
Abstract
It is generally accepted that collaboration yields better performance than when the same task is performed by individuals operating alone. Is collaboration always superior to individual performances, or do its benefits depend on whether collaborating individuals have the same or different roles? Here, participants performed tasks to transport a virtual beam while keeping it horizontal. Twin robotic manipulanda were coupled to the simulated beam with either symmetric or asymmetric dynamics. In paired modes, two participants grasped the robot handles, whereas in solo modes, individual participants grasped both handles with their two hands. Computational simulations revealed that this task could be successfully performed regardless of whether roles were the same or different for each beam dynamics. However, pairs performed better than individuals after two participants spontaneously assumed different roles, co-adapting by learning in asymmetric dynamics, with one participant leading the movement and exerting more force than the other. In contrast, individuals performed better than pairs if pairs did not assume different roles. Our findings suggest that better performance by pairs than individuals is not a general phenomenon, but that collaboration only yields better performance if role specialization spontaneously emerges in such interactions.
Keywords
Novelty Statement
This study marks a significant contribution to understanding human–human physical cooperation by revealing, for the first time, that superior performance in paired individuals (dyads) than in single agents (solos) is not a general phenomenon, as suggested by previous literature using pre-determined roles, but rather that collaboration yields better performance only when role specialization emerges spontaneously in dyadic interactions. This finding contributes to the field of human–robot interaction by showing that modulation of physical cooperation conditions through robotic interfaces can significantly impact task performance by either hindering or facilitating spontaneous emergence of roles. This novel framework for exploring underlying mechanisms of physical human–human collaboration can be used to design algorithms to improve performance of human–robot interactions in various settings, including industrial and clinical applications where smooth, high-level collaborations are required.
1. Introduction
The field of human–robot interaction has been identified as one of the grand challenges in robotics (Yang et al., 2018). The growing field of human–robot interaction research has leveraged principles from psychology, biomechanics, motor, and cognitive neuroscience. Although significant advances have been made in this field to date, human–robot interactions are far from resembling natural human–human interactions (Kragic and Sandamirskaya, 2021).
For this challenge to be successfully addressed, research has focused on solving several interrelated problems (Lorenzini et al., 2023; Sciutti et al., 2018; Thomaz et al., 2016), including, but not limited to inferring human intention (Breazeal et al., 2009; Butterfield et al., 2009), co-adaptation of robotic and human actions (Ehrlich and Cheng, 2018; Van Zoelen et al., 2021), and designing actuators and sensors that enable safe and natural interactions (Cherubini and Navarro-Alarcon, 2021; Cid et al., 2014; Tiboni et al., 2022). Research on human ability to interact seamlessly with each other has inspired research on controllers for human–robot interactions (Beyan et al., 2020; Liu et al., 2023), for example, theory of the mind (Premack and Woodruff, 1978; Sebanz et al., 2006; Tomasello et al., 2005), and reinforcement learning (Lee et al., 2012). Related research has been devoted to improving collaboration between two agents. Therefore, discovering factors that optimize performance of human–robot pairs remains a major research goal. Human–human collaboration research has been extensively investigated in the past two decades in a variety of contexts, ranging from social to physical interactions (Jarrassé et al., 2012; Konvalinka and Roepstorff, 2012; Schmidt et al., 1998; Takagi et al., 2017; Varlet et al., 2014). Understanding factors that impact task performance when two or more individuals cooperate has significant implications, not only for understanding sensorimotor mechanisms, but also in contexts in which a human operator interacts with a robot.
It is generally assumed that human–human collaboration can lead to better performance than efforts made by individuals. This phenomenon, also known as the “assembly bonus effect” (Collins and Guetzkow, 1964), in which group performance exceeds the performance of individual group members, has been reported in various types of collaboration, for example, decision-making tasks (Bahrami et al., 2010), language acquisition (Storch, 1999), cards sorting tasks (Fawcett and Garton, 2005), and programming tasks (Balijepally et al., 2009).
In the domain of haptic collaboration, that is, actions performed jointly by physically-interacting individuals, investigations regarding whether collaborating participants can outperform individual participants have yielded mixed results (Wahn et al., 2018). Some studies demonstrated that paired individuals perform better than single participants. Reed et al. (2006) found that pairs can perform a target acquisition task with a rotational crank faster than individuals. Ganesh et al. (2014a), using a two-dimensional continuous target-tracking task, found that paired performances resulted in smaller target tracking errors than solo performances. However, other studies did not find performance advantages in dyads over solos. Van der Wel et al. (2011) reported that pairs performed a pendulum swing task at the same level as solos. In tasks that require keeping an object stable against gravity, several studies have found that pair performance was, on average, similar to individual performance (Burstedt et al., 1997; Mojtahedi et al., 2017a; Solnik et al., 2016).
There are several factors that potentially account for differences in these findings. One may be the number of effectors involved in the task. Pairs tend to outperform individuals if the task can be performed unimanually by one individual (Feth et al., 2009; Ganesh et al., 2014a; Oosterhout et al., 2018; Reed et al., 2006). This can be explained by biomechanical advantages associated with involvement of more effectors during paired cooperation, that is, two arms from two participants versus one arm from one participant. Another factor might be the extent to which paired individuals can focus on control of different aspects of the task, that is, role specialization (Wahn et al., 2018). This factor could be particularly important when two effectors are required to perform a given task. For instance, Oosterhout et al. (2018) assigned two participating arms asymmetric and independent roles: one arm controlled the vertical motion of the object, whereas the other arm controlled the horizontal motion in a teleoperation task. This study demonstrated superior performance by pairs over bimanual individuals when roles are pre-assigned. However, it remains unknown whether role specialization that emerges spontaneously during collaboration benefits pairs more than individuals. Spontaneous role specialization has been observed in different physically coupled motor collaboration tasks (Bosga et al., 2010; Groten et al., 2009; Melendez-Calderon et al., 2015; Reed and Peshkin, 2008; Takai et al., 2023), but these studies did not examine individual performance of bimanual tasks.
To address these unknowns, we designed an experimental study involving virtual beam transport tasks that had to be completed by two effectors belonging to one participant (bimanual solo mode) or two participants (paired mode) to minimize the beam’s tilt. Importantly, dynamics of the virtual beam could be altered to give the two participating effectors symmetric or asymmetric implicit mechanical leverage. One of our recent studies on pairs showed that when asymmetric mechanical leverage was given, stable and consistent leader–follower role specialization emerged spontaneously by co-adaptation through learning among all pairs and gradually improved task performance (Takai et al., 2023). In that previous study, we used a task that allowed a large variety of successful solutions that provided implicit gradients for optimizing paired motor behavior. The very existence of such optimization gradients led to gradual co-adaptation, culminating with convergence to a consistent strategy. Following systematic exploration of the solution space, asymmetric mechanical leverage favored role emergence and led pairs to co-adapt and converge upon similar solutions whereby one of the participants (the one assigned a shorter moment arm) always spatiotemporally led the other participant (the one assigned a longer moment arm). These asymmetric contributions by pairs suggest that role specialization occurred when the short-moment-arm participant assumed the leader’s role, according to the common lead-lag definition of leaders in joint motor tasks (see literature review in Supplemental Material, Quantitative Identification of Participant Roles). Moreover, the short-moment-arm participant also exerted greater force and exhibited more corrective force than the long-moment-arm participant. These findings reveal that consistent leader–follower relationships emerge by co-adaptation through learning from implicit haptic interactions. Importantly, computational simulations of dyadic interactions under these asymmetric dynamics revealed that role specialization was not forced as a mechanically obligatory phenomenon (Takai et al., 2023; see solution space density maps in Supplemental Figure 3). Furthermore, another recent study of ours demonstrated that no consistent role specialization was observed when symmetric mechanical leverage was given to pairs (Mojtahedi et al., 2022). Therefore, we hypothesized that spontaneous role specialization is a necessary condition for superior performance, which only occurs with asymmetric, but not symmetric dynamics in our task. We tested this hypothesis by investigating the performance of paired and individual participants under asymmetric versus symmetric dynamics.
Consequently, the novelty and importance of this study lie in examining performance in relation to role specialization. Although spontaneous role specialization has been observed in various tasks, it remains unclear whether this benefits paired performance more than solo performance. Previous studies have been unable to correlate roles with performance due to a lack of control conditions. To address this, we purposefully designed two task conditions: one that inherently favors asymmetric roles and one that does not. This experimental design enabled us to directly contrast paired and individual performances in scenarios in which role specialization may or may not spontaneously arise. As a result, we found that the “assembly bonus effect” occurred only when clear role specialization emerged after learning by pairs. Individuals outperformed pairs when no clear role specialization was seen. Therefore, we conclude that a key factor underlying superior performance in pairs is spontaneous role specialization that emerged through adaptive coordination. Finally, we emphasize that this conclusion can be obtained using only data from asymmetric dynamics, because only late trials with different roles showed the assembly bonus effect. Thus, symmetric dynamics results are not essential for the main conclusion of this study.
2. Methods
2.1. Participants performed a virtual beam transportation task under symmetric and asymmetric dynamics
We implemented a virtual beam transportation task (Figure 1(a)–(c)), which was controlled by moving a pair of robotic manipulanda (TVINS, Figure 1(d)–(h)) (Ganesh et al., 2014a, 2014b; Honda et al., 2013). In the paired mode, both participants grasped the robot handle with their right hands. Robots and seated participants were separated by a wall such that they could not see each other (Figure 1(g)). In the bimanual individual mode, robots were placed close to each other to allow individual participants to grasp robot handles with both hands (Figure 1(h)). All experiments in this study were based on physical simulation of the virtual beam, and they did not employ a real physical object. Positions of the two handles and the force applied to them controlled the movement of the virtual beam. Simulated physical interaction force provided haptic feedback to participants through the manipulanda (see Supplemental Material, Virtual Beam Equations). Visual rendering of beam movement was projected on a display surface located above the robot and participant arms. Moreover, robots were programmed to move in only one direction (forward or backward with respect to participants; y-direction, Figure 1(c)). Task, manipulanda, and experimental settings. (a) Task dynamics of the left-pivoted beam (asymmetric dynamics). The virtual beam, shown as a solid black line, behaved as if its rotational point (pivot, dashed black circle) was located at the left far end. Participant hands (red and blue dotted circles) are connected to the beam ends (black dots) via simulated spring-dampers. The distance between each handle was 1 m. The distance from the pivot to the left handle was 9 m. We defined this experimental situation as “asymmetric” based on the unequal distance between each handle and the pivot. (b) Task dynamics of the center-pivoted beam (symmetric dynamics). This virtual beam behaved as if its pivot was located at the center (the pivot-to-hand distance for both hands was the same); therefore, we defined this experimental situation as “symmetric.” (c) Visual feedback of the task. The visualized beam was shown as a solid black line. Participants transported the beam from the start position (black dots) to the target location (green circles). Beam tilt was rendered as a yellow dot sliding along the beam (sensitivity: 0.04 m/degree). (d) A single robotic manipulandum consisted of an air-magnetic, floating, direct-driven parallel link with a force sensor at the end-effector handle. The opaque table used to display visual feedback is not shown in these photos (d, e, f, h). (e, f) Twin visuomotor and haptic interface system (TVINS) used for paired modes. Two manipulanda were grasped independently by two participants. (g) A black board was placed between the two participants to prevent visual interaction. Participants were informed of the task goal and criteria (the maximum absolute beam tilt and the range of transportation time) and they were told that they were going to perform different tasks. (h) Two manipulanda were moved to the center table and were grasped by one participant in individual mode. Individual participants were informed of the task goal and criteria as done with participants performing in paired modes.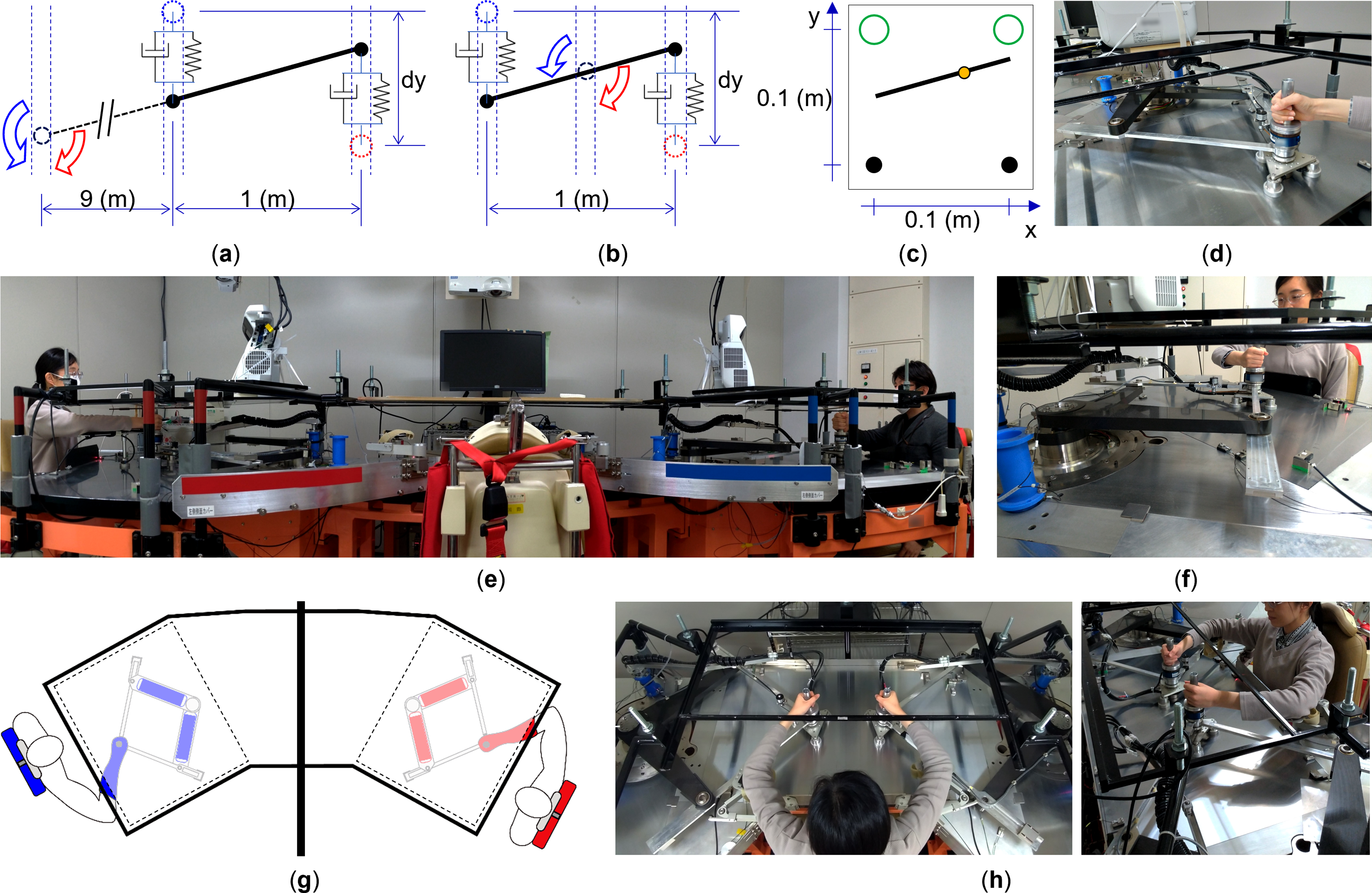
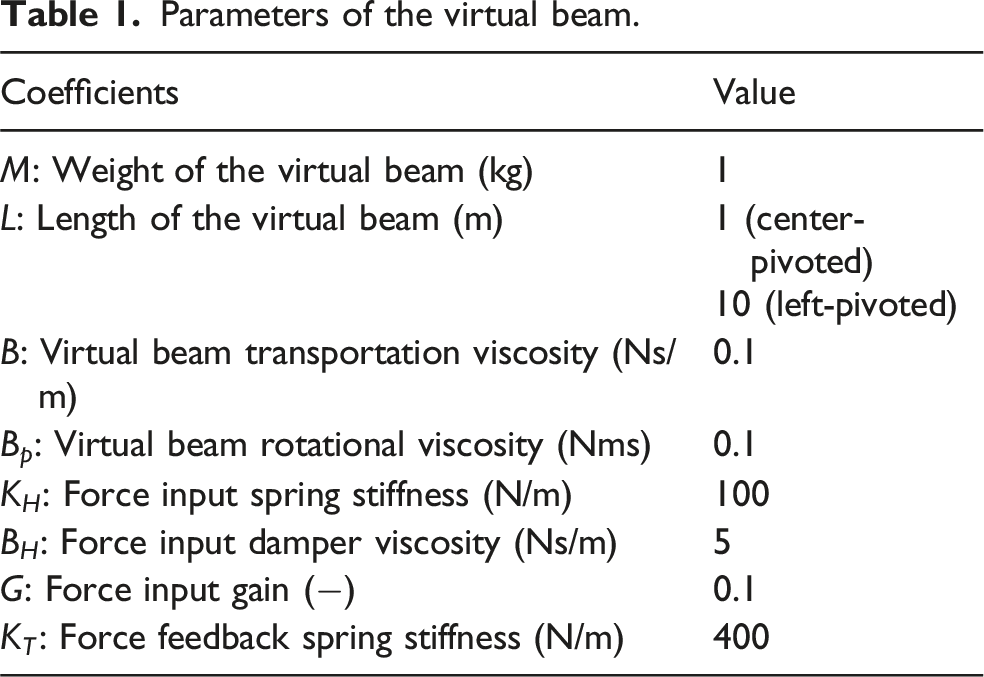
Parameters of the virtual beam.
For all task settings, participants were asked to move the virtual beam forward over the target location and to keep the beam as horizontal as possible. Participants were instructed that the task would stop if the tilt exceeded a certain threshold determined by the experimenter (see Supplemental Material, Experimental Procedure). The experiment continued until participants performed 40 successful trials or a maximum of 250 total trials.
Participants in paired modes were told that they were going to perform different tasks. Specifically, after both participants entered the room, the experimenter gave the following instructions regarding the task in Japanese: “During the experiment, you will not be able to see each other. You will wear headphones so that you cannot hear each other. You will be conducting the experiment simultaneously, but since these are different experiments, please proceed and ignore the other person. We cannot provide detailed explanations about the experiment during the session. We appreciate your understanding.” We gave these instructions because we wanted participants to focus on task performance, rather than encouraging them to cooperate. Furthermore, this instruction to participants in paired modes matches how the participants individually approached performance of the bimanual task in individual modes.
3. Results
All participants were able to perform 40 successful trials before reaching the maximum number of 250 trials (see Supplemental Material, Experimental Procedure, Supplemental Figure 1(c)). The maximum number of trials was 123 (one pair, symmetric dynamics), whereas the smallest number of trials was 40 (one pair, asymmetric dynamics). The mean number of trials ±standard deviation for all subjects was 50.6 ± 13.1. Extension 1 shows the performance of the virtual beam transport task by a pair in symmetric dynamics. Performance success or failure, defined by complying with both temporal and beam angle requirements, was displayed immediately after each trial through a text message. For transportation time, participants received feedback only at the end of each trial, with no real-time feedback during the task. For beam angle, participants received two types of feedback regarding trial failure: (1) Participants were explicitly informed through a text message whether a trial was successful only at the end of each trial. This provided clear, trial-level feedback after task completion. (2) Throughout the trial, participants could continuously monitor the yellow dot (Figure 1(c)) representing the beam angle in real-time. These constraints and performance feedback were used to encourage participants to find a strategy that would enable them to adjust their movements on a trial-to-trial basis. Extension 2 shows the same pair performing the task in asymmetric dynamics. Although visualization of the virtual beam was the same for participants, the underlying dynamics differed from those shown in Extension 1. Shortly after several failed trials, participants learned to transport the virtual beam while minimizing its tilt to a greater extent than in symmetric dynamics. Extension 3 shows one participant from the pair shown in Extension 1 performing individual trials in symmetric and asymmetric dynamics. In individual modes, single participants held the robot handles with both hands to match the number of effectors (two human arms) involved in paired modes.
3.1. Pairs performed better than individuals in asymmetric, but not in symmetric dynamics
Figure 2 shows the beam angle from representative trials from all experimental settings (individual vs paired and asymmetric vs symmetric). When participants performed the task in individual modes, the maximum absolute beam angle was smaller in a more consistent fashion across trials in asymmetric than symmetric dynamics (Figure 2(a) and (b), respectively). In contrast, pairs exhibited a gradual trial-by-trial improvement in performance by co-adaptation through learning in asymmetric dynamics (Figure 2(c)). Later trials colored red exhibited smaller beam angles than earlier trials colored blue, and also rainbow-like color distributions showed a gradual decrease of the beam angle. This was not observed in symmetric dynamics, which were also characterized by larger beam angles (Figure 2(d)). From these representative examples, we made three important observations. First, performance improved with trials by co-adaptation through learning only in paired, asymmetric settings, consistent with our previous study (Takai et al., 2023). Second, for symmetric dynamics, individuals outperformed pairs. Third, for asymmetric dynamics, pairs outperformed individuals. Representative task performance for asymmetric and symmetric dynamics. Data are from successful trials performed by one pair and one single participant from the same pair who holds the handle connected to the long-moment arm beam end. (a) Beam angle in the individual asymmetric setting. (b) Beam angle in the individual symmetric setting. (c) Beam angle in the paired asymmetric setting. (d) Beam angle in the paired symmetric setting. Trials from the 1st to the 40th are color coded (blue to red; see inset below). Rainbow-like color distribution along the number of trials, seen only in (c): Paired asymmetric, indicates that co-adaptation through learning occurred and that the maximum absolute beam angle decreased as co-adaptation proceeded. Each plot shows data from 40 successful trials.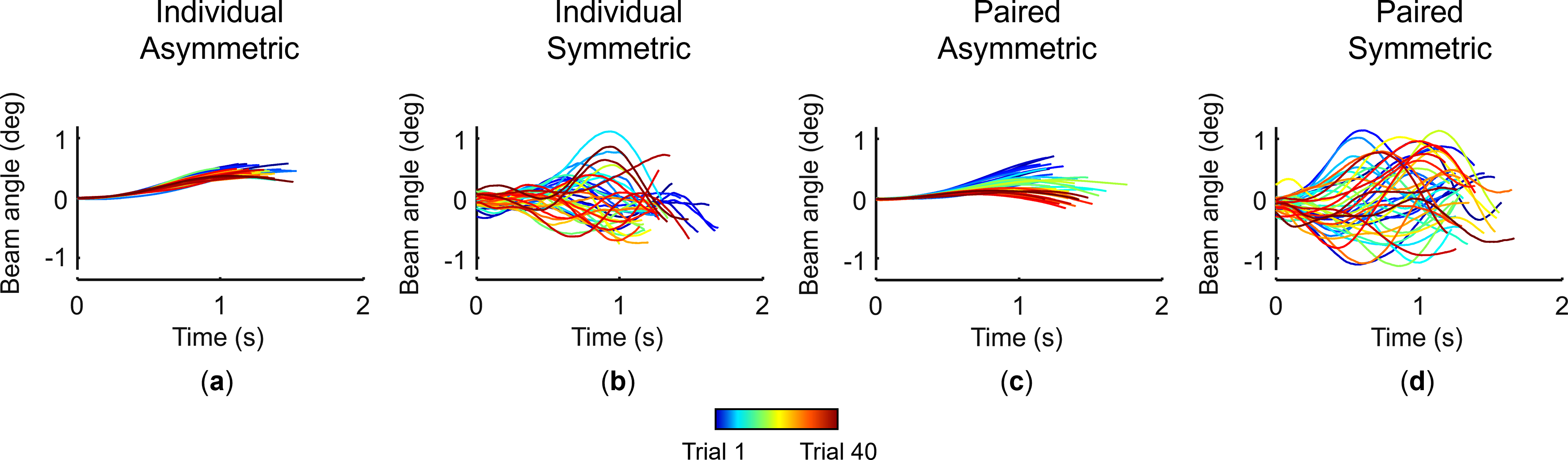
To compare spatial performance of the task between individuals and pairs systematically and comprehensively, we examined differences in maximum absolute beam angles. Pairs tilted the beam less than individuals in asymmetric dynamics, but tilted it more than individuals in symmetric dynamics (main effect of Dynamics: F (1,13) = 104.49, p < 0.001, Figure 3(a)). Furthermore, for asymmetric dynamics, one-sample t-tests revealed that pairs exhibited significantly smaller tilt than individuals in the last five successful trials (t (13) = −4.27, p = 0.007), but not in the first five. In contrast, pairs exhibited significantly larger tilt than individuals in symmetric dynamics for both the first and last five successful trials (t (13) = 5.51, p = 0.001 and t (13) = 4.71, p = 0.003, respectively). Paired and individual performances in asymmetric and symmetric dynamics. (a) Between-condition differences (paired minus individual) in the mean maximum absolute beam angle in the first and last five successful trials in each dynamics (Asymmetric or Symmetric). (b) Between-condition differences in mean transportation times in each dynamics. (c) Between-condition differences in the total number of trials in each dynamics. Data shown in panels are from 16 participants (eight pairs). Gray dots denote each participant’s difference in the metric (individual performance was subtracted from his/her paired performance). Asterisks denote differences statistically non-zero (n.s.: no difference). *, **, and *** denote p < 0.05, 0.01, and 0.001, respectively. Whiskers extend to minimum and maximum values.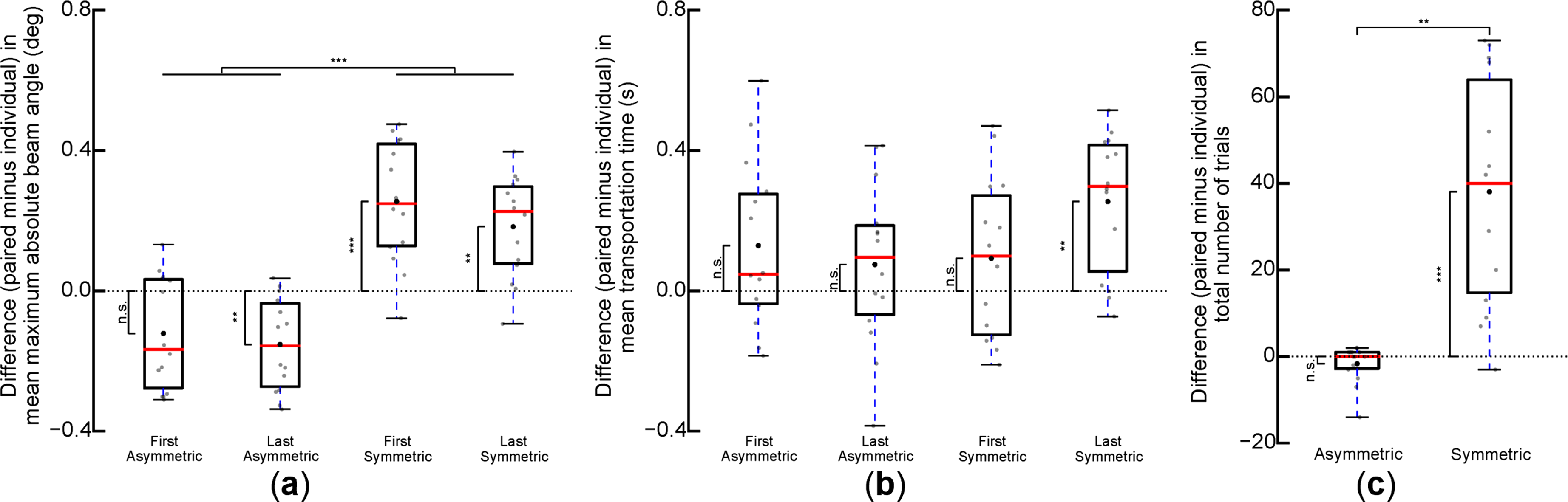
To compare temporal performances of individuals and pairs during the task, we examined differences in beam transportation time. We found a significant Dynamics × Trial interaction (F (1,13) = 6.33, p = 0.026, Figure 3(b)). However, post-hoc comparisons did not reveal significant differences between conditions after Bonferroni correction. One-sample t-test showed that transportation time for pairs was significantly longer than individuals only in the last five trials of symmetric dynamics (t (13) = 4.79, p = 0.003), and no difference was found in other conditions.
Lastly, for overall task performance, the total number of trials required to accomplish 40 successful trials did not differ significantly between individuals and pairs in asymmetric dynamics. However, pairs required more trials than individuals in symmetric dynamics (Figure 3(c)). One-sample t-tests revealed a significantly larger number of trials in the paired than in individual performance of the symmetric dynamics (t (13) = 5.41, p < 0.001). Furthermore, the total number of trials of pairs compared to individuals in symmetric dynamics was significantly larger than in asymmetric dynamics (W (13) = 0.00, p = 0.003). Pairs achieved significantly less beam tilt than individuals, although there did not appear to be differences in transportation time or number of trials, in asymmetric dynamics. Pairs generally performed better than individuals in asymmetric dynamics, whereas individuals performed better than pairs in symmetric dynamics.
3.2. Role specialization emerged during performance under asymmetric, but not under symmetric dynamics
Although individuals and pairs performed similar movements in our beam transport task, their movement execution differed in task asymmetric and symmetric dynamics. Figures 4 and 5 show representative time plots of paired performances regarding position and force in asymmetric and symmetric dynamics, respectively. A difference in the participant position is also apparent (Figure 4(a) and (b)), that is, the difference between movements of two participants gradually became more evident over time in asymmetric dynamics. Specifically, for asymmetric dynamics, long-moment-arm and short-moment-arm participants applied forces in opposite directions (Figure 4(c)). In contrast, these differences disappeared in symmetric dynamics (Figure 5(a)–(d)). These observations indicate that two distinct coordination strategies were observed despite the two task dynamics requiring achieving the same task goal: one involving role differentiation and the other where no role differentiation emerged. We conducted computational simulations to confirm that role specialization is not an obligatory consequence of asymmetric dynamics. Hand position and velocity are defined by the Minimum Jerk model (Supplemental equations (6) and (7)) in simulations. Simulated hand’s positions are substituted into equation (4) as the force input to the virtual beam, solving the equation of motion of the beam under the same conditions as the experiment. While the position and velocity of the hand are considered, the force sensor value in equation (4) is set to zero, considering only kinematics (see simulation details explained in Supplemental Material, Computational Simulation of Feasible Coordination Strategies). Representative handle position and force trajectories from paired performances: asymmetric dynamics. (a) Short-moment-arm (left) and long-moment-arm (right) handle positions. (b) Difference between left- and right-handle positions. (c) Forces applied to the left and right handles. (d) Difference between forces applied to the left and right handles. Each plot shows data from 40 successful trials. Data from the same pair as in Figure 2. (e) Relationships between mean hand position difference (abscissa) and maximum absolute beam angle (ordinate) for simulated (gray area) and experimental results (symbols). The positive zone of the abscissa indicates that short-moment-arm participants are leaders, while the negative zone indicates that long-moment-arm participants are leaders. The zero zone indicates that there was no leader–follower relationship. The gray area shows successful simulated trajectories sampled from 571,536 simulations. For experimental results, each pair (n = 7) is denoted by a different symbol. Each experimental data point represents a mean of blocks of five trials. Warmer colors indicate later trials. Representative handle position and force trajectories from paired performances: symmetric dynamics. (a)–(d) show experimental data from the same representative dyad shown in Figure 4 performing our task in symmetric dynamics. (e) shows experimental and simulated results in the same format as Figure 4(e).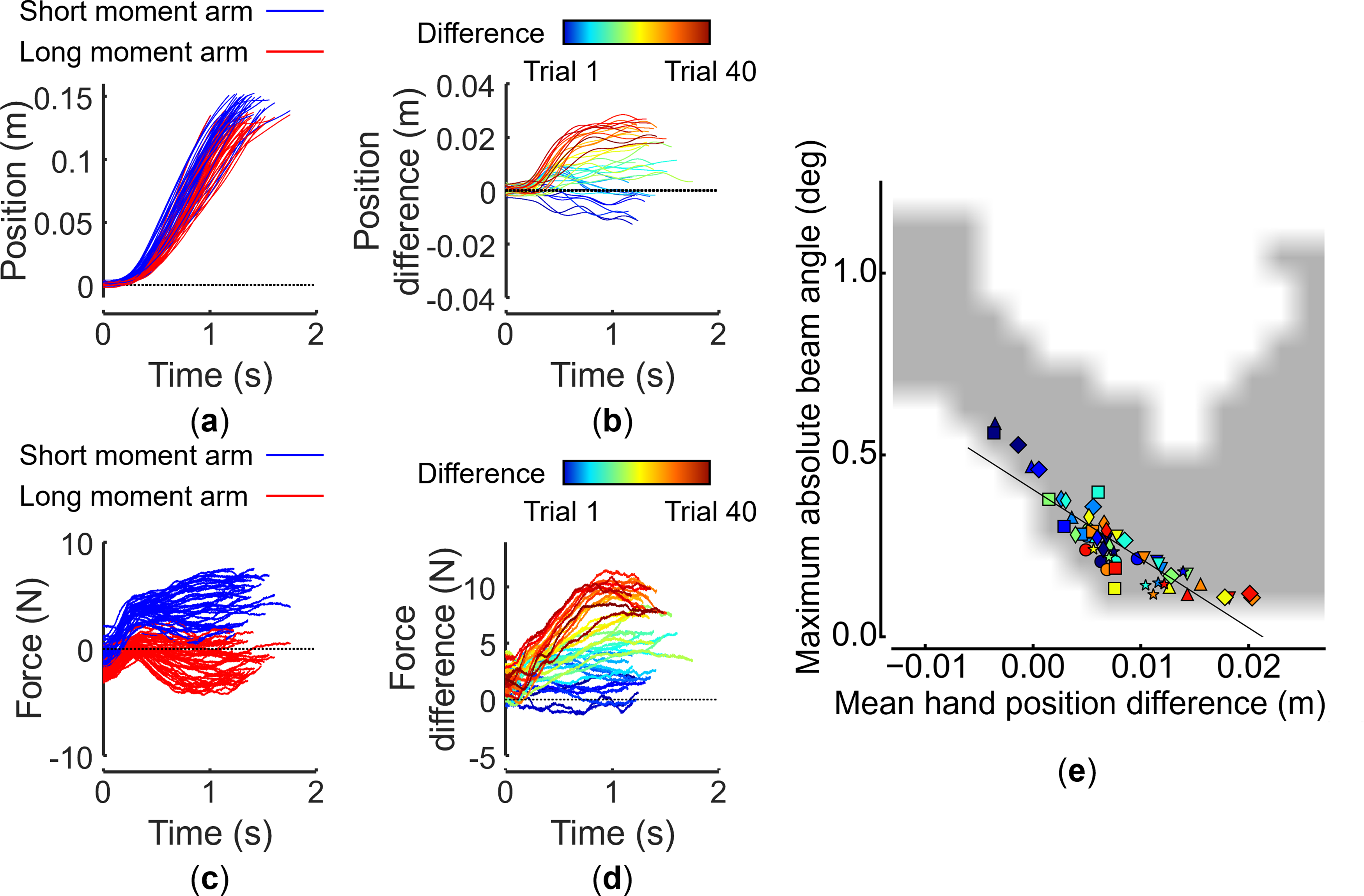
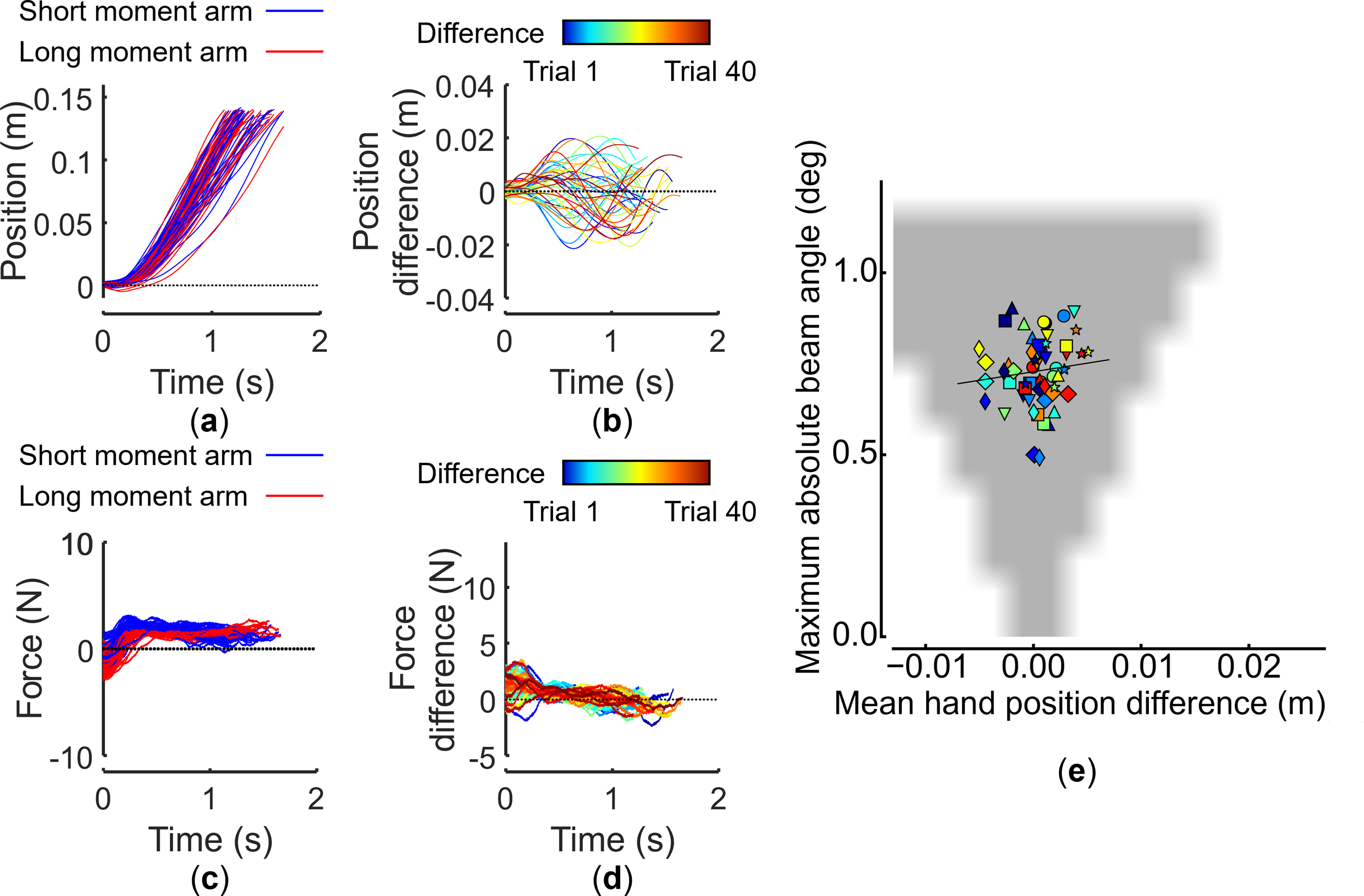
Computational simulation of these settings revealed that the two dynamics (symmetric and asymmetric) allow a large number of viable coordination strategies. This phenomenon is depicted in Figures 4(e) and 5(e) as widely distributed simulated successful trials (gray area), demonstrating the broad range of feasible solutions, that is, two participants could have behaved equally (no role-taking), or either participant could have taken the role of leader or follower for both symmetric and asymmetric dynamics. Therefore, these results indicate that successful performance of our task could have been accomplished regardless of role specialization in either task dynamics. However, in asymmetric dynamics (Figure 4(e)), although most participants chose the same action strategy in earlier trials (blue symbols), they shifted to taking different roles and contributing to the task using different actions in later trials by co-adaptation through learning (red symbols). In contrast, in symmetric dynamics (Figure 5(e)), participants randomly chose the same or different strategies in earlier and later trials.
To quantify spatial coordination between two hands, we subtracted the right handle position from that of the left handle (left minus right) so that a positive value would denote left-handle leading. We show that the short-moment-arm (left) participant’s handle in pairs and the left hand of individual participants led the right handle in asymmetric dynamics, but not in symmetric dynamics, during the last five successful trials (Figure 6(a)). In contrast, during the first five successful trials, small position differences between two hands were found with similar magnitude for both symmetric and asymmetric dynamics, in both paired and individual modes. Experimental results indicate that role specialization was not necessary to enable successful task performance. Spatial coordination and measured handle forces: asymmetric and symmetric dynamics. (a) Across-mode (paired vs individual) comparisons of mean differences between left and right handle positions of the first and last five trials for each task dynamics (asymmetric and symmetric). (b) Across-mode (paired vs individual) comparisons of mean differences between left and right absolute handle forces of the first and last five trials for each task dynamics (asymmetric and symmetric). Data shown in panels are from 14 participants (seven pairs). Gray dots are data from each paired and individual trial. Asterisks denote statistically significant differences between trials and dynamics (*, **, and *** denote p < 0.05, 0.01, and 0.001, respectively). Whiskers indicate minimum and maximum values.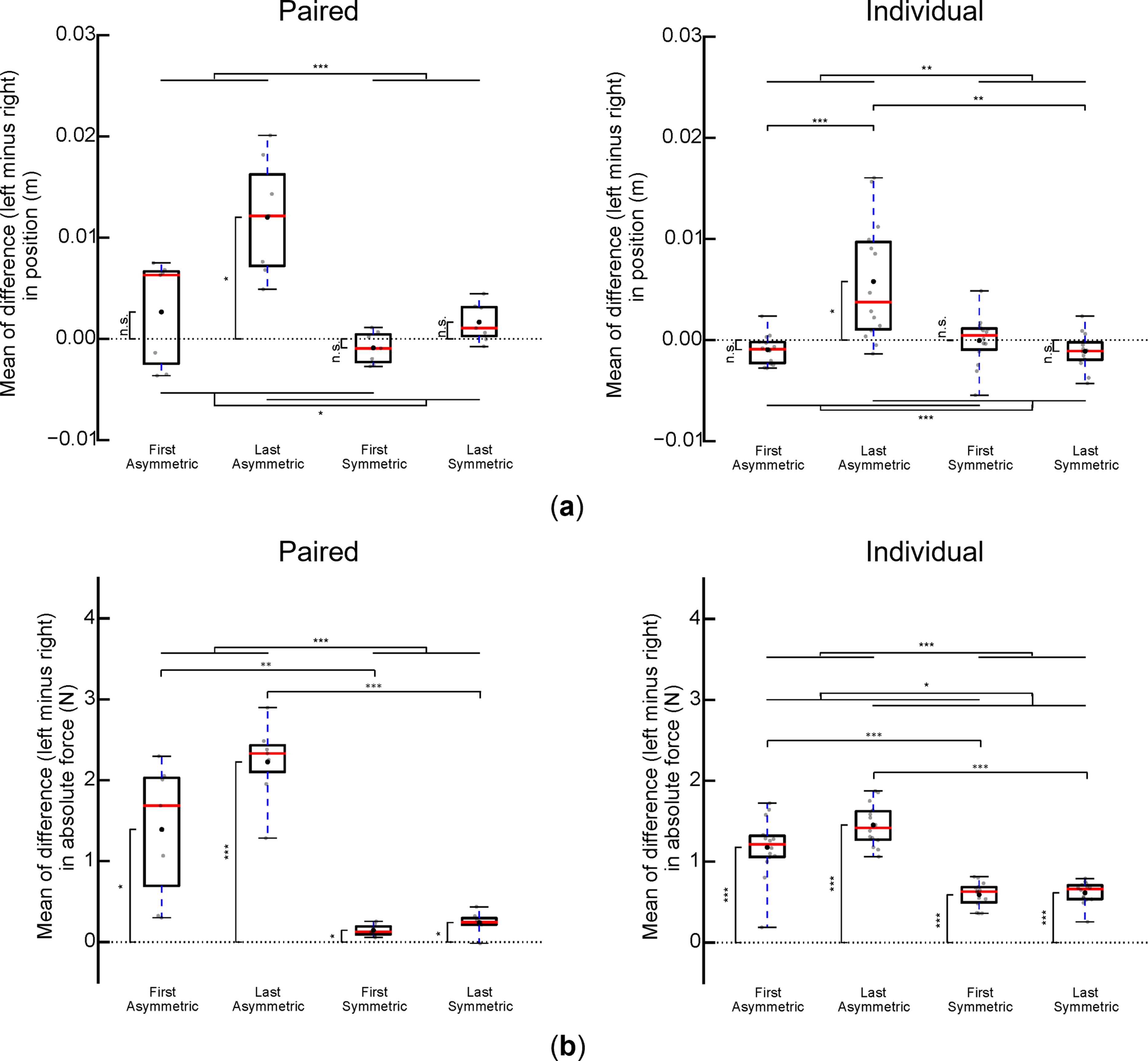
The above observations were confirmed by statistical analysis. For paired modes, we found significant main effects of Trial (F (1,6) = 11.62, p = 0.014) and Dynamics (F (1,6) = 56.36, p < 0.001). For individual modes, we found a significant Trial × Dynamics interaction (F (1,13) = 24.79, p < 0.001). Post-hoc tests revealed that spatial left-lead was greater during the last than the first five successful trials in asymmetric dynamics (t (13) = 5.14, p = 0.001), and that spatial left-lead was greater in asymmetric than symmetric dynamics during the last five successful trials (t (13) = 4.46, p = 0.003). Lastly, one-sample t-tests further confirmed this phenomenon, with only the spatial lead during the last five trials in asymmetric dynamics being significantly different from zero (t (6) = 5.43, p = 0.013 and t (13) = 3.67, p = 0.023 for paired and individual modes, respectively).
To quantify the relative force contribution from both hands, we computed the mean of differences in absolute force between two handles (left minus right). We found greater force differences in asymmetric than symmetric dynamics during the first and last five trials for both paired and individual modes (Figure 6(b)). Specifically, the short-moment-arm participant (or hand) used more force than the long-moment-arm participant (or hand) in asymmetric dynamics. Statistical analysis confirmed the significant main effect of Dynamics for both paired and individual modes (F (1,6) = 127.15, p < 0.001 and F (1,13) = 192.73, p < 0.001, respectively). This suggests a greater left-right force difference in asymmetric than symmetric dynamics. We also found a significant main effect of Trial for individual modes (F (1,13) = 5.71, p = 0.033), and one-sample t-tests showed that left-right force difference in all settings were significantly greater than zero (p < 0.001).
In summary, we found differences in handle position and force when comparing task dynamics between individuals and pairs. Specifically, when either performed asymmetric dynamics, one (left) handle consistently led and exerted more force than the other (right). In contrast, no clear position or force differences were found when they performed in symmetric dynamics.
Based on the observation that pairs outperformed individuals, particularly in the asymmetric dynamics, and that both pairs and individuals consistently led and applied more force with the left handle during the last five successful trials, we conclude that there may be a correlation between spontaneous emergence of role specialization and improved paired performance by co-adaptation through learning. The relationship between role specialization and superior performance was supported by statistical analysis (Figures 4(e) and 5(e)). The beam angle (ordinate) is large when the positional difference of participants (abscissa) is near zero at the beginning, as shown by the dark blue markers (early trials) representing experimental data (Figure 4(e)). Subsequently, the positional difference increases across trials, as warmer color markers indicate, as the beam angle decreases. This trend shows that role convergence and performance improvement develop together by co-adaptation through learning in a trial-by-trial fashion in asymmetric dynamics. The negative linear trend in asymmetric dynamics is obvious in pairs (
4. Discussion
In asymmetric dynamics, there was no significant difference in transportation time between pairs and individuals (Figure 3(b)). However, pairs achieved a significantly smaller beam angle than individuals (Figure 3(a)). In contrast, in symmetric dynamics, individuals achieved significantly shorter transportation times, smaller beam angles, and fewer trials. That is, pairs performed better in asymmetric dynamics, the only experimental condition in which role differences were observed. In fact, in symmetric dynamics, in which pairs did not exhibit clear role differences, the beam was wobbly, and pairs required more transportation time and trials than individuals. Overall, these results support our hypothesis that spontaneous role specialization enables pairs to outperform individuals, rather than better performance arising from two participants cooperating to attain a common goal per se. Therefore, two are not always better than one when performing collaborative tasks. Below, we discuss implications of our findings in the context of previous research on human–human interaction, applications to human–robot cooperation, and future research avenues.
4.1. Spontaneous role specialization emerged in physical joint actions: Pair modes
In collaborative physical actions, two agents can be assigned different roles based on asymmetric constraints imposed by visuomotor control interfaces. For example, robotic teleoperation tasks may be constrained by different types of physical operations afforded by robotic slaves (Oosterhout et al., 2018), or different fields of view (Gromov et al., 2012) each agent may have access to. In these cases, pre-determined role specialization is often designed to match task constraints, for example, one agent controls the vertical position of the load with a crane whereas the other controls the horizontal position of the load with a robot (Oosterhout et al., 2018). However, many other collaborative tasks may not explicitly impose asymmetric constraints on how each agent can perform the task, and participating agents are not directly assigned different roles. In these cases, role specialization may emerge spontaneously, and this phenomenon has been quantified in task-specific ways. Reed and Peshkin (2008) showed that two agents may specialize in performing the acceleration and deceleration phase of a fast crank rotation task. Groten et al. (2009) quantified the force contribution in a haptic collaborative tracking task and found that the contribution was asymmetric between two agents, but that the asymmetry was not consistent across trials for each pair. Moreover, Melendez-Calderon et al. (2015) showed that the torque contributions from two agents in a similar haptic collaborative wrist-tracking task can be classified into different categories based on distinct asymmetric patterns. However, these patterns were not consistent within or across pairs. Bosga et al. (2010) asked pairs to move a rocking board to track different prescribed oscillatory motions by full-body motion and demonstrated that one agent is more likely to temporally lead the pair movement.
Notably, task conditions in the above-mentioned studies were symmetric. That is, the same control input to the task from each agent would have the same effect on the task. However, it has been argued that the extent to which role specialization can emerge in these tasks can result from idiosyncratic differences in individual sensorimotor capabilities, for example, speed, accuracy, strength, which may not require pair-wise co-adaptation to make asymmetric contributions. For example, differences in reaction time and movement speed could lead pairs to exhibit spatiotemporal asymmetry in joint reaching (Takagi et al., 2016). Therefore, previous work has not found consistent role specialization in symmetric tasks and has often been quantified on a pair-to-pair basis, making it difficult to assess common patterns among pairs. In contrast, our recent work (Takai et al., 2023) demonstrated that consistent role specialization can spontaneously and gradually emerge across trials to take advantage of the implicit asymmetric dynamics.
In our previous work, computational simulation of hand movements associated with successful performance of our virtual task (Figure 4(e)) predicted that the asymmetric beam movement task has a large solution space for pair coordination (Takai et al., 2023). Therefore, it is not necessary for pairs to assume different roles to successfully perform the task. In fact, in our previous work, when asymmetric dynamics were performed without haptic feedback, no role specialization was found and peak beam tilt was larger than with haptic feedback (Takai et al., 2023). Therefore, it is likely that pairs exploited the asymmetry of the mechanical leverage in this task to improve performance through a haptically mediated co-adaptation process, that is learning acquisition of the leader–follower relationship. With regard to symmetric dynamics, participants explored, on a trial-to-trial basis, coordination patterns of viable strategies (Figure 5(e)). Nevertheless, and in contrast to asymmetric dynamics, they did not fully converge to consistent solutions.
Experimental results of asymmetric dynamics in the current study are consistent with findings of our previous work, showing that the left handle spatially led the right handle (Figure 4(a)), and the force applied on the handles was uneven (Figure 4(c)). In our previous work (Takai et al., 2023), these metrics successfully captured leader–follower relations in paired interaction and co-adaptation in asymmetric dynamics (see details in Supplemental Material, Quantitative Identification of Participant Roles). The spatiotemporal lead of the short-moment-arm participant and the larger force measured at the left handle are consistent with the definition of leaders in joint motor studies (Amazeen et al., 1995; Groten et al., 2009; Reed and Peshkin, 2008).
Furthermore, we also showed that individuals adapted to asymmetric dynamics using a similar inter-limb coordination strategy. In contrast, when performing the task in symmetric dynamics, role specialization between limbs was much weaker in both pairs and individuals. Edelson and colleagues identified responsibility aversion, defined as subject propensity to avoid assuming responsibility for others’ outcomes, as an important factor for emergence of leader–follower roles in individuals and groups performing non-motor tasks (Edelson et al., 2018). Our previous (Takai et al., 2023) and current studies suggest that asymmetry in collaborative contexts is an additional factor that could account for the emergence of leader–follower relationships in motor and non-motor tasks in individuals, pairs, and society.
4.2. Spontaneous role specialization emerged in physical joint actions: Individual modes
As noted in the Introduction, previous work on human–human haptic collaboration has provided mixed results regarding the superiority of task performance by pairs over individuals. We identified the higher number of effectors used in paired tasks as one of several factors that could have caused conflicting results. To rule out this potential confounder, we compared paired with bimanual individual performance of the same haptic task.
When an individual performs a visuomotor task bimanually, two hands may contribute differently even if the task context is symmetric. Such role specialization could be attributed to differences in motor control of dominant and non-dominant hands. Because the brain has direct access to internal models of each hand, for example, strength and variance, the contribution of each hand to the task can be determined in an optimal fashion (Salimpour and Shadmehr, 2014; O’Sullivan et al., 2009). Furthermore, the dynamic dominance framework (Sainburg, 2014) proposes that control of the dominant and non-dominant hand is predominantly mediated by predictive and reactive control, respectively. Therefore, it is possible that role specialization can directly emerge from allowing each hand to perform in its specialized mode. In one of our previous studies, we showed that the dominant (right) hand can move faster than the non-dominant (left) hand in a symmetric continuous object balance task (Mojtahedi et al., 2022). Moreover, there is evidence that bimanual coordination may be organized by the dominant hemisphere, as cross-limb coupling is often asymmetric (De Poel et al., 2007; Hu and Newell, 2011; Treffner and Turvey, 1996). However, we did not find strong evidence for unequal contributions by the two limbs when individuals performed symmetric dynamics. Instead, we found stronger role specialization in asymmetric dynamics, which was acquired gradually after repeated exposure to dynamics. It is unlikely that this role specialization resulted from intrinsic differences between the limbs because limb dominance effects do not require practice to be observed. Instead, emergence of asymmetric contributions in individual modes in our results could be explained by a motor learning process in which participants found better coordination strategies to leverage asymmetric dynamics.
Whereas both pairs and individuals learned to perform asymmetric dynamics with specialized roles for each hand, the underlying learning process may differ. When learning as an individual, somatosensory feedback from both hands can be integrated with visual feedback about object movement to infer task dynamics, which remain invariant. In contrast, when learning how to perform the task as a member of a pair, the haptic feedback acquired by an individual results from a combination of task dynamics and motor actions of the other participant (Supplemental Material, equation (5)), who may change the motor actions on a trial-to-trial basis. Nevertheless, the extent to which such mixed information drives pairs to adopt observed coordination patterns remains to be investigated.
4.3. Spontaneous role specialization may facilitate paired performance
Previous studies using tasks with explicitly assigned asymmetric roles revealed that pairs performed better than individuals. For example, when two participants were assigned to control different dimensions of object movement, pairs could complete the tasks faster (Oosterhout et al., 2018; Wahn et al., 2016). In the present study, we found that task performance, that is, the tilt angle of the beam, was smaller for pairs than for individuals for asymmetric dynamics in which consistent pair role specialization emerged spontaneously. In contrast, pair performance for symmetric dynamics was worse than for individuals. This observation suggests a potential confounding effect related to the difference in inherent task difficulty between symmetric and asymmetric dynamics. The beam angle in symmetric dynamics varies widely within a single trial, whereas the beam angle in asymmetric dynamics is smooth and barely varies (Figure 2). Additionally, the number of total trials to achieve 40 successful trials was larger in symmetric than asymmetric dynamics (Supplemental Figure 1(c)). However, the following considerations indicate that the higher difficulty of symmetric dynamics alone was not the main driver of our findings. First, both asymmetric and symmetric dynamics afford many feasible coordination patterns (Figures 4(e) and 5(e)). This means that the symmetry of the tasks does not inherently limit what the agents can do, and movements characterized by large within-trial variability were still successful. Second, although individuals needed more trials to achieve 40 successful trials in symmetric dynamics, the overall success rate was still quite high (approximately 84% and 92% for symmetric and asymmetric dynamics, respectively). In contrast, the overall success rate for pairs was much lower in symmetric than in asymmetric dynamics (approximately 51% and 95% for symmetric and asymmetric dynamics, respectively). These results suggest that task symmetry influences task difficulty much more for pairs than for individuals. Therefore, this phenomenon can only be explained by the inability of pairs to develop consistent role specialization in symmetric dynamics.
Our results are consistent with the proposition that group benefit may originate from allowing each agent to make contributions that are consistent with individual capabilities (Wahn et al., 2018), this phenomenon manifesting as role specialization. Furthermore, our results can be explained by differences in sensorimotor constraints for controlling bimanual versus joint motor actions. Bimanual coordination might be constrained by inter-limb coupling effects such that greater neural resources may be engaged during asymmetric than symmetric bimanual coordination (Rueda-Delgado et al., 2014; Swinnen and Gooijers, 2015; Swinnen and Wenderoth, 2004). Therefore, it is conceivable that bimanual performance in our symmetric dynamics might have been cognitively less demanding than in asymmetric dynamics. An opposite scenario occurred when pairs were asked to perform our task in symmetric dynamics. Here, spatiotemporal synchronization is difficult because sensorimotor delays and noise challenge each agent’s ability to match the movement characteristics of the other. In contrast, when asymmetric actions are produced through role specialization, spatiotemporal synchronization is less important and each individual can focus on a unimanual sub-task, leading to lesser cognitive demand for each agent.
The extent to which our findings might generalize to tasks involving more agents or more complex tasks, as well as their underlying mechanisms, task specificity or the number of repetitions, remains to be investigated. Furthermore, investigating the potential for optimal actions through effort exerted by each agent could open new avenues for effectively utilizing spontaneous role specialization in multi-agent collaboration and human–robot interactions (below).
4.4. Leveraging leader–follower relations to optimize task performance in human–robot interactions
For human–robot interactions to be successful, it is important for the robot to infer the partner’s actions via sensory cues. These phenomena have been extensively studied in scenarios involving the inference of movement direction of reaching movements (Mojtahedi et al., 2017b) or the direction of a robot leading a human partner walking (Regmi et al., 2022) by using stiffness cues, or humans leading movement sequences during dancing with a robotic system using haptic cues (Chen et al., 2017), teleoperations using impedance measured from human operator forces and torques (Laghi et al., 2020; Li et al., 2018) or controlling an exoskeleton using muscle activity (Ison and Artemiadis, 2015; Peternel et al., 2017). It is also important to relay sensory information from slave robots to humans that humans need for interaction by simulating the difference in force sense that occurs during human manipulation (Howe, 1992). However, most studies either focus on tasks in which pre-determined roles were given to the robot and human, or tasks in which symmetric dynamics do not inherently favor emergence of specific roles.
Our results contribute to the field of human–robot interaction by showing that modulation of physical cooperation dynamics (symmetric vs asymmetric) through robotic interfaces can significantly impact task performance by interfering with, or facilitating, spontaneous role emergence. These findings have important implications for design of human–robot interaction control algorithms for various applications, for example, exoskeletons, robotic surgery, and physical substitution. In the context of robotic-assisted rehabilitation for stroke patients using exoskeleton robots, multiple agents (rehabilitation robot and patient) move the patient’s paretic limb together when constrained by the limb’s physics and the exoskeletal robot. In one case, patient and rehabilitation robot interaction through exoskeletons is physically asymmetric due to asymmetric pivot structures in the robot. In other cases, pivot positions or arm lengths can be adjusted to create either symmetric or asymmetric dynamics. Based on our findings, such mechanical design choices can be used to induce spontaneous leader–follower emergence or follower–leader relationships between robots and patients. This approach allows elicits role emergence depending on whether the rehabilitation goal is for the robot or the human to lead the physical interaction.
Given that our findings were derived from human–human interactions, it is crucial to investigate how these results might translate to human–robot interactions. Future research should specifically explore how asymmetric dynamics, which foster leader–follower dynamics, could similarly enhance performance in human–robot settings. In the scenario of robotic-assisted rehabilitation, although many active-assistance strategies have been proposed, an ultimate strategy that optimally enhances therapeutic outcomes has not yet been identified. A previous study (Colombo et al., 2007) demonstrated the importance of adherence to the rehabilitation task by both the rehabilitation robot and the patient, showing positive effects on patient motivation and therapeutic outcomes. The study gradually decreased assistive input of the robot to achieve a tracking task based on residual abilities of stroke patients, significantly shifting the assigned roles from passive to active. More recent studies (Ivanova et al., 2022; Short et al., 2023) identified improved performance in pairs depending on the partner’s ability, the stiffness of the virtual connection, and the haptic interaction force from the partner in 1-DoF and 2-DoF tracking tasks.
For spontaneous role emergence to occur, goal sensing for reactive control is necessary, but not sufficient. Both robot and human agents also must use the inferred partner’s action to predict future interactions. Furthermore, a robot must know when to take or delegate a role to the human agent, which may require the robot to infer intrinsic asymmetries in task dynamics.
5. Conclusions
This study reports the relationship between role specialization and performance in virtual, haptically-coupled, paired and individual tasks. Pairs outperformed individuals, but only when clear roles emerged through control of different aspects of a task in asymmetric dynamics. In contrast, individuals outperformed pairs when role specialization did not occur in dynamics that favored control of the same aspects by two participants. Our findings do not support the “assembly bonus effect” reported by previous work while highlighting a necessary condition for the performance advantage of paired versus individual participants: role specialization. Therefore, we conclude that role specialization, rather than paired interaction per se, is a key factor underlying superior performance in pairs.
Our work also provides an important framework and foundation for exploring a broad range of human–robot interactions, including clinical applications. One of the factors that should be investigated to improve task performance and that is common to both paired and individual modes is the magnitude of the contribution of each subject or arm, respectively, that is, role assignment. Participant cooperation with a training program is highly likely to be essential and beneficial based on our results and those of Colombo et al. (2007). These applications would leverage dyadic tasks with human therapists or robotic devices designed to elicit role emergence to improve sensorimotor function in patients affected by neurodegenerative disorders. The goal of these clinical applications will be to modulate active control of the task by the patient from a “follower” to a “leader” role. Therefore, the perspective offered by the present work paves the way for future research in human–robot interactions, as well as neuroscience and neural engineering, to identify neural mechanisms underlying human–human and human–robot interactions.
Supplemental Material
Supplemental Material - Role specialization enables superior task performance by human dyads than individuals
Supplemental Material for Role specialization enables superior task performance by human dyads than individuals by Asuka Takai, Qiushi Fu, Yuzuru Doibata, Giuseppe Lisi, Toshiki Tsuchiya, Keivan Mojtahedi, Toshinori Yoshioka, Mitsuo Kawato, Jun Morimoto, Marco Santello in The International Journal of Robotics Research
Supplemental Material
Supplemental Material - Role specialization enables superior task performance by human dyads than individuals
Supplemental Material for Role specialization enables superior task performance by human dyads than individuals by Asuka Takai, Qiushi Fu, Yuzuru Doibata, Giuseppe Lisi, Toshiki Tsuchiya, Keivan Mojtahedi, Toshinori Yoshioka, Mitsuo Kawato, Jun Morimoto, Marco Santello in The International Journal of Robotics Research
Supplemental Material
Supplemental Material
Supplemental Material
Footnotes
Acknowledgments
We thank Dr. Heni Ben Amor for his comments on an earlier version of the manuscript.
Author contributions
Conceptualization: AT, GL, MK, JM, MS.
Methodology: AT, GL, TT, KM, TY, MK, JM, MS.
Investigation: YD, TT, TY.
Visualization: AT, QF.
Funding acquisition and project administration: JM, MS.
Supervision: MK.
Writing—original draft and writing—review and editing: AT, QF, MK, JM, MS.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Science Foundation (NSF) grant [BCS-1827752], the Japan Agency for Medical Research and Development (JP22he2202017), the Commissioned Research of National Institute of Information and Communications Technology (NICT) (18701), the Kayamori Foundation of Informational Science Advancement, the New Energy and Industrial Technology Development Organization (JPNP20006), the Innovative Science and Technology Initiative for Security, and Acquisition, Technology & Logistics Agency (ATLA) (JPJ004596).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article
Data Availability Statement
Data and code of the current study are available from the corresponding author, Asuka Takai, upon request.
Supplemental Material
Supplemental material for this article is available online.
Index to Multimedia Extensions
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.