
Abstract
Background:
Substance use disorder emerges in a small proportion of drug users and has the characteristics of a chronic relapsing pathology.
Aims:
Our study aimed to demonstrate and characterize the variability in the expression of the rewarding effects of cocaine in the conditioned place preference (CPP) paradigm.
Methods:
A cocaine-CPP paradigm in male Sprague–Dawley rats with an extinction period of 12 days and reinstatement was conducted. A statistical model was developed to distinguish rats expressing or not a cocaine-induced place preference.
Results:
Two groups of rats were identified: rats that did express rewarding effects (CPP expression (CPPE), score >102 s) and rats that did not (no CPP expression (nCPPE), score between −85 and 59 s). These two groups did not show significant differences in a battery of behavioral tests. To identify differentially expressed genes in the CPPE and nCPPE groups, a whole-transcriptome ribonucleic acid-sequencing analysis was performed in the nucleus accumbens (NAc) 24 h after the CPP test. Four immediate early genes (Fos, Egr2, Nr4a1, and Zbtb37) were differentially expressed in the NAc of CPPE rats after expression of CPP. Variability in cocaine-induced place preference persisted in the CPPE and nCPPE groups after the extinction and reinstatement phases. Transcriptomic differences observed after reinstatement were distinct from those observed immediately after expression of CPP.
Conclusion:
These new findings provide insights into the identification of mechanisms underlying interindividual variability in the response to cocaine’s rewarding effects.
Keywords
Introduction
Substance use disorder (SUD) emerges in a small proportion of drug users and is characterized as a chronic relapsing pathology. Cocaine is one of the most widely used drugs of abuse, and cocaine use disorder represents a major health concern with social and economic consequences (Toledo et al., 2017). Among cocaine users, 15%–19% become addicted within the first 2 years of cocaine use (Toledo et al., 2017). This suggests that interindividual variability in the response to cocaine exists and determines the likelihood of developing abuse and addiction.
Characterizing the underlying mechanisms of this interindividual variability is important to understand the neuroadaptations induced by cocaine that could lead to addiction. For this purpose, the use of animal models provides enormous potential through a coordinated analysis of brain function and behavior. Models of addiction have evolved over the last two decades to provide increasing validity through anthropomorphizing rodent behavior (Crombag et al., 2008), but the markers responsible for individual susceptibility to SUD are still unclear.
Stimuli commonly associated with cocaine use elicit craving and relapse to drug taking (Huston et al., 2013), a process likely supported by the development of learned, classically conditioned associations between the effects of cocaine (unconditioned responses) and the environment in which cocaine is experienced (conditioned stimulus; Piazza et al., 2000). These associations are modeled using the conditioned place preference (CPP) paradigm, an associative memory model linking drug reward with neutral environment cues. Conditioned responses play an important role in eliciting relapse, even long after an individual has been drug-free (Crombag et al., 2008). The addition of an extinction phase and a reinstatement test allowed the CPP paradigm to become one of the most widely used paradigms to evaluate the development, expression, and persistence of drug-induced behavioral and neurological adaptations (Huston et al., 2013). Moreover, it is well documented that repeated cocaine administration induces different behavioral, cellular, and molecular modifications based on the cocaine administration pattern (Piazza et al., 2000; Puig et al., 2012, 2014; Unterwald et al., 2001). Although noncontingent drug administration is involved, the CPP paradigm is appropriate for studies of interindividual variability in rats receiving the same dose of cocaine and having the same experience (Tzschentke, 2007).
The nucleus accumbens (NAc) is critically involved in the attribution of incentive motivation and the acquisition and expression of reward-dependent behavior; Hyman et al., 2006). However, the CPP amplitude is quite variable between rats, suggesting that this amplitude can be an interesting criterion to explore the origins of the interindividual variability of the response to cocaine (Niedzielska-Andres et al., 2019; Zaitsu et al., 2014).
In this study, we first developed a statistical model to classify rats as more or less vulnerable to cocaine based on their score in the CPP test. We also performed a battery of behavioral control tests to control for the effects of extraneous behaviors on CPP performance. We then performed a global transcriptomic analysis of the NAc following cocaine-CPP expression (CPPE) and cocaine-primed reinstatement and examined the obtained transcriptomic profiles with respect to the sensitivity to cocaine reward. This model allows the identification of subgroups of rats displaying variability in the response to cocaine-induced rewarding effects. The transcriptomic results suggested a number of neurobehavioral and molecular dimensions of relevance to the addiction acquisition process. Similar experiments were performed with CPP induced by a natural reward (food).
Methods
See the Supplemental Methods File for complementary methods.
Animals
Male Sprague–Dawley rats (Charles River, Écully, France), weighing 250–300 g at the start of the experiment (n = 401), were used. Animals (three per cage) were housed for 7 days before the experiment in an environment maintained at 20°C ± 1°C with controlled humidity and a light-dark cycle. These studies were limited to males, as cycling levels of estrogen and progesterone have been demonstrated to affect both dopamine neurotransmission (Becker, 1999; Becker and Rudick, 1999; Calipari et al., 2017) and cocaine-seeking (Anker et al., 2007; Larson and Carroll, 2007; Roberts et al., 1989). Food and water were provided ad libitum. All experiments were carried out in accordance with the European Communities Council Directive, French Law, and standard ethical guidelines under the control of the institutional ethical committee of Université de Paris (no. 18-154).
Drugs
Cocaine hydrochloride (Francopia, Antony, France) was dissolved in sterile saline solution (0.9% (w/v) NaCl) to achieve a concentration of either 20 or 5 mg/ml, based on the experiment. Rats received the cocaine solution (1 ml/kg of body weight) by the intraperitoneal (i.p.) route at 20 or 5 mg/kg, depending on the experiment.
Behavioral measurements
Cocaine place preference paradigm
The apparatus consisted of two conditioning compartments (45 cm × 45 cm × 30 cm) separated by a neutral compartment (18 cm × 36 cm × 30 cm). Two distinct sets of sensory cues differentiated the compartments: the wall color (black or stripes) and the floor texture (grid or smooth). The combination was black wall/grid floor and striped wall/smooth floor. The neutral compartment had gray walls and floor. Rat movements and location were recorded by computerized monitoring Viewpoint® software (France).
The protocol consisted of three phases (Figure 1(a)). (i) Preconditioning phase (pretest): drug-naive rats had free access to the three compartments for 18 min, and the time spent in each compartment was recorded during the day. The spontaneous time and proportion distribution between compartments are presented Supplemental Figure 1(a) and (b), the mean time ± standard error of the mean (SEM) spent in the black compartment is 372.1 ± 4.7 s and in the striped compartment is 403.1 ± 4.6 s. The proportion of preference for each compartment is as follows: 39.5% of rats preferred black compartment and 60.5% of rats preferred striped compartment. (ii) Conditioning phase: 4 days during which each conditioning chamber was closed. On the morning of the first conditioning day, the animals were treated with saline and individually placed in one of the conditioning environments for 18 min. After 6 h, the animals were given cocaine (20 mg/kg, i.p.) in the opposite compartment, and this sequence alternated over the next 3 days. The control animals received saline twice a day and were submitted to the same alternating sequence between the two compartments. Animals were be assigned, randomly in each treatment group. For each pair of animals, the animal showing the most bias toward one of the compartments was cocaine- or saline-paired with the non-preferred compartments. The second animal was then cocaine- or saline-paired with opposite chamber regardless of its preference to avoid a biased experimental design. Thus, in each treatment group, there were animals conditioned to their initially preferred and some to their initially non-preferred compartment and there were an equal number of animals paired with each compartment (black vs striped). Supplemental Figure 1(c) showed the balanced spontaneous preference distribution between compartments. Fisher test showed no relationship between groups and compartments (initially preferred or non-preferred) p = 0.48. (iii) Testing phase (test): conducted the day after the last conditioning session and identical to the preconditioning phase. The results are expressed in scores calculated as the difference between the postconditioning and preconditioning time spent in the compartment associated with the conditioning drug. The time spent in the middle compartment and the scores for each groups is showed Supplemental Figure 1, there is significant difference of the time spent in the neutral compartment at the pretest between groups “(p = 0.69, one-way analysis of variance (ANOVA)”).
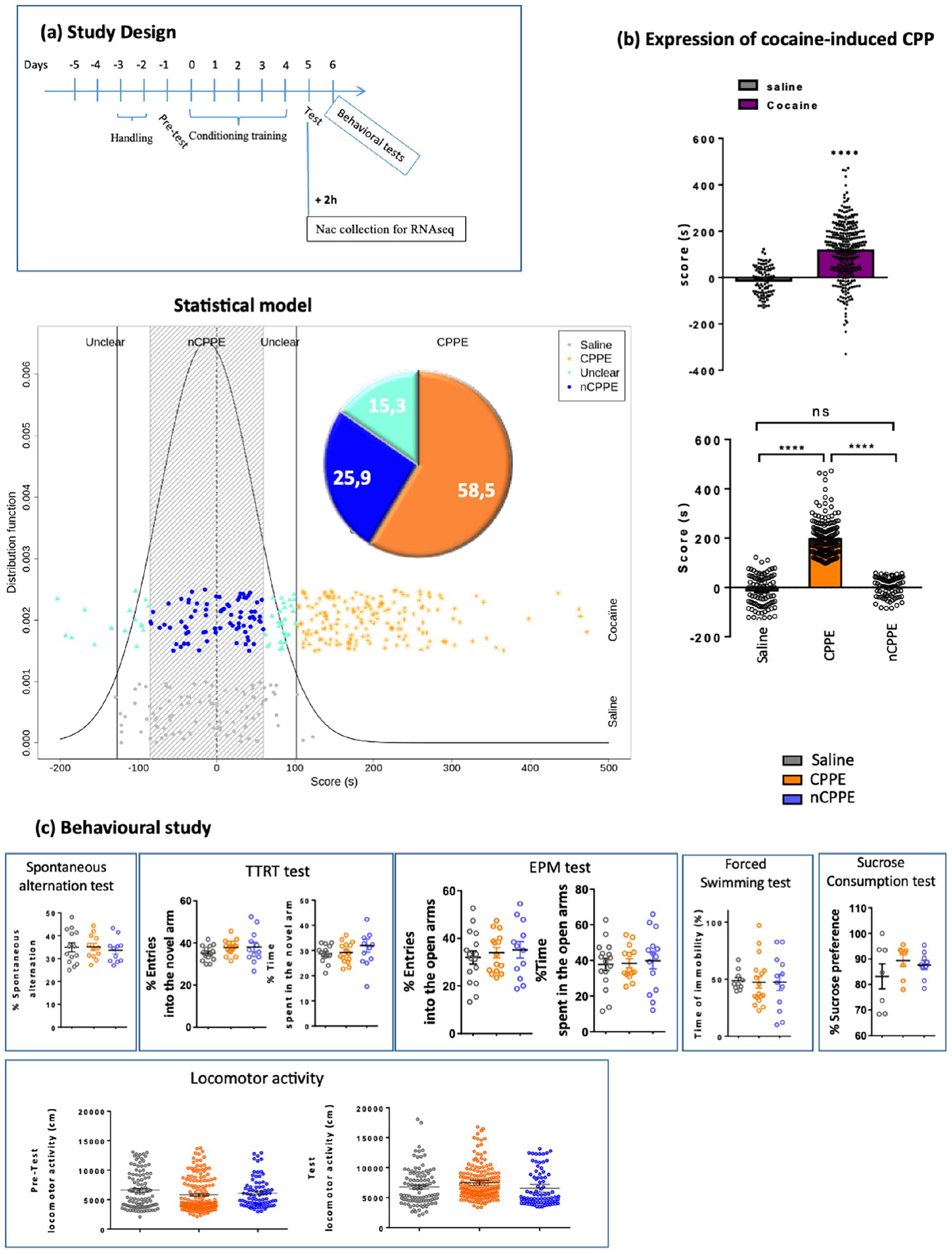
Expression of cocaine-induced CPP. (a) Study design. (b) Expression of cocaine-induced CPP in the total population and statistics. Values are expressed as the mean ± SEM (saline, n = 100; cocaine, n = 301, CPPE = 177, nCPPE = 78). (c) Cognitive and emotional behavioral assessments 24 h after CPPE in saline-treated, CPPE, and nCPPE rats. Values are expressed as the mean ± SEM (n = 7–16 for each group, spontaneous alternation: saline = 14, CPPE = 11, nCPPE = 11; TTRT test: saline = 14, CPPE = 15, nCPEE = 11; EPM test: saline = 16, CPPE = 15, nCPPE = 12; forced swimming test: saline = 10, CPPE = 16, nCPPE = 12; sucrose consumption test: saline = 7, CPPE = 8, nCPPE = 8). Comparisons were performed using t-tests or one-way ANOVA followed by post hoc tests with Bonferroni correction.
Food place preference paradigm
A similar protocol was used for the natural reward-induced CPP assessments as a control. The same CPP procedure and apparatus as cocaine CPP were used for food CPP (Dias et al., 2012). In the preconditioning phase, food-restricted rats had free access to the three compartments for 15 min, and the time spent in each compartment was recorded. The conditioning phase consisted of 8 days in which each conditioning chamber was closed. On the first conditioning day, the animals were placed in one compartment with 40 Froot Loops in a transparent food tray, and on alternate days, they were confined in the other compartment with an empty food tray. Each conditioning session lasted 25 min. The control animals were confined with an empty tray each day and were submitted to the same alternating sequence between the two compartments. The testing phase was conducted the day after the last conditioning session and was identical to the preconditioning phase.
In the extinction phase, which was after the postconditioning phase, the time spent by each rat in the CPP apparatus was measured every day for 14 days. Food-induced reinstatement of CPP was performed by placing a single Froot Loop in the middle compartment where the rats were placed at the beginning of the session. The time spent in each compartment was recorded for 15 min during the test, extinction, and reinstatement days.
Locomotor activity was performed during the pre and post conditioning test. After each series of cocaine CPP, the assessments of either anxiety-like behavior (elevated plus-maze (EPM) test), sucrose consumption, spatial recognition memory (two-trial recognition task (TTRT) and spontaneous alternation test), or behavior in the forced swim test were performed with saline- and cocaine-treated rats after the expression and reinstatement phases (see Supplemental Data) as previously described (Atehortua-Martinez et al., 2019). Each rat was subjected to only one test, randomly chosen.
Ribonucleic acid (RNA) extraction, library preparation, and sequencing
For all groups, 24 h after the last drug administration, brains were removed and sectioned on ice in a brain block (1-mm thick), and micropunches of the NAc (core and shell) were snap frozen on dry ice and stored at −80°C in Qiazol to prevent degradation (Qiagen, France) until use. RNA extraction, library preparation, and sequencing parameters are detailed in Supplemental Data and were done as previously described (Vorspan et al., 2020). Total RNA was extracted from six rats from six CPP experiments.
Library preparation and Illumina sequencing were performed at the École normale supérieure genomic core facility (IBENS, Paris, France) as previously described (Vorspan et al., 2020). For CPPE and reinstatement studies, messenger (polyA+) RNAs from three pools of two NAc samples from six saline-treated, six CPPE, or six nCPPE rats were purified from 500 ng of total RNA using oligo (dT).
Read filtering, mapping, alignment filtering, read quantification, normalization, and differential analysis were performed using the Eoulsan pipeline (Jourdren et al., 2012). Reads were then aligned against the Rattus norvegicus genome from Ensembl version 84 using STAR (version 2.4.0k; Dobin et al., 2013). Normalization, statistical treatments, and differential analyses were performed using DESeq 1.8.3 and DESeq2 (Love et al., 2014).
Reverse transcription and real-time quantitative PCR
Reverse transcription of RNA was performed as previously described (Vorspan et al., 2020). Sequences of the primers used for amplification are presented in Supplemental Table 1. Amplification was detected by SYBR Green fluorescence on an ABI PRISM 7900HT sequence detection system (Applied Biosystems, France)). Details on the normalization and quantification are described in Supplemental Material.
Statistical analysis
All analyses were performed using R software, version 3.5.0 (R Core Team, 2020), and the appropriate packages as described below, except behavioral analysis.
Expression of cocaine-induced CPP
Scores of rats from the saline group were used to build (β, γ)-tolerance intervals of the scores in absence of cocaine, assuming a Gaussian distribution. The Assumption was checked graphically using normal probability plots. In addition, classical normality tests (Shapiro-Wilk, Shapiro-Francia, Anderson-Darling, and Liliefors) were used A (β, γ)-tolerance interval is an interval centered has a γ probability of containing a proportion β of the values of the population of interest (Burrows, 1963; Krishnamoorthy and Mathew, 2009). Two intervals were built: a [90, 95] interval (which should contain 90% of the scores without cocaine) and a [70, 95] interval (70% of the scores). Rats from the cocaine group were then classified as follows: rats with a score outside the [90, 95] interval, in favor of the cocaine chamber, as CPPE having an unlikely score should cocaine have no effect; rats with a score in the [70, 95] interval, as nCPPE having a score typical of the saline group; and the remaining rats, as unclear and discarded from further analysis. Tolerance intervals were built using the tolerance package for R (Young, 2010). The total number of rats is 401 rats pooled from 13 series of CPP “(301 cocaine-treated and 100 saline).”
Score evolution over time (extinction phase)
Daily CPP scores were analyzed using a mixed effects linear model, with the rat as a random effect on both the intercept and the slope, the day as a quantitative fixed effect (assuming a linear change over time), and the group as a fixed effect. Both the error term and the random effects were assumed to be Gaussian distributed. Assumptions were checked graphically using quantile-quantile plots and other standard methods. The model was fitted using maximum likelihood. Asymptotic likelihood-ratio tests were used to test the significance of the day and the group coefficients. This analysis was performed using the lme4 package (Bates et al., 2015).
Score comparison (reinstatement phase)
This analysis was performed on the (D13−D12) score differences using ANOVA.
Behavioral tests analysis
All data are expressed as the mean ± SEM. Student’s t-tests or two-way ANOVA followed by appropriate post hoc tests for multiple comparisons with Bonferroni correction were performed using GraphPad Prism version 7.00 (GraphPad Software, La Jolla, CA, USA). p-Value < 0.05 were considered significant.
RT-qPCR data analysis
ΔΔCq results are presented as a log2 fold change with respect to the designated group. Data were analyzed using one-way ANOVA between subjects, followed by Tukey’s test for post hoc comparisons. The level of significance was set at p < 0.05.
In addition, graph analysis from the RT-qPCR experiments was performed with the SARP.compo package using previously described methods (Curis et al., 2019). Briefly, all genes were considered nodes of a graph. Two nodes were connected if the ratio of the corresponding gene expression did not significantly change between groups. Genes belonging to different subgraphs were considered differentially expressed. With 23 genes and three groups of three samples, the optimal threshold to cut edges was set at 0.35 (95% confidence interval [0.3495; 0.3689], 10,000 simulations) to ensure a conservative Type I error rate below 5%.
Results
Expression of cocaine-induced CPP
Figure 1(b) shows the expression of cocaine-induced CPP. On average, rats receiving cocaine had significantly higher scores than saline-treated rats (116.7 ± 7.2 s vs −12.8 ± 6.1 s, respectively; t = 13.75; p < 0.0001, Student’s t-test corrected for unequal variances (df = 343.6)).
Statistical model for classification
The [70, 95]- and [90, 95]-tolerance intervals derived from the saline group were [−85.1, 59.4] and [−128, 102], respectively (Figure 1(b)). Hence, rats from the cocaine group with a score higher than 102 s were classified as expressing a strong CPP (“CPPE” group), whereas rats with a score between −85 and 59 s were classified as not expressing a CPP (“nCPPE” group). This classification led to 177 CPPE (58.8%), 78 nCPPE (25.9%), and 46 unclear (15.3%) rats in the cocaine group. The unclear group was composed of aversive rats (n = 6), a continuum between CPPE and nCPPE rats (n = 31), and a continuum between nCPPE and aversive rats (n = 9). To highlight the interindividual variability to the rewarding effects of cocaine the two contrasted CPPE and nCPPE groups were chosen and “unclear” rats were not considered further in the analysis.
Behavioral studies
The results of the cognitive and emotional behavioral assessments of the saline, CPPE and nCPPE groups are shown in Figure 1(c). Using one-way ANOVAs, none of these tests shown significant difference between the three groups of rats (CPPE cocaine-treated rats, nCPPE cocaine-treated rats, and saline-treated rats)—spontaneous alternation test (number of alternations: F(2, 34) = 0.193, NS), TTRT test (entries: F(2, 38) = 1.384, NS; time spent in the novel arm: F(2, 38) = 1.212, NS), EPM test, considering either the entries (F(2, 19) = 0.199, NS) or the time spent in the open arms: F(2, 19) = 1.516, NS), forced swimming test (immobility time: F(2, 20) = 1.090, NS), sucrose consumption test (sucrose preference: F(2, 21) = 1.026, NS), and locomotor activity during either the pretest (F(2, 354) = 2.401, NS) or the test phase (F(2, 354) = 3.356, NS).
Reinstatement of cocaine-induced CPP
The average time spent by each group in the cocaine- or saline-associated compartments during the extinction and reinstatement phases is shown in Figure 2(b). There was a large amount of variability between the rats both on the intercept (estimated standard deviation: σ = 71.8 s, 95% CI [59.7; 85.9]) and the slope ( σ = 9.2 s/day, [7.5; 11.1]). However, on average, the score remained stable during the 12-day period (slope: −1.4 s/day [−3.4; 0.59], p = 0.1664) in all three groups. The average score was 8.69 s in the saline group [−15.4; 32.8], which was significantly different than that in the CPPE group (+136.8 s [+104.9; +168.7]) but not in the nCPPE group (−22.8 s [−66.5; +11.7]).
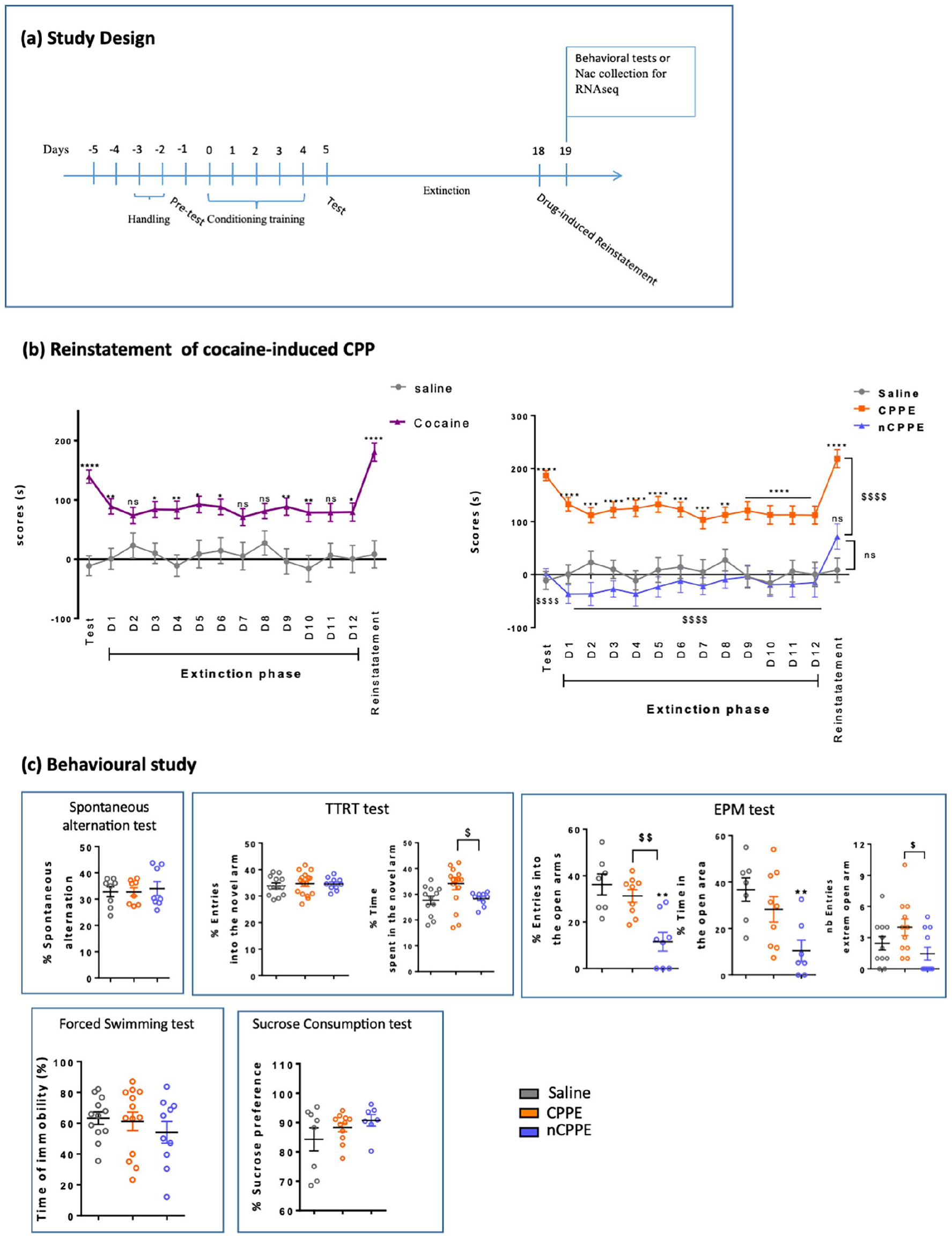
Reinstatement of cocaine-induced CPP. (a) Study design. (b) Extinction and reinstatement of CPP. Values are expressed as the mean ± SEM (saline, n = 45; CPPE, n = 54; nCPPE, n = 25). (c) Results of cognitive and emotional behavioral assessments 24 h after CPP reinstatement in saline-treated, CPPE, and nCPPE rats. Values are expressed as the mean ± SEM (n = 7–16 for each group, spontaneous alternation: saline = 8, CPPE = 8, nCPPE = 8; TTRT test: saline = 13, CPPE = 16, nCPPE = 10; EPM test: saline = 7, CPPE = 9, nCPPE = 8; forced swimming test: saline = 12, CPPE = 12, nCPPE = 10; sucrose consumption test: saline = 8, CPPE = 11, nCPPE = 7). Comparisons were performed using one-way or two-way repeated measures ANOVA followed by post hoc tests with Bonferroni correction.
Behavioral study
The results of the cognitive and emotional behavioral assessments of the saline, CPPE and nCPPE groups are described in Figure 2(c). Using one-way ANOVA, in the TTRT test, the time spent in the new arm was significantly increased in the CPPE group compared to the nCPPE group (F(2, 35) = 9.583, p = 0.0005), but the number of entries in the new arm did not significantly differ (F(2,35) = 0.188, NS); in the EPM test, the entries into the open arms and the extreme open arms were significantly increased in the CPPE group compared to the nCPPE group (F(2, 20) = 9.621, p < 0.001; F(2,20) = 3.361, p < 0.05; respectively), and entries and time were decreased in the nCPPE group compared to the saline group (F(2,20) = 9.621, p < 0.001; F(2, 20) = 3.361, p < 0.001; respectively). Results of the other tests were not significantly different between the three groups—spontaneous alternation test (alternation: F(2, 21) = 0.123, NS), forced swimming test (immobility time: F(2, 23) = 1.465, NS), and sucrose consumption test (sucrose preference: F(2, 20) = 0.151, NS).
Expression and reinstatement of Froot Loops-induced CPP
Supplemental Figure 1(a) shows the expression of Froot Loops-induced CPP. In average, rats receiving Froot-Loops had significantly higher scores than saline rats (83.2 ± 9.5 s vs −6.5 ± 11.5 s, t = 5.87; p < 0.0001, Student t-test corrected for unequal variances (df = 81.72)).
Statistical model for classification
The (70.95)- and (90.95)-tolerance intervals derived from the saline group were, respectively, [−38.4; 33.4] and [−167.3; 162.3]. Hence, rats from the Froot Loops group having a score higher than 147.5 s were classified as expressing a strong CPP (“CPPE” group), whereas rats with score between −60 and 49.7 s as not expressing any particular CPP (“nCPPE” group). This classification led to 25 CPPE (38.5%), 21 nCPPE (32.3%), and 19 unclear (29.2%) rats in the Froot Loops group. “Unclear” rats were not considered further in the analysis.
Average time spent by each group in the Froot Loops or saline-associated compartments during the extinction and the reinstatement phases is shown in Supplemental Figure 1(a). There was a large variability between rats both on the intercept (estimated standard deviation: σ = 32.0 s (95% CI [14.1; 55.0]) and the slope ( σ = 5.5 s, [3.6; 8.0]). In average, the score slightly decreased along the 14-days period (slope: −3.3 s/day, [−5.7; −0.9], p = 0.006921) in all three groups. The average score was 19.1 s in the saline group ([−6.1; 44.3]); it was significantly different in the CPPE group (+144.3 s [+108.3; +180.4]), but not in the nCPPE group (−20.2 s [−62.4; +22.1]).
After reinstatement, the score was not significantly increased in the Saline group (−12.5 s [−100.1; +75.1]), the CPPE group (+31.5 s [−23.8; +86.8]), and nCPPE groups (+12.3 s [−84.2; +108.7], p = 0.648, one-way ANOVA).
Differentially expressed genes after CPPE
RNA-seq exploration
Among the 22,961 expressed genes in the NAc, 371 were significantly differentially expressed between the nCPPE versus saline-treated rats (Figure 3(a)). The list of the top 20 annotated differentially expressed genes (DEGs) in this comparison, based on the adjusted p-value, is presented in Table 1(a). No Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway was found significantly enriched in this comparison. Although the CPPE and nCPPE rats received the same cocaine treatment, the expression levels of 1797 genes were significantly different between these two groups. The top 20 annotated DEGs in this comparison, based on the adjusted p-value, are presented in Table 1(b). Supplemental Table 2 displays the 20 most significantly enriched KEGG pathways related to the DEGs in the CPPE versus nCPPE rats. Among them, pathways such as the Ras signaling pathway, the mitogen-activated protein kinase (MAPK) pathway, and serotonergic, glutamatergic, and gamma-aminobutyric acid (GABA)ergic synapses have previously been associated with cocaine addiction. However, based on the adjusted p-value, only four genes were found to be differentially expressed in the NAc of the CPPE rats compared to the saline-treated rats: Egr2 (early growth response 2), Nr4a1 (nuclear receptor subfamily 4), Junb (jun B proto-oncogene), and Zbtb37 (zinc finger and bric-a-brac, tramtrack, broad complex (BTB) domain containing 37; Table 1(c)). Although all these genes are immediate early genes (IEGs), no KEGG pathway was found significantly enriched in this comparison. As shown in the Venn diagrams (Figure 3(a)), no common DEG was identified across the three comparisons, however the four DEGs in the CPPE versus saline group comparison were also found in the CPPE versus nCPPE group comparison (Figure 3(a)).
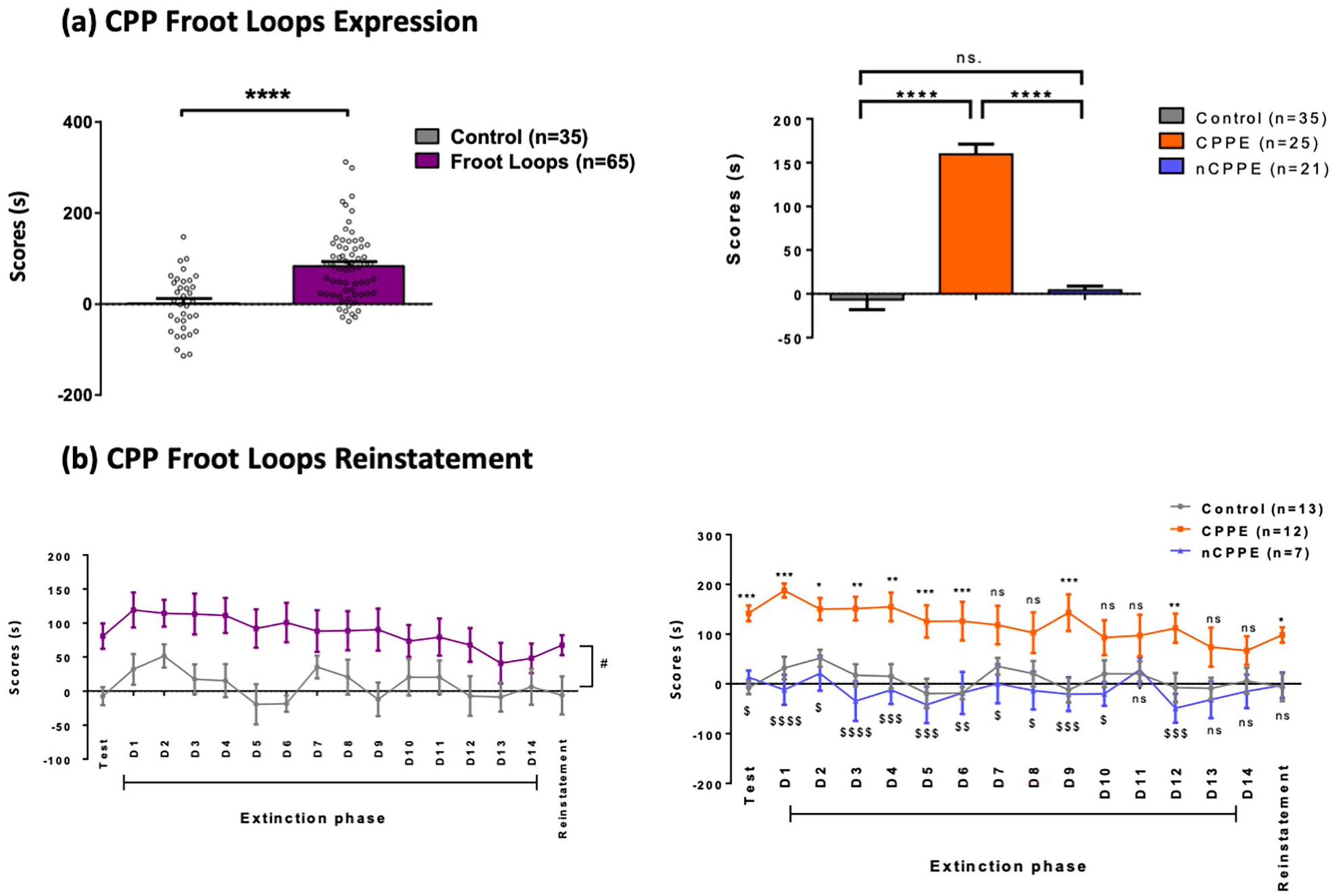
(a) Froot Loops-induced CPPE and (b) reinstatement. Values are expressed as the mean ± SEM.
List of the DEGs in the three comparisons after cocaine-induced CPPE.
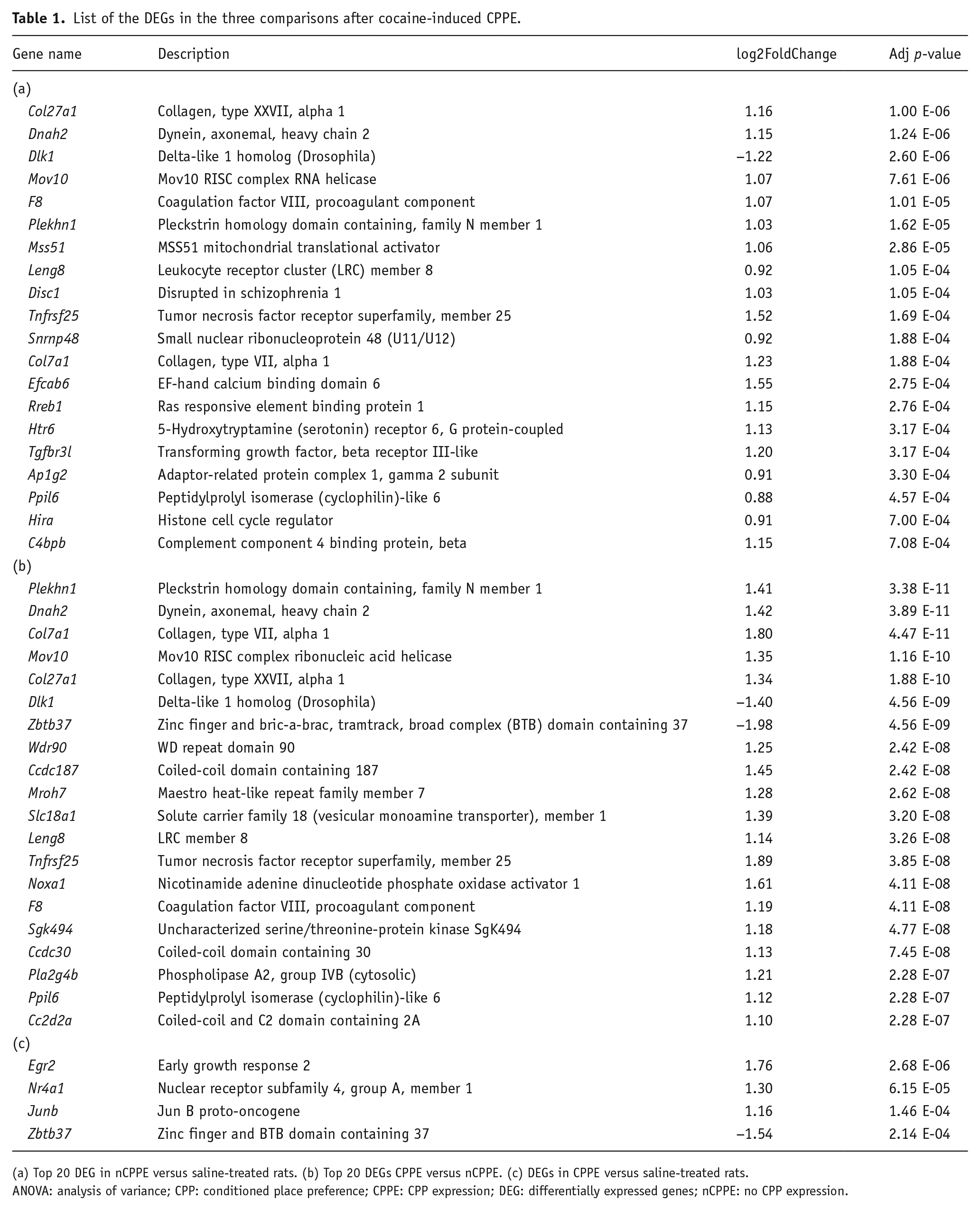
(a) Top 20 DEG in nCPPE versus saline-treated rats. (b) Top 20 DEGs CPPE versus nCPPE. (c) DEGs in CPPE versus saline-treated rats.
ANOVA: analysis of variance; CPP: conditioned place preference; CPPE: CPP expression; DEG: differentially expressed genes; nCPPE: no CPP expression.
Confirmation of DEGs after CPPE using RT-Qpcr
RT-qPCR experiments were performed in the same samples to validate the observed RNA-seq gene expression modulation. About 17 genes were chosen based on the adjusted p-value obtained in RNA-seq experiments and on the literature (Table 2(a)). For the majority of the 17 genes tested, the results obtained using qPCR were in accordance with the RNA-seq data. Of the four genes identified using RNA-seq in the comparison between CPPE and saline-treated rats, three were validated by qPCR. The modulation of seven genes from the nCPPE versus saline-treated rat comparison was validated, and two genes (F8 and Htr6) were found modulated in the opposite direction relative to the RNA-seq results. Finally, 15 out of the 17 tested genes from the CPPE versus nCPPE comparison were validated, with 1 gene (Dlk1) found modulated in the opposite direction relative to the RNA-seq results. The results of the univariate analysis were confirmed by a disjoint graphs network analysis that showed a group of six genes (Egr1, Arc, JunB, Fos, Egr2, and Nr4a1) with similar modulation in the CPPE versus saline comparison (Figure 3(c)). In the nCPPE versus saline comparison, eight genes (Col27a1, F8, DNAh2, Leng8, Mss51, Mov10, Ap1g2, and Plekhn1) were similarly modulated (Figure 3(c)). In the nCPPE versus CPPE comparison, the regulation of one gene (Dlk1) was found to be different from that of all the other tested genes (Figure 3(c)). Of note, the same analysis applied on the RNASeq results for these genes lead to the same graph structure (Supplemental Figure 2). The results of the disjoint graphs analyses in RT-qPCR and RNA-seq after CPPE indicate that, even if the regulation of some genes is not confirmed with the univariate analyses, the relative expression levels of these genes observed with the two techniques are comparable.
Validation of mRNA regulation in the NAc of saline-treated, CPPE, and nCPPE rats using RT-qPCR.
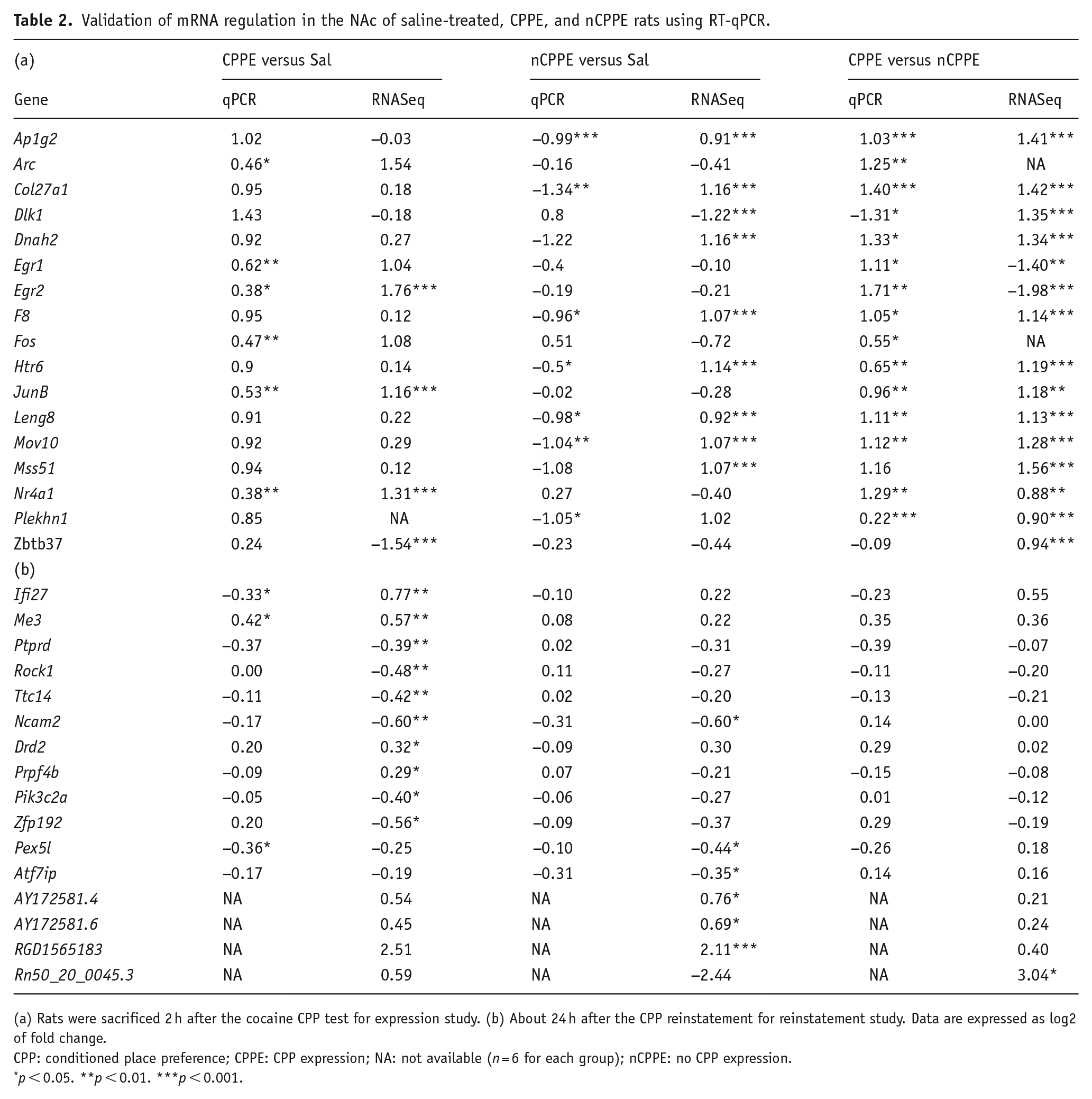
(a) Rats were sacrificed 2 h after the cocaine CPP test for expression study. (b) About 24 h after the CPP reinstatement for reinstatement study. Data are expressed as log2 of fold change.
CPP: conditioned place preference; CPPE: CPP expression; NA: not available (n = 6 for each group); nCPPE: no CPP expression.
p < 0.05. **p < 0.01. ***p < 0.001.
DEGs after CPP reinstatement
RNA-seq exploration
Of the 32,403 expressed genes observed in the NAc, only a few were found differentially expressed in the three comparisons (Figure 4(a)). Six genes were differentially expressed in the NAc between the nCPPE versus saline-treated rats after p-value adjustment for multiple comparisons (Supplemental Table 4). Interestingly, only one (Rn50_20_0045.3) was differentially expressed between the two cocaine-treated groups (CPPE vs nCPPE comparison). There were 10 DEGs in the NAc in the CPPE versus saline-treated rat comparison (Supplemental Table 4). As shown in the Venn diagrams, no common DEG was identified across the three comparisons (Figure 4(a)). Ncam2 was the only gene found significantly downregulated in the CPPE versus saline and nCPPE versus saline comparisons. KEGG pathway enrichment analyses of the DEGs for the three comparisons did not identify any significantly enriched pathway (Figure 5).
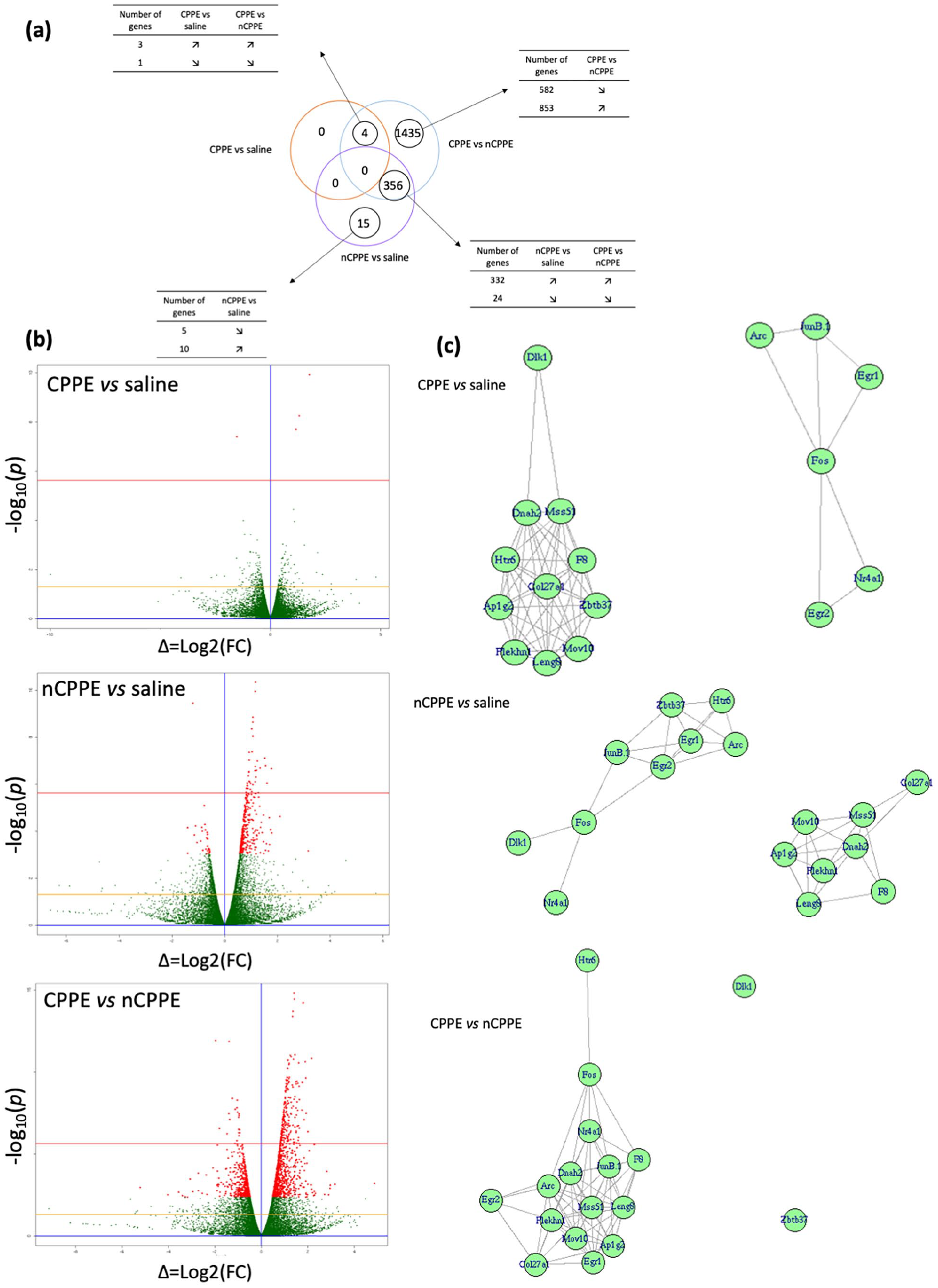
Expression of cocaine-induced CPP. (a) Venn diagram of the DEGs in the NAc. Modulation of the DEGs in the three tested comparisons after cocaine-induced CPPE are detailed. (b) RNA-Seq results with Vulcano Plot. DEG in CPPE versus saline, nCPPE versus saline, and nCPPE versus CPPE comparisons were identified in red p < 0.05; yellow line p = 0.05 and red line p = 0.05 with Bonferroni correction (n = 6 for each group). (c) Graph made from coexpression data of DEGs. Graph obtained from SARP.compo (see Methods Section for details).
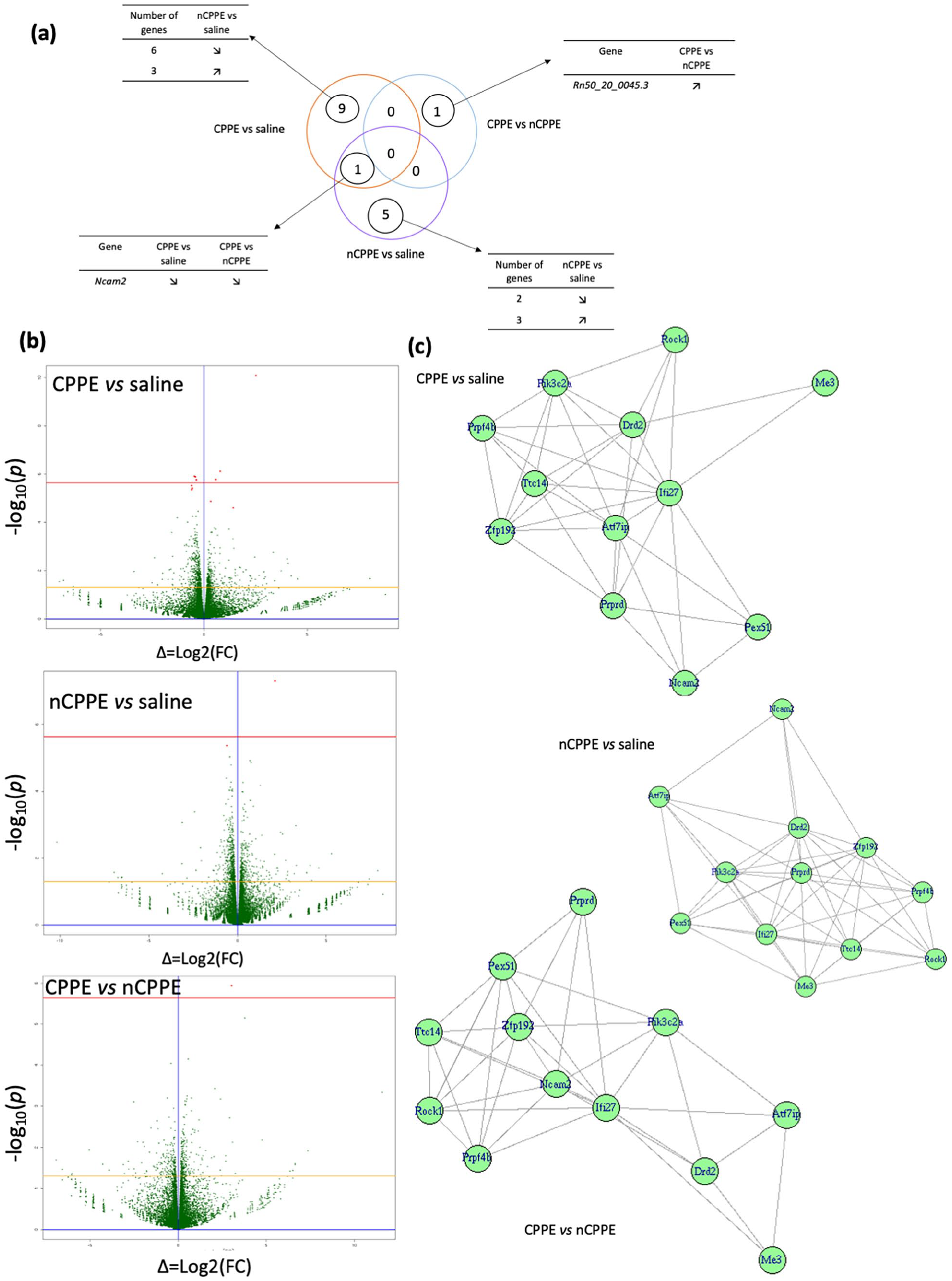
Reinstatement of cocaine-induced CPP. (a) Venn diagram of the DEGs in the NAc. Modulation of the DEGs in the three tested comparisons after 24 h cocaine-induced CPP reinstatement are detailed. (b) RNA-Seq results with Vulcano Plot. DEG in CPPE versus saline, nCPPE versus saline, and nCPPE versus CPPE comparisons were identified in red p < 0.05; yellow line p = 0.05 and red line p = 0.05 with Bonferroni correction (n = 6 for each group). (c) Graph made from coexpression data of DEGs. Graph obtained from SARP.compo (see Methods Section for details).
Confirmation of DEGs after CPP reinstatement using RT-qPCR
RT-qPCR experiments were performed to validate the RNA-seq results after reinstatement. All 16 DEGs identified in at least one of the three comparisons were tested (Table 1(b)). Four genes (AY172581.4, AY172581.6, RGD1565183, and Rn50_20_0045.3) displayed very low expression levels and could not be quantified. Therefore, the only DEG identified in the CPPE versus nCPPE comparison (Rn50_20_0045.3) could not be validated using RT-qPCR. Only 1 of the 10 DEGs identified using RNA-seq in the CPPE versus saline-treated rat comparison (Me3) was validated by RT-qPCR, while Ifi27 showed a significant yet opposite regulation. The trend for a downregulation of Pex5l in the CPPE rats relative to the saline-treated rats was confirmed with a significant downregulation. None of the six significant regulations observed in the nCPPE versus saline-treated rat comparison could be validated using RT-qPCR. A disjoint graphs network analysis did not detect, for any of the three tested comparisons, groups of genes with differential expression/modulation (Figure 4(c)); this was also the case when applied to the RNA-Seq data for these genes.
RT-qPCR Froot Loops after expression and reinstatement
Expression modulations of the genes identified in the cocaine-induced CPPE and reinstatement experiments were also analyzed after Froot loops-induced CPPE and reinstatement using RT-qPCR. In the CPPE versus Sal comparison, two genes were found differentially expressed (Mov10 and Plekhn1) while Mov10 was the only gene differentially expressed in the nCPPE versus Sal comparison after food CPPE (Supplemental Table 3(a)). However, disjoint graphs network analysis did not detect, for any of the three tested comparisons, groups of genes with differential expression/modulation as seen in Supplemental Figure 3(a)–(c). As shown in Supplemental Table 3(b) none of the genes (identified in the cocaine-induced CPP) examined were found significantly regulated in the food reinstatement. Similarly, the disjoint graph network analysis did not detect genes (identified in the cocaine-induced CPP) with significant differential expression in the Froot loops-induced reinstatement (Supplemental Figure 3(d)–(f)).
Discussion
This study aimed to demonstrate and characterize the variability in the expression of the rewarding effects of cocaine in the CPP paradigm. We report three main findings: (i) despite receiving the same cocaine regimen in the CPP paradigm and presenting no differences in other behavioral tests, at least two groups of rats, those that do (CPPE) and do not (nCPPE) present rewarding effects, can be identified; (ii) gene expression observed in the NAc at the time of expression of CPP revealed specific differential effects on mRNA of IEGs in the CPPE rats; and (iii) behavioral and transcriptomic differences can be detected after extinction and reinstatement phases. These findings are of interest in the context of identifying the mechanisms underlying individual variability in the response to cocaine’s rewarding effects.
In the drug-induced CPP paradigm, the results are usually expressed by averaging the scores, to reflect the performance of rats treated with the studied drug, and comparing them to the average results from the control group. However, some previous studies have raised the issue that computing a group average may hide differences between population subgroups in the acquisition or expression of CPP. dela Cruz et al. (2009) established a criterion for the expression of a cocaine CPP in male Sprague–Dawley rats based on the time spent in the initially less-preferred chamber during the test session that effectively discriminated control from cocaine-conditioned rats. Differences in population subgroups have also been observed in 3,4-methylenedioxymethamphetamine (MDMA)-induced CPP, with some animals showing a strong CPP and others showing an aversion to the MDMA-paired chamber (Daza-Losada et al., 2007). This has also been shown for morphine CPP, with the scores in rats being related to urinary and blood plasma metabolic profiles (Zaitsu et al., 2014). These reports suggested the need for an analysis method for CPP data that allows the effective identification of subgroups of subjects within a treatment group without additional behavioral or biochemical tests. The present study validated such a method for the expression of the rewarding effects of cocaine in the CPP paradigm, and also characterized this response variability. The proposed method used the results of a CPP study in a control (saline) group to determine the expected range of scores for “no effect” drugs and then classified rats receiving the challenge drug, here cocaine, into a group of rats experiencing a substantial CPP (“vulnerable” rats) and a group of rats experiencing a CPP similar to the controls (“nonvulnerable” rats). We have also shown that this model can also be applied to a natural reward (see Supplemental Data).
The set of basic behavioral tests performed after the CPP test showed that the behavioral responses of the CPPE and nCPPE rats were similar in classic anxiety and hedonic tests. In the same way, after the CPP test, CPPE and nCPPE rats, without being under the influence of drugs, showed similar results in the TTRT (memory test). Therefore, the observed differences in cocaine-CPPE scores could not be associated to differences in these basic behaviors and at memory effects.
A whole-transcriptome analysis of the differences in the rats identified as CPPE and nCPPE was carried out in the NAc, a brain structure critically involved in the expression of reward-dependent behavior, especially CPP (Bardo, 1998). A key finding of this study is the small number of genes that were differentially expressed in the NAc between CPPE and saline-treated rats. Moreover, all the identified DEGs were IEGs. We validated the differential regulation of six IEGs (Arc, Egr1, Egr2, Fos, JunB, and Nr4a1) in the CPPE group using RT-qPCR. IEGs have been repeatedly shown to be regulated by cocaine in several paradigms (Bisagno and Cadet, 2019; Harlan and Garcia, 1998; Hope, 1998). The results obtained in the NAc of the CPPE group in the current study are consistent with the previously reported significant increase in Fos and Egr1 in the NAc of mice after cocaine-CPPE (Muñiz et al., 2017). Our results extend this regulation to other IEGs only in the CPPE group. IEGs have been extensively used to identify neuronal populations activated by a stimulus and have been linked to the formation of long-term memory (Minatohara et al., 2015). Fos, the most studied IEG in the context of addiction, has been shown to be involved in learning, memory, and motivational processes in the case of cocaine (Zhang et al., 2006). We showed that IEGs were also differentially expressed between the CPPE and nCPPE groups, suggesting that they might be implicated in the expression of CPP and anticipation of rewarding effects. It is important to note that food-CPPE did not identify any change in IEG expression (see Supplemental Data). Taken together, these results suggest that the observed regulation of IEGs were specific to rats that express cocaine place preference in the CPP paradigm.
The large number of DEGs between the CPPE and nCPPE rats was a rather unexpected result. Indeed, these two groups of rats received the same cocaine treatment during CPP conditioning. Most of the significantly enriched KEGG pathways in this comparison have been associated with cocaine-induced effects in the CPP paradigm (Bardo, 1998; Beninger and Gerdjikov, 2004; McBride et al., 1999). Specifically, MAPK signaling in the NAc has been repeatedly shown to be important for cocaine CPP (Brami-Cherrier et al., 2005; Janes et al., 2009; Mannangatti et al., 2015; Miller and Marshall, 2005). Calcium signaling has also been shown to play an important role in CPP with cocaine (Alaghband et al., 2018; Liang et al., 2017). Similarly, glutamatergic signaling has been demonstrated to be involved in cocaine reinstatement (Briand et al., 2016). Cannabinoid receptors have been found to modulate cocaine effects (Gobira et al., 2019). GABAergic signaling plays a crucial role in cocaine-CPPE (Zhang et al., 2018). Cocaine reinstatement has been shown to be influenced by circadian stimuli (Sorg et al., 2011). Taken together, the pathways and genes identified in the CPPE versus nCPPE comparison might represent the most crucial ones regarding cocaine’s rewarding effects in the different phases of the CPP paradigm.
The behaviors of the two subgroups CPPE and nCPPE evolved differently during the extinction phase and after reinstatement. Given the importance of IEGs in memory processes, this suggested that these genes may be important for the mechanisms involved in these long-lasting behavioral differences. Indeed, the battery of behavioral tests performed after reinstatement showed that CPPE rats have greater memory expression based on the TTRT test compared to the nCPPE group without altered spatial orientation while nCPPE group did not display cognitive impairment. This is consistent with the reported greater efficacy and velocity to learn spatial tasks in memory tests of Lewis rats (vulnerable to addiction; Cadoni, 2016; Miguéns et al., 2013) compared to Fischer 344 rats (less vulnerable to addiction; Fole et al., 2017; van der Staay et al., 2009). In the EPM test, the nCPPE group had a lower percentage of entries into the open arms and a lower number of visits to the extremes arms than the CPPE group, reflecting anxious behavior. This result corroborated previous results suggesting in various models of vulnerability that resilient animals exhibited anxiety-like behaviors (Dellu et al., 1996; Flagel et al., 2014). Behavioral traits, such as locomotor activity, have been used to predict the development of stronger addiction-like behaviors (Allen et al., 2007; Dellu et al., 1996; Piazza et al., 1989, 2000). Locomotor activity, measured after CPPE, was similar between CPPE and nCPPE groups which excludes basal locomotor activity as an explanation for the differences found in EPM results after reinstatement. However, it has been shown that cocaine-associated contextual cues are capable of eliciting anxiogenic-like behavior in the absence of the drug (DeVries and Pert, 1998). Our results are concordant with this anxiogenic-like behavior after reinstatement for nCPPE group.
Another key finding was that after reinstatement, only a few genes were found modulated in the NAc of the three groups of rats. The study was carried out 24 h after the reinstatement which could explain the small number of modulated genes. However, in the CPPE group, the 10 genes found differentially expressed compared to the saline group might play an important role in reinstatement to cocaine in the CPP test. Among those genes, Drd2 encodes the D2 subtype of the dopamine receptor, and was upregulated in the CPPE group after reinstatement. An upregulation of this gene has been proposed to be involved in neural plasticity mechanisms underlying the strengthening of learning the association between drug and environment in mice (Muñiz et al., 2017). A clinical study showed that genetic variations associated with DRD2 expression affected explicit memory, specifically for rewarding stimuli related to cocaine addiction memories (Richter et al., 2017), and another clinical study provided evidence for the involvement of DRD2 in the early stages of addiction and supported the notion that genetically driven interindividual differences in dopaminergic transmission mediate reward sensitivity and risk of smoking (Macare et al., 2018).
After reinstatement, Ncam2 (neural cell adhesion molecule 2) was found to be significantly decreased in both the CPPE and nCPPE groups compared to the saline group in the RNA-seq experiments, while the RT-qPCR experiments revealed a trend. This gene is involved in the formation and maintenance of dendritic and axonal compartmentalization in the olfactory system (Winther et al., 2012). In the cerebral cortex, Ncam2 has been shown to regulate neurite outgrowth as well as synapse formation and maintenance (Leshchyns’ka and Sytnyk, 2016; Parcerisas et al., 2020; Sheng et al., 2015, 2019). The observed downregulation of Ncam2 expression in the two cocaine-treated groups (CPPE and nCPPE) might be a long-term consequence of cocaine treatment independent of the associated drug compartment or rewarding effects and suggests an effect on neural plasticity. Unfortunately, the only DEG between the CPPE and nCPPE groups was not detected in the RT-qPCR experiments and is not annotated (Rn50_20_0045.3), as this could be a key marker to better understand the differences in vulnerability to the rewarding effects of cocaine.
Conclusion
We propose here a method that allows for the dichotomous categorization of individual subjects within a cocaine (data with food included in the Supplemental Data) treatment group as CPP expressing or non-CPP expressing as a complementary approach to consider interindividual variability in responses among subjects. Moreover, we also followed the two groups of rats during an extinction and a reinstatement phase. To our knowledge, this is the first study to explore, in rats, the NAc whole transcriptome in cocaine-CPPE tests and reinstatement. However, our study performed with one dose and only on male rats also presents limitations. (i) We have a continuum between the CPPE and nCPPE categories, with approximately 15% of the cocaine rats who were unclear. This biostatistical model could be improved in subsequent experiments. (ii) Due to the small sample size in the transcriptomic analyses, we may have lacked the power to identify regulation of certain genes in the CPPE versus saline comparison. (iii) It would be interesting to extend the present study to female rats, to different time points and to other brain regions involved in addiction. Nonetheless, the unique biostatistical animal model presented here can be used to identify genes and mechanisms involved in vulnerability and resilience to addiction.
Supplemental Material
sj-docx-1-jop-10.1177_02698811221123047 – Supplemental material for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence
Supplemental material, sj-docx-1-jop-10.1177_02698811221123047 for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence by Luisa Alessandra Atehortua Martinez, Emmanuel Curis, Nawel Mekdad, Claire Larrieu, Cindie Courtin, Laurent Jourdren, Corinne Blugeon, Jean-Louis Laplanche, Bruno Megarbane, Cynthia Marie-Claire and Nadia Benturquia in Journal of Psychopharmacology
Supplemental Material
sj-tif-2-jop-10.1177_02698811221123047 – Supplemental material for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence
Supplemental material, sj-tif-2-jop-10.1177_02698811221123047 for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence by Luisa Alessandra Atehortua Martinez, Emmanuel Curis, Nawel Mekdad, Claire Larrieu, Cindie Courtin, Laurent Jourdren, Corinne Blugeon, Jean-Louis Laplanche, Bruno Megarbane, Cynthia Marie-Claire and Nadia Benturquia in Journal of Psychopharmacology
Supplemental Material
sj-tif-3-jop-10.1177_02698811221123047 – Supplemental material for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence
Supplemental material, sj-tif-3-jop-10.1177_02698811221123047 for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence by Luisa Alessandra Atehortua Martinez, Emmanuel Curis, Nawel Mekdad, Claire Larrieu, Cindie Courtin, Laurent Jourdren, Corinne Blugeon, Jean-Louis Laplanche, Bruno Megarbane, Cynthia Marie-Claire and Nadia Benturquia in Journal of Psychopharmacology
Supplemental Material
sj-tif-4-jop-10.1177_02698811221123047 – Supplemental material for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence
Supplemental material, sj-tif-4-jop-10.1177_02698811221123047 for Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence by Luisa Alessandra Atehortua Martinez, Emmanuel Curis, Nawel Mekdad, Claire Larrieu, Cindie Courtin, Laurent Jourdren, Corinne Blugeon, Jean-Louis Laplanche, Bruno Megarbane, Cynthia Marie-Claire and Nadia Benturquia in Journal of Psychopharmacology
Footnotes
Acknowledgements
The authors thank Astrid Morillo for animal care and PharmAnima Platform, UMS 3612 CNRS—US25 Institut National de la Santé et de la Recherche Médicale, Faculty of Pharmacy, University of Paris Cité, Paris, France, for animal care. C. Marie-Claire is supported by the Centre National pour la Recherche Scientifique (CNRS).
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Institut National de la Santé et de la Recherche Médicale and the IdEx Université de Paris Cité, ANR-18-IDEX-RM11J20IDXB3.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.