
Abstract
Objectives
Construction of adenoviral vectors can be complicated and time-consuming. The aim of this study was to construct an adenoviral vector expressing human bone morphogenetic protein 2 (BMP-2).
Methods
An adenoviral vector expressing human BMP-2 was constructed using the Gateway™ technique based on site-specific recombination. Briefly, BMP-2 cDNA was obtained by polymerase chain reaction, inserted into the pMD19-T cloning vector, and subcloned into a shuttle vector. The DNA sequence encoding BMP-2 was then subcloned from pEC3.1-BMP2 into the pAd/BLOCK-iT™-DEST adenoviral vector genome, which was then linearized and used to transfect 293 cells for adenovirus packaging. Adenovirus particles were generated with a titer of 1 × 1010 infectious units/mL.
Results
Potent expression of BMP-2 mRNA and protein was detected in adenovirus-infected 293 cells, confirming that the adenoviral vector encoding BMP-2 was successfully constructed.
Conclusions
This recombinant adenoviral vector encoding BMP-2 can be applied in future studies to explore the roles of BMP-2 in various cell types and tissues.
Introduction
Bone morphogenetic protein 2 (BMP-2) belongs to the transforming growth factor β (TGF-β) superfamily and plays critical roles in embryogenesis, 1 development, 2 cell proliferation3,4 and differentiation.5,6 The critical role of BMP-2 in osteogenesis and chondrogenesis has been demonstrated by an increasing number of studies.7,8 However, the native form of BMP-2 is expressed at low levels in vivo and has a short half-life, making it difficult to maintain its long-term effect. With the development of molecular biotechnologies, potential clinical applications of BMP-2 gene therapies are emerging. A variety of gene delivery methods have been developed to serve specific clinical needs. Adenoviral vectors have been widely used for gene delivery because of their high transduction efficiencies both in vitro and in vivo. By contrast with lentiviruses or retroviruses, adenoviral DNA does not integrate into the host genome and therefore adenoviral vectors have been considered as a safer option for gene therapy. 9 Nevertheless, traditional adenovirus production procedures have several weaknesses including long production cycles and tedious, complicated preparation steps. Hence, more efficient construction of recombinant adenoviral vectors is a major goal of many research groups.
We constructed a recombinant adenoviral vector encoding BMP-2 using Gateway™ technology with significantly reduced production time and increased efficiency.10,11 Our work makes high level gene expression of BMP-2 possible and lays the foundation for future applications of BMP-2 in gene therapies for orthopedic conditions including bone defects12,13 as well as in bone tissue engineering.14,15
Material and methods
Primer design
Cloning primers were designed based on the mRNA sequence of BMP-2 available in Genbank (accession number BC069214) using Primer Premier 5.0 software. The sequences of cloning primers were: BMP-2-f (including BamHI restriction site): 5′-
PCR amplification of BMP2
PCR amplification of BMP-2 was performed using pOTB7-BMP2 vector (ATCC, Manassas, VA, USA) as the template. The following components were added to the reaction mixture: 2.5 μL of 10× LA PCR Buffer (containing Mg2+), 0.25 μL of LA Taq, 1 μL of dNTPs (2.5 mM), 0.25 μL of BMP-2-f primer (10 μM), 0.25 μL of BMP-2-r primer (10 μM), 1 μL of pOTB7-BMP2, and dH2O to a final volume of 25 μL. Reaction conditions were as follows: 94°C for 5 minutes; 30 cycles of 94°C for 20 seconds, 53°C for 25 seconds, and 72°C for 75 seconds; 72°C for 3 minutes. The PCR reaction was stored at 4°C. PCR products were separated by electrophoresis on a 1% agarose gel. DL 2000 Marker (Takara, Japan) was used as the molecular weight standard. The agarose gel was imaged under UV light and photographed, after which DNA was recovered using a DNA gel extraction kit (ZD Biotech, Shanghai, China).
Subcloning of BMP2 into pMD19-T cloning vector
The purified PCR product was ligated into the pMD19-T vector (Invitrogen, USA). DH5α cells (Biontex, Munich, Germany) were transformed with the ligation product. Plasmid DNA extracted from single colonies was digested with BamHI and EcoRI. The digested DNA was separated on a 1% agarose gel and clones showing DNA fragments of the expected size were sent to Invitrogen (Shanghai, China) for sequencing. Sequencing results were analyzed using Sequencher (Gene Codes Corporation, Ann Arbor, MI, USA) and BLAST to confirm that the DNA sequence encoding BMP-2 was correctly inserted into pMD19-T. The pMD19-T plasmid bearing the BMP-2 insert was designated as TS-BMP-2.
Subcloning of BMP2 into shuttle vector pEC3.1(+)
The TS-BMP-2 and pEC3.1(+) vectors were digested with BamHI and EcoRI. The digested DNA was separated on a 1% agarose gel. The BMP2 insert was purified and ligated into the pEC3.1(+) vector using T4 DNA ligase. DH5α cells were transformed with the ligation product. Plasmid DNA purified from single colonies cultured on ampicillin-containing agar plates was digested with HindIII and EcoRI. A clone showing a DNA fragment of the expected size was designated as pEC3.1-BMP-2.
Transfer of BMP-2 expression cassettes into pAd/BLOCK-iT™-DEST adenoviral expression vector
The BMP-2 expression cassette was transferred into the pAd/BLOCK-iT™-DEST adenoviral expression vector via in vitro homologous recombination mediated by Gateway LR Clonase™ II. The reaction components were as follows: 3 μL of pEC3.1-BMP2, 0.5 μL of pAd/BLOCK-iT™-DEST adenoviral vector (0.25 μg/μL), 2 μL of Gateway LR Clonase™ II Enzyme Mix, and Tris-ethylenediaminetetraacetic acid buffer (pH 8.0) to a final reaction volume of 10 μL. The reaction was incubated at 25°C for 1 hour, then 1 μL of proteinase K was added to digest the recombinase at 37°C for 15 minutes. DH5α cells were transformed with 5 μL of the reaction mixture, then spread onto agar plates containing ampicillin and incubated at 37°C overnight. Single colonies were used to inoculate LB medium containing ampicillin and grown at 37°C overnight. Plasmids were extracted using a Plasmid Extraction Kit (Qiagen, Valencia, CA, USA) and identified via their banding patterns following restriction enzyme digestion. Digested DNA products were separated on a 1% agarose gel and the recombinant vector bearing the BMP-2 expression cassette was designated as pAd-BMP2.
Adenovirus packaging and production
To ensure the replication and packaging efficiency of pAd-BMP2, the plasmid was linearized with PacI and purified successively using phenol, chloroform and isopentanol. After centrifugation, the supernatant was mixed thoroughly with ethanol to precipitate DNA, and the pellet was washed and dissolved in dH2O. HEK293 cells (American Type Culture Collection, USA) were transfected with linearized adenoviral DNA using METAFECTENE™ transfection reagent (Biontex, Germany). HEK293 cells were observed for cytopathic effect (CPE) for 7 days post-transfection. When more than 50% cells had detached from the bottoms of culture flasks, the cells were collected, subjected to three freeze-thaw cycles, and stored at −74°C.
Amplification and characterization of recombinant adenoviral vector
HEK293 cells was transduced with supernatants from lysed cells. When CPE was observed, cells were freeze–thawed, centrifuged and their supernatants collected. This step was repeated several times to obtain high-titer adenovirus. DNA from infected HEK293 cells was extracted and used as template for PCR amplification with BMP-2-specific primers. Titration of adenoviruses was performed using the Adeno-X™ Rapid Titer Kit (Takara, Shiga, Japan).
Results
PCR amplification of BMP2 gene and cloning into pMD19-T vector
Human BMP2 was amplified by PCR using an existing pOTB7-BMP-2 vector as template. The expected PCR amplicon size was 1.2 kb and electrophoresis showed a band around 1 kb (Figure 1a). The purified PCR product was ligated into the pMD19-T Simple Vector. Following digestion with EcoRI and BamHI to release the inserted DNA fragment, the banding pattern of DNA subjected to agarose gel electrophoresis indicated successful cloning (Figure 1b). The inserted DNA fragment was also sequenced and analyzed using BLAST (data not shown). Its sequence was consistent with the GenBank entry for BMP2. These results confirmed that the TS-BMP-2 recombinant vector harboring the human BMP2 gene was successfully constructed.

Agarose gel electrophoresis showing PCR amplification of human BMP2 and cloning into the pMD19-T vector. (a) PCR product amplified using BMP-2 cloning primers. Lane 1: DL2000 Marker; Lane 2: BMP2 PCR product. (b) Lane 1: DL2000 Marker; Lane 2: double restriction enzyme digestion of recombinant TS-BMP-2 vector.
Construction of recombinant pEC3.1-BMP-2 vector
The DNA fragment derived from double digestion of the TS-BMP-2 vector was inserted into the pEC3.1(+) shuttle vector, yielding the recombinant pEC3.1-BMP-2 vector. The expected sizes of DNA fragments released from this vector by double digestion with HindIII and EcoRI were 4400 bp and 1200 bp. Following digestion, we observed DNA fragments around 4400 bp and 1000 bp (Figure 2), confirming that the recombinant pEC3.1-BMP-2 vector was successfully constructed.
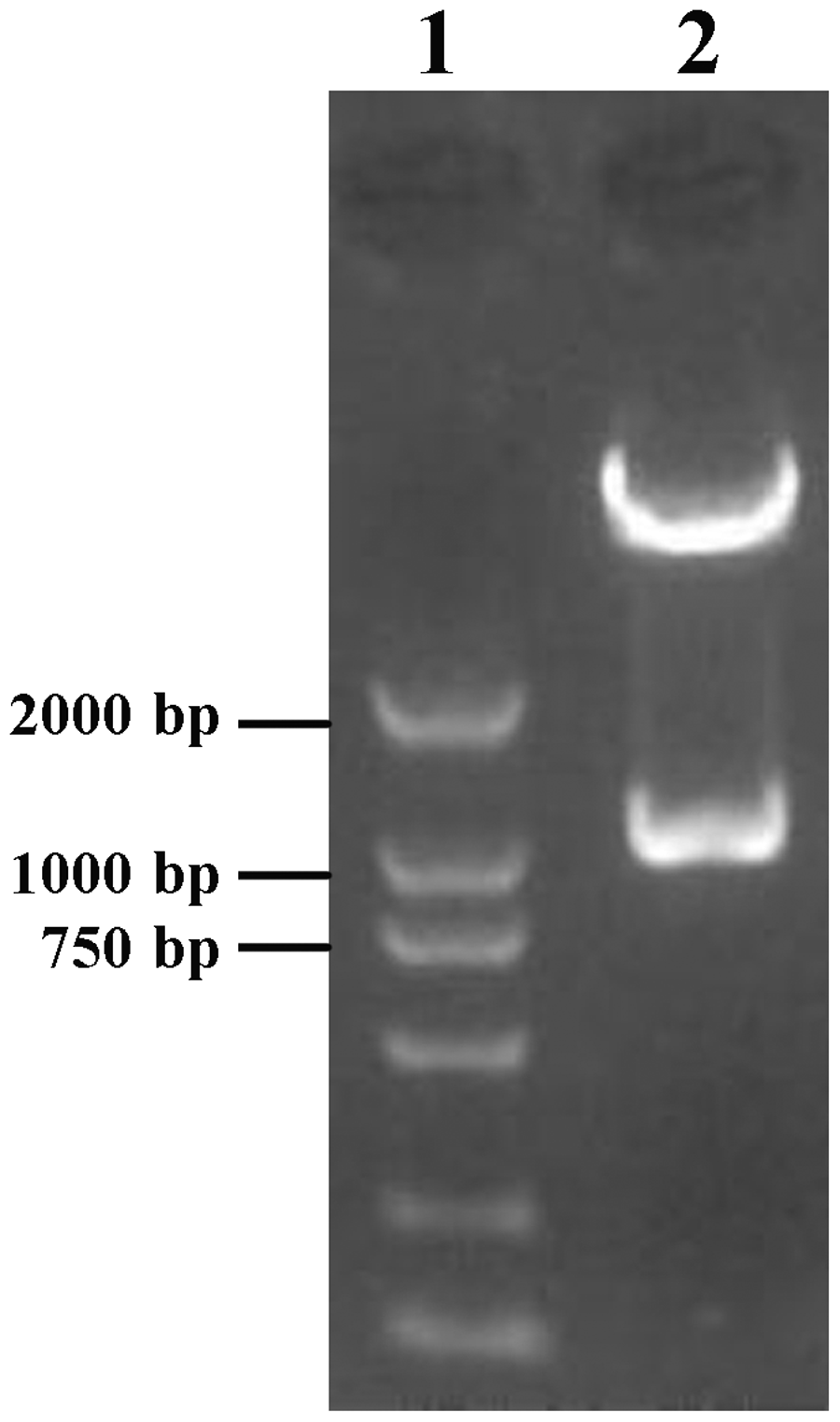
Agarose gel electrophoresis of recombinant pEC3.1-BMP-2 vector double-digested with HindIII and EcoRI. Lane 1: DL2000 Marker; Lane 2: DNA fragments following double restriction enzyme digestion.
Construction of recombinant pAd-BMP-2 vector
The pEC3.1-BMP-2 and pAd/BLOCK-iT™-DEST adenoviral vectors underwent homologous recombination mediated by LR Clonase™ II. DH5α competent cells were transformed with the resulting recombinant vector. Adenoviral DNA was extracted from single ampicillin-resistant colonies, and BMP2 was amplified by PCR (Figure 3a). The recombinant adenoviral vector was also digested with XbaI, resulting in one DNA fragment larger than 15 kb and one of approximately 3 kb, consistent with the expected sizes (Figure 3b). These results confirmed that the recombinant pAD-BMP-2 vector was successfully constructed.
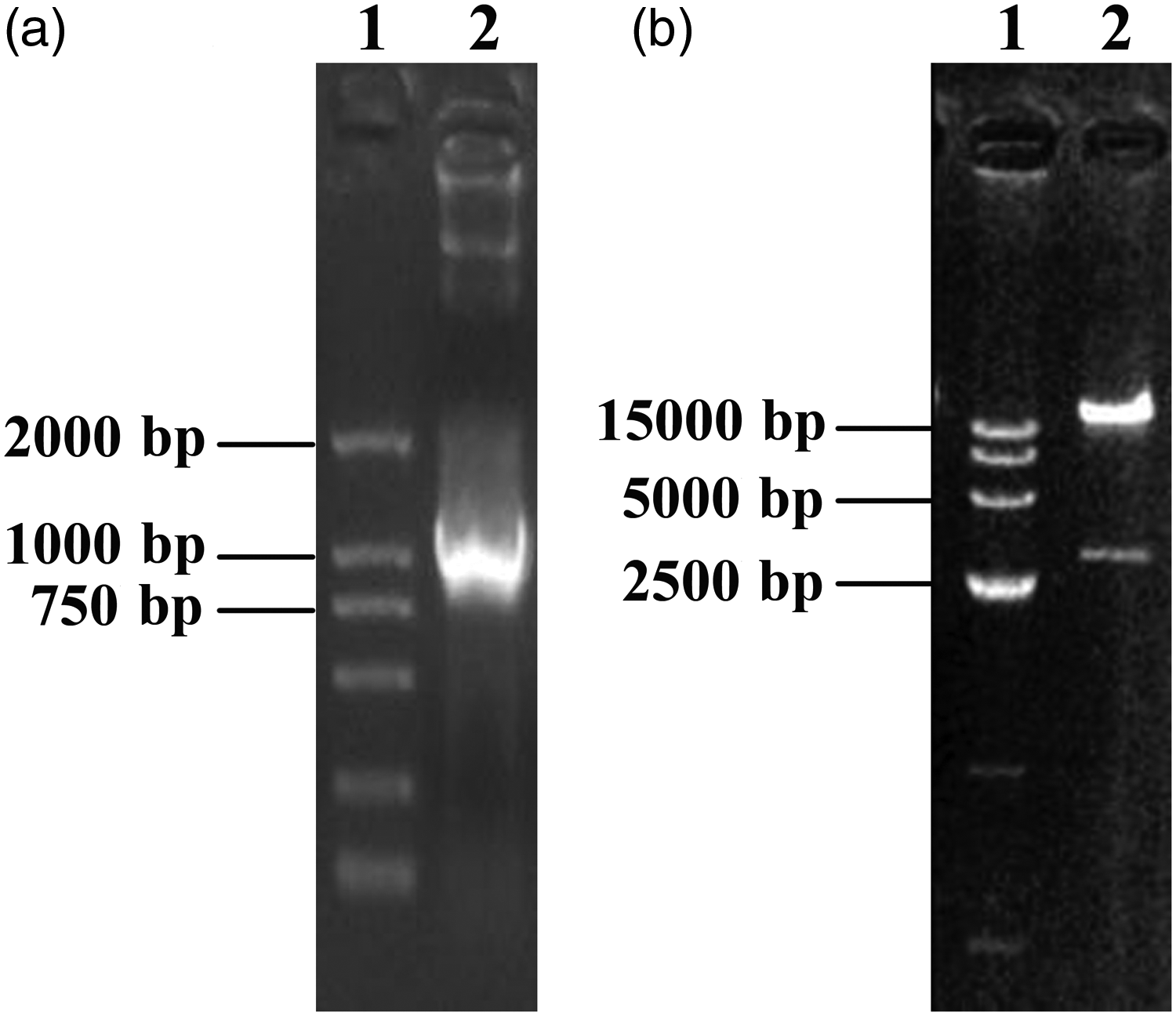
Agarose gel electrophoresis confirming subcloning of the BMP-2 expression cassette into the adenoviral vector. (a) Lane 1: DL2000 Marker; Lane 2: PCR amplification of BMP2 from the recombinant adenoviral vector. (b) Lane 1: DL15000 Marker; Lane 2: recombinant adenoviral vector digested with XbaI.
Preparation and characterization of the recombinant adenovirus
The pAd-BMP-2 vector was linearized by PacI digestion and used to transfect HEK293 cells. The transfected cells were observed for CPE. Adenovirus particles were purified from culture medium and viral DNA was precipitated with ethanol. The presence of BMP2 in adenoviral DNA was confirmed by PCR. A 1.2 kb PCR amplicon was observed by electrophoresis, suggesting that purified adenoviral DNA contained the BMP-2 expression cassette (Figure 4a). Lysates from infected HEK293 cells were analyzed by western blotting, which showed a single band corresponding to the size of BMP-2 (Figure 4b). The recombinant adenoviral vector was designated Ad-BMP-2, and had a titer of 1 × 1010 infectious units/mL. These results indicated that a potently infective adenoviral vector harboring the human BMP2 gene was successfully generated.

Agarose gel electrophoresis (a) and western blotting (b) confirming cloning of the BMP-2 expression cassette into the adenoviral vector. (a) Lane 1: DL15000 Marker; Lane 2: PCR amplification of BMP2 from adenoviral DNA. (b) Lane 1: HEK293 cells infected with an adenoviral vector lacking DNA sequences encoding BMP-2; Lane 2: HEK293 cells infected with Ad-BMP-2.
Discussion
Methods for recombinant adenoviral vector construction are typically complicated and time-consuming, with low success rates. 16 Developed by Invitrogen, the BLOCK-iT™ adenovirus construction system, based on phage λ site-specific recombination, applies Gateway™ technology. Compared with previous methods, this technology achieves fast gene cloning in different expression systems and offers higher recombinant adenovirus construction efficiency. 17
Accumulating studies on BMPs indicate that these proteins can induce mesenchymal stem cells to differentiate into chondrocytes or osteoblasts.18,19 BMPs also play an important role in healing of bone defects or fractures. BMP-2 is a critical regulator of many physiological processes and represents a promising potential target for future clinical applications. However, recombinant BMP-2 is easily degraded by proteases in vivo and thus is only able to maintain its biological activity for a short period. One proposed mechanism of BMP-2-induced chondrogenesis/osteogenesis is that BMP-2 protein produced in vivo triggers the release of more BMP-2 from chondrocytes and osteoblasts. Secreted BMP-2 accumulates in the cartilage matrix and is absorbed by chondrocytes. Together with exogenous BMP-2, elevated expression of endogenous BMP-2 induces mesenchymal stem cells to differentiate into chondrocytes/osteoblasts, which form new bones. 20 Maintaining consistent, steady expression of BMP-2 in vivo has been an obstacle to clinical applications and adenoviral vectors are being explored as gene delivery tools to solve this problem. 21 Adenoviruses enjoy the advantages of high in vitro and in vivo infection efficiency, tropism for various types of cells, successful infection of both non-dividing and dividing cells, and non-incorporation into the host genome. 22 Therefore, adenoviral vectors have been widely applied in gene therapies for various diseases.
Three main types of strategies are currently available for recombinant adenoviral vector construction: (i) the Adeno-X system developed by Clontech based on direct in vitro ligation; (ii) the AdEasy system developed by He et al, 23 in which a linearized shuttle vector and backbone vector are used to co-transform Escherichia coli and subsequently undergo homologous recombination; and (iii) the AdMax system, in which recombination occurs in HEK293 cells. Direct in vitro ligation is a traditional method with low recombination efficiency. Additionally, the limited number of enzyme cleavage sites for cloning in adenoviral genomes restricts applications of the Adeno-X system. The AdEasy and AdMax systems both rely on homologous recombination but have the disadvantages of complicated protocols, time-consuming clone screening, low recombination efficiency and the potential generation of wild-type adenoviruses. Therefore, development of novel, convenient, and efficient adenoviral vector construction strategies is urgently needed.
In our study, we employed Gateway™ technology, a system different from the recombination strategies mentioned above. Gateway™ technology is based on λ phage site-specific recombination (attL × attR→ attB × attP). In the presence of LR Clonase II, attL and attR undergo precise site-specific recombination, yielding a plasmid product that contains attB and attP sites. We successfully subcloned the BMP2 gene fragment into the shuttle vector pEC3.1(+), which contains two attL sites (attL1 and attL2). Subsequently, this shuttle vector encoding BMP2 (designated as pEC3.1-BMP2) underwent recombination with the replication-deficient adenoviral backbone vector pAd/BLOCK-iT™-DEST. In this reaction, recombination occurred between the attL sites (attL1 and attL2) in pEC3.1-BMP2 and the attR sites (attR1 and attR2) in pAd/BLOCK-iT™-DEST, resulting in the insertion of BMP2 into the pAd/BLOCK-iT™-DEST vector and replacement of the attR gene that the vector originally contained. This reaction, which leaves open reading frames unchanged, is strictly conservative and eliminates the possibility of vector construction failure resulting from base mutations. The reaction is highly efficient, requiring only a 1 hour incubation at 25°C. This technology also uses the attR selection method: when recombination occurred between the pAd/BLOCK-iT™-DEST vector and the pEC3.1-BMP2 plasmid, the ccdB fragment in pAd/BLOCK-iT™-DEST was replaced by BMP2, yielding pAd-BMP2 and a byproduct plasmid carrying attP. DH5α cells were transformed with the reaction products and cultured on ampicillin-containing agar plates. Only cells bearing the pAd-BMP2 plasmid, but not those bearing pAd/BLOCK-iT™-DEST or the recombination byproduct, were able to grow on the ampicillin plates. Compared with other established methods, this procedure avoids complicated screening for recombinant clones and repeated purification steps, making isolation of the recombinant Ad-BMP2 vector fast and simple. Using Gateway™ technology, we constructed the Ad-BMP2 vector quickly and efficiently in preparation for future applications of BMP-2 in development of gene therapies for orthopedic diseases. We estimate that Gateway™ technology saved us about 1 to 3 weeks compared with traditional cloning methods. Gateway™ technology relies on homologous recombination and therefore does not require restriction endonuclease digestion or ligation of DNA fragments, which saves significant time by circumventing multiple cloning and subcloning steps. Additionally, after homologous recombination mediated by LR Clonase II, only bacteria bearing plasmids encoding the gene of interest will survive. Empty vectors bear the ccdB gene, which encodes gyrase poison. Hence, bacteria bearing empty vector cannot grow on agar plates. This greatly enhances the efficiency of screening for positive clones, as the effort required to pick and sequence positive clones is significantly shortened.
We chose adenoviral vectors over other vectors because of their numerous advantages. We used second-generation adenoviral vectors, bearing deletions of the E1 and E3 genes and thus having decreased viral genome sizes, for our experiments. This not only improves biosafety but also enables insertion of larger (up to 6.5 kb) exogenous gene fragments. Thus, second-generation vectors are more adaptable and stable. Following transduction, the adenoviral vector we used resulted in expression of the target protein for 1 to 2 weeks,17,24 rendering it suitable for in vivo studies. Despite the advantages mentioned above, adenoviral vectors have weaknesses such as high immunogenicity,25,26 which remain to be solved in the future.
Footnotes
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Ethics approval and consent to participate
The study was approved by the Ethics Committee of The First Hospital Affiliated of Shenzhen University, Shenzhen Second People’s Hospital.
Funding
This work was supported by the Shenzhen Science and Technology Innovation Council (Nos. JCYJ20150330102401096, GCZX2015043017241191, and JCYJ20160226192924528), the National Natural Science Foundation of China (Nos.81672234 and 81902303), the Fund for Health and Family Planning Commission of Shenzhen Municipality (No. SZBC2017022) and the Shenzhen Science and Technology Project (No. 201606018).