
Abstract
Human migration requires developing new relationships with plants. In Aotearoa New Zealand (AoNZ), settled by Polynesians c. 800 years ago, myriad linguistic and conceptual clues in oral tradition invite examination of this process and the contributions of new and familiar knowledge, to understand the intertwining of social and natural landscapes. We used one enduring form of Māori oral tradition, whakataukī (ancestral sayings), to examine biocultural relationships between humans and plants in AoNZ. We asked first whether plant species mentioned are representative of plant family phylogenies in AoNZ, and globally. We then examined the occurrence frequency of introduced and native plants in whakataukī to tease out plant knowledge that connects to prior experience through the Pacific, including cultivation, and consider how people-plant relationships might be influenced by cultural histories. We found that at the level of plant order, the number of plant species recognized in AoNZ is strongly correlated with the number of species recognized globally. Whakataukī tend to mention plants from orders that have a high number of species present in AoNZ but with some notable outliers. The enrichment of whakataukī references to the Solanales and ferns is driven by the importance of kūmara and aruhe/rarauhe, both key cultivated food crops. Cultivated plants as a group are well represented - both those that traveled with the early settlers, and also native species, highlighting the importance of cultivated plants in constructing a new human ecodynamical landscape. Plants used for ceremonial, medicinal, and food purposes also occur, as do gymnosperms and other forest trees with high cultural value, such as tōtara, rātā, and kauri. We consider our findings in the context of a rapid shift from small islands at tropical latitudes to large temperate islands with a diversity of biomes, and the innovation and adaptability of those creating lives in a new land.
Keywords
Introduction
Successful human settlement in new lands is underpinned by the development of new relationships between people and plants, including hitherto unknown species. Environmental conditions in these places may differ markedly from the point of embarkation and therefore may require the development of new knowledge, even when the plant species themselves are already known. Indeed, developing and enhancing human–plant relationships in these new lands is often critical for survival. Migration to new environments thus presents opportunity for cultural innovation, triggered by biological, environmental, and cultural factors (d’Errico et al. 2017), to create a foundation for lasting, dynamic human-nature relationships.
In addition to developing knowledge of native species in a place, transportation is a recurring and enduring event in the history of human migration and settlement (e.g. Hiroa 1949; Huebert and Allen 2020; Wehi and Roa 2023). Within East Polynesia, more than 35 root and tree crops are likely to have been translocated (Whistler 2009) to support human survival and flourishing. Introduced species accompanying humans generally include domesticated plants and animals, but also game animals, ornamental species, and diseases (Crosby 1986, Whistler 2009), and are often described as a “portmanteau biota” within European contexts (Crosby 1986) or a “transported landscape” in the Pacific (Kirch 1982). Such introductions may be particularly important for people where, for example, local plant foods with high energy carbohydrate content are rare (Huebert and Allen 2020).
The actions of organisms, such as humans, that result in selective environments for themselves and other organisms are sometimes described as niche construction (O’Brien and Laland 2012; Quintus and Cochrane 2018). These actions are founded on growing knowledge of, for example, plant or pollinator characteristics, preferred habitats or niches, and soil chemical and physical properties (Huebert and Allen 2020). In addition, cultural praxis, including an ethic of responsibility that links humans to the spiritual realm, may require horticulturists to contribute to holistic landscape management, with detailed knowledge of plants, soils, sites, and crops passed down from one generation to the next through a range of mechanisms, as has been the case for Māori people in Aotearoa New Zealand (Roskruge 2011). For people such as Māori who have strongly developed oral traditions, knowledge of such connections between organisms and place, and their context, may be placed in stories, ancestral sayings and aphorisms, song, and genealogical lineages or networks, as well as other repositories of knowledge (e.g. Black 2014).
One way to accelerate human niche construction, and develop relationships with the native species of a new land, is by drawing from past knowledge of phylogenetic relatives. In oceanic islands, such as Hawai’i, taxa typically belong to the same genera as other places globally, although they may be endemic at species level (Kirch 2007). Recognizing native or endemic plants as relatives of ones that are already known can facilitate human settlement and thereby contribute to the unfolding biocultural relationships in new lands; for example, phylogenetically related plants may be similar in their chemical properties (Salis-Lagoudakis et al. 2012). The linguistic transfer of names to plants of similar morphology or use in new lands, such as has occurred among Pacific islands, also suggests that bestowing species names that reference prior knowledge from elsewhere acts as a memory aid for new, related native species (Turner et al. 2003; Berkes 2012).
The success of introductions also requires knowledge of local conditions, that improves their establishment and long-term flourishing (see, for example, Jones and Biggs 1995; Lincoln and Vitousek 2017; Quintuds and Cochrane 2018; Lincoln et al 2023). Indeed, Leppard (2019) described the arrival of domesticated ecosystems as an extremely strong perturbation in islands (as well as other sensitive habitat types). In Polynesia, Huebert and Allen (2020) highlighted both the pathway and value of cultivation practices, modified from ancestral populations over many generations of Pacific voyagers and settlers. The modification process might begin shortly after human arrival on a new island, via relocation of organisms to a new habitat, with existing forest replaced by domesticated species, or through perturbation where, for example, soil conditions for domesticated species might be tested and improved for growth (e.g. Lincoln and Ladefoged 2014). Both relocation and perturbation are embedded in the history of human landscape engineering or niche construction, albeit with varying degrees of success for different species. Some species may wane in importance, and even disappear if environmental conditions are marginal in some way and these difficulties cannot be mitigated or overcome. As one example, aute (Broussonetia papyrifera) was unable to establish in the cooler climate of Aotearoa New Zealand (hereafter AoNZ) after the arrival of Polynesian settlers from a more tropical Pacific. Transported species such as aute that were unsuccessful transplants are also of interest; aute is used to create tapa cloth in much of the Pacific, but in AoNZ other sources of fiber for clothing were developed using new technologies, cultivated, and traded (Hiroa 1924; Wehi 2006).
Documenting human relationships with plants after arrival, including successful and unsuccessful attempts at relocation, cultivation, plant storage, and seed retention, is essential to understand place-based relationships, and cultural and societal development. Yet although some of these relationships and processes may be painstakingly identified through archeological materials and methods, a comprehensive understanding of past environments can also be frustrating elusive. Evidence of early settlement may be extensively explored through middens, house construction remains, soil changes, and other material remains or topographical signs, but there are limitations: these materials may deteriorate at different rates, be recoverable only in part, or be restricted to certain spaces or time periods for example. Indigenous knowledge is a further pathway that can provide “expert witness” in understanding the workings of socio-ecological systems (Fitzhugh et al. 2019), and act as a valuable window into human-other species relationships and the development of culture in new lands. This is particularly the case for oral tradition in pre-literate peoples who are or were well accustomed to passing knowledge down verbally, as well as through other means. Such knowledge may have been recorded by ethnographers during or after colonization by other peoples (in AoNZ, for example, by Colenso (1880), and Best (1930)), be recovered through experimentation, and/or remain part of cultural knowledge and practice in contemporary communities.
Aotearoa New Zealand (AoNZ) is an archipelago in Remote Oceania first settled by the Polynesian ancestors of the Indigenous Māori people around 800 years ago, followed by European colonization and settlement from around 1800 AD. Climate in AoNZ varies from sub-tropical through to sub-Antarctic, creating challenging conditions for people arriving from tropical Polynesia. At first, Polynesian settlement in New Zealand was largely coastal, although evidence suggests much of the country was explored within the first few centuries of settlement (Walter et al. 2010; Prebble et al. 2019; Bunbury et al. 2022). Early fires created substantial ecosystem change, particularly before 1600 (Wilmshurst 1997, McWethy et al. 2014), with increasing agricultural intensification following (Quintus and Cochrane 2018) and a second abrupt change to environmental conditions after European settlement disrupted the traditional food systems and land use in place (Perry, Wilmshurst and McGlone 2014). Despite these disruptions, Māori communities today maintain and value practices that build on traditional relationships with plants (see for example Bond et al. 2019). Diverse forms of oral tradition, as well as continuing practice, store ecological information about these relationships (Williams and Chrisp 1992; Wehi 2009).
The relatively short and discrete time scale of human history in AoNZ as well as high levels of species endemism, the prominence of colonization records, latitudinal spread of settlement from tropical Polynesia to temperate AoNZ, and relative isolation from the rest of Polynesia (Holdaway et al. 2019) mean that AoNZ is well situated for examination of human ecodynamics and interactions with nature. With these factors in mind, we have previously explored ecological knowledge through the lens of whakataukī, a body of ancestral sayings or aphorisms in Māori oral tradition, that includes both specific knowledge of AoNZ birds, and broader perspectives of extinction (see Wehi et al. 2013, 2018; Whaanga et al. 2018). Here, we specifically examine biocultural relationships with plants observed in whakataukī to further investigate socio-ecological relationships with plants across this relatively recent human history.
We hypothesized that whakataukī would be a rich source of references to Māori relationships with plants. We first asked if some plant species are cited more often in whakataukī, focusing on overall reference patterns for both transported and native plants in whakataukī. We asked whether plant species that are represented in the whakataukī are representative of plant family phylogenies globally, as well as those found in AoNZ itself, and investigated the ratio of native to introduced plant species in whakataukī. We hypothesized that the importance of cultivated plant species transported from Polynesia to AoNZ would result in a high frequency of occurrence for transported (introduced) species, perhaps because of the vastly different climatic and environmental conditions across much of the country. We hypothesized that cultivated species might appear most often early after settlement, during the period when developing knowledge of plant cultivation and flourishing in these new conditions was of utmost importance. However, given the importance of cultivated plants in human ecodynamics, we also expected that they would continue to feature heavily throughout the time period, with the exception of species that did not flourish, such as aute. We expected waning or extinct / unsuccessfully transported plant species to appear in very low numbers in whakataukī, and early in the time sequence; or perhaps alternatively to occur frequently if used as an exemplar with archetypal meaning, as with the extinct moa (Order Dinornithiformes; see Wehi et al. 2018). Finally, we investigated content referring to native species and their ecological relationships.
Methods
Whakataukī Dataset
Māori have a strong tradition of oral literature with a number of formal patterns including whakataukī or ancestral sayings, passed down inter-generationally, with a wide currency of use. Many whakataukī were recorded in archives of oral traditions by 19th and twentieth century ethnographers. We used the same dataset as previously, built from 4500 whakataukī collected by Neil Grove and Hirini Moko Mead (2001) in their seminal book Ngā Pepeha a ngā tīpuna, with the addition of a further 1000 whakataukī from Supplemental material. Previous work using this dataset has focused on marine and freshwater species, and birds (e.g., Wehi et al. 2013, 2018; Whaanga et al. 2018).
Phylogeny
We categorized all plant species in the whakataukī dataset by Family and then tested the proportion of species in each Family represented in whakataukī against the proportions found in Families in the NZ flora generally. The official phylogeny (APG IV) of the Angiosperm Phylogeny Group (Angiosperm Phylogeny Group et al. 2016) was adopted as a visualization framework, supplemented with entries for ferns and gymnosperms. The number of species known to occur globally in each plant order was taken from the Angiosperm Phylogeny Website v. 14 (Stevens 2017), while the number of species found in New Zealand was taken from Wilton and Breitwieser (2000). Māori plant names in the whakataukī were identified to scientific species and classified relative to the official angiosperm phylogeny. Visualization and statistics on the phylogeny were undertaken in R v. 3.5.3 (R Development Core Team 2023) using packages ape v. 5.3 (Paradis & Schliep 2019), geiger v. 2.0.6.1 (Harmon et al. 2008), and phytools v. 0.6–60 (Revell 2012).
Regressions against species counts were calculated using Phylogenetic Generalized Least Squares to account for variable relationships in the phylogeny (i.e., Galton's problem). As the Angiosperm Phylogeny IV taxonomy has known topology but unknown branch lengths, branch distances were defined as the number of steps in the phylogeny using the R package phylogram v. 2.1.0 (Wilkinson and Davy 2018). Phylogenetic Generalized Least Squares was undertaken using R packages ape v. 5.3 (Paradis and Schliep 2019) and nlme v. 3.1–139 (Pinheiro et al. 2019), with the phylogenetic correlation structure defined by Pagel's λ (Pagel 1999).
Whakataukī Occurrence, Dating, and Content
We counted all occurrences of plant names in the whakataukī dataset and also identified whakataukī that included more than one species (both plant-animal and plant-plant). We also investigated which plant species/Families were missing from the whakataukī dataset, but present in AoNZ. We assigned all whakataukī with plant mentions to categories that reflected plant phylogeny and other characteristics, such as whether they were native or introduced, and cultivated or uncultivated, based on ethnographic and archeological records and oral tradition. Finally, we considered the type of information conveyed in the whakataukī.
We estimated the distribution of whakataukī through time using linguistic markers and codified knowledge so that we could assign an approximate time span for the origin of each whakataukī (see Wehi et al. 2018 for detailed methods). Although there is uncertainty in this process, correct assignment is likely high (see Supplement, Wehi et al. 2018) and provides a broad estimate of change in overall patterns that allows examination of development in ecological knowledge and socioecological relationships. Here, we extend this time distribution work to examine whakataukī that specifically refer to plant species.
Results
Overall, 265 whakataukī in the total dataset directly or metaphorically referred to plants or plant parts. We identified around 63 plants to genus or species. Introduced species were mentioned in 54 (20%) and native plant species in 194 (75%) of whakataukī overall, although not all whakataukī referred to clearly identifiable species or plant groups.
Plant Phylogeny and Frequency
Plant species mentioned in whakataukī appear representative of plant family phylogenies both in AoNZ and globally. At the level of plant order, the number of plant species that occur in AoNZ is tightly correlated with the number of species recognized globally (Phylogenetic Generalized Least Squares; β = 0.78, λ = 0.030, t64 = 9.7, p < 3.5 × 10−14). Similarly, whakataukī tend to mention plants from orders that have a high number of species present in AoNZ (β = 0.46, λ = 0.039, t64 = 4.1, p < 1.1 × 10−4). Nonetheless, there are notable outliers to these patterns (Figure 1). The enrichment of whakataukī references to the Solanales and ferns is driven by the importance of kūmara (sweet potato, Ipomoea batatas) and rarauhe/aruhe (bracken fern root, Pteridium esculentum). Tall forest trees, including gymnosperms, are also frequently mentioned (Figure 2). They represent a number of dominant species such as tōtara (Podocarpus totara), rātā (Metrosideros robusta), and kauri (Agathis australis), that have a range of uses (e.g. timber, kauri resin). Other plant orders are notable by their absence; grasses are seldom mentioned in whakataukī despite New Zealand being rich in grass species (Poales).

Taxonomic relationships of plant orders, with circles (left to right) scaled proportional to the number of species recognized globally, the number of species present in New Zealand, and the number of times species are mentioned in the whakataukī.
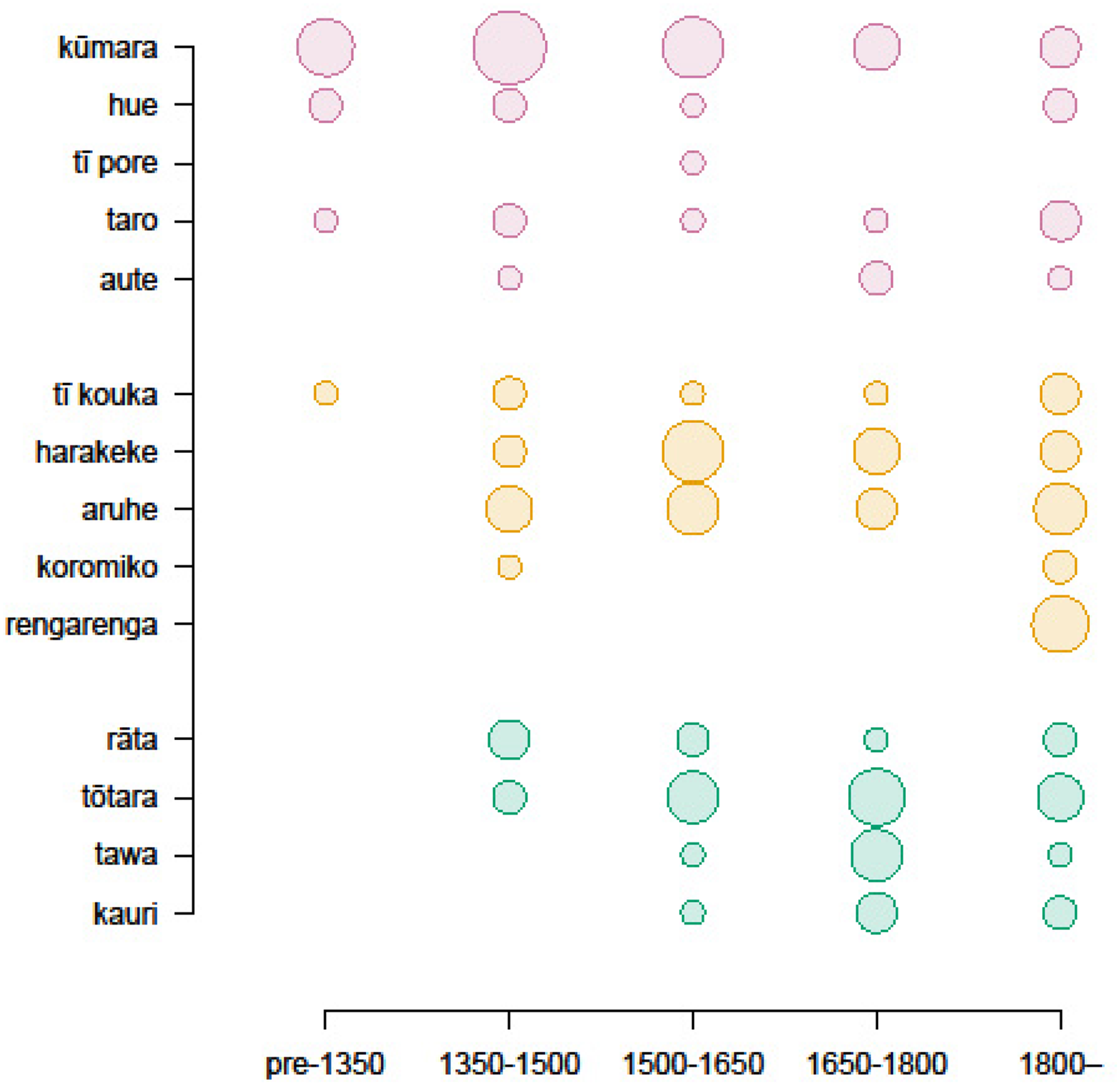
Time distributions of whakataukī that refer to introduced “waka” species (kūmara (Ipomoea batatas), hue (Lagenaria siceraria), tī pore (Cordyline fruticosa), taro (Colocasia esculenta), and aute (Broussonetia papyrifera)); native cultivated and domesticated species (tī kouka (Cordyline australis), harakeke (Phormium tenax), aruhe (Pteridium esculentum), koromiko (Veronica stricta), and rengarenga (Arthropodium cirratum)); and a sample of known native forest trees (rātā (Metrosideros robusta), tōtara (Podocarpus totara), tawa (Beilschmidia tawa), and kauri (Agathis australis)). Size of the dot is relative to the number of sayings in that time period.
Transported and Cultivated Species Relationships
Occurrence for introduced (transported) species in whakataukī had a significantly different time distribution from the overall pattern of all whakataukī, with a higher frequency of early mentions in whakataukī that we dated. This pattern was driven entirely by kūmara (
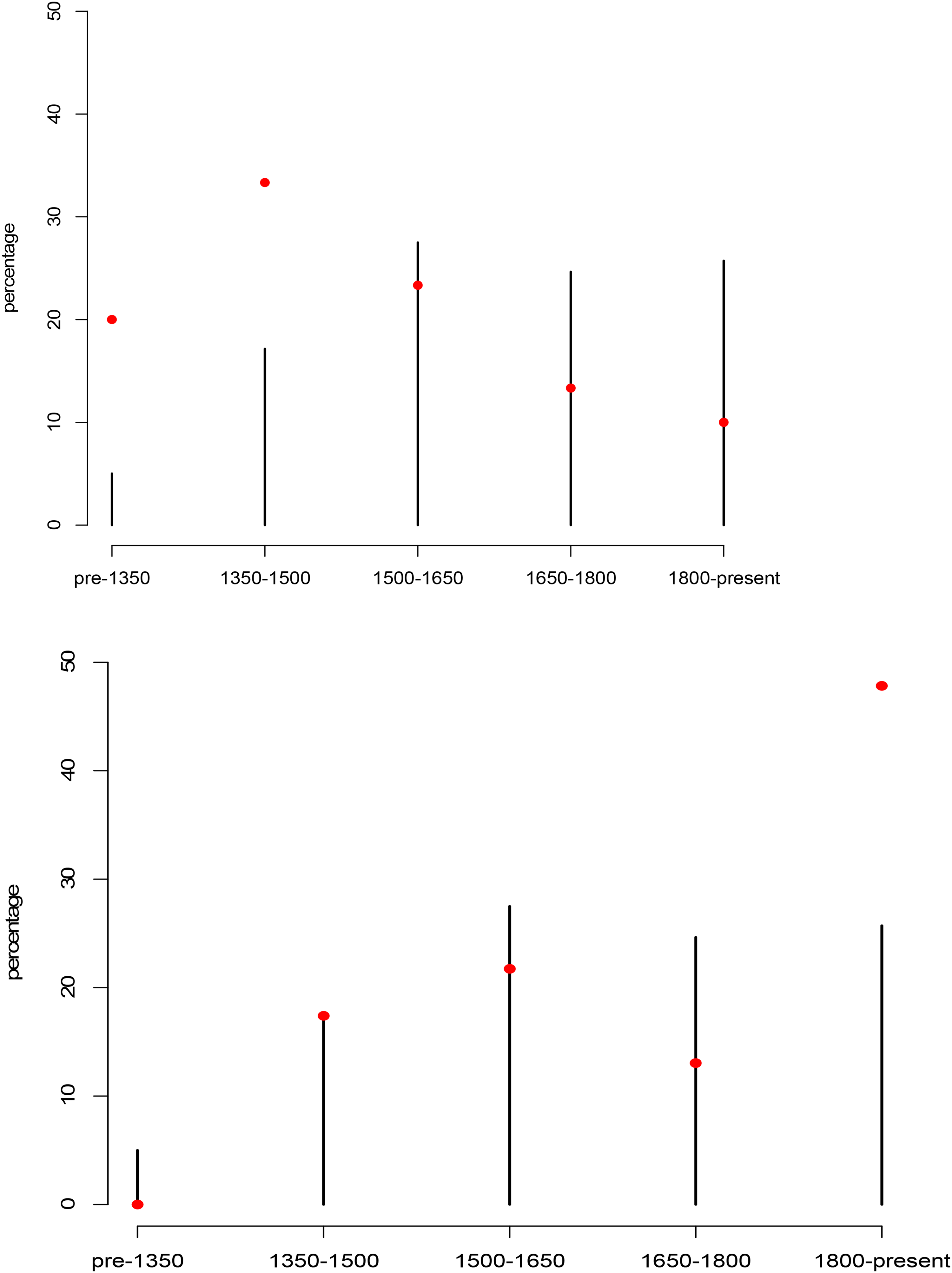
(a) Kūmara (Ipomoea batatas) whakataukī (round points, n = 31) appear earlier than the distribution of whakataukī as a whole (bars). This pattern is not seen for other introduced “waka” plant species—aute (Broussonetia papyrifera), hue (Lagenaria siceraria), tī pore (Cordyline fruticosa), and taro (Colocasia esculenta) (n = 30; i.e., this difference does not reflect a difference in power). Most kūmara whakataukī (round points) were dated as being in circulation pre-settlement (i.e., accompanying Maori settlement) or immediately post-settlement. (b) Distributions for rengarenga (Arthropodium cirratum) and aruhe (Pteridium esculentum) whakataukī are later compared to the distribution for all whakataukī. Round points represent the number of whakataukī that mention rengarenga and aruhe and that have been classified as belonging to that time period.
The importance of cultivation for societal development is underscored by the 116 whakataukī (44%) that refer to cultivations or cultivated species, both introduced and native. Content for cultivated species focuses on a wide range of practical topics such as planting conditions, pests and how to manage them, and the value of these crops (Table 1). As a group, we did not see a difference in the time distribution of cultivated introduced plant species brought by the Polynesian ancestors of Māori (“canoe” or “waka” plants kūmara, aute, hue (Lagenaria siceraria, tī pore (Cordyline fruticosa) and taro (Colocasia esculenta)) compared to native plant species that were cultivated or domesticated in AoNZ after Māori arrival (e.g. harakeke (Phormium tenax), rengarenga (Arthropodium cirratum) or tī (Cordyline spp.). However, two of the most important cultivated native plants, rengarenga and aruhe, occur in a significantly different time distribution from that for all whakataukī (
Whakataukī that Mention Cultivated Species have Similarities in their Content among Species that are Both Introduced (Kūmara) and Native (Aruhe, Harakeke, Rengarenga) to AoNZ.
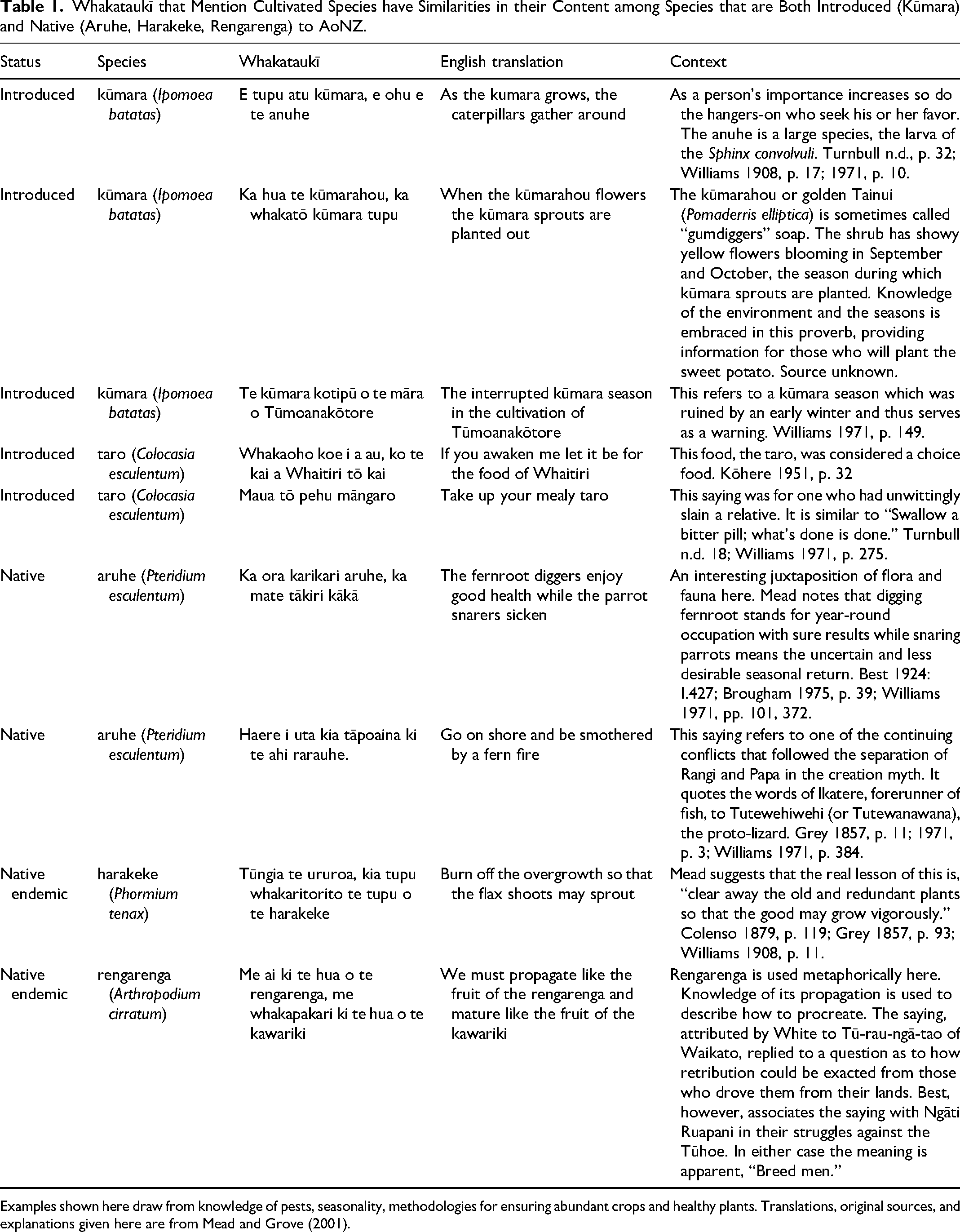
Examples shown here draw from knowledge of pests, seasonality, methodologies for ensuring abundant crops and healthy plants. Translations, original sources, and explanations given here are from Mead and Grove (2001).
Extinct Species
The only extinct plant species represented in whakataukī is the aute, brought to AoNZ from the tropical Pacific where it continues to thrive. The time distribution of aute (round points) is not significantly different from the time distribution of all whakataukī (bars in the graph) but, with only four whakataukī that explicitly refer to aute, it is impossible to draw conclusions with such a small sample size (see Supplemental, Figure S1). Visual inspection suggests that aute was important early on but was not widely discussed post-European arrival; these whakataukī refer to its disappearance, such as in the words “Te aute tē whawhea” [transl. “the aute, used for tapa cloth, will not go around”; see Mead and Grove 2001; also Hiroa 1924).
Endemic and Native Species Content
Whakataukī referring to native species contain a range of details relevant to ecological relationships. Some read as a guide to the forest, such as “Taute tītoki, whero te rātā i te waru” [Tītoki (Alectryon excelsus) matures and rātā (Metrosideros robusta) flowers are red in the eighth month; [Mead & Grove, 2001]; whereas others speak much more directly to use such as “Ka haere te tōtara haemata, ka takoto te pukatea (Laurelia novae-zelandiae) wai nui” [The strong-growing tōtara floats while the water-logged pukatea sinks]; and yet others embed references to cultural histories, such as the sighting of red pōhutukawa (Metrosideros excelsa) flowers when voyagers first arrived in AoNZ [“E, kua nui ake te kura o tēnei kāinga i te kura o Hawaiki, ka panga hoki ahau i aku kura ki te wai”; The red plume is more plentiful in this country than in Hawaiki].
Large forest trees that dominate AoNZ lowland forest include tōtara (Podocarpus totara), rātā (Metrosideros robusta, kauri (Agathis australis), and tawa (Beilschmiedia tawa). Trees such as these all feature in whakataukī, although the characteristics mentioned do not necessarily refer to their dominance in the canopy. These whakataukī mention characteristics such as resin used as chewing gum, flowering seasonality, and the burning properties of the wood. Tōtara bucks this trend and is described in majestic terms, often metaphorically referring to chiefs. However, whakataukī that mention these trees do not differ significantly from the time distribution of all whakataukī and are therefore not obviously associated with early settlement as we had hypothesized. A great deal of information that refers to both everyday life and the relationships between humans and these trees is embedded in these whakataukī.
Interaction networks between different plant species in the whakataukī show that most whakataukī refer to only one species at a time, but that some species pairs are present (see Supplemental for all networks). Rengarenga and kawariki (described by Mead and Grove (2001) as Ranunculus sp., but also identified as Coprosma grandifolia; Beever 1991) is one such common pairing. Where plant species occur together, most often both are native species. If both an introduced species and a native one occur together, the connections reveal the process of learning new seasonal calendars; for example, kūmara is linked with the tree kūmarahou (Pomaderis kūmaraho) as an indicator of the timing of seasonal planting when the tree flowers. One of the most interesting flora-fauna groupings is for kūmara and its pests, both native and introduced; caterpillars, the kiore or Pacific rat (Rattus exulans), and the swamphen or pūkeko (Porphyrio porphyrio) are all highlighted in relation to kūmara.
Discussion
Examination of the flora referenced in whakataukī reveals patterns that reflect the importance of cultivated plants. Analysis of plant phylogenies shows that the AoNZ flora is typical of others around the world at the level of Order, and the frequency at which plants occur in whakataukī is largely representative, with outstanding exceptions for Solanales that demonstrates the bias towards cultivation in the dataset. The large number of cultivated species, both introduced and native, that are referred to, highlight that whakataukī convey important information and concerns associated with flourishing in a new land. They emphasize the active human agency observed in niche construction (O’Brien and Laland 2012) and many of the connections also seen in whakapapa frameworks (Roberts et al. 2004). Early European ethnographers confirm the development of horticultural practices undertaken for plants to flourish; for example, Colenso (1844) reported that during a journey down the East Coast, taro plantations were large, in nice condition, and looked very neat, the plants being planted in true quincunx order, and the ground strewed with fine white sand. As well, small screens formed of branches of manuka, to shelter the young plants from the violence of the winds, intersected the grounds in every direction. Yen (1961) also highlighted technical adaptations in propagation and storage that allowed such transported species such as kūmara to thrive in this new environment.
Seen from another perspective, the data on both transported and native species in whakataukī align with Smith and Hutchings’ (2024) observation that “diverse whānau groupings across the islands thrived on the māhika kai (wild foods) from the ngāhere (forests), awa (rivers), and moana (ocean), as well as cultivated foods all of which are the atua domains of Haumia-tiketike, Tāne-Mahuta, Tangaroa, and Rōngomātane respectively, all offsprings of primordial parents, Ranginui and Papatūānuku.” They go on to say that “Harvesting or cultivating kai involved karakia to ngā atua, recalling the actions and identities of tūpuna, and ensuring that all practices followed the appropriate tikanga in order to maintain order and balance between tangata, whenua and atua.” The names of atua are seen in whakataukī, as are names of people and places associated with plants; Roskruge (2011) affirms the practice of applying names to sites, as well as treasured items, including special trees and plants.
Most of AoNZ's approximately 2000 native species are not explicitly seen in the whakataukī. Nonetheless, native plants used for ceremonial, medicinal, and food purposes all occur in the whakataukī dataset, and some, such as the cultivated rengarenga and harakeke, occur often, although references tend to be metaphorical rather than descriptive of use. Knowledge of plants and their uses is transmitted in contemporary and past cultures through a range of different mechanisms; for example, by practising medicinal experts as part of their healing toolbox. Yet even plants mentioned rarely in the whakataukī provide information on Māori understandings of environmental conditions. One example is wī or wā (Carex sp. or Juncus sp.) which is referenced as an indicator of scarcity, but also indicates a limitation in seasonal wetness where it grows on open ground (Roskruge 2007). As yet, other forms of oral tradition such as waiata (song), whakapapa (layers of relationships, often referred to as genealogy) and pūrakau (stories), and the text archives of nineteenth-century ethnographers, have not been comprehensively examined to explore the range of human–plant relationships. This might be a useful path to take in future.
It remains unclear how much native plant use in AoNZ was based on prior experience with relatives in the Pacific, but the transfer and use of names such as kawakawa (Piper excelsum) and kiekie (Freycinetia banksii) for both morphologically and /or phylogenetically similar plant species in the Pacific draw attention to the shared origins of Polynesian culture. Other work suggests common discovery processes in many cultures that are isolated from each other globally (Salis-Lagoudakis et al. 2012), but this aspect of plant use has not yet been explored in depth in AoNZ.
The notable preponderance of introduced “waka” species mentioned in the whakataukī dataset indicate the importance of this transported portmanteau to successful settlement. Key transported food crops such as kūmara are very well represented, and similarly also appear in important stories cherished in oral tradition (e.g., Jones and Biggs 1995). The high number of early mentions in the dataset for kūmara appears to indicate vigorous discussion about this root crop and its cultivation from the early settlement period, at least in northern AoNZ, perhaps reflecting urgency of kūmara cultivation during this period. Its importance in niche construction is certainly reflected in the propagation and storage methods successfully developed in AoNZ, that are not generally used elsewhere in the Pacific (Yen 1961). The narrative of Whakaotirangi, from Tainui, details her successful attempts to improve soils and growing conditions in the populous coastal area where the Tainui waka, or ocean-traversing vessel, landed (Jones and Biggs 1995; Wehi and Roa 2023; also see Barber 2010). The connection of Whakaotirangi's story to the arrival of Tainui squarely places niche construction for kūmara growth at the start of Tainui settlement. The concepts of relocation, perturbation, and feedback as identified in trajectories of agricultural change (see Huebert and Allen 2020) are all present in both these narratives and in whakataukī. We acknowledge kūmara narratives from other regions such as Northland that embed similar experiences and knowledge.
In the whakataukī dataset, cultivation implicitly references the long history of experience in the Pacific but also highlights new developments relevant to a new temperate environment, and the effects of pests (see Leppard 2019). Unpacking the details of whakataukī for cultivated species often reveal knowledge of growing conditions or ecology, that in turn lay a foundation for intensification or expansion of agricultural strategies associated with manipulating planting environments and land cover change (Huebert and Allen 2020). In addition, the ways that plants may be prepared or cooked as food are also referenced. Landscape-changing agents such as fire (see Table 1 for examples) are mentioned in association with native species aruhe and harakeke, both of which were domesticated.
The whakataukī record important details of developing human and more-than-human relationships in a new land. We note however that regional differences in the timing and spread of cultivation are evident in the archeological record, and that some of the findings here contrast with archeological observations, for example, on taro. Taro is well-known from Whakaotirangi's gardens at Aotea after the arrival of the Tainui waka, one of the early seafaring vessels (McIvor and Forde 2021). It is also recorded as transported on other waka such as the Aotea and Nukutere (Best 1904), and taro plantations are described in the ethnographic literature (Dieffenbach 1841; Colenso 1844) as well as being the subject of intense archeological interest. Yet taro is not frequently referenced in whakataukī.
Archeological evidence from the island of Ahuahu indicates that taro cultivation was especially important in early northern New Zealand (Ferdon 1988; Prebble et al. 2019; but also see Barber 2020). Taro in AoNZ had a restricted distribution, with climatic conditions unconducive to its flourishing in the south of the country. Moreover, plantations were not as productive as elsewhere in Polynesia (Furey 2006: 13). Archeological work by Prebble and his co-authors demonstrate a likely shift in taro production after 1500 CE on Ahuahu, given the absence of taro pollen, and argue that this is linked to the abandonment of taro cultivation, and a shift to dryland kūmara production. That is, they suggest that kūmara production may have been less important in the early settlement period. Barber (2020), on the other hand, draws attention to taro production in the far north in the early period. The whakataukī that we see linked to taro emphasize that it was a valued food, but whakataukī referencing taro are not especially visible in the early periods, in contrast to kūmara. Nonetheless, the whakataukī dataset is generally unlinked to specific areas and is not comprehensive across all regions of AoNZ, so we expect work in other disciplines and on other datasets will help unravel these tangled lines of evidence. Uwhi or yam (Discorea sp.) is a further Polynesian introduction that was transported to AoNZ (Best 1931) but that is not referenced in the whakataukī set analysed here.
In the whakataukī, there is a steady but small increase in sayings that mention aruhe through to around 1650. The later prevalence of both aruhe and rengarenga in the sayings align with the continuing development of agricultural land, where areas had been cleared and were regenerating, as well as with fallow gardens. On Ahuahu, wetland conditions and fires were recorded until 1600, leading to an increase through time in fire-adapted ground fern, and bracken (Prebble et al. 2019); high proportions of ferns are recorded in the paleoecological record, suggesting continued island-wide burning after 1600–1650. These data align reasonably well with the whakataukī dataset, particularly those that refer to burning for the purposes of cultivation. There is also a large peak in sayings that refer to aruhe and rengarenga after European settlement. It is not entirely clear, however, whether this late peak represents a commentary on increasing bracken and ground fern, perhaps associated with garden abandonment after European colonization (Prebble et al. 2019), or something else. Rengarenga is used as a metaphorical indicator of prosperity in the whakataukī, for example in the whakataukī “Me ai ki te hua o te rengarenga, me whakapakari ki te hua o te kawariki” meaning that humans should propagate like the fruit of the rengarenga and mature like the fruit of the kawariki, a saying that could have become especially pertinent with European colonization.
In another line of evidence that adds context to the importance of rengarenga in whakataukī, Shepherd et al. (2016) found that rengarenga has undergone several genetic bottlenecks, indicating its domestication and later translocation, and thus its value as a food plant. While rengarenga is also thought to have a natural distribution north of around 38°S (the southern distributional limit for a number of plant species restricted to the northern North), it is found in the south associated with archeological sites, including gardens.
Plant-plant juxtapositions are uncommon within sayings, but nevertheless occur between food staples, particularly transported staples such as kūmara and taro; between fragrant plants from ferns to a subalpine speargrass; and in forest or other groupings (for example, tōtara, kauri, and harakeke) that emphasizes metaphorical allusions to chieftainship or family, and functional relationships. Many specific uses of plants are recorded in the comprehensive online database of Maori plant uses, Nga Rauropi Whakaoranga https://rauropiwhakaoranga.landcareresearch.co.nz/, drawn from the writings of early ethnographers. Although we see some seasonal linkages in the whakataukī, such as between the bushy flowering kūmarahou and kūmara planting, and the flowering of rātā and titoki, seasonal indicators are not common in this dataset. We suggest that seasonal knowledge may be more logically attached to maramataka, seasonal calendars, used by practitioners, or taught in other types of transmitted knowledge such as astronomy. Within whakataukī, nonetheless, there appear to be many different ways of conveying knowledge and making it memorable, from allusions to the embedding of stories to references to deities, connections with other species, and of course, to human endeavor.
As with our earlier analysis of birds in whakataukī, extinct (to AoNZ) species were not evident in the dataset pertaining to flora, with aute the only representative. There appear to be few extinct plant species (whether waka species or native species) in AoNZ's recent history relative to birds (Sax et al. 2002), so a low representation of extinct plant species in the dataset is not unexpected. We note however that the aute, represented in only four whakataukī, is also present in other forms of oral tradition such as place names that are associated with regional histories (Hiroa 1924), emphasizing its importance in the story of migration and settlement. The aute remains a vibrant part of culture elsewhere in the Pacific, allowing learnings from these places with different climatic growing conditions.
Taken together, the plant references in whakataukī support a narrative of large-scale environmental change where Maori have modified environments, as others have done throughout Polynesia (Huebert and Allen 2020). Rapid development of knowledge has accompanied the environmental modifications that have assisted management of agroeconomic systems. This growing environmental knowledge demonstrates dynamic innovation, as relationships with previously unknown plants became established, but also reflects the strong Polynesian horticultural basis for food systems (Kirch 2007). They also demonstrate, in Kirch's terminology, direct impacts that rely on conscious decision making (such as plant collecting, or forest clearance through fire) as well as indirect impacts stemming, for example, from transported species like kūmara, along with its pests. Nonetheless, the whakataukī dataset has limitations. For example, the sayings themselves largely represent the energies of the nineteenth and early twentieth century ethnographers who recorded them, meaning that regions are very unevenly represented, and some regions are rarely or never mentioned. Environmental restrictions that limit the distribution of the plants themselves, add another layer of complexity in interpreting the dataset and the spread of data, for example, in terms of plant phylogenies. The gender and interests of ethnographers also played a part in determining which whakataukī might be shared or recorded in the sources we studied. Nonetheless, study of the past increasingly draws from a range of disciplines, and whakataukī have a valuable role to play in illuminating both environmental practices and human concerns and attitudes, in this case toward plants. Importantly, whakataukī continue to be used across AoNZ in Māori communities, so the knowledge that they reveal continues to contribute to Indigenous revitalization in our fast-changing world.
Supplemental Material
sj-xlsx-1-ebi-10.1177_02780771241303902 - Supplemental material for Plant Biocultural Landscapes in Māori Oral Tradition
Supplemental material, sj-xlsx-1-ebi-10.1177_02780771241303902 for Plant Biocultural Landscapes in Māori Oral Tradition by Priscilla M Wehi, Murray P Cox, Hēmi Whaanga and Tom Roa in Journal of Ethnobiology
Footnotes
Acknowledgments
We thank Kahurangi Tipene, Bart Cox, and Niwa Wehi for assistance with dataset cleaning and tidying. The work of this group was supported by an award from the Marsden Fund. PMW was also funded by a Rutherford Discovery Fellowship 14-LCR-001. We also thank Te Pūnaha Matatini Centre of Research Excellence in Complex Systems for funding a writing retreat in 2021.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Royal Society Te Apārangi, Te Pūnaha Matatini, Marsden Fund (grant number RDF 14-LCR-001).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.