
Abstract
Objectives
Human herpesvirus 8 (HHV-8) is transmissible and causes Kaposi’s sarcoma and other malignancies. This study analyzed the seroepidemiology and phylogeny of HHV-8 among 515 injection drug users (IDUs) and 229 men who have sex with men (MSM) in Taiwan.
Methods
Blood and peripheral mononuclear cells were analyzed for HHV-8 seroprevalence using enzyme-linked immunosorbent and immunofluorescence assays. Viral loads were measured using a real-time PCR assay. Phylogenetic analysis of the K1 gene was performed using nested PCR and DNA sequencing.
Results
HHV-8 infection rate was higher in MSM (24.9%) than in IDUs (3.8%). The rate of HHV-8 infection was higher in HIV-1-positive patients (32.8%, MSM; 5.5%, IDUs) than in HIV-1 negative patients. HHV-8 load was not significantly different between HHV-8 seropositive and seronegative patients. HHV-8 genotypes C and A variants were detected at frequencies of 80% and 20%, respectively, among IDUs; and genotypes C, D, E, and A were detected at frequencies of 55.6%, 11.1%, 11.1%, and 5.6%, respectively, among MSM. Variants of K1 amino acid residues 54–84 were detected in most IDUs and MSM.
Conclusions
HHV-8 prevalence was significantly higher among MSM than among IDUs. Evolution of the K1 gene occurred in HHV-8 variants of IDUs and MSM.
Introduction
Human herpesvirus 8 (HHV-8), also known as Kaposi’s sarcoma-associated herpesvirus, causes Kaposi’s sarcoma, primary effusion lymphoma, and multicentric Castleman’s disease.1–4 HHV-8 coevolves within humans and spreads horizontally. HHV-8 prevalence varies geographically as follows: 5% in Asia, northern Europe, Australia and the Americas and >20% in Mediterranean countries and sub-Saharan Africa. 5 The distribution of HHV-8 genotypes based on K1 gene sequences (A–F and Z) also varies geographically.6–9 Genotypes A and C are prevalent in Europe, Australia, and the United States. Genotype D infects aboriginal people of the Pacific Rim.10,11 Genotypes A, B, C, and F are detected in patients infected with human immunodeficiency virus-1 (HIV-1), and genotype B correlates with the prognosis of Kaposi’s sarcoma. 12
The association of HHV-8 infection with drug-injecting behavior demonstrates that sharing syringes is a significant mode of transmitting HHV-8, similarly to HIV-1, hepatitis B virus, and hepatitis C virus.13–16 The prevalence of HHV-8 antibodies and genotypes is important for public health but is infrequently studied in injection drug users (IDUs). The Increase in HHV-8 seroprevalence is significantly associated with the duration of drug injection, although demographics vary.17,18 Further, the high HHV-8 seroprevalence in Greek IDUs is significantly associated with poor personal hygiene related to drug injection. 19 We demonstrated that the prevalence of HHV-8 is higher in HIV-1-positive MSM (10.6%) compared with that of HIV-1 positive IDUs (7.1%) in Taiwan, indicating that sex and HIV load are associated with HHV-8 infection of HIV-1-positive patients. 20 Sequence analysis of the open reading frame (ORF) of the K1 gene is used to analyze the subtypes or genotypes of HHV-8, 12 although little information is available on the distribution of subtypes or genotypes among IDUs.
Here we performed analyses of seroprevalence, viral load, and the phylogeny of HHV-8 among IDUs and MSM in Taiwan. Antibodies against HHV-8 lytic antigens in IDUs and MSM were detected using an enzyme-linked immunosorbent assay (ELISA), and samples with antibody titers = 1:40 were confirmed using an immunofluorescence assay. The virus load of peripheral mononuclear cells (PBMCs) was quantitated using a real-time PCR assay with HHV-8 K6-specific primers. The hypervariable regions (K1) of the HHV-8 genome were determined using direct nucleotide sequencing.
Materials and methods
Study population
The study recruited 744 patients (515 IDUs and 229 MSM) treated at the Taipei-Municipal Venereal Disease Control Institution or the Taipei-veterans General Hospital in northern Taiwan. All participants provided verbal informed consent and agreed to our use of their demographic data and medical records. The Institutional Review Board of National Yang-Ming University approved this study (Institutional Review Board No. 2010A007). The demographic data and medical records included age, sex, family history, height, weight, dates of HIV-1 detection, disease onset, administration of antiviral drugs, virus load, CD4+ T cell count, hepatitis virus B and C markers, blood glucose, low-density lipoprotein, triglycerides, blood pressure, and opportunistic infections. All patients with long-term stable disease provided venous blood and PBMCs every 6 months for HHV-8 detection assays. The samples were stored at –80°C.
Serological testing and CD4+ T cell counts
The HHV-8 Whole Virus Lytic IgG ELISA (Advanced Biotechnologies, Inc.; Columbia, MD, USA) exhibited high sensitivity and specificity with no cross-reactivity with Epstein-Barr virus or other human herpesviruses.21–23 The ELISA was used to rapidly screen for HHV-8 antibodies against viral lytic antigens. Each serum sample was diluted 40-fold according to the protocols provided by the source. The cutoff point for seropositivity was defined as an optical density at 450 nm >0.05. An HHV-8 immunofluorescence assay (Advanced Biotechnologies Inc.) was used for to confirm the detection of antibodies against HHV-8 lytic antigens in most patients.21–23 A 40-fold dilution of each serum sample was prepared for the immunofluorescence assay, and slides with >5 fluorescent cells were used to determine seropositivity. CD4+ T cell counts were determined using a FACSCalibur (Becton, Dickinson and Co., Franklin Lakes, NJ, USA) after incubating venous blood with Tritest CD3/CD4/CD45 Reagent (Becton, Dickenson, and Co.) according to the manufacturer’s instructions.
Preparation of DNA
DNAs were extracted from PBMCs using the Qiamp Blood Kit (Qiagen Gmbh, Hilden, Germany).4,24 DNA from HHV-8-infected cells (primary effusion B lymphoma cells, BC-1) (American Type Culture Collection [ATCC], Manassas, VA, USA) served as the positive control, and DNA isolated from the T-lymphocyte HH cell line (CRL-2105) served as the negative control.
Real-time quantitative PCR
HHV-8 DNA was quantitated using a real-time PCR assay with HHV-8 K6-specific primers as described in previous studies.25,26 The amount of endogenous retrovirus-3 in each sample was used to assess DNA quantity and quality. The reactions were performed using an Applied Biosystems Prism 7000 Sequence Detection System (Perkin Elmer, Foster City, CA, USA).
HHV-8 K1 nested PCR
An 841-bp fragment of the K1 gene was amplified using nested PCR.4,7 DNA (1 µg) was added to the first-cycle PCR reaction, and 1 µL of the PCR product was placed into the second-round PCR reaction mixture. Both assays included 20 pmol/µL of each primer, 125 µM of each dNTP, 10x PCR buffer (MgCl2), and 0.5 U Taq polymerase, and first- and second-round PCR reactions were performed as previously described.27,28
Phylogenetic analysis
The fragments amplified using nested PCR were directly sequenced using the ABI PRISM BigDye Primer Cycle Sequencing Ready Reaction Kit and an ABI Prism 377 DNA Sequencer (both from Perkin Elmer). The nucleotide sequences were submitted to GenBank (accession numbers FJ866506 to FJ866528). Alignments of K1 gene sequences of HHV-8 variants were examined using the DNASTAR MegaAlign clustal method (DNASTAR, Madison, WI, USA). The sequence of HHV-8 type M (GenBank locus U75698) served as a positive control. Phylogenetic neighbor-joining trees were built based on 1000 bootstrap replicates with Kimura’s z-parameter distance matrix using MEGA (Version 3.0) and PHYLIP (Version 3.6) software.29,30 PHYLIP parsimony and maximum-likelihood methods were verified using the topologies of taxa shown in the trees. 31
Statistical analysis
Distributions of the characteristics of MSM and IDUs were compared using the χ2 test. Fisher’s exact test was used for small sample sizes. Odds ratios (OR) and 95% confidence intervals (CIs) were used to evaluate the association between HIV-1 and HHV-8, and the Breslow-Day method was used to test the homogeneity of ORs between MSM and IDUs. Because the distribution of the HHV-8 viral load was extremely skewed, the median is presented, and the Wilcoxon rank sum test was used to compare the median values of MSM and IDUs. SAS statistical analysis software Version 9.1 (SAS Institute, Inc.; Cary, NC, USA) was used for all analyses. P < 0.05 indicates statistical significance. 32
Results
Demographics and seroepidemiological analysis
Analysis of patients’ demographics revealed that 67.0% of IDUs and 54.6% of MSM were HIV-1 positive, 83.2% of IDUs were male, and 42% of IDUs were 30 to 39 years (Table 1). The majority of IDUs were junior high school graduates (53.9%), and one-third of IDUs were laborers. In the HIV-1(+)MSM group, 48% were 30 to 39 years, and 68.0% were college graduates. Approximately 50% of HIV-1(+) MSM did not report their occupations. The seroprevalence of HHV-8 infections in IDUs and MSM is shown in Table 2. Among MSM and IDUs, HHV-8 seropositive rates were 12.8% and 6.2% among HIV-1(+) and HIV-1(−) patients, respectively (Table 2). The seroprevalence of HHV-8 was 3.8% in IDUs and 24.9% in MSM. Among IDUs, HIV-1(+) patients had a significantly higher rate of HHV-8 infection compared with HIV-1(−) patients (5.51% vs 0.6%, OR = 9.85, 96%CI = 1.31–74.2). Among MSM, HIV-1(+) patients had a significantly higher rate of HHV-8 infection compared with HIV-1(−) patients (32.8% vs 15.4%, OR = 2.68, 95%CI = 1.40–5.15). However, the ORs of MSM and IDUs were not significantly different (Table 2).
Patients’ demographic characteristics

IDUs, injection drug users; MSM, men who have sex with men.
The association between HIV-1 and HHV-8 among MSM and IDUs in Taiwan

IDUs, injection drug users; MSM, men who have sex with men.
HHV-8 viral load in PBMCs
The CD4+ T cell counts were not significantly different between patients with and without HHV-8 infection. MSM had significantly higher CD4+ T cell counts compared with those among IDUs (500 vs 439, P = 0.0221). The distribution of the HHV-8 viral load was extremely skewed. Therefore, the Wilcoxon rank sum test was used to judge the difference between median values of MSM and IDUs. IDUs had significantly higher median HHV-8 viral loads compared with those of MSM (5170 vs 500, P = 0.0174). However, the HHV-8 viral loads were not significantly different between HHV-8 seropositive and seronegative patients (Table 3).
CD4+ T cell counts and HHV-8 viral loads in MSM and IDUs with and without anti-HHV-8 antibodies
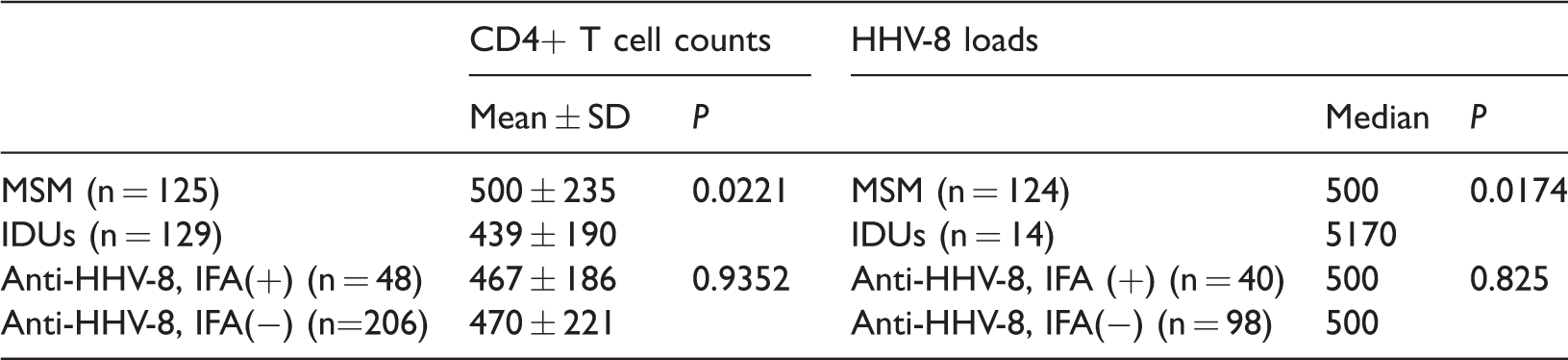
IDUs, injection drug users; MSM, men who have sex with men; IFA, immunofluorescence assay; SD, standard deviation.
K1 nested PCR and phylogenetic analyses
The HHV-8 K1 sequences of nested PCR products from 5 IDUs (Taiwan Yang-Ming University Kaposi’s sarcoma associated virus [TYKS] 1, 5, 17–19) and 18 MSM (TYKS 2–4, 6–16, 20–23) were directly sequenced. In addition, 46 published sequences were used to construct the phylogenetic tree (Figure 1). Of the 5 IDU sequences, 1 was genotype A (TYKS 1), and the other 4 (TYKS 5, 17–19) were genotype C. Further, 10 (55.6%) sequences of the 18 HHV-8 variants of MSM were genotype C (TYKS 3, 4, 6–16), 2 (11.1%) were genotype D (TYKS 20, 21), 2 (11.1%) were genotype E (TYKS 22, 23), and 1 (5.6%) was genotype A (TYKS 2).
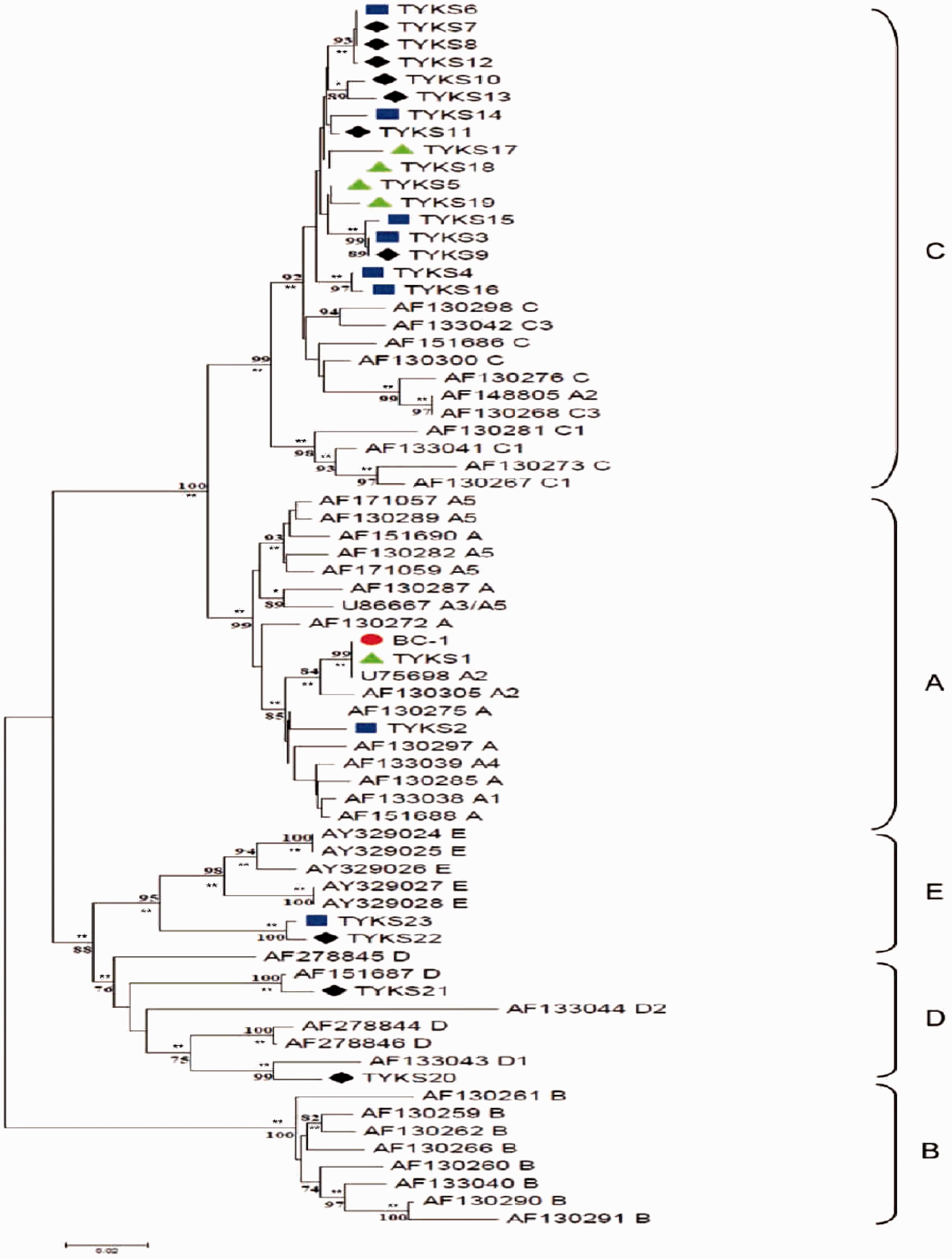
Predicted phylogenetic distribution of a 561-bp segment of HHV-8 K1 amplified from MSM and IDUs with HIV(+) among a background of GeneBank sequences representing the major ORF K1 subtypes. Linear unrooted phylogenetic dendrograms were generated using the neighbor-joining method. Bootstrapping of 1000 replicates is indicated as a percentage at major branch points, and P values are labeled on each node (*P < 0.05 and **P < 0.01). The genetic distance, size scale = 0.02 (2% divergence) is indicated. •: BC-1 (primary effusion B lymphoma cells, positive control); ▲: IDUs (TYKS 1, 5, 17–19); ♦: MSM (TPE-VGH, TYKS 7–13, 20–22); ▪: MSM (MVDCI, TYKS 2–4, 6, 4–16, 23); TYKS, Taiwan Yang-Ming University Kaposi’s sarcoma associated virus; MVDCI, Taipei-Municipal Venereal Disease Control Institution; TPE-VGH, Taipei-Veterans General Hospital.
Variability of the K1 gene of HHV-8 variants
The deduced amino acid sequences of the K1 gene were based on the HHV-8 variants (18 MSM and 5 IDUs) (Figure 2). Positive control (BC-1), U75698, and TYKS 1 patients had 99% amino acid sequence similarity with all clones of IDU patients. TYKS 2 patients from MSM had nine notable amino acid residue changes in strains of BCBL-R (T37N, F58L, P60L, E62K, T63K, F66L, P67G, and T69N) in 85% of the clade’s genes (Figure 2). Compared with amino acid residues 54–84 of the prototype (BCBLR, AF133038), most Taiwanese isolates (TYKS 1–23) had 8–19 changes in the K1 variable region.

Sequence analyses of the K1 variable region, VRI loop, identify genotypic diversity in HHV-8 detected in IDUs and MSM. The K1 region was PCR-amplified from PBMC DNAs, sequenced, and aligned with representative K1 genotypes 11 . Dashes indicate identities.
Discussion
The present study demonstrates that HHV-8 infections of IUDs and MSM were significantly associated with HIV-1 infection (Table 2), revealing that coinfection with HHV-8 and HIV-1 was transmitted through sexual behavior and blood exposures. Among patients with HIV-1 infection, the HHV-8 infection rate of MSM was significantly higher compared with that of IDUs (32.8% vs 5.51%) (Tables 2, 3). IDUs had a higher median HHV-8 viral load compared with that of MSM (5170 vs 500) (Table 3). However, the HHV-8 viral load was not significantly different between HHV-8 seropositive and seronegative patients. The route of sexual contact, but not injecting drugs, is significantly associated with the prevalence of HHV-8 in HIV-1-positive patients. 20 A similar finding was made here in that there was a difference in the seroprevalence of HHV-8 between MSM who were IDUs or not.
Sexual risk factors are associated with the higher seroprevalence of HHV-8 among IDUs,13,33 and there is a significant association of HHV-8 infection with individual risk factors in citizens of Amsterdam, such as sexual routes or needle sharing. 16 HHV-8 viremia in IDUs causes repeat exposure of IDUs who are HHV-8-negative; thus the injecting drug route is a high risk factor for blood-borne transmission of HHV-8 compared with that of blood donors. 15 Similarly, HIV-positive IDUs have a higher incidence of Kaposi’s sarcoma compared with that of the general population. 34 Further, there is controversy about the correlation of the HHV-8 viral load with antibody titers against HHV-8 lytic antigens.35,36 The present study did not find this correlation (Table 3). This may be explained by the high percentage of specimens from MSM with low HHV-8 viral loads.
The high level of intratypic variability between amino acid residues 54 and 84 within the VR loop of ORF-K1 protein is mainly genotype specific (Figure 2). The VR loop of ORF-K1 protein is structurally similar to those of several hypervariable loops within HIV-1 gp160 (e.g. V3 and V4). 11 All subtype ORF-K1 protein patterns are hypervariable within VR1 and particularly dominant within the A and C genotypes (Figure 2). However, fewer substitutions at individual residues, but not distinct short in-frame deletions or repeats, were identified within the VR2 region (data not shown). Phylogenetic analysis of the K1 sequence indicates that the frequencies of HHV-8 genotype-C variants were 80% in IDUs and 55.6% in MSM (Figure 1). The result is consistent with the frequencies of patients residing in the United States, Europe, Australia, Asia, and Brazil. 6
In the present study, single infection with HHV-8 was detected in 16 (7%) MSM and one (0.2%) IDU (Table 2), indicating that these HHV-8 strains spread through different transmission routes, differing from the virological and genetic properties of those coinfected with HHV-8 and HIV-1. However, the sample size was low and therefore insufficient to design a robust experiment with statistical significance. Importantly, salivary contact may represent a classical route of HHV-8 transmission. 37 Therefore, our future studies will evaluate the significance of differences between HHV-8 strains in single- and co-infected patients, including transmission routes (blood, sex, and salivary contact), antibodies against lytic and latent antigens, and viral loads in plasma and PBMCs.
Conclusion
In summary, a higher prevalence of HHV-8 was discovered among MSM compared with IDUs in northern Taiwan. Coinfection with HHV-8 and HIV-1 was detected among the MSM and IDUs. Further, the phylogenetic tree of the ORF-K1 protein indicates its high polymorphism, providing important insights into the epidemiology of HHV-8 infection among MSM and IDUs. Our findings reveal the evolution of HHV-8 K1 in the life cycle of HHV-8.
Footnotes
Authors’ contributions
LYM analyzed data and prepared the manuscript. HPS analyzed the data and revised the manuscript. LCW designed and coordinated the study and wrote the manuscript. All authors corrected the final version of the manuscript.
Acknowledgments
We thank the staff of the Genome Research Center at National Yang-Ming University for their technical support, and our colleagues at the AIDS prevention and Research Center of National Yang-Ming University and Division of Clinical Virology, Department of Pathology and Laboratory Medicine, of Taipei-VGH for helpful discussions and support.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
This study received grants from the Republic of China (ROC) Taipei-Veterans General Hospital (VGH: C327, V95A-077, VGHUST 98-P6–27); the ROC Centers for Disease Control (Number 03734246); National Yang-Ming University Hospital, Yilan (RD2010–021); the Ministry of Education, Aim for the Top University Plan; and the China Medical University (CMU106-ASIA-06, and CMU106-BC-1).