
Abstract
Objective
Osteoarthritis (OA) is a chronic degenerative arthropathy characterized by articular cartilage degeneration, subchondral osteosclerosis, and hyperosteogeny. MicroRNAs (miRNAs) play an important regulatory role in its pathological development, so this study explored the effect and potential mechanism of miR-146a-5p in interleukin (IL)-1β-induced OA cartilage injury.
Methods
The human chondrosarcoma cell line SW1353 and normal human chondrocytes C28/I2 were stimulated by IL-1β to construct the OA chondrocyte model. miR-146a-5p and thioredoxin interacting protein (TXNIP) expression levels were detected by quantitative real-time (qRT)-PCR and western blot. Their expression was modified by transfecting an miR-146a-5p inhibitor, mimic, and pcDNA-TXNIP. The relationship between miR-146a-5p and TXNIP was analyzed by the dual-luciferase assay, while cell viability, apoptosis, and inflammatory expression were determined by cell counting, TUNEL staining, and ELISA, respectively.
Results
miR-146a-5p expression was upregulated in SW1353 and C28/I2 cells stimulated by IL-1β. miR-146a-5p knockdown significantly enhanced cell activity, inhibited inflammatory factor expression, and reduced cell apoptosis. The dual-luciferase assay revealed TXNIP as a target gene of miR-146a-5p and suggested that miR-146a-5p promotion of OA damage could be reversed by upregulating TXNIP.
Conclusion
These results suggest that miR-146a-5p inhibits cell proliferation and promotes apoptosis and the inflammatory response in OA cartilage injury by modulating TXNIP.
Introduction
Osteoarthritis (OA) is a chronic bone and joint disease characterized by articular cartilage degeneration and arthritis. Typical clinical manifestations are chronic pain and joint movement disorders, which seriously affect the quality of life of affected patients. 1 , 2 Early drug therapy and middle and late joint replacement are the main treatments for OA but these can be associated with adverse reactions. 3
The degeneration of articular cartilage in OA is thought to be caused by a decrease in the number of chondrocytes (the only cell type of articular cartilage) and cytokine and growth factor-stimulated degradation of the extracellular matrix (ECM). 4 , 5 Therefore, understanding the molecular mechanism involved in the initiation and progression of OA is important to identify early diagnostic markers and biotherapeutic targets of OA.
MicroRNAs (miRNAs) are a class of highly conserved, endogenous non-coding RNAs containing 19 to 25 nucleotides. They mainly bind to the 3′-untranslated region (UTR) of target mRNA, which is then degraded or silenced, thus affecting cell proliferation, differentiation, apoptosis, and ontogeny. 6 , 7 Several studies have shown that miRNAs play an important role in the pathological development of OA, especially in maintaining homeostasis of the endochondral environment. There is also a significant difference in miRNA expression between OA patient and healthy chondrocytes, which leads to an imbalance of chondrocyte synthesis and catabolism and the occurrence and development of OA. 8
As an example, miR-373 expression was reported to be downregulated in OA patient chondrocytes, while its overexpression promoted chondrocyte proliferation by inhibiting the expression of P2X7 receptor and inflammatory factors. 2 Interleukin (IL)-1β also promoted the occurrence of OA and degradation of the ECM through regulating the mitogen-activated protein kinase/nuclear factor-κB pathway by inhibiting miR-27a-3p. 9 Recently, miR-146a-5p was shown to play an important regulatory role in a variety of tumors, including gastric cancer, lung cancer, and prostate cancer.10–12 Furthermore, its expression was significantly upregulated in the cartilage tissues and serum of OA patients compared with healthy controls.13,14 However, to the best of our knowledge, few studies have investigated the role and mechanism of miR-146a-5p in OA.
In this study, two types of chondrocytes (SW1353 and C28/I2 cells) were stimulated with IL-1β to establish the OA model. Subsequently, cell viability, apoptosis, and the expression of inflammatory factors were assessed by the cell counting kit (CCK)-8 assay, TUNEL staining, and ELISA to determine the role of miR-146a-5p in OA. The regulatory relationship between miR-146a-5p and thioredoxin-interacting protein (TXNIP) was also explored as TXNIP has been shown to be significantly decreased in OA cartilage. 15 Our results suggest a novel biological target for the prevention and treatment of OA.
Methods
Cell culture and treatment
SW1353 and C28/I2 cells were purchased from the American Type Culture Collection (ATCC; Rockville, MD, USA). Cells were cultured in Dulbecco’s modified Eagle’s medium (HyClone, Logan, UT, USA) containing 10% fetal bovine serum (HyClone), 100 U/ml penicillin, and 100 μg/ml streptomycin in a humidified atmosphere of 95% O2 and 5% CO2 at 37°C. To construct the OA model, both of these cell types in logarithmic growth phase were inoculated in 96-well plates at a density of 1 × 104 cells/well. When they had reached 80% to 90% confluence, they were cultured in serum-free medium for 24 hours then treated with 10 ng/ml IL-1β for 24 hours. Five replicates were used for every group.
Cell transfection
The TXNIP gene was artificially synthesized by Genscript Biotechnology Co., Ltd. (Nanjing, China) and we cloned it into the pcDNA3.1 vector (Invitrogen, Carlsbad, CA, USA) to construct the reporter vector pcDNA-TXNIP. miR-146a-5p mimics, an miR-146a-5p inhibitor and corresponding scrambled control were obtained from GENEWIZ (Shanghai, China). These plasmids were transfected into cells using Lipofectamine 2000 (Invitrogen) following the manufacturer’s instructions. Transfection efficiency was detected after 48 hours.
Quantitative real time (qRT)-PCR analysis
Total RNA was extracted by TRIzol reagent according to the manufacturer’s instructions (Thermo Fisher Scientific, Carlsbad, CA, USA) and reverse-transcribed into cDNA using the PrimeScript™ II 1st Strand cDNA Synthesis Kit (TaKaRa, Dalian, China). Real-time PCR analysis was performed on the ABI PRISM 7300 system (Applied Biosystems, Waltham, MA, USA) using SYBR Premix ExTaq (TaKaRa). GAPDH was used as the internal reference gene. The relative expression of TXNIP and miR-146a-5p was determined following the 2−ΔΔCt method (Livak & Schmittgen). 16
CCK-8 assay
Cells grown in logarithmic phase were inoculated in 96-well plates at a density of 1 × 104 cells/well for conventional culture. After 24 hours, 10 μl CCK-8 solution was added to each well. Cells were placed in the dark for 1 hour, then absorbance at a 450-nm wavelength was detected by a microplate reader (Bio-Rad, Hercules, CA, USA).
TUNEL staining
Apoptosis was detected by TUNEL staining using the One-Step TUNEL Apoptosis Kit according to the manufacturer’s instructions (Merck, Darmstadt, Germany). The nuclei of apoptotic cells stained brown-yellow, and this was observed and analyzed by inverted fluorescent microscopy.
ELISA assay
Cells in logarithmic phase were inoculated in 6-well plates at a density of 5 × 104 cells/well and cultured for 24 hours. Cell supernatants from each group were collected and centrifuged at 2700 ×g for 10 minutes at 4°C. The supernatants were then aspirated and the expression of inflammatory factors (tumor necrosis factor [TNF]-α, IL-1β, and IL-6) was detected by ELISA (Elascience, Wuhan, China) according to the manufacturer’s instructions.
Dual-luciferase reporter assay
pcDNA-TXNIP containing the wild-type 3′-UTR binding site or pcDNA-TXNIP containing the mutant 3′-UTR binding site were cotransfected into HEK293T cells using Lipofectamine 2000 with miR-146a-5p mimic or negative control. The luciferase reporter assay kit (Promega, Madison, WI, USA) was used to detect luciferase activity.
Western blot analysis
Total cell protein was extracted with radioimmunoprecipitation assay buffer (CWBIO, Beijing, China) and the protein concentration was detected by the BCA kit (CWBIO). Proteins were then separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes. These membranes were blocked with 5% skimmed milk powder for 1 hour, washed several times with Tris-buffered saline and Tween 20, then incubated with primary antibodies against TXNIP (dilution 1:1000; Cell Signaling Technology Inc., Danvers, MA, USA) and β-actin (dilution 1:1000; Cell Signaling Technology Inc.) at 4°C overnight. They were then incubated with secondary antibodies conjugated to horseradish peroxidase (dilution 1:5000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at room temperature for 2 hours, and protein bands were visualized with ECL reagent (GE Healthcare, Chicago, IL, USA) and imaged by the typhoon scanner (GE Healthcare). β-actin was used as an internal reference.
Bioinformatics
Predicted miRNA target genes were determined using the TargetScan website (http://www.targetscan.org). Briefly, ‘miR-146a-5p’ was input into the ‘Enter a microRNA name’ box on the website.
Statistical analysis
SPSS statistical software (version 19.0; IBM, Armonk, NY, USA) was used for statistical analysis. Experimental data are expressed as means ± standard deviations. The Student’s t-test was used to compare data between two groups, and one-way analysis of variance followed by Tukey’s post hoc test was used to compare multigroup data. P < 0.05 was considered statistically significant.
Results
Effect of miR-146a-5p knockdown on proliferation and apoptosis of IL-1β-stimulated chondrocytes
To investigate the role of miR-146a-5p in OA cartilage injury, we first determined the expression of miR-146a-5p in SW1353 and C28/I2 cells stimulated by IL-1β. qRT-PCR analysis showed that IL-1β stimulation significantly increased the expression of miR-146a-5p in SW1353 and C28/I2 cells compared with the IL-1β (10 ng/mL)-0h group (p < 0.001; Figure 1a). Next, gene knockdown experiments were performed to determine the effect of miR-146a-5p on SW1353 and C28/I2 cells stimulated with IL-1β. qRT-PCR analysis showed that gene knockdown was effective at significantly reducing miR-146a-5p expression in the miR-146a-5p inhibitor group compared with the IL-1β-negative control (NC) inhibitor group (p < 0.001; Figure 1b). CCK-8 analysis revealed that the viability of SW1353 and C28/I2 cells stimulated with IL-1β was significantly lower than that of the control group, while the viability of miR-146a-5p inhibitor cells was significantly higher than that of IL-1β-NC inhibitor cells (p < 0.05; Figure 1c). TUNEL staining showed that IL-1β stimulation greatly promoted the apoptosis of SW1353 and C28/I2 cells compared with the control group, whereas apoptosis was greatly reduced by miR-146a-5p inhibitor transfection (Figure 2d).

Effect of miR-146a-5p knockdown on proliferation and apoptosis of IL-1β-stimulated chondrocytes. (a) Intracellular miR-146a-5p expression detected by qRT-PCR after IL-1β treatment. (b) miR-146a-5p expression detection by qRT-PCR after transfection with miR-146a-5p inhibitor and NC inhibitor. (c) Cell viability as determined by CCK-8. (d) Apoptosis as determined by TUNEL staining. ***p < 0.001 vs. IL-1β (10 ng/ml)-0h; #p < 0.05, ##p < 0.01, and ###p < 0.001 vs. IL-1β-NC inhibitor
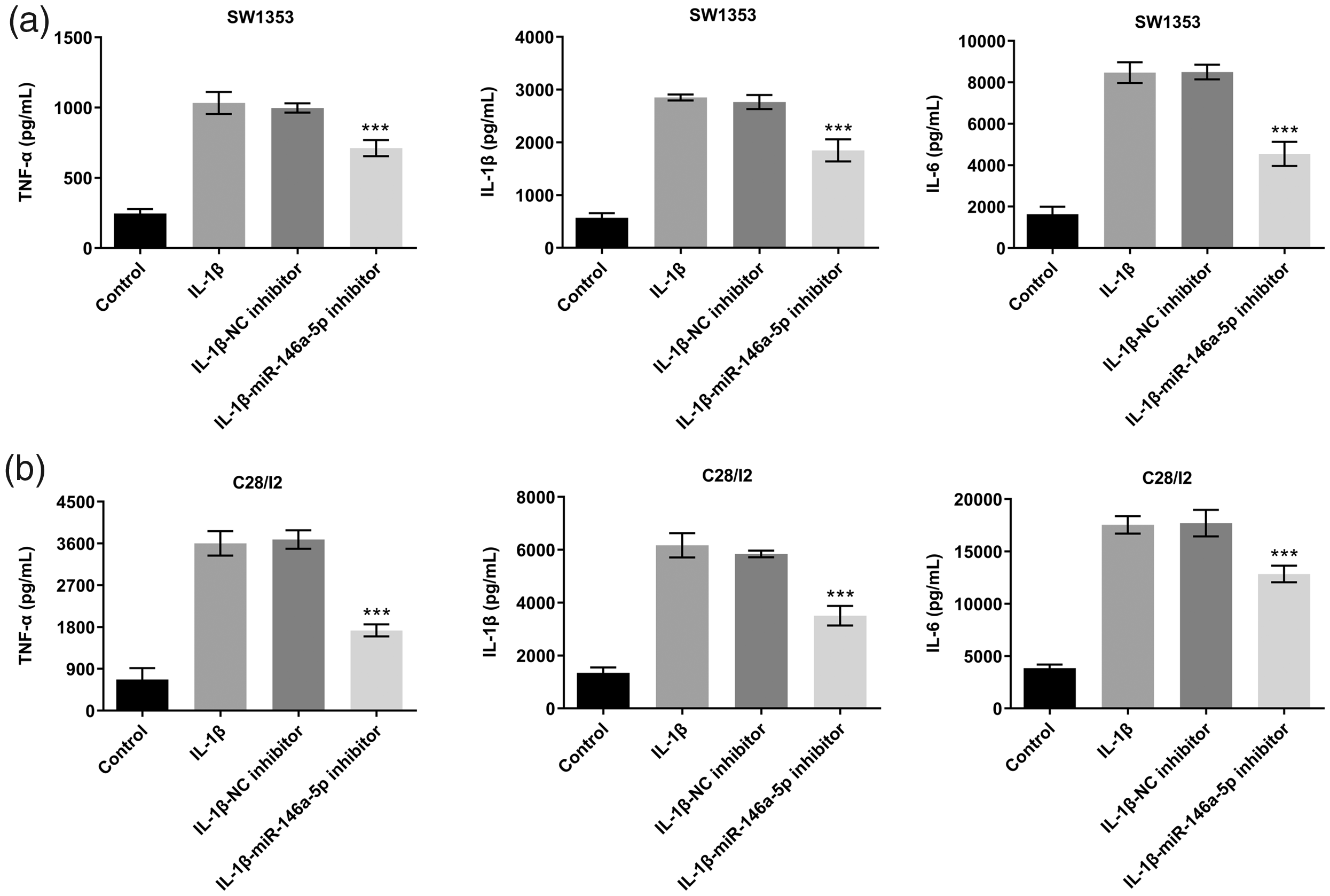
Effect of miR-146a-5p knockdown on inflammatory cytokine expression in IL-1β-stimulated chondrocytes. (a) ELISA detection of TNF-α, IL-1β, and IL-6 expression in IL-1β-stimulated SW1353 cells. (b) ELISA detection of TNF-α, IL-1β, and IL-6 expression in IL-1β-stimulated C28/I2 cells. ***p < 0.001 vs. IL-1β-NC inhibitor.
Effect of miR-146a-5p knockdown on inflammatory cytokine expression in IL-1β-stimulated chondrocytes
We next examined the effect of miR-146a-5p on the expression of proinflammatory factors TNF-α, IL-1β, and IL-6. As shown in Figure 2, their expression increased significantly after IL-1β treatment (p < 0.001). Moreover, miR-146a-5p knockdown reduced the enhancing effect of IL-1β on pro-inflammatory factors and alleviated the inflammatory response (Figure 2a and 2b).
miR-146a-5p directly targeted TXNIP
To further study the injury mechanism of miR-146a-5p in IL-1β-induced chondrocyte damage, we screened the target gene TXNIP through the TargetScan website and investigated its binding site with miR-146a-5p (Figure 3a). The dual-luciferase reporter assay showed that the luciferase activity of SW1353 cells cotransfected with the miR-146a-5p mimic and wild-type TXNIP was significantly lower than that of SW1353 cells cotransfected with miR-NC and wild-type TXNIP (p < 0.001). However, there was no change in the luciferase activity of SW1353 cells cotransfected with miR-146a-5p mimic/NC and mutant TXNIP (Figure 3b). A similar result was seen in C28/I2 cells. Compared with the miR-NC group, miR-146a-5p overexpression significantly decreased the expression of TXNIP (p < 0.001; Figure 3c). Additionally, the expression of TXNIP in SW1353 and C28/I2 cells stimulated with IL-1β was significantly downregulated (p < 0.001; Figure 3d). Overall, miR-146a-5p directly targeted TXNIP and negatively regulated its expression.
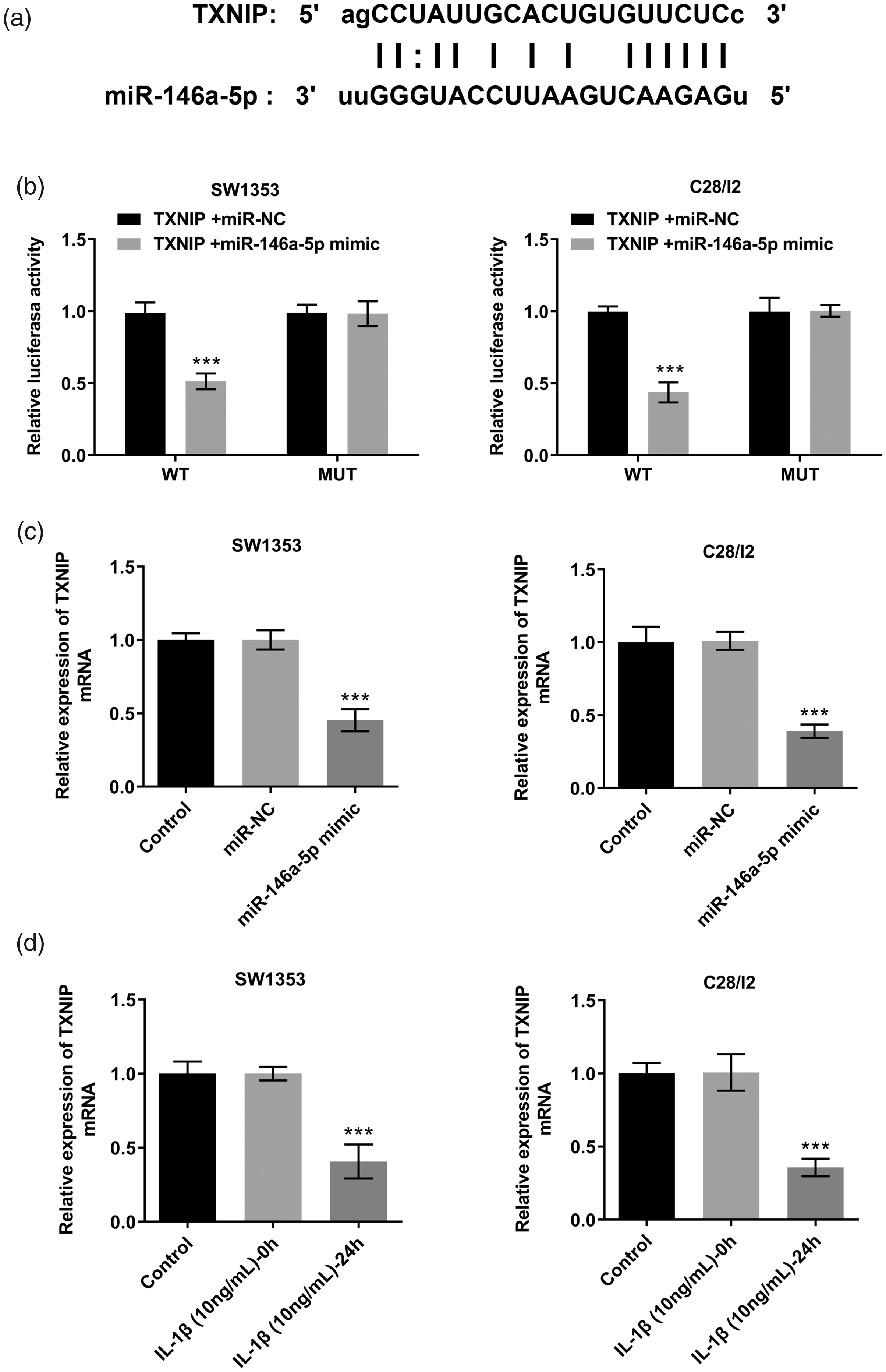
miR-146a-5p directly targeted TXNIP. (a) The predicted binding site between the 3′-UTR of TXNIP miRNA and miR-146a-5p. (b) Dual-luciferase reporter assay analysis. ***p < 0.001 vs. TXNIP+miR-NC. (c) qRT-PCR detection of TXNIP expression after miR-146a-5p mimic and miR-NC transfection into SW1353 and C28/I2 cells. ***p < 0.001 vs. miR-NC. (d) qRT-PCR detection of TXNIP expression after treatment with IL-1β. ***p < 0.001 vs. IL-1β (10 ng/ml)-0h.
TXNIP overexpression alleviated the effect of miR-146a-5p on the proliferation and apoptosis of IL-1β-stimulated chondrocytes
Next, we examined whether TXNIP was involved in miR-146a-5p-mediated IL-1β-induced SW1353 and C28/I2 cell injury. The miR-146a-5p mimic, pcDNA-TXNIP, and corresponding negative controls were transfected into SW1353 and C28/I2 cells. Western blot analysis confirmed that transfection had been effective (Figure 4a). CCK-8 and TUNEL staining results showed that miR-146-5p overexpression significantly inhibited cell proliferation and induced apoptosis, compared with the negative control group, and that these changes could be alleviated by TXNIP overexpression (p < 0.05; Figure 4b and 4c). Taken together, miR-146a-5p overexpression enhanced IL-1β-induced SW1353 and C28/I2 cell injury by downregulating TXNIP.
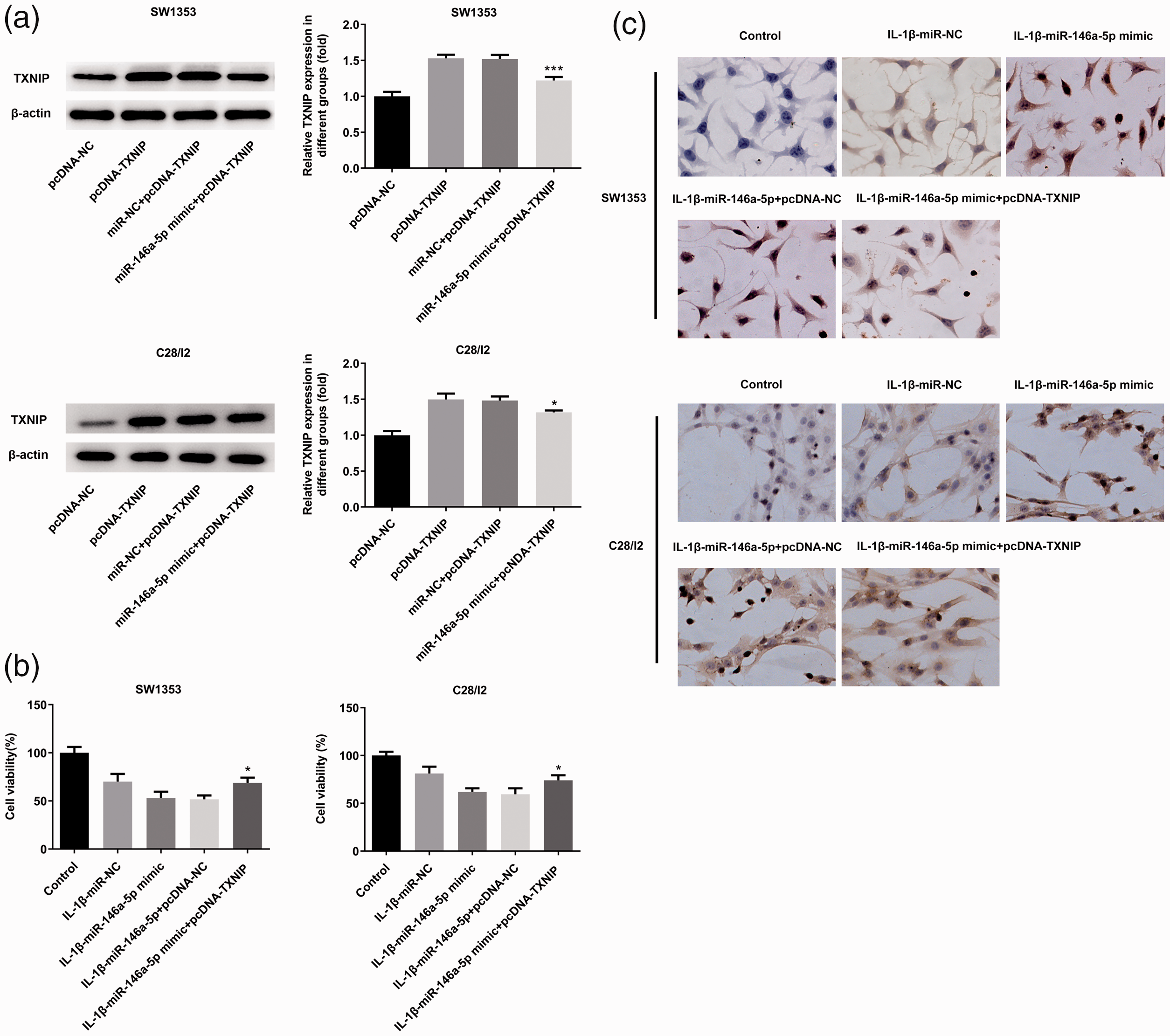
TXNIP overexpression alleviated the effect of miR-146a-5p on IL-1β-stimulated chondrocyte proliferation and apoptosis. (a) Western blot detection of TXNIP expression after miR-146a-5p mimic, pcDNA-TXNIP, and corresponding scrambled control transfection into SW1353 and C28/I2 cells. (b) Cell viability as determined by CCK-8. (c) Apoptosis as determined by TUNEL staining. *p < 0.05 and ***p < 0.001 vs. miR-NC+pcDNA-TXNIP.
TXNIP overexpression alleviated the effect of miR-146a-5p on inflammatory cytokine expression in IL-1β-stimulated chondrocytes
Finally, we tested the effect of TXNIP overexpression on the expression of proinflammatory factors TNF-α, IL-1β, and IL-6 in SW1353 and C28/I2 cells stimulated with IL-1β. Compared with the IL-1β-miR-NC group, miR-146a-5p overexpression significantly increased the expression of TNF-α and IL-1β, and IL-6, and this could be partially reversed by TNXIP overexpression (p < 0.001; Figure 5a and 5b).

TXNIP overexpression alleviated the effect of miR-146a-5p on IL-1β-stimulated chondrocyte inflammatory cytokine expression. (a) ELISA detection of TNF-α, IL-1β, and IL-6 expression in IL-1β-stimulated SW1353 cells. (b) ELISA detection of TNF-α, IL-1β, and IL-6 expression in IL-1β-stimulated C28/I2 cells. **p < 0.01 and ***p < 0.001 vs. IL-1β-miR-146a-5p+pcDNA-NC.
Discussion
Our results show that IL-1β stimulation can induce damage and increase the expression of miR-146a-5p in SW1353 and C28/I2 cells. Moreover, we found that miR-146a-5p knockdown significantly promoted cell proliferation and inhibited apoptosis and the inflammatory response, eventually improving the cellular injury induced by IL-1β. Additionally, TXNIP was shown to be a target gene of miR-146a-5p and to significantly reduce the inhibitory effect of miR-146a-5p overexpression on SW1353 and C28/I2 cell activity.
As the most common degenerative joint disease, OA has maintained a high incidence in middle-aged and older people. 17 Furthermore, with the recent intensification of social aging and unsatisfactory geriatric nursing, OA has become an issue of global concern. 18 Current intra-articular sodium hyaluronate injections effectively alleviate relieve pain but do not provide a complete cure, while opioid analgesics are addictive and have other side effects meaning that they cannot be widely used. 19
Articular cartilage destruction, hardening, and hyperosteogeny are the main causes of joint pain and motor damage in OA. 20 IL-1β plays a key role in this by promoting the expression of matrix metalloproteinases, increasing the levels of inflammatory mediators prostaglandin E2 and nitric oxide, and triggering an inflammatory response. 21 It is therefore commonly used in the construction of an in vitro OA model. We showed that IL-1β stimulation significantly enhanced SW1535 and C28/I2 cellular injury, which was characterized by decreased cell viability and increased apoptosis and inflammation.
Collagen produced by chondrocytes is an important component of the cartilage matrix, and the chondrocyte metabolic balance is disrupted in the OA microenvironment, leading to cartilage injury. 22 Several studies have documented the important role of miRNAs in regulating IL-1β-induced chondrocyte injury. For example, miR-502-5p targets TNF receptor associated factor 2 to alleviate IL-1β-induced chondrocyte injury, 23 while miR-34a regulates the proliferation and apoptosis of chondrocytes by targeting the sirtuin 1/p53 pathway. 24 Additonally, Soyocak et al. 25 reported significantly higher miR-146a-5p expression in the serum of OA patients compared with healthy controls, while miR-146a was found to participate in OA pain through IL-1 receptor-associated kinase-1, and then affect nerve cell differentiation and pain production. 26 However, the specific role and mechanism of miR-146a-5p in chondrocyte injury induced by IL-1β had not been fully studied.
In this study, miR-146a-5p was shown to be upregulated in IL-1β-stimulated SW1353 and C28/I2 cells, and miR-146a-5p knockdown significantly promoted proliferation and inhibited apoptosis and the expression of inflammatory factors in both cell types. The observed expression of miR-146a-5p is consistent with previous clinical findings, 25 and with the report that miR-146a-5p promotes chondrocyte apoptosis induced by IL-1β, 27 suggesting the potential of miR-146a-5p as a regulator of OA injury.
To further study the regulatory mechanism of miR-146a-5p in SW1353 and C28/I2 cell injury induced by IL-1β, we used bioinformatics analysis and the dual-luciferase reporter assay to identify TXNIP as the downstream regulatory gene of miR-146a-5p. TXNIP combines with thioredoxin to promote the production of reactive oxygen species (ROS) and eventually induce inflammation or apoptosis. ROS promotes TXNIP transfer from the nucleus to the cytoplasm, and induces the production of apoptosis signal-regulating kinase-1, caspase-1, IL-1β, and other molecules associated with inflammation or apoptosis. 28 In OA progression and development, the level of inflammatory mediators is generally increased, but it was unclear whether TXNIP participated in the development of OA. One study showed that TXNIP expression was significantly decreased in OA cartilage and that the regulated in development and DNA damage/TXNIP complex was necessary for autophagy activation of chondrocytes. 15
We speculate that miR-146a-5p affects IL-1β-induced SW1353 and C28/I2 cell injury by regulating the expression of TXNIP. In the current study, we found that TXNIP overexpression attenuated the effects of miR-146a-5p overexpression on cell proliferation, apoptosis, and inflammation. These results suggest that miR-146a-5p promotes the damaging effect induced by IL-1β on SW1353 and C28/I2 cells by downregulating TXNIP.
In conclusion, our study confirmed that miR-146a-5p was highly expressed in SW1353 and C28/I2 cells stimulated with IL-1β, and showed that miR-146a-5p knockdown mitigated IL-1β-induced damage of SW1353 and C28/I2 cells. Additionally, miR-146a-5p promoted IL-1β-induced chondrocyte injury by targeting TXNIP. These findings offer insights into novel treatments and prognosis biomarkers for OA.