA ameliorates endothelial dysfunction in diabetic mice.
Oxidative stress is closely associated with the pathogenesis of diabetes-related cardiovascular complications. 21 CGA, one of the most abundant polyphenol compounds in the human diet, belongs to a group of phenolic secondary metabolites produced by certain plant species and is a major component of coffee. 22 CGA has been shown to attenuate oxidative stress both in models of oxidized low-density lipoprotein-induced endothelial dysfunction and streptozotocin-induced diabetes in rats. 23 Additionally, the protective effects of CGA against oxidative stress-induced cell damage were associated with the Nrf2 signaling pathway. 24 However, whether CGA protects against hyperglycemia-induced endothelial cell injury was unclear, and its effects on endothelial dysfunction in diabetes were unknown. In the present study, we showed that CGA increased nuclear Nrf2 expression in a dose-dependent manner. Absorption and metabolism of CGA have been studied in healthy volunteers and human tissues. 25 Because there is no evidence that these dietary conjugates are absorbed in vivo and true bioavailability data are incomplete, 26 previously determined concentrations of CGA may be inaccurate. Despite these caveats, high levels of CGA are potentially achievable in human blood. Our study is the first to assess the effects of CGA on Nrf2 in endothelial cells.
Nrf2, a transcription factor with a high sensitivity to oxidative stress, binds to antioxidant responsive elements in the nucleus and promotes transcription of a wide variety of antioxidant genes. 27 HO-1, NQO1, and GPx1 are the major downstream antioxidant effectors of Nrf2. Upregulation and activation of Nrf2 increases the expression of HO-1, NQO1, and GPx1 through positive transcriptional regulation. In the present study, we found that CGA upregulated the expression of HO-1, NQO1, and GPx1 in addition to Nrf2. These results suggest that CGA enhances expression of anti-oxidative genes including HO-1 via upregulation of Nrf2. This finding is consistent with a previous study demonstrating that CGA activated the Nrf2/HO-1 anti-oxidative pathway in osteoblastic cells. 28
Instead of relying on a single antioxidant effector, Nrf2 activation results in the coordinated upregulation of several antioxidant enzymes and cytoprotective genes, making it an attractive target for treatment of diabetic complications. 29 Our in vitro studies showed that CGA attenuated ROS production, preserved NO levels, and decreased P22phox, P47phox, P67phox, and nitrotyrosine levels in HUVECs under HG conditions. Consistently, previous studies demonstrated that NADH/NADH oxidase can be activated via upregulation of P22phox, P47phox, and P67phox, and represents the predominant source of ROS in the vasculature. 30 To confirm the role of Nrf2 in HUVECs, we used an Nrf2 siRNA that substantially decreased the expression of Nrf2. We found that the protective effects of CGA were abolished by silencing of Nrf2. Studies of Nrf2−/− and matched wild-type mice further confirmed these results. CGA pretreatment inhibited ROS generation and preserved NO levels in the endothelium of wild-type mice, but not Nrf2−/− mice. Endothelium-dependent relaxation, but not endothelium-independent relaxation, was impaired under HG conditions in wild-type mice. Pretreatment with CGA preserved endothelium-dependent relaxation in wild-type mice, but not Nrf2−/− mice.
Because db/db mice recapitulate key features of human type 2 diabetes, 31 we used db/db mice to investigate the effects of dietary CGA on diabetes-related endothelial dysfunction. After 12 weeks of dietary CGA administration, db/db mice showed attenuated ROS production, increased NO levels, and improved endothelium-dependent relaxation of aortic rings. These improvements were associated with increased Nrf2 expression and decreased P22phox, P47phox, P67phox, and nitrotyrosine levels following CGA treatment.
In conclusion, our findings suggest that CGA prevents hyperglycemia-induced endothelial dysfunction via the Nrf2 pathway.
Footnotes
Authors’ Contributions
PW, PZ: conception and design; DW, JH, JW, YY, SL, and WL: collection and/or assembly of data; DW, JH, XL, XD, PZ, PW: data analysis and interpretation, manuscript writing, and manuscript revision; DW, JH, JW, YY, SL, XL, WL, XD, PZ, PW: final approval of manuscript.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
This work was supported by grants from the National Natural Science Foundation of China (81970262 to P.W.), the Central Funds Guiding the Local Science and Technology Development of Sichuan Province (2020ZYD036 to P.W.), the Innovation Team Project Department of Education of Sichuan Province (18TD0030 to P.W.), and the Major Project of Education Department in Sichuan Province (16ZA0293 to X.L.).