
Abstract
This paper introduces a new construct, the ‘pivotal mental state’, which is defined as a hyper-plastic state aiding rapid and deep learning that can mediate psychological transformation. We believe this new construct bears relevance to a broad range of psychological and psychiatric phenomena. We argue that pivotal mental states serve an important evolutionary function, that is, to aid psychological transformation when actual or perceived environmental pressures demand this. We cite evidence that chronic stress and neurotic traits are primers for a pivotal mental state, whereas acute stress can be a trigger. Inspired by research with serotonin 2A receptor agonist psychedelics, we highlight how activity at this particular receptor can robustly and reliably induce pivotal mental states, but we argue that the capacity for pivotal mental states is an inherent property of the human brain itself. Moreover, we hypothesize that serotonergic psychedelics hijack a system that has evolved to mediate rapid and deep learning when its need is sensed. We cite a breadth of evidences linking stress via a variety of inducers, with an upregulated serotonin 2A receptor system (e.g. upregulated availability of and/or binding to the receptor) and acute stress with 5-HT release, which we argue can activate this primed system to induce a pivotal mental state. The pivotal mental state model is multi-level, linking a specific molecular gateway (increased serotonin 2A receptor signaling) with the inception of a hyper-plastic brain and mind state, enhanced rate of associative learning and the potential mediation of a psychological transformation.
Introduction
One way of explaining quantum change experiences is that they represent a kairos, a turning point in the life journey where major change simply must occur because the person in unable or unwilling to continue in his or her present course. It is a point of desperation, a breaking point where ‘something has to give’ – and it does.
Psychological transformation (defined here as rapid, marked and enduring psychological change, where ‘psychological’ refers to perception, cognition and action or behaviour) has been the focus of previous psychological and philosophical texts (Miller and C’de Baca, 2001; Paul, 2014) as well as influential therapeutic programmes (Wilson and Cohen, 2015), but it has received surprisingly little formal scientific investigation and past definitions have been vague. This paper aims to rectify this by proposing a multi-level, biologically informed, context-dependent and process-based approach to the phenomenon. Advancing previous work, we introduce a potentially useful new construct, the pivotal mental state (PiMS). Focusing on psychological transformation as a process is a simple but important aspect of our approach that enables us to offer a potential explanation for how transformative experiences (experiences mediating psychological transformation) can manifest into extremely divergent outcomes, such as positively life-changing spiritual breakthrough versus descent into a potentially life-long psychotic illness. More concretely, we propose that however divergent the nature of the transformations themselves, many can be traced to somewhat consistent triggering conditions, with chronic stress being a primer and acute stress a trigger. In what follows, we highlight striking similarities in the conditions of induction, neuropharmacology, neurobiology and psychology of transformative experiences. We demonstrate that these similarities are most compelling when one focuses on the states preceding and mediating psychological transformations, but become obscured when one selectively attends to the products of the transformations themselves.
In the same way that traumatic experiences can trigger post-traumatic stress disorder (PTSD) or post-traumatic growth (Tedeschi, 1999), we hypothesize that intense periods of psychological crisis can serve to kindle conditions for major, potentially lasting, psychological change, pivotable either towards illness or wellness (Figure 1). This process will later be linked to the phenomenon of bifurcation as described in dynamical systems theory (Kielhöfer, 2011). PiMSs share many parallels with so-called ‘quantum change’ experiences, which have been defined as ‘vivid, surprising, benevolent, and enduring personal [psychological] transformation[s]’ (Miller and C’de Baca, 2001). As will become clear, however, we offer a more precise state (as opposed to outcome) focused, neurobiologically grounded definition of PiMSs that links their hypothesized neuropharmacology and systems-level neurophysiology with their phenomenology. Our broad definition of PiMSs is that they are transient, intense hyper-plastic mind and brain states, with exceptional potential for mediating psychological transformation. We sharpen this definition by suggesting three key identifying criteria: (a) elevated cortical plasticity, (b) an enhanced rate of associative learning and (c) a unique capacity to mediate psychological transformation.
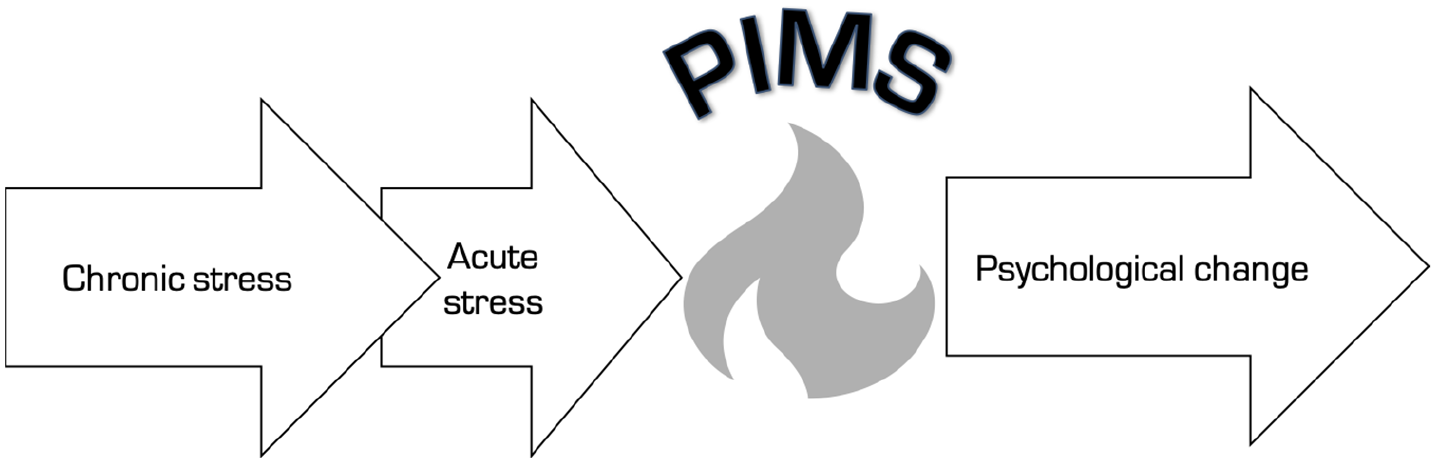
Process-based representation of the pivotal mental state (PiMS). Chronic stress is the primer and acute stress is the trigger of a PiMS, which functions as a mediating state for rapid psychological change.
The study of the nature and causes of PiMSs and their close association with psychological transformation is a central focus of this review. Our approach is multi level and integrative, addressing the neurobiology, pharmacology and physiology of PiMSs as well as their psychology. In keeping with popular contemporary psychological perspectives (Hayes, 2019; Hayes et al., 2013) as well as the biopsychosocial approach to mental health (Engel, 1977), we are mindful of the essential role played by context in shaping the quality and influence of PiMSs. It is our view that the principle of qualifying the longer-term impact of PiMSs by the context(s) or ‘relational frames’ (Hayes, 2019) in which they arise subside and potentially recur is vital for resolving the wellness versus pathology paradox, namely whether positive or negative changes in mental health follow from a PiMS.
Unlike past literature on quantum change, our PiMSs model does not favour positive outcomes over negative, speaking to an essential context and relational dependency, where context refers to both biological (e.g. polygenic) predisposition as well as the individual’s immediate and remote environmental context. Thus, the term ‘context’ is used here in an extended biopsychosocial and temporal way (Figure 2).
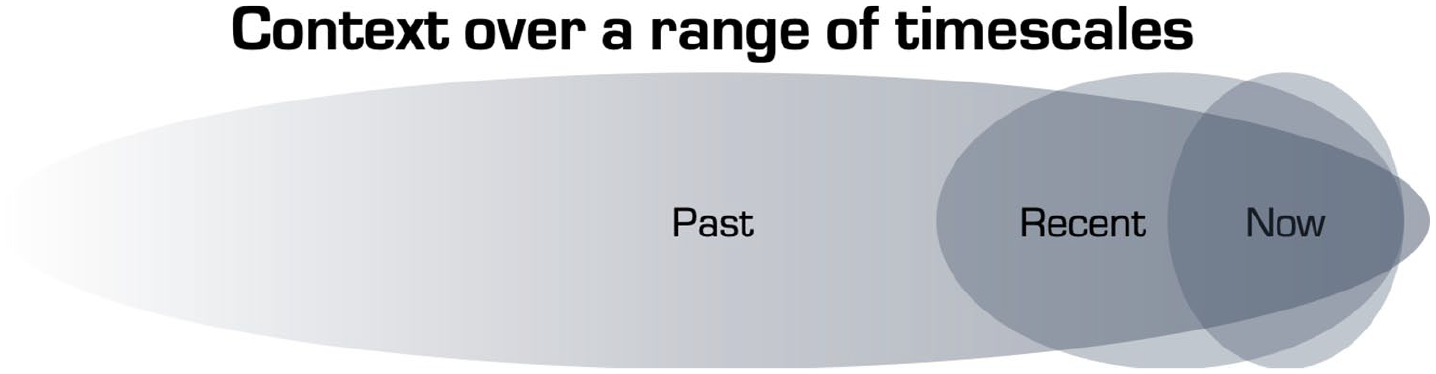
Schematic that acknowledges the breadth of timescales over which contextual factors can influence one’s current state. The boundaries between the three timeframes are arbitrary but one may think of ‘past’ in terms of, for example, an enriched versus adverse childhood or any past trauma but it can also include what is inherited (i.e. genetically encoded). The ‘recent’ past might include such things as a recent relationship breakdown or a bereavement, ongoing cultural or social crisis, or recent success or cause for celebration. What is happening in one’s immediate environment (i.e. ‘now’) might be easier to observe, describe and manipulate but is not necessarily the most potent influencer of one’s present state. This schematic is intended to acknowledge the temporal range of contextual influences that can impinge on the quality and outcome of a pivotal mental state (PiMS).
If we are correct that context plays a key role in shaping the outcome of a PiMS, an important implication is that contextual factors need to be treated with special attention and therapeutic care (where possible) if an individual’s psychological wellbeing is to be safeguarded, whenever a PiMS arises or seems imminent (Carhart-Harris et al., 2018c; Hartogsohn, 2016; Hayes, 2019; Johnson et al., 2008; Leary et al., 1963). This said, we recognize that certain emergent PiMSs – such as manic psychotic states – may, however, be particularly difficult to manage via the manipulation of current environmental context alone, particularly during the active PiMS itself. Thus, we do not advocate for environmental contextual management as an exclusive therapeutic strategy and neither do we argue against pharmacological intervention.
The psychopharmacology of PiMSs takes centre stage in this article. We place special focus on the serotonin system and its 2A receptor (5-HT2AR) subtype in particular, which has been shown to be particularly implicated in biology x environment interactions (Chang et al., 2017; Dressler et al., 2016; Fiocco et al., 2007; Jiang et al., 2016; Jokela et al., 2007; Lebe et al., 2013; Mellman et al., 2009; Parade et al., 2017; Salo et al., 2011). Various acute stressors appear to be reliable and robust inducers of serotonin release (Adell et al., 1997; Amat et al., 1998, 2005; Bastani et al., 2017; Beitia et al., 2005; Bland et al., 2003; Cohen et al., 2015; Ferres-Coy et al., 2013; Fuenmayor and Garcia, 1984; Fujino et al., 2002; Gardner et al., 2005; Hale et al., 2011; Higuchi et al., 2019; Ishida et al., 1997; Ishiwata and Greenwood, 2018; Johnson et al., 2005; Keeney et al., 2006; Kelly et al., 2011; Li et al., 2015; Myers and Beleslin, 1971; Nakajima et al., 2009; Neugebauer, 2020; Paul et al., 2011; Rex et al., 2005; Yoshioka et al., 1995) and different types of stressor, such as sleep deprivation (Elmenhorst et al., 2012; Maple et al., 2015; Zhao et al., 2019), hypoxia (Anju et al., 2011), chronic tryptophan depletion (Cahir et al., 2007), inflammation (Zhang et al., 2001), tonic pain (Kupers et al., 2009), repeated forced swim (Takao et al., 1995), repeated shock (Dwivedi et al., 2005), administration of stress hormones (Jitsuiki et al., 2000; Kuroda et al., 1992, 1994), amygdala stimulation (Kalynchuk et al., 2006), time-dependent stress (Harvey et al., 2003), novelty stress (Aloyo and Dave, 2007), maternal separation (Benekareddy et al., 2010, 2011; Godar et al., 2019; Vazquez et al., 2000; although see Ohta et al., 2014), isolation rearing (Preece et al., 2004; Rilke et al., 1998), isolation housing (Günther et al, 2008; Schiller et al., 2003; although see Bibancos et al., 2007; Schiller et al., 2003) and social defeat (Berton et al., 1997; McKittrick et al., 1995; although see Visser et al., 2014) all appear to upregulate 5-HT2AR expression, particularly in the cerebral cortex (Anju et al., 2011; Benekareddy et al., 2010, 2011; Berton et al., 1997; Cahir et al., 2007; Dwivedi et al., 2005; Elmenhorst et al., 2012; Godar et al., 2019; Günther et al, 2008; Harvey et al., 2003; Jitsuiki et al., 2000; Kalynchuk et al., 2006; Kupers et al., 2009; Kuroda et al., 1992, 1994; Maple et al., 2015; McKittrick et al., 1995; Ossowska et al., 2001; Preece et al., 2004; Rilke et al., 1998; Takao et al., 1995; Vazquez et al., 2000; Zhao et al., 2019; see also Table 1), which is well known to be massively expanded in humans.
The 5-HT2AR mediates responses to various types of stress.
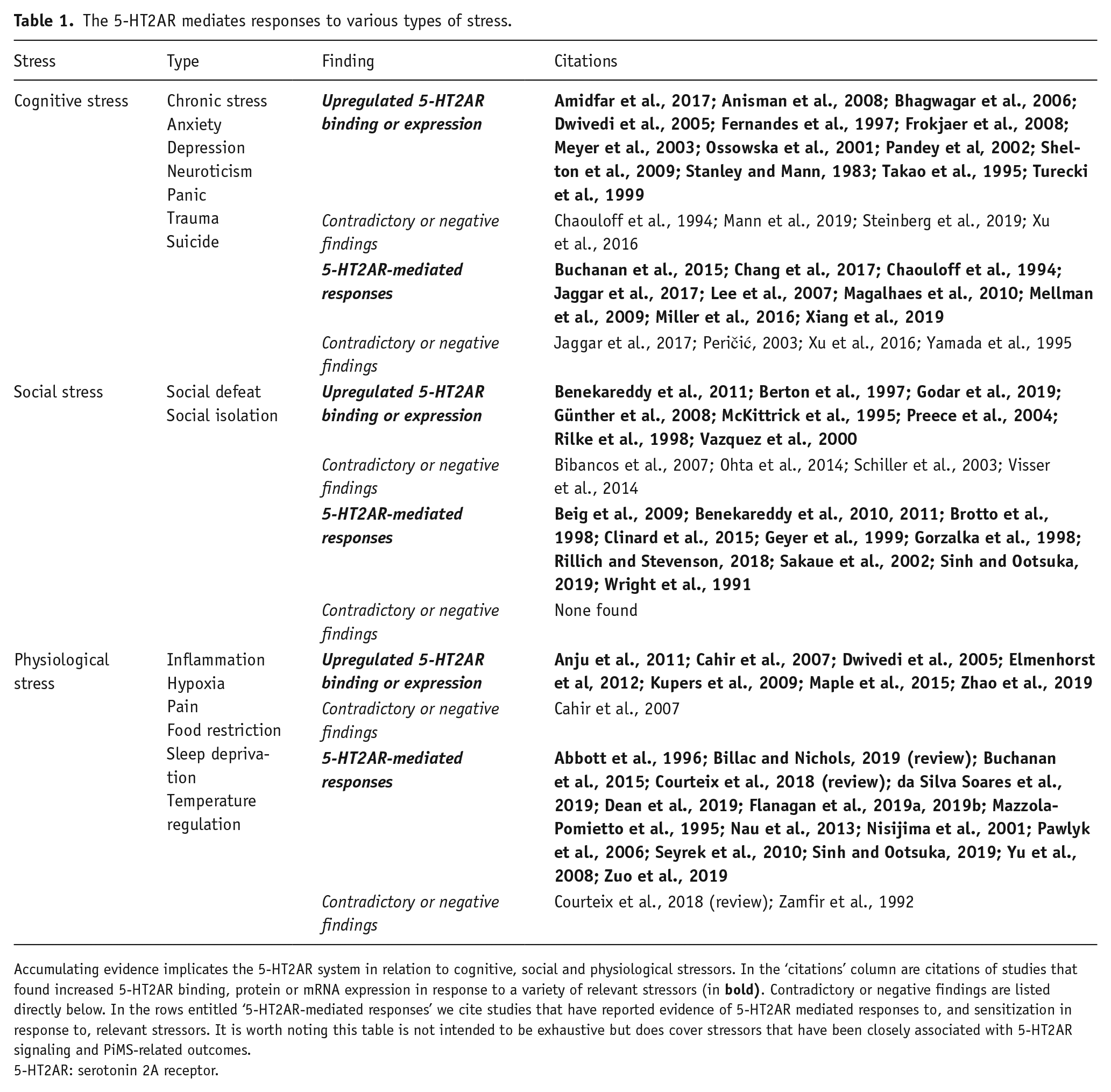
Accumulating evidence implicates the 5-HT2AR system in relation to cognitive, social and physiological stressors. In the ‘citations’ column are citations of studies that found increased 5-HT2AR binding, protein or mRNA expression in response to a variety of relevant stressors (in
5-HT2AR: serotonin 2A receptor.
Social isolation and defeat also reliably sensitize behavioural responses to 5-HT2AR agonists (Benekareddy et al., 2010; Brotto et al., 1998; Gorzalka et al., 1998; Sakaue et al., 2002; Sood et al., 2018; Wright et al., 1991; see also Table 1). The role of stress and the 5-HT2AR system has been the focus of two recent review papers (Carhart-Harris and Nutt, 2017; Murnane, 2019). Supplementing these findings, direct agonism of the 5-HT2AR via psychedelic drugs can sometimes induce psychological states exhibiting phenomena that mimic those seen in extreme stress states, for example, enhanced associative learning and a significant capacity for mediating psychological transformation (Briere et al., 2015; Hefferon et al., 2009; Joëls et al., 2006), as can occur via traumatic encounters. Psychedelics have also been shown to increase the release of stress hormones (Alper, 1990; Calogero et al., 1989; Dos Santos et al., 2012; Hasler et al., 2004; Owens et al., 1991; Schmid et al., 2015; Strajhar et al., 2016; see Schindler et al., 2018 for review).
We highlight how certain natural inducers of stress such as social isolation, starvation, atypical breathing, sleep deprivation, extreme body temperature and pain can be intentionally manipulated for the purposes of personal and/or spiritual development, presumably by promoting endogenous psychedelic-like signaling and we review several cross-cultural, historical and modern examples of such manipulation (Camporesi, 1988; Farré-i-Barril, 2012; Garrett et al., 2011; Grof and Grof, 2010; Janssen et al., 2016; Macmillan, 2013; Naor and Mayseless, 2017).
In keeping with principles of bidirectional translation (Jia, 2016), research with psychedelic (‘mind-manifesting’) drugs can inspire research into the pharmacology and phenomenology of endogenously occurring PiMSs, which, in turn, helps inspire hypotheses on the function of 5-HT2AR signaling, including how it relates to mental illness and its treatment, such as via psychedelic therapy (Carhart-Harris and Goodwin, 2017), plus other methods of manipulating PiMSs for psychotherapeutic ends (Fosha, 2000; Grof and Grof, 2010; Kuijpers et al., 2007). All classic serotonergic psychedelic drugs have direct agonist properties at the 5-HT2AR (Nichols, 2016), and the key role this particular receptor plays in mediating their signature behavioural effects is supported by: (a) affinity-by-potency relationships in animals and humans (Glennon et al., 1984; Rasmussen et al., 1986; Sadzot et al., 1989); (b) a plethora of antagonist pre-treatment studies (e.g. Preller et al., 2017; Quednow et al., 2012; Vollenweider et al., 1998); and (c) evidence of a 5-HT2AR occupancy by subjective effects relationship (Madsen et al., 2019). A wealth of evidence now exists that, via their action at the 5-HT2AR (Madsen et al., 2019), psychedelics, at relevant dose ranges, reliably trigger conditions conducive to psychological transformation (Carhart-Harris and Goodwin, 2017).
Thus, identifying the 5-HT2AR as a key trigger site for inducing PiMSs, we propose that psychedelics hijack the same neurochemical mechanisms that are engaged during, and likely exist for, situations where a hyper-plastic state and associated psychological change is felt as needed. Developing an understanding of endogenously occurring PiMSs can thus shed light on the evolutionary function of brain 5-HT2AR, as well as the action of psychedelics themselves. We propose that the mechanisms underlying PiMSs have evolved to aid rapid and deep learning in situations of perceived or actual existential threat or crisis for the ultimate purpose of catalyzing psychological change when (perceived) circumstances demand this. Somewhat consistent ideas have been expressed in the past (Jaynes, 1976), and more recently in two separate reviews (e.g. Carhart-Harris and Nutt, 2017; Murnane, 2019), but are more fully developed here.
We conclude this article by restating the principle that although PiMSs may be associated with major psychological change, the quality of such change is neither consistent nor pre-determined. We propose that the surrounding context and relational frame in which a given PiMS occurs is a vital determinant of how it manifests (Hayes et al., 2013). Thus, we end this paper by discussing how engineering of optimal contextual frames (as far as this is possible), including not just the containing environment for the PiMS itself but also prior intentions and integration work after the event (Carhart-Harris et al., 2018c; Kornfield, 2001) can enable PiMS-related psychotherapy to be delivered most safely and effectively.
We focus on psychedelic therapy as a prototypical PiMS-focused intervention but there are other relevant examples. Indeed, PiMS-focused therapeutic work can be viewed as a more fundamental therapeutic approach than psychedelic therapy, with the latter merely representing one (particularly potent) example (Carhart-Harris, 2018a). We believe that a growing appreciation of PiMSs could inspire a healthy pivot in mental healthcare and research, more firmly towards the biopsychosocial model (Engel, 1977). Psychedelic therapy, and the PiMS-based model more generally, are quintessentially ‘biopsychosocial’ (Deacon, 2013) as they recognize how social, psychological and biological factors interact in bidirectional, synergistic ways to determine health and illness. The PiMS model purports to explain how biopsychosocial synergies can be harnessed for the delivery of improved mental healthcare. Such improvements are needed if the significant burden of mental illness is to be properly addressed (World Health Organization, 2017). Elevating an awareness of PiMSs as key states of mind and brain, with a heightened potential for mediating lasting change, may have implications for the scientific study of a range of PiMSs-related approaches, such as ‘breathwork’ (Grof and Grof, 2010), meditation (Kuijpers et al., 2007) and accelerated psychotherapies (Fosha, 2000), which should serve to develop their shared validity.
Before beginning our detailed review of relevant literature pertaining to the PiMSs construct, it feels necessary to flag some of its complications early on. For example, it is an implication of the ‘outcome agnosticism’ of the PiMS model that these states be capable of fomenting iatrogenic outcomes, that is, a worsening or triggering of psychopathology (Erritzoe et al., 2017; Strassman, 1984), if the surrounding context is negative, for example, as is often the case in the aetiology of psychosis (Varese et al., 2012b) and perhaps even more plainly, PTSD (Kilpatrick et al., 2003). This matter is highly relevant to inappropriate and/or unsupported psychedelic drug use, as well as malpractice in any kind of paediatric, pastoral or healthcare sector where individuals exhibiting elevated brain plasticity are implicated (Carhart-Harris et al., 2018c; Kornfield, 2001; Schlosser et al., 2019). A wealth of evidence from psychedelic drug, child psychology and mental health research can be cited to highlight the importance of contextual factors, such as: (a) preparedness, (b) intentions and expectations, (c) inter-personal trust/therapeutic alliance, (d) community support and (e) other forms of psychological integration, for safeguarding against harm and enhancing positive therapeutic outcomes (Carhart-Harris et al., 2018c; Kessler et al., 2010).
We also wish to recognize the stabilizing influence that certain implicit assumptions can have on one’s mental state, even if they are symptomatic of mental illness. One should therefore be mindful of potential risks entailed by destabilizing such beliefs, either via the direct action of psychedelic drugs or other means; see Letheby (2016) for a relevant discussion paper. Delusional beliefs are a good example of psychologically stabilizing, but plainly pathological, beliefs. However, we also highlight evidence supporting a role for destabilization as mechanism of therapeutic change (Olthof et al., 2019). Themes of psychological flexibility (Zhang et al., 2018) and experiential acceptance (Rochefort et al., 2018; Watts et al., 2017) are also relevant here.
Relatedly, the position that PiMSs are ripe mediators of psychological transformation, whether towards pathology or away from it, could be viewed as unjustly dismissive or neglectful of the important contribution made by factors such as polygenic predisposition (Davis et al., 2016) and early life adversity (Szyf, 2013; Varese et al., 2012b) in shaping mental health. Such apparent neglect is not intentional, and indeed we consider the highlighted factors part of the overall contextual/relational frame shaping the onset and outcome of a PiMS, where context is a phenomenon that stretches over a broad timescale (Figure 2). Although the recent and current context surrounding a PiMS might be easier to manipulate in favour of positive outcomes, more remote contextual factors such as polygenic predisposition or childhood trauma could still be accounted for, for example, a high polygenic risk for psychotic symptoms might contraindicate psychedelic therapy and thus be used to inform screening for such therapy.
A fork in the road or river analogy is often used to reflect a bifurcation process, namely a ‘cross-roads’, where the trajectory of a system can rapidly destabilise and complexify at a point of bifurcation (Kielhöfer, 2011). In the schematic below (Figure 3), the analogy is used in relation to the PiMS. Contextual factors can be viewed as biasing currents in the river influencing particular outcomes, in the same way that cambers or slopes bias outcomes along a solid path. Briefly, bifurcation theory describes the occurrence of sudden changes in the trajectory of a system. Typically, energy introduced into a system causes the destabilization of a previously dominant trajectory or state (e.g. represented by the single path that precedes the fork). The critical destabilization creates new potential states or trajectories that the system can enter or follow. For simplicity, ‘wellness’ and ‘illness’ are presented here as discrete binary states but we recognize that this is an oversimplification and that mixed features are also possible, for example symptoms consistent with post-traumatic stress and growth can co-exist or ‘flip-flop’ after a traumatic episode, and similarly, mixed euphoric and dysphoric states can occur in manic psychoses. The unifying principle, however, is that change, whatever its nature, is more likely after such pivotal events.

Use of the ‘fork in the river analogy’ to communicate the nature and effect of pivotal mental states (PiMSs). The boat, named ‘5-HT2A’, represents the molecular gateway leading to the occurrence of a PiMS, that is, entry into a hyper-plastic state in which the likelihood of major psychological change is enhanced, and the nature of that change is especially context dependent. The fork in the road reflects a bifurcation and thus can be expected to fall under the principles of bifurcation theory more generally (Kielhöfer, 2011); the details of this are beyond the scope of this article.
Pivotal mental states and their divergent outcomes
Throughout this paper we argue that PiMS can mediate divergent outcomes that strongly depend on the surrounding context in which they arise. Here we apply this principle to a classic controversy in psychology and psychiatry: namely, the relationship between spiritual experiences and psychosis. It has long been noted that certain spiritual and early and acute psychotic experiences exhibit similar features such as anomalous self-experience, magical thinking and perceptual aberrations (Baldacchino, 2016; Buckley, 1981; Cangas et al., 2008; Crespi et al., 2019; Grof and Grof, 1989; Hunt, 2000, 2007; Jackson, 1997; Jaynes, 1976; Luhrmann, 2017; Lukoff, 1985, 2007, 2018; Murray et al., 2012; Parnas and Henriksen, 2016; Perry, 1977; Polimeni, 2018; Powers and Corlett, 2018; Ross and McKay, 2018; Willard and Norenzayan, 2017). All three phenomena are reliably induced by 5-HT2AR agonist psychedelics (Carhart-Harris, 2007; Carhart-Harris and Friston, 2010; Kraehenmann et al., 2017; Letheby, 2016; Millière, 2017; Nour et al., 2016), thus implying their relationship to a more fundamental state – the 5-HT2ARR-mediated PiMS.
According to the model presented here, the framing of psychosis as pathological and spiritual experience as psychologically beneficial, obscures their common relationship to the PiMS (Jackson, 1997). Indeed, a negative or positive outcome is the paramount criterion for distinguishing between mystical and psychotic experience, respectively (DeHoff, 2015; James, 2003 [1902]). Moreover, scales used to measure trait schizotypy and spirituality also contain items relating to social and emotional adjustment, behaviour and appearance (Davidson et al., 2016; Garssen et al., 2016; Raine, 1991; Raine et al., 1994), which capture divergent responses to ego-disturbance, perceptual aberration and magical thinking (Crespi et al., 2019). It is our view that certain psychoses and spiritual or religious conversions share a common heritage in the PiMS. We also argue that this connection is often overlooked because of a tendency to selectively focus on outcomes rather than the processes that lead up to them (Figure 4). The mediational states themselves are often relatively brief and intense, but they can also be more protracted. We describe them as ‘pivotal’ based on the principle that a consistent root state can mediate strongly divergent outcomes, such as spiritual or religious epiphany or conversion versus the acquisition of a psychotic delusion. The Oxford definition of a pivot (as a noun) is a central point or person from which a mechanism turns or oscillates (Oxford English Dictionary, pivot, def. 1., n.d.) and the adjective ‘pivotal’ is defined as ‘[a thing] of crucial importance in relation to the development or success of something else’. We therefore use the adjective here in reference to the principal properties and function of PiMSs, that is, a heightened ability to mediate major psychological change. We propose that a focus on the root state preceding a given outcome should reveal the presence of the defining properties of PiMSs.

How an outcome-biased or outcome-focused perspective can obscure the role of a common root state preceding a bifurcation that led to the outcome. Although a particularly stark outcome may manifest (e.g. the development of a psychotic episode or disorder), the implication is that this may not have been an inevitability and measures can be taken to either shepherd the pivotal mental state (PiMS) in a particular direction or even seek to avoid it altogether, for example, in cases where there are polygenic vulnerabilities.
In this paper, we use a recent definition of stress as ‘the body’s multi-system response to any challenge that overwhelms, or is judged likely to overwhelm, selective homeostatic response mechanisms’ (Day, 2005). The cited paper is a useful reference for reviewing alternative definitions. Discussing different definitions of stress is beyond the remit of the present paper but it is relevant to note that most definitions appear to agree that stress is a multi-level response to an apparent threat to an organism’s present state. This consistent definition of stress is useful, as it acknowledges how stress can engage adaptive mechanisms – such as heightened plasticity – that is, the quality of being more shapeable (Oxford English Dictionary, plasticity entry, n.d.). Appreciating the link between stress and adaptability can help us understand the etiological and evolutionary function of PiMSs. As we show below, stress is a particularly robust and reliable primer, trigger and potentiator of PiMSs (van der Steen et al., 2017).
Stress is typically construed of and felt as an aversive phenomenon and recognizing this can help us understand why naturally arising PiMSs often lead to negative outcomes (Figure 5). For example, if a PiMS is not expected and the experience is protracted, then the net effect of this is likely to be distressing (Rekhi et al., 2019; van der Steen et al., 2017). Adverse conditions are, almost by definition, stressful and classic examples such as unintentional social isolation, urban-environment-related stress, poor socioeconomic status and childhood adversity are likely to mediate and/or potentiate PiMS, and have all been significantly linked with psychotic disorders (Selten and Cantor-Graae, 2005, 2007; Selten et al., 2013, 2016). In contrast, spiritual experiences appear to be more likely to arise in positive environmental contexts, such as in nature (Anderson et al., 2018).
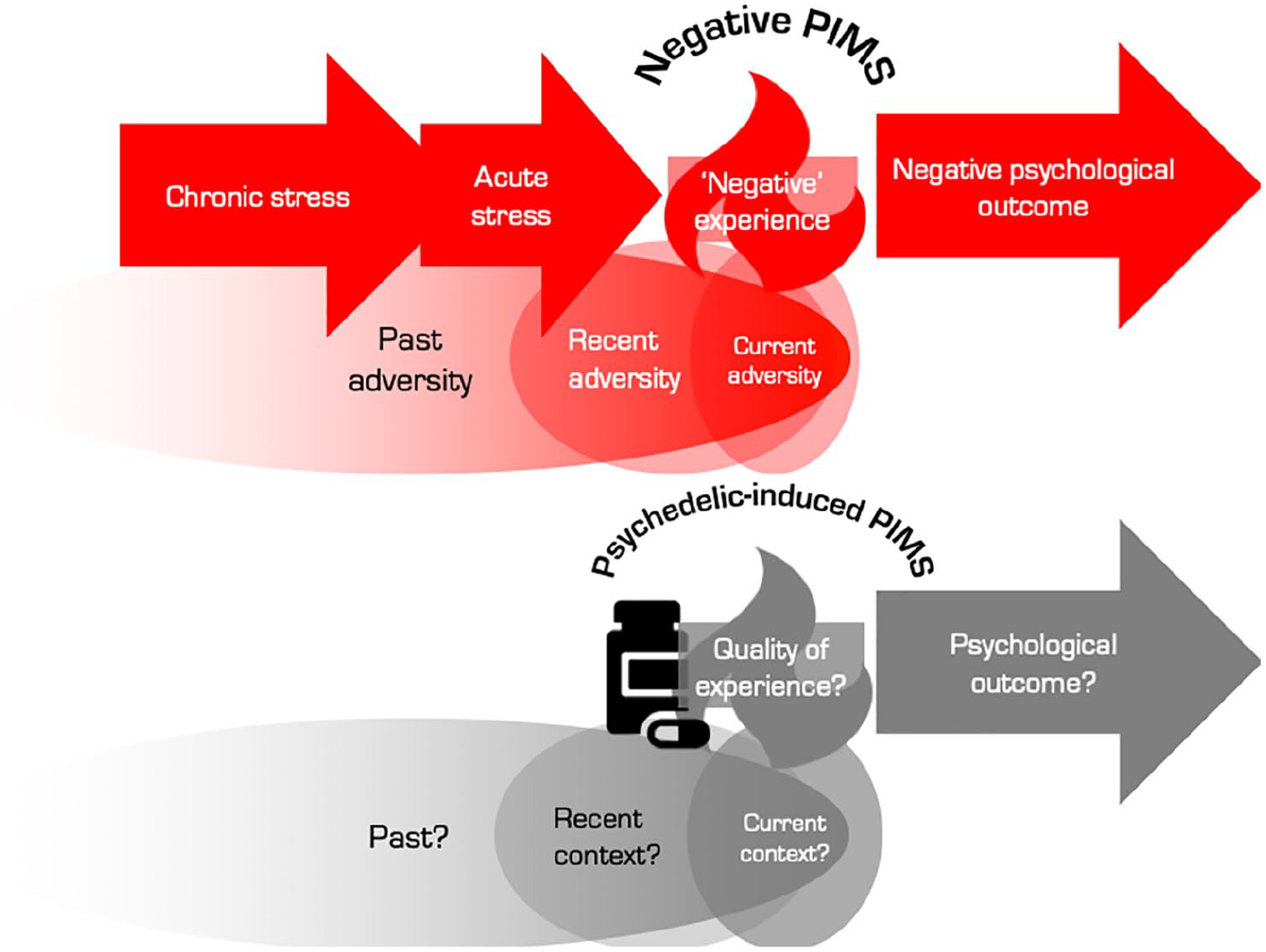
The upper process is perhaps the most typical scenario for a naturally occurring pivotal mental state (PiMS) but also the worst possible scenario as far as long-term mental health is concerned. It is the scenario that, we believe, most often accompanies the development of a psychosis. The lower process can be described as ‘outcome agnostic’ in the sense that the quality of the PiMS itself and its subsequent impact on mental health is contingent on the surrounding context and relational frame into which is arises. If the recent, current and post-PiMS context can be shaped favourably, then one can expect the longer-term impact of the PiMS to be favourable also. However, more remote contextual factors (e.g. polygenic make-up or a history of trauma) are, understandably, harder to manipulate and may, on occasion, merely signal that a PiMS could be especially risky, for example, in terms of its outcomes. This said, it is entirely plausible that naturally occurring PiMS can yield favourable psychological outcomes.
The bidirectional and often mutually reinforcing relationship between belief and affect is reflected in emotionally antithetical experiences of self-dissolution in spiritual versus psychotic experiences. In severe and enduring psychosis, self-fragmentation (Millière, 2017; Parnas and Henriksen, 2016) is often felt as invasive and torturous, whereas sensations of ‘mystical union’ or ‘inter-connectedness’ (Carhart-Harris et al., 2018b) appear to lie at the heart of positive experiences of ego-dissolution in naturally occurring (Stace, 1960) and psychedelic-induced spiritual experiences (Millière, 2017; Nour et al., 2016; Roseman et al., 2018). See Parnas and Henrikson (2016) for another detailed comparison of mystical and psychotic experience. We are mindful, however, that manic episodes can also have a euphoric quality and thus, euphoric states are not therapeutic themselves (Masters, 2010).
Increased emotional tone is a common feature of PiMSs that likely plays an important role in modulating their immediate nature and longer-term impact. Recent work with psychedelics has revealed that ‘emotional breakthrough’ is an important and distinct mediator of long-term positive outcomes after psychedelic therapy (Roseman et al., 2019). Relatedly, feelings of ‘awe’ in nature have been found to mediate improvements in wellbeing via nature exposure (Anderson et al., 2018). The active suppression of emotion is associated with the nature and severity of symptomatology in psychosis and PTSD (Laloyaux et al., 2016; Roemer et al., 2001; Tull et al., 2018) and the duration of negative affect (but not its intensity) has been found to predict negative long-term outcomes after psychedelic drug use (Carbonaro et al., 2016).
A logical explanation for the impact of intense emotion on long-term psychological outcomes from PiMSs is that the felt emotion modulates the nature of associations made during the pivotal state and intensifies their influence, allowing for beliefs and perspectives to be affected. In predictive processing terms, emotion can be thought of as prediction error modulating the ‘precision’ (inverse variance) of ‘posteriors’ or ‘priors’ (probability distributions in the Bayesian sense, i.e. the expected likelihood of a set of possibilities). Expressed in a way to serve readers’ intuition: at a high hierarchical level, the precision of a prior relates to one’s confidence in a given perspective or belief. If the affective tone of a PiMS is positive and intense, this may drive a ‘de-weighting’ or relaxation of a negatively held belief. For example, a negative (cognitive) bias characterizing a depressive disorder (Disner et al., 2011) would be felt less confidently if it were relaxed. However, if the affective tone accompanying a PiMS is intense, negative and unresolved, this may contribute to an instilling and/or reinforcement of negative beliefs – it would add precision to the (negative) belief (Roseman et al., 2019). See Carhart-Harris (2019) for a discussion of related themes.
Increased emotional tone coupled with hyper-plasticity and enhanced associative learning is a potent mix for moderating or reinforcing deeply held beliefs and perspectives. Symbolic associations may become increasingly oriented to one’s underlying emotional state, encouraging the formation of ‘affect-laden’ worldviews. Indeed, if emotion is intensified during a PiMS but its valence is unpredictable, it may explain how a consistent root state can mediate extremely divergent outcomes. The intensity, duration, psychological preparedness and degree of psychological resolution may all contribute to the nature and influence of emotion on the quality and impact of a PiMS (Haijen et al., 2018). Negative affect predominates in schizophrenic psychosis and is associated with delusions of persecution (Paolini et al., 2016) and general cognitive disorganization (Carrigan and Barkus, 2017). Conversely, positive affect predominates in western conceptions of spiritual experience (DeHoff, 2015; Hardy, 1979) as well as psychedelic drug experiences (Liechti et al., 2017), all of which usually feature some degree of positive context.
Manic states could be seen as a challenge to this rule, however. Relatedly, we recognise that persecutory and grandiose thinking and associated negative and positive mood states are not mutually exclusive and can exist in parallel and/or interchangeably, as exemplified by apocalyptic and religious delusions (Iyassu et al., 2014; Wessinger, 2000) and manic psychoses (Picardi et al., 2018). Perceptions of interconnectedness and self-transcendence in manic states bear resemblance to the phenomenological qualities of spiritual states, including those induced by psychedelics (e.g. see Carhart-Harris et al., 2013). Manic states are often triggered by stressful events and culminate in chronic psychotic disorders. Comorbidity of bipolar, schizophrenia and schizoaffective disorders reflects a shared polygenic vulnerability to psychotic symptoms (Cardno and Owen, 2014; Craddock et al., 2009; Laursen et al., 2009) and possibly, by extension, PiMSs. Context is rarely, if ever, either entirely ‘positive’ or ‘negative’ and the same is true for how it is received, that is, context is inherently relational and subjective. Moreover, we recognize the paradox that highly positively valanced mood states, for example euphoric states in manic psychoses or a psychedelic drug experience, do not naturally imply positive long-term outcomes for mental health. The phenomenon of ‘spiritual bypassing’ is relevant in this regard (Masters, 2010). For example, the zealous promotion of self/ego transcendence in the absence of subsequent psychological integration could be regarded as a (subtly) negative, or at least ‘imperfect’, context (Kornfield, 2001).
Psychotic episodes and spiritual experiences are both commonly preceded by a dissatisfaction with reality and one’s place within it (Hardy, 1979; James, 2003 [1902]; Miller and C’de Baca, 2001). In the prodrome to psychosis (i.e. a pre-psychotic period that precedes a conspicuous psychotic episode) an individual’s presentation may often resemble that of a depressive phenotype (Corcoran et al., 2011; DeVylder et al., 2014; Rietdijk et al., 2013). Indeed, an individual entering a PiMS (whether interpreted as psychotic, spiritual, both or neither) may perceive their world as unreal, thus spurring belief in a ‘meaningful’ but concealed other reality (James, 2003 [1902]: 149–155). One should be mindful how transition into a psychotic episode or spiritual experience can be construed as a ‘manic defence’ (Winnicott, 1935), that is, an escapist ‘flight from reality’. See again Masters (2010) on the topic of ‘spiritual bypassing’.
In longer prodromal states, a loss of interest in key activities and pursuits (e.g. education and vocation) and pleasures (e.g. food, sex, social interactions) can coexist with an emerging interest in supernatural paranormal, religious and ritual domains (Møller and Husby, 2000, 2003). Thoughts may begin to feel unrelated to the self and emotionally distant (Parnas and Handest, 2003). Individuals may find themselves examining their thoughts and behaviours from a third-person perspective (Nelson et al., 2016), as aware yet dissociated subjects (Parnas and Handest, 2003). A dissociation of subjective awareness from one’s body and/or thoughts evokes common religious/spiritual notions of the incorporeal soul, ‘true-self’ (Ātman) and ‘no-self’ (Anātman) (Parnas and Henriksen, 2016) as well as the related notions of ‘non-dual awareness’ (Josipovic, 2014) and the ‘unitive experience’ (Stace, 1960).
Many religious traditions place value on understanding the nature of the self, its relationship to the world and its transformation. One potentially relevant theme here is ‘salvation’, which, outside of theological contexts, has been defined as a ‘preservation from destruction or failure’ and ‘deliverance from danger or difficulty’ (Merriam-Webster, salvation entry, def. 3., n.d.). Thus, the ‘salvific’ and/or adaptive function of extraordinary personal experiences has historically been framed in a religious way and religion may function to ease human suffering via other plausible mechanisms. For example, rituals may mitigate anxiety and facilitate healing (Csordas and Lewton et al., 1998; Lang et al., 2015) and absolutist religious beliefs may protect against a basal existential uncertainty. Thus, religion and non-denominational spirituality (whether secular or otherwise) offer potentially useful frameworks/belief systems from which to positively frame experiences that in other contexts might be readily construed as pathological.
Serotonin, stress and the 5-HT2AR
Serotonin, coping and adaptation
Serotonin is an endogenous monoamine found throughout the body, particularly in the gastrointestinal system (Gershon, 2013), lungs (Castro et al., 2017) and, to a lesser extent, the central nervous system. Despite its more modest prevalence in the brain, serotonin neurotransmission is known to play an important modulatory role in several key aspects of mind and behaviour, including brain development (Azmitia, 2001), mood (Garcia-Garcia et al., 2017), cognition (Meneses, 1999) and sleep (Jouvet, 1999). Serotonin is a particularly complex neuromodulator, with a broad range of receptor subtypes (i.e. at least 14 different subtypes have been identified to date (Hoyer, 2019), some of which have opposing functions on activation (Andrade, 2011; Araneda and Andrade, 1991)). Previous attempts at a unifying model of the function of brain serotonin have tended to focus on its role in moderating anxiety states (Charney et al., 1987; Deakin and Graeff, 1991; Piszczek et al., 2015) as well as impulsivity/impatience and aggression (Brown and Linnoila, 1990; Fairbanks et al., 2001; Fonseca et al., 2015; McDannald, 2015; Miyazaki et al., 2012, 2014, 2018; Ranade et al., 2014). The most reliable inducers of 5-HT release appear to be stress (Fujino et al., 2002), pain (Harvey et al., 1975) and uncertainty (Miyazaki et al., 2018), see Table 1. Thus, it has been proposed that serotonin’s ‘serenic’ effects (Olivier and Mos, 1990), particularly via non-5-HT2AR (i.e. most notably postsynaptic 5-HT1A receptors in stress circuitry), may be a perceived as an adaptive response to adversity, for example, aiding a type of resilience one might call ‘fortitude’, ‘passive coping’ or an enhanced ability to endure adversity and thus, ‘get by’ (Carhart-Harris and Nutt, 2017).
However, meeting stress with an intention to merely endure may not be an optimal long-term strategy. For example, efforts to suppress and thereby avoid, stress may not be conducive to the revision of (potentially problematic) internal models, such as those linked to cognitive biases in depression, for example. Thus, it seems reasonable to ask: does there not exist an alternative adaptive mechanism, sufficiently different to the stress avoidance/mitigation strategy just described (Carhart-Harris and Nutt, 2017; Wallace, 1956), perhaps one that becomes triggered when adverse conditions surpass a critical threshold of severity and/or chronicity (Dwivedi et al., 2005) such that mere endurance is not enough?
It has been proposed before that a principal function of the serotonin 2A receptor subtype is to induce a state of cortical hyper-plasticity conducive to major adaptive change (Carhart-Harris and Nutt, 2017). The present paper extends this previous work to highlight how chronic stress primes the serotonin (2A receptor) system for the elicitation of a PiMS: a hyper-plastic state in which prior assumptions are relaxed, enabling an enhanced sensitivity to potential new information, consistent with rapid and deep learning. In psychosis, this process may result in the maladaptive formation of delusional beliefs that nevertheless help make sense of a frightening world. In spiritual experiences, individuals may report sudden moments of clarity and insight (e.g. epiphanies) servicing positive self-development and renewed perspective (C’De Baca and Wilbourne, 2004; Miller and C’de Baca, 2001). Highly consistent themes can be found in reports of post-traumatic growth after recovery from psychosis and other severe conditions (Slade et al., 2019), near-death experiences (Geiger, 2009; Khanna and Greyson, 2015) and in cases of clinical breakthrough with psychedelic therapy (Roseman et al., 2019; Watts et al., 2017). See also Miller and C’de Baca (2001).
Addressing the following questions can help us develop our hypotheses on the role of stress and the 5-HT2AR in PiMSs: 1. Are certain stressors linked with the occurrence of states meeting the definition of a PiMS (e.g. incipient psychosis or spiritual experience)? 2. What types of stress and stressors appear to upregulate the 5-HT2AR system most robustly and reliably? 3. Does increased 5-HT2AR activity facilitate psychotic or spiritual states or traits? 4. Are 5-HT2AR-induced PiMSs associated with major psychological change or transformation? These questions will be addressed in the following sections, starting with an examination of various stressors that facilitate self-transformation and upregulate 5-HT2AR signaling.
Cognitive stress and the 5-HT2AR
A perceived lack or loss of control is a well-known cause of stress (Steptoe and Poole, 2016) and a potential transdiagnostic factor in a variety of mental illnesses (McEvoy and Mahoney, 2011). Perceived uncontrollability of thoughts correlates with intensification of pathology in obsessive compulsive disorder (OCD) and schizophrenia (García-Montes et al., 2006). In general, negative thoughts are more likely to be misattributed to sources other than the self, reflecting less felt ownership (Swiney and Sousa, 2013). Abnormal meta-cognition (‘thinking about thinking’) is a characteristic of psychosis (Sellers et al., 2017) and schizotypal individuals are also significantly more likely to show abnormal metacognition (Chan et al., 2015). The schizotypal mind may relieve (or reward) itself by hypothetically solving matters of uncertainty via escape into fantasy or delusion, akin to the relief served by compulsive rituals in OCD. A similar function may be served by ritual in religion (Lang et al., 2015). Indeed, the over-weighting of priors (excessive ‘precision’, or inverse variance, synonymous with excessive confidence), particularly in response to perceived uncertainty and loss of control, may be a transdiagnostic feature of psychological suffering (Boswell et al., 2013; Carhart-Harris, 2019; McEvoy and Mahoney, 2011), namely the proposed (defensive) function of many symptoms of psychopathology may be to mitigate uncertainty through the over-weighting of beliefs and/or excessive reinforcement of certain specific behaviours (Boswell et al., 2013; Carhart-Harris, 2019; Carhart-Harris and Friston, 2019).
Neuroticism and depression, indicative of chronic cognitive stress, regularly precede and coexist with psychotic disorders (Corcoran et al., 2011; DeVylder et al., 2014; Lönnqvist et al., 2009; Rietdijk et al., 2013; Van Os and Jones, 2001) and depression and despair also often precede religious experiences (Hardy, 1979). Most studies suggest that chronic stress upregulates 5-HT2AR binding and expression (Dwivedi et al., 2005; Fernandes et al., 1997; Ossowska et al., 2001; Takao et al., 1995; although see Xu et al., 2016) and the 5-HT2AR is implicated in physiological and behavioural responses to chronic stress in humans (Chang et al., 2017; Fiocco et al., 2007; Parade et al., 2017) and animal models (Jaggar et al., 2017; Xu et al., 2016; although see Jaggar et al., 2017). Various studies show upregulated 5-HT2AR transcript and protein expression in neuroticism (Frokjaer et al., 2008) and depression (Amidfar et al., 2017; Bhagwagar et al., 2006; Meyer et al., 2003; Shelton et al., 2009; although see Steinberg et al., 2019), particularly in relationship to dysfunctional attitudes (Baeken et al., 2014; Meyer et al., 2003) and suicide (Anisman et al., 2008; Pandey et al., 2002; Stanley and Mann, 1983; Turecki et al., 1999; although see Mann et al., 2019; Steinberg et al., 2019).
Given the close association between depression and PiMS-related outcomes (i.e. spiritual and psychotic experiences), it is natural to surmise that cognitive-stress-induced upregulation of 5-HT2AR expression may be an important biology x environment interaction through which both spiritual and psychotic experiences manifest via a subsequent increase in 5-HT2AR activation.
Consistent with the PiMS model, baseline neuroticism is associated with elevated cortical 5-HT2AR expression (Frokjaer et al., 2008), predicts thought disturbance, blackout and challenging experiences under psychedelics (Barrett et al., 2016, 2017; Lienert and Netter, 1996; although see Studerus et al., 2012) and yet may be reduced after psychedelic therapy (Erritzoe et al., 2018), presumably because of positive contextual manipulation during the acute hyper-plastic state, as well as afterwards. In relation to naturally occurring PiMSs, both uncontrollable stress (Amat et al., 2005; Bland et al., 2003) and punishment (Cohen et al., 2015; Faulkner and Deakin, 2014) are associated with 5-HT release (Amat et al., 2005; Bland et al., 2003; Cohen et al., 2015; Faulkner and Deakin, 2014).
Psychological trauma predisposes certain individuals towards dissociation, hallucination and other psychotic-like features (Kilcommons and Morrison, 2005; McCarthy-Jones and Longden, 2015; Varese et al., 2012a). There exists an especially high comorbidity between psychotic and PTSD symptoms (Grubaugh et al., 2011; Ng et al., 2016). The 5-HT2AR may mediate altered mind and brain functioning in relation to traumatic events. Consistently, 5-HT2AR variants have been associated with PTSD (Lee et al., 2007; Mellman et al., 2009), symptom severity in PTSD, and the degree of default mode network connectivity amongst people with PTSD (Miller et al., 2016), a network implicated in the action of psychedelics (Carhart-Harris et al., 2012a) that is rich in 5-HT2AR (Beliveau et al., 2017). Animal models implicate the 5-HT2AR in anxiety (Chaouloff et al., 1994; Magalhaes et al., 2010) and anxiety following exposure to trauma (Xiang et al., 2019). Indeed, traumatic experiences are likely to meet all three of the key criteria for a PiMS.
Social stress and the 5-HT2AR
Social stress and other relevant factors such as urban stress, ethnic minority status (Kirkbride et al., 2012; Veling et al., 2008), migration, childhood trauma, poor cognitive aptitude and drug abuse all contribute to a sense of social defeat and have been linked with schizophrenia (Selten and Cantor-Graae, 2005, 2007; Selten et al., 2013, 2016; Zammit et al., 2010). Social defeat is a phenomenon relative to one’s immediate social surroundings and expectations for the future (Jones et al., 1993; Reininghaus et al., 2008), that is, one’s specific relational frame (Hayes et al., 2013). Social stress has also been linked with higher rates of religious engagement (Aydin et al., 2010; Friedman and Saroglou, 2010; Ghorpade et al., 2006). Loss of close social connections and social isolation have been linked with cognitive-perceptual abnormalities such as a hallucinated sensed presence (Geiger, 2009; Steffen and Coyle, 2010), and loneliness has also been linked with facets of magical thinking such as anthropomorphization (Bartz et al., 2016; Epley et al., 2008; Eyssel and Reich, 2013), which is a feature of religious belief (Guthrie and Guthrie, 1995). Perceived social disconnection, social isolation and social withdrawal regularly precede psychotic and spiritual experiences (Mishlove and Chapman, 1985; Seeman, 2017) and solitary confinement, conceived initially as a method of spiritual rehabilitation, often leads to perceptual distortions, hallucinations, cognitive deficits and paranoia (Haney, 2018). It is also worth highlighting that the classic 5-HT2AR agonist psychedelics quite reliably induce experiences of vivid sensed presence (e.g. Timmermann et al., 2019) as well as magical thinking more broadly (Carhart-Harris et al., 2014a; Kraehenmann et al., 2017).
In preclinical modelling studies, 5-HT2AR antagonism impairs acquisition of conditioned defeat (Clinard et al., 2015; Rillich and Stevenson, 2018) and suppresses hyperthermic response to social defeat (Beig et al., 2009; Sinh and Ootsuka, 2019), whereas agonist administration into the basolateral amygdala increases acquisition of conditioned defeat (Clinard et al., 2015). Acute social defeat increases 5-HT levels (Beitia et al., 2005; Gardner et al., 2005; Higuchi et al., 2019; Keeney et al., 2006; Paul et al., 2011), and region-specific increases in 5-HT2AR expression have been found in chronically subordinate animals (Berton et al., 1997; McKittrick et al., 1995). Acute social defeat has not been shown to upregulate 5-HT2AR expression (Visser et al., 2014) or produce phenotypes indicative of 5-HT2AR sensitization, such as that seen after recurring defeat (Hayashida et al., 2010). These observations are consistent with the present model, which acknowledges that the positive relationship between various stressors and 5-HT2AR upregulation is dependent on the chronicity and severity of that stress.
Maternal separation stress in rodents potentiates the effects of 5-HT2AR agonists (Benekareddy et al., 2010; Sood et al., 2018), and 5-HT2AR antagonists reduce maternal separation-induced anxiety (Benekareddy et al., 2011), aggression and bradycardia (Godar et al., 2019). Preliminary evidence also suggests that maternal separation upregulates 5-HT2ARs (Godar et al., 2019) and 5-HT2AR mRNA expression (Benekareddy et al., 2011; Vazquez et al., 2000; although see Ohta et al., 2014). Isolation rearing likewise seems to upregulate 5-HT2AR expression (Preece et al., 2004; Rilke et al., 1998) and potentiate the effects of 5-HT2AR agonists (Wright et al., 1991). Pre-pulse inhibition deficits displayed by isolation-reared animals are reduced by 5-HT2AR antagonism (Geyer et al., 1999)
The effects of isolation housing on 5-HT2AR expression is inconclusive and time variable, e.g., one study found that 5-HT2AR expression is decreased at 4 weeks (Schiller et al., 2003) whereas others found increases at 6 and 12 weeks (Günther et al., 2008; Schiller et al., 2003), while another found decreased 5-HT2AR mRNA expression at 6 weeks (Bibancos et al., 2007). More evidence is needed to clarify these time-dependent relationships and how they relate to 5-HT2AR mRNA versus protein expression. The effects of isolation on 5-HT2AR sensitization appears more straightforward, with isolation housing in mature animals potentiating 5-HT2AR agonist-induced wet dog shakes (Brotto et al., 1998) and head twitch (Sakaue et al., 2002).
Physiological stress and the 5-HT2AR
Chronic inflammation, excitotoxicity, hypoxia, metabolic dysfunction, starvation, sleep deprivation and pain are all physiological processes linked to stress. Brain inflammation and excitotoxicity are putative risk factors for psychosis and likely contribute to neurodevelopmental abnormalities (e.g. reduced grey matter volume) in schizophrenia (Plitman et al., 2016; Watkins and Andrews, 2016; Zhang et al., 2016b). Various studies show that 5-HT2AR activation has neuroprotective and anti-inflammatory effects (Billac and Nichols, 2019; Fanibunda et al., 2019; Flanagan et al., 2019a, 2019b; Flanagan and Nichols, 2018; Nau et al., 2013; Yu et al., 2008) and inflammation is a common feature of stress-related disorders (Liu et al., 2017) including psychosis (Fraguas et al., 2019). The general neuroprotective and anti-inflammatory functions of the 5-HT2AR may explain why this receptor is upregulated by and mediates responses to so many stressors (Figure 3) and lends additional support to its candidacy as a therapeutic target (Flanagan and Nichols, 2018).
Hypoxia (oxygen deficiency) may be particularly relevant, as respiratory complications are associated with increased rates of psychosis (Kalucy et al., 2013; Partti et al., 2015). Hypoxia is also implicated in near-death experiences (NDEs) (Klemenc-Ketis et al., 2010; Van Lommel et al., 2001), ‘runners’ high’, high-altitude-induced psychosis (Hüfner et al., 2018), panic attacks (Klein, 1993; Roth et al., 2002) and spiritual practices and experiences (Nivethitha et al., 2017). Hypercapnia (increased CO2 in the bloodstream) may mediate the psychedelic-like effects of naturally occurring hypoxic conditions (Klemenc-Ketis et al., 2010). Indeed, there is a history of CO2 ‘carbogen’ therapy (Meduna, 1950) that overlaps in some regards with psychedelic therapy, such as with an emphasis on the induction of cathartic experiences.
Hypoxia upregulates cortical 5-HT2ARs (Anju et al., 2011) and the 5-HT2AR agonist psychedelic N,N-Dimethyltryptamine (DMT) exerts neuroprotective effects in cells exposed to hypoxic conditions, leading scholars to suggest endogenous DMT may be released as an adaptive response to physiological stress (da Silva Soares et al., 2019; Szabo et al., 2016). A recent study observed increased DMT levels in rat visual cortex following cardiac arrest (Dean et al., 2019), a finding that supports the hypothesized role of endogenous DMT in NDEs (Strassman, 2000; Timmermann et al., 2018). The specificity of this release needs to be considered, however, given that concentrations of 5-HT and other neurotransmitters are also massively increased during asphyxiation, cardiac arrest and hypercapnia and may compete with DMT at the 5-HT2AR (Johnson et al., 2005; Li et al., 2015; Nichols and Nichols, 2019). Stimulation of the 5-HT2AR may also precipitate psychotic symptoms during panic attacks (Galynker et al., 1996; Goodwin and Davidson, 2002; Masdrakis et al., 2017), which often involve a physiological disturbance (e.g. hypercapnia) coupled with cognitive misinterpretation (Clark, 1986; Vollmer et al., 2015). Preliminary evidence suggests the 5-HT2AR mediates CO2-induced arousal (Buchanan et al., 2015; Smith et al., 2018), a popular model of panic attack, as well as anxiety responses to corticotrophin-releasing factor receptor 1 stimulation (Magalhaes et al., 2010). That CO2 inhalation can induce panic or pleasant psychedelic-like experiences (Meduna, 1950), with activation of 5-HT2ARs likely playing a role, is supportive of the PiMS model, including its emphasis on the context dependency of outcomes.
Metabolic dysfunction and starvation are associated with psychotic phenotypes (Morylowska-Topolska et al., 2017; Pillinger et al., 2017; Prabakaran et al., 2004). Chronic tryptophan depletion (3 weeks) selectively increases cortical 5-HT2AR binding (Cahir et al., 2007). Fasting also increases serotonin-induced intracellular calcium cation concentration, a proposed correlate of 5-HT2AR function in the brain (Sudo et al., 1997), as stimulation of 5-HT2ARs induces intracellular calcium release (Raote et al., 2007). Increases in cortical brain-derived neurotrophic factor (BDNF), as well as the antidepressant and anti-inflammatory effects of fasting (Cui et al., 2018; Fond et al., 2013), are consistent with the effects of 5-HT2AR activation (Carhart-Harris et al., 2016a; Cui et al., 2018; Flanagan et al., 2019b; Jaggar and Vaidya, 2018). Acute fasting and intermittent religious fasting have been shown to increase 5-HT levels and metabolism (Bastani et al., 2017; Fuenmayor and Garcia, 1984; Ishida et al., 1997; although see Bubenik et al., 1992) whereas longer-term tryptophan depletion or starvation may decrease 5-HT levels (Cahir et al., 2007; Haider and Haleem, 2000). Agonists and antagonists of the 5-HT2AR reliably decrease and increase feeding, respectively (see Gorwood et al., 2018 for review).
There is some evidence of increased 5-HT2AR expression in overweight individuals (Erritzoe et al., 2008; although see Chaouloff et al., 1995) and reduced 5-HT2AR expression in anorexia nervosa (AN) (Audenaert et al., 2003; Bailer et al., 2004; Frank et al., 2002; Kaye et al., 2013; although see Bailer et al., 2007). That patients recovered from AN continue to display reduced 5-HT2AR expression (Kaye et al., 2013) might be interpreted as suggesting that decreased 5-HT2AR expression is an inherited trait marker of AN, but we suggest a compensatory long-term downregulation of 5-HT2ARs in AN could also occur in response to chronic overactivation of 5-HT2ARs associated with restricted feeding. Enduring but state-specific decreases in 5-HT2AR expression, as well as epigenetic modifications of the 5-HT2AR, have been observed in schizophrenia (Abdolmaleky et al., 2011; Cheah et al., 2017; Rasmussen et al., 2016); see below.
Sleep deprivation can serve as a model of psychosis and the relationship between sleep deprivation/disorders and psychosis is well established (Meyhöfer et al., 2017; Reeve et al., 2018; Waters et al., 2018). Sleep deprivation is associated with a rapid upregulation of 5-HT2AR expression in rodents (Maple et al., 2015; Zhao et al., 2019) and increased cortical 5-HT2AR binding in humans (Elmenhorst et al., 2012). Antagonists and agonists of the 5-HT2AR promote sleep and wakefulness, respectively (see Monti et al., 2018 for review), and both increases and decreases in 5-HT levels following sleep deprivation have been observed (see Menon et al., 2019 for review), although increased 5-HT levels may be more likely if combined with malnourishment (Alfaro-Rodríguez et al., 2006).
Severe and chronic pain and pain-processing abnormalities are associated with psychosis (Koyanagi and Stickley, 2015; Minichino et al., 2016). The function of 5-HT2AR is involved in nociception and antinociceptive response in peripheral tissue, spine and brain in ways that are too nuanced for generalizations or coverage here (Abbott et al., 1996; da Silva Soares et al., 2019; Seyrek et al., 2010; Zuo et al., 2019; see Cortes-Altamirano et al., 2018; Courteix et al., 2018 for review). Acute pain quite reliably increases 5-HT release (see Neugebauer, 2020 for review of serotonin modulation of pain) and increased 5-HT2AR binding in various cortical areas is associated with tonic pain ratings in humans (Kupers et al., 2009).
The 5-HT2AR also plays a role in body temperature regulation, with agonists reliably increasing core body temperature (Blessing and Seaman, 2003; Friedman and Hirsch, 1971; Klock et al., 1975; Liskow, 1971; Mazzola-Pomietto et al., 1995, Murakami et al., 1980; Pawlyk et al., 2006; Salmi and Ahlenius, 1998) and antagonists blocking this effect (Mazzola-Pomietto et al., 1995; Nisijima et al., 2001). Antagonism of the 5-HT2AR also reduces body temperature (Pawlyk et al., 2006) and blocks hyperthermic response to social defeat (Sinh and Ootsuka, 2019). Some evidence suggests that exposure to heat or cold increases 5-HT levels (Hale et al., 2011; Ishiwata and Greenwood, 2018; Kelly et al., 2011; Myers and Beleslin, 1971), but other studies found no relationship between temperature manipulations and brain 5-HT levels (Ishiwata et al., 2004; Nakagawa et al., 2016; Saito et al., 2005). We are not aware of any direct evidence showing that body temperature manipulation influences 5-HT2AR activity (Zamfir et al., 1992), but given the intentional manipulation of body temperature in spiritual and therapeutic practice (see below), there are logical reasons to suspect there may be a relationship.
Preliminary evidence suggests that decreased brain pH is an endophenotype for schizophrenia and bipolar disorders (see Hagihara et al., 2018 for review). Acidosis (low pH) could be caused by hypercapnia due to respiratory complications, high altitude, breathing techniques, as well as ketoacidosis associated with starvation, metabolic dysregulation and alcoholism. It has been proposed that brainstem and midbrain 5-HT neurons act as chemoreceptors sensitive to extracellular pH (Richerson, 2004; Teran and Richerson, 2020), potentially explaining how homeostatic imbalance caused by stress might generally engage the serotonergic system and elicit psychedelic-like subjective effects via heightened 5-HT2AR agonism. Experimentally exploring this relationship, for example by testing the capacity of a 5-HT2AR antagonist to block the effects of putative CO2-induced psychedelic-like experience (Meduna, 1950), would be a relevant and potentially fruitful future research avenue for the PiMS hypothesis.
The link between stress and psychosis is well established and consistent with the ‘diathesis-stress’ model of psychopathology in which pre-existing vulnerabilities (diathesis = predisposition) combine with stress to catalyse transition into illness (Belsky and Pluess, 2009). The causal link between stress and spiritual experience may be less obvious – but it is, in fact, supported by a wealth of evidence – as we shall see more clearly in the next section. In summary, a large variety of chronic and acute intense stressors upregulate and activate the 5-HT2AR and are associated with PiMS-relevant phenomena.
Intentional stress-induced pivotal mental states?
Is it possible that humans have intuited how to hijack or ‘hack’ their own physiology for the purpose of self-development? Asceticism, or the withdrawal from sensory stimulation and dedication to a simple but disciplined lifestyle, has an ancient history of association with altered states of consciousness (Hof and Rosales, 2011; Kotler and Wheal, 2017; Wimbush and Valantasis, 2002). Our perspective is that self-manipulated and intended ‘stress’ leading to increased 5-HT2AR signaling and associated PiMSs is a relevant candidate mechanism here. Various evidence supporting this view is provided below.
Intentional social isolation, often in nature, is associated with spiritual and transformative experiences (Naor and Mayseless, 2017). Many religious narratives report that religious exemplars (e.g. Moses, Jesus, Mohammed, Siddhartha, Lao Tzu) sought extreme solitude – often in conjunction with the discovery of their benchmark philosophies. With regard to cognitive stressors, meditation on sin, guilt, death and suffering play a role in the spiritual exercises of various religious traditions (Giustarini, 2012; Loyola, 2007). Celibacy is another ascetic practice (Olson, 2008) that may create tension and stress.
Fasting, or extreme moderation of food intake, is a common religious practice (Diamond, 2003; Dugan, 1995; Eskildsen, 1998; Laidlaw, 2005; Sakr, 1975). Bouts of extreme fasting, such as those reportedly undertaken by Jesus in the wilderness or Prince Siddhartha (the ‘Buddha’) before enlightenment, often precede important spiritual or religious revelations. Spiritual ‘athletes’ practice sleep restriction, promote night-time and/or early morning spiritual exercises, and in extreme cases, physically disable themselves from lying down (Farré-i-Barril, 2012; Macmillan, 2013). Endurance activities, such as the epic kaihōgyō (1000 marathons in 1000 days) or sun dance (Rhodes, 1987; Spier, 1921), are sometimes used for spiritual development, as is self-inflicted bodily pain and self-mortification (Camporesi, 1988). Manipulations of respiration and body temperature, as evidenced by yogic breathing (Brown and Gerbarg, 2005) and the Native American sweat lodge (Garrett et al., 2011), also play roles in spiritual practices and ceremonies.
Modern therapeutic techniques such as breathwork (Brown and Gerbarg, 2005; Grof and Grof, 2010) and whole-body hyperthermia (Hanusch et al., 2013; Janssen et al., 2016) continue to reinforce the idea that physiological stress or dysregulation can lead to meaningful changes in mental states and have been found to interface with the serotonin system (Anju et al., 2011; Buchanan et al., 2015; Pawlyk et al., 2006). The cross-cultural and recurring practice of asceticism cannot be explained as a purely culturally contingent phenomenon. As reviewed by Singh (2018) and noted by Winkelman and White (1987), ‘shamans’ (or their cultural equivalents), in various societies, engage in ascetic practices such as social isolation, food, sleep and sex restriction. Combining multiple ascetic practices with meditation, prayer or ritual and a background of stress may (perhaps inadvertently) create ideal synergistic conditions for the natural upregulation of 5-HT2AR signaling (Alfaro-Rodríguez et al., 2006) and thus, the emergence of a PiMS.
A crucial distinction between ascetic practices and unintentional counterparts, such as social exclusion or involuntary celibacy, is that the ascetic retains a sense of control over the stressors and a willingness towards self-transformation. Although some ascetics may be predisposed towards harm avoidance in the domains of social interaction or sexual relationships, for example, a positive estimation of this behaviour m0ay be protective against the derogatory and persecutory evaluations so integral to psychosis. This raises the important question of whether ascetic behaviour directly drives the occurrence of PiMSs, or whether the onset of PiMS biases a person towards social withdrawal, celibacy, starvation, as well as other behaviours associated with asceticism. As is often the case with such ‘chicken and egg’ type questions, the dilemma may be solved by invoking bidirectional causality.
While acknowledging the relevant causal pathways may operate bidirectionally and be mutually reinforcing, additional evidence for ascetic-like behaviour being causative of PiMSs is provided by secular uses of stress to induce altered states. For example, militaries regularly subject new recruits to psychosocial and physiological stressors in efforts to break down and reform the individual into a community-minded soldier (McGurk et al., 2006). It is also debated whether various forms of torture involving sleep deprivation, social isolation and pain may render an individual more suggestible to the planting and/or imprinting of new/false beliefs and memories (Bloche, 2016; O’Mara, 2015). Indeed, it seems logical, given the already cited associations between stress and the 5-HT2AR, that such torture techniques would elevate brain plasticity via upregulation and activation of 5-HT2ARs (Ly et al., 2018). It is therefore relevant and noteworthy that a significant history exists of combining torture, coercion and interrogation techniques with the administration of psychedelic drugs such as LSD (Lee and Shlain, 1992), which are known to promote suggestibility (Carhart-Harris et al., 2015).
Returning to the question at the beginning of this section, there are good reasons to surmise that, at some given period in our evolutionary development, a perhaps universal intuition arose amongst humans that physiological manipulation can induce hyper-plastic mind and brain states; a realization that has since been exploited throughout the ages for purposes of self-development and spiritual or religious growth (Kotler and Wheal, 2017). In the next section, we will begin to address in more detail some candidate physiological and neuropharmacological processes that are likely to serve an important causal and/or mediational role in the induction of PiMSs. Given the special reliability with which they can induce PiMSs, we focus on the action of psychedelic drugs.
Psychedelics, pivotal mental states and the 5-HT2AR
Psychedelics, psychopathology and spiritual experiences
As discussed earlier, certain psychotic states (e.g. early and acute psychosis) are considered important examples of naturally occurring PiMSs. The psychotomimetic (psychosis-mimicking) effects of classic 5-HT2AR agonist psychedelics have been well documented (Carhart-Harris et al., 2016b; Gonzalez-Maeso and Sealfon, 2009; Quednow et al., 2020; Vollenweider et al., 1997, 1998). Importantly, psychedelics are felt to be useful models of incipient psychotic states that may be more likely to display psychedelic-like phenomena (Bowers and Freedman, 1966), such as changes in perception, cognition and ego functioning (Bercel et al., 1956; Bowers and Freedman, 1966; Carhart-Harris et al., 2013; Fischman, 1983; Gouzoulis-Mayfrank et al., 2005; Savage, 1955; Savage and Cholden, 1956). Conversely, established psychotic disorders such as schizophrenia are more likely to feature characteristics of rigid cognition such as fixed delusions (Paolini et al., 2016; Rajapakse et al., 2011). Selective 5-HT2AR antagonism attenuates the main characteristic subjective effects of LSD, psilocybin and ayahuasca (Preller et al., 2017; Quednow et al., 2012) and the intensity of psychedelic states is reliably predicted by 5-HT2AR occupancy (Madsen et al., 2019).
Beyond the hypothesized involvement of the 5-HT2AR in the induction of psychotic states (Carhart-Harris et al., 2014b; Geyer and Vollenweider, 2008), it is generally thought that dopaminergic (DA) dysregulation serves as a ‘final common pathway’ underlying chronic psychoses (Howes and Kapur, 2009). In support of this notion, tolerance to sustained 5-HT2AR agonism develops quickly (Geyer and Vollenweider, 2008) and more selective 5-HT2AR antagonists lacking any appreciable DA antagonism properties have not proven to be efficacious antipsychotics (De Paulis, 2001; Garay et al., 2016; Meltzer et al., 2004). However, some evidence does suggest that 5-HT2AR binding and blockade may contribute to the superior efficacy of atypical antipsychotics (particularly clozapine) in attenuating positive and negative symptoms (Aringhieri et al., 2018, Meltzer and Massey, 2011; Richtand et al., 2007). Pimavanserin, a selective 5-HT2AR inverse agonist, has been licensed for psychotic symptoms in relation to Parkinson’s disease psychosis (Cummings et al., 2014; Mohanty et al., 2019), with some additional evidence of efficacy for psychotic symptoms in Alzheimer’s disease (Ballard et al., 2019) and schizophrenic psychoses unresponsive to clozapine (Nasrallah et al., 2019). It has also been demonstrated that, via antagonism of the 5-HT2AR, clozapine ameliorates the psychotomimetic effects of N-methyl-D-aspartate receptor (NMDAR) antagonism in animal models (Schmid et al., 2014, Szlachta et al., 2017). Similar findings have also been observed with risperidone (which has appreciable 5-HT2AR antagonist properties) and ketamine (an NMDAR antagonist) in humans (Joules et al., 2015); see (Carhart-Harris et al., 2013) for a critique of NMDAR antagonist drug models of psychosis.
Serotonin 2A antagonism has less marked effects on cognition than 5-HT2AR agonism but there is some evidence it can impair learning (Welsh et al., 1998; Zhang and Stackman, 2015) and promote compulsive behaviour (Kim et al., 2019; Schirmbeck and Zink, 2012) as well as sleep (Monti et al., 2018). Thus, it is possible that some of the therapeutic effects of 5-HT2AR antagonist antipsychotics may be due to a generic negative action on learning-related cognition and wakefulness. Given the central thesis of this paper, the deployment of 5-HT2AR antagonists early in the etiology of a psychotic disorder may conceivably prevent conversion to psychosis via suppressing the occurrence of a mediating pre- or peri-psychotic PiMS. Consistently, 5-HT2AR antagonism has been found to blunt or reduce cortical plasticity (Inaba et al., 2009; Jitsuki et al., 2011; Lombaert et al., 2018; Xu et al., 2012). Whether such actions have net positive or negative repercussions for long-term mental health is a complicated question, but see (Whitaker, 2011) for an interesting and relevant perspective.
Most post-mortem studies that have assessed 5-HT2AR levels have reported downregulated 5-HT2AR mRNA and protein expression in the cortex of individuals who had schizophrenia, and in vivo imaging of 5-HT2AR binding in schizophrenia also suggests decreased cortical receptor densities (see Quednow et al., 2020 for review). Decreased 5-HT2AR expression in schizophrenia may be state specific, as decreased 5-HT2AR binding in first-episode psychosis correlates with severity of positive symptoms (Rasmussen et al., 2010) and healthy monozygotic siblings of affected persons do not display decreases in 5-HT2AR binding (Rasmussen et al., 2016). It is plausible that in prolonged psychoses, the 5-HT2AR becomes adaptively downregulated after an initial period of heightened activity, consistent with agonist-induced downregulation of the 5-HT2AR (Erritzoe et al., 2011). Evidence of epigenetic modification of the 5-HT2AR in schizophrenia (Abdolmaleky et al., 2011; Cheah et al., 2017) is also relevant here.
It seems plausible that a cortical abnormality mediated by increased 5-HT2AR signaling and related glutamatergic activity (Aghajanian and Marek, 2000; Moreno and González-Maeso, 2018) and featuring abnormal plasticity (Kavanagh et al., 2015; Stephan et al., 2009) and associative learning – as mediators of major psychological change – is an important early component of the psychotic process in schizophrenia; whereas 5-HT2AR-mediated dysregulation of dopamine activity is a consequential, and perhaps defining component, namely ‘the final common pathway’ (Howes and Kapur, 2009; Pehek et al., 2006; Pehek and Hernan, 2015; Stahl, 2018). If we allow ourselves to be instructed on the pharmacology of the psychotic process via its phenomenology, then an initial state characterized by ego-disturbance and cognitive and perceptual disturbance preceding subsequent inflexible or perseverative cognitive and behavioural styles (Boulougouris et al., 2008; King et al., 1974; Murphy-Beiner and Soar, 2020) might fit with an initial serotonergic (5-HT2AR) component (although see Boulougouris et al., 2008), followed by a final pathway that is dominated by a hyperactive mesolimbic dopamine system (Stahl, 2018). However, converging evidence also suggests that upregulation (Chiu et al., 2014; Hámor et al., 2018; Napier and Istre, 2008), sensitization (Chiu et al., 2014; Hámor et al., 2018; Napier and Istre, 2008), and direct agonism (Soman et al., 2019) of 5-HT2AR contributes to dopamine-induced psychoses (Cummings et al., 2014; Meltzer et al., 2010). Indeed, manic states might precipitate incipient psychoses in a similar fashion (Correll et al., 2007; Jauhar et al., 2017). It may be relevant in this sense that some shared features have been found in the phenomenology of spiritual experiences and positive psychotic symptoms and the psilocybin experience whereas manic symptoms appear to be mimicked more reliably by pro-DA stimulant drug experiences (Carhart-Harris et al., 2013).
Beyond psychosis, psychedelics offer a reliable model of ‘peak’ (Roseman et al., 2018), ‘god-encounter’ (Griffiths et al., 2019), ‘mystical-type’ (Griffiths et al., 2006, 2008, 2011), near-death (Strassman, 2000; Timmermann et al., 2018) and other such anomalous experiences (Carbonaro et al., 2017) that bear an undeniable resemblance to experiences designated as ‘religious’ or ‘spiritual’. Trait absorption is one of the most reliable predictors of sensitivity to the subjective effects of psychedelics (Haijen et al., 2018) and has also been found to predict spiritual experiences induced via various means (Lifshitz et al., 2019). The 5-HT2AR rs6313 TT genotype is associated with trait absorption and altered time-perception in humans, both of which are implicated in spiritual experiences and NDEs (Cant et al., 2012; Ott et al., 2005; Sysoeva et al., 2010). NDEs and psychedelic experiences feature ‘time-dilation’ (Yanakieva et al., 2019) and altered time perception has also been reported in psychotic states (see Thoenes and Oberfeld, 2017 for review) and PTSD (Ahmadi et al., 2019).
The psychology of the psychedelic state
We begin this section with a focus on the psychology of the psychedelic experience before turning our attention to its neurobiology, with specific reference to human brain imaging studies with psychedelics. Psychedelics induce cognitive-perceptual instability, relax one’s normal sense of agency and ownership over perceptual objects, including one’s own body and at higher doses, reliably produce different degrees of ‘ego-dissolution’ (Millière, 2017; Nour et al., 2017). Psychedelics also elicit an increased emotional lability – not inconsistent with infancy, early psychosis and spiritual experiences (Carhart-Harris et al., 2016b). The recently developed RElaxed Beliefs Under pSychedelics (REBUS) model proposes that, via a 5-HT2AR-induced entropic effect on cortical activity (Carhart-Harris and Friston, 2019), psychedelics relax the precision weighting (i.e. inverse variance) of high-level priors (internal predictive models), thereby allowing bottom-up information (‘prediction error’) to flow more freely up the brain’s functional hierarchy to impress on high-level cortices and enter conscious awareness. It is proposed that this process is necessary for the effective revision of priors or beliefs (Carhart-Harris and Friston, 2019). This model has recently received support from the application of travelling wave analyses to DMT electroencephalogram (EEG) data, where a rapid shift from top-down to bottom-up dominating waves coincided very closely the onset and subjective intensity of the DMT experience (Alamia et al., 2020).
The REBUS model is consistent with much of the phenomenology of the psychedelic experience, including intense spiritual, ‘peak’ or ‘mystical-type’ experiences (see Barrett and Griffiths, 2017; Roseman et al., 2018). In these (typically high-dose) experiences, discriminative beliefs (e.g. A is different to B) are often moderated and replaced by a sense of reciprocal interconnectedness, which is referred to as the ‘unitive experience’ in studies of ‘mystical’ or ‘spiritual’ experiences (Stace, 1960). Such experiences may account for lasting psychological changes seen with psychedelics, such as enduring increases in the personality trait ‘openness’ (Erritzoe et al., 2018; MacLean et al., 2011; Nour et al., 2017) as well as improvements in wellbeing (Haijen et al., 2018). Thus, during and after profound psychedelic experiences, specific beliefs and the emotional valences attached to them seem particularly susceptible to change, consistent with so-called ‘quantum change experiences’ (C’De Baca and Wilbourne, 2004; Miller and C’de Baca, 2001).
Returning to the phenomenology of the psychedelic state, peak, mystical-type or spiritual experiences and associated feelings of oceanic boundlessness have been found to reliably predict positive outcomes in psychedelic therapy (Griffiths et al., 2016; Haijen et al., 2018; Roseman et al., 2018; Ross et al., 2016). However, they may also represent an extreme experience only achievable (to most people) through pharmacological manipulation. Achieving the same quality and intensity of experience may be difficult via spiritual practice alone; although a recent study comparing the interaction between psychedelics and meditation has suggested a potential synergistic effect (Smigielski et al., 2019; see also Millière et al., 2018). Given our focus on stress-induced PiMSs and associated psychological transformation, it is worth taking a closer look at the common themes of psychological struggle and breakthrough in psychedelic experience (Roseman et al., 2019).
Through the relinquishment of top-down control effected by high-level priors (Carhart-Harris and Friston, 2019), psychedelics may free suppressed emotions and memories so they may more easily percolate into consciousness awareness (Alamia et al., 2020; Carhart-Harris et al., 2014a). Such emotions and memories may be felt as being emotionally challenging (Barrett et al., 2016). Feelings of anxiety are common during and after psychedelic experiences (Barrett et al., 2016; Belser et al., 2017; Carbonaro et al., 2016; Eisner and Cohen, 1958; Gasser et al., 2015; Roseman et al., 2018, 2019) as are symbolic/archetypal themes (Hill, 2013; Malone et al., 2018; Masters and Houston, 2000 [1966]; Shanon, 2002). Intense personal and transpersonal themes can rise to the forefront of awareness (Belser et al., 2017). Values can be ‘remembered’ (Belser et al., 2017) and affect-laden beliefs that previously seemed ‘abstract’ (e.g. ‘love is fundamental’) can be deeply felt (Belser et al., 2017; Pollan, 2019). Cathartic release under psychedelics (Roseman et al., 2018, 2019) may foster an emotional re-evaluation of cognitive (Lyons and Carhart-Harris, 2018b) and philosophical perspectives (Lyons and Carhart-Harris, 2018a) that may have previously been closely tied in with a person’s pathology.
Appropriately managed, psychedelic experiences can lead to an increased willingness and an ability to engage with emotionally difficult psychological material (Roseman et al., 2018; Watts et al., 2017). In therapeutic contexts, this experience is generally positive and is associated with an enhanced sense of emotional empathy (Dolder et al., 2016; Pokorny et al., 2017) and pro-social feelings and behaviour (Griffiths et al., 2018). It warrants stating, however, that extreme negative affect, including paranoid ideation and occasional injurious behaviour, can also arise during psychedelic experiences (Carbonaro et al., 2016; Coid et al., 2016; Honings et al., 2016; Strassman, 1984), particularly if the contextual frame in which the experience occurs is not sufficiently well controlled and supportive.
The long-term psychological effects of psychedelics
The following psychological factors have been found to be altered in an enduring way with psychedelics: (a) personality (Erritzoe et al., 2018; Lebedev et al., 2016; MacLean et al., 2011); (b) mental health and wellbeing (Argento et al., 2017; Bogenschutz et al., 2015; Carhart-Harris et al., 2017, 2018b; Gasser et al., 2015; Griffiths et al., 2006, 2008; Haijen et al., 2018; Hendricks et al., 2015; Johnson et al., 2014; Osório et al., 2015; Moreno et al., 2006; Ross et al., 2016); (c) political perspective (Lyons and Carhart-Harris, 2018a; Nour et al., 2017); (d) lifestyle preferences (Forstmann and Sagioglou, 2017); and (e) feelings towards the environment/nature (Forstmann and Sagioglou, 2017; Lyons and Carhart-Harris, 2018a; Kettner et al., 2019).
The neurobiological effects of psychedelics
Functional brain imaging studies of the acute psychedelic state using a variety of psychedelics have observed decreases in the functional modularity of a broad range of functional modules, including high-level networks such as the default-mode and fronto-parietal control network (Carhart-Harris et al., 2016c; Lebedev et al., 2015). Moreover, decreases in the integrity of these networks (Carhart-Harris et al., 2016c; Lebedev et al., 2015) as well as their reduced presence at any given point in time (Lord et al., 2019) appear to relate to high-level aspects of the drug-induced subjective experience, including ‘ego-dissolution’. Under psychedelics, the normal functional segregation or specialization in the brain is instead replaced by a globally interconnected profile (Carhart-Harris et al., 2012a, 2016c; Lord et al., 2019; Petri et al., 2014; Preller et al., 2018; Smigielski et al., 2019; Tagliazucchi et al., 2014). It is natural to speculate that this increase in global connectivity in the brain under psychedelics relates to the weakening of discriminative cognition (e.g. A versus B), exemplified by the unitive experience as well as ‘non-dual awareness’. It is easy to see how something akin to the unitive experience may occur in early psychosis, in which compromised ego-boundaries are reported (Carhart-Harris et al., 2012b; Parnas and Handest, 2003) and similar changes in functional connectivity have been seen in early psychosis as with psychedelics (Carhart-Harris et al., 2013). Decreased orthogonality between high-level networks (e.g. the default mode and salience networks) may also be a sub-acute effect of ayahuasca (Pasquini et al., 2020), although this effect has not been seen consistently (sub-acutely) with psilocybin (Barrett et al., 2020; Carhart-Harris et al., 2017).
Psychedelics have been shown to promote extinction-learning through 5-HT2AR agonism (Cameron et al., 2018; Catlow et al., 2013; Young et al., 2017) as well as low-level associative learning (Gimpl et al., 1979; Harvey, 1996, 2003; King et al., 1974; Morici et al., 2018; Romano et al., 2010; Welsh et al., 1998; Zhang et al., 2013, 2016a; Zhang and Stackman, 2015), for example, as has been nicely demonstrated by the catalysing influence of LSD (a 5-HT2AR agonist) on learning rate versus the impairing influence of ritanserin, a 5-HT2AR antagonist (Harvey, 2003).
Consistently, psychedelics have been shown to potently increase cortical neuroplasticity (Benekareddy et al., 2012; Cavus and Duman, 2003; Frankel and Cunningham, 2002; Gewirtz et al., 2002; Jaggar and Vaidya, 2018; Jones et al., 2009; Meller et al., 2002; Niitsu et al., 1995; Vaidya et al., 1997). A doubling of BDNF mRNA was found in the parietal cortex after administration of the 5-HT2AR agonist DOI (Vaidya et al., 1997) and markedly increased functional and structural neuroplasticity has been found after DMT, LSD and 2,5-Dimethoxy-4-iodamphetamine (DOI) (Ly et al., 2018) – leading the senior author to introduce the term ‘psychoplastogen’ for these and any other relevant compounds that can rapidly elicit appreciable increases in neuroplasticity (Olson, 2018). We have preliminary evidence of increased visual evoked long-term potentiation (LTP) in healthy individuals under the influence of psilocybin (Olson, 2018), and long-term changes in network functionality have also been observed 1 day (Carhart-Harris et al., 2017), 1 week (Barrett et al., 2020) and 1 month after psilocybin (Lyons, 2020) in clinical (Carhart-Harris et al., 2017) and non-clinical populations (Carhart-Harris et al., 2012a).
Increased (spontaneous) brain (activity) entropy or complexity is a highly reliable marker of the acute functional brain effect of psychedelics (Abasolo et al., 2015; Carhart-Harris, 2018b; Dolan et al., 2018; Fernández et al., 2013; Lebedev et al., 2016; Lyons, 2020; Vivot et al., 2020). Increased brain complexity has also been observed during rapid eye movement (REM) sleep (Abasolo et al., 2015), meditation (Vivot et al., 2020), musical experiences (Dolan et al., 2018) and certain psychotic states, with medication status, age and the stage of the psychotic process the key parameters to consider when assessing this (Fernández et al., 2013).
Acute brain entropy under LSD, measured with functional magnetic resonance imaging (fMRI), was found to predict psychological changes 2 weeks later, namely increased trait ‘openness’ (Lebedev et al., 2016) and in a more recent study, increased brain complexity under psilocybin was found to predict both long-term changes (1 month later) in brain network functional connectivity (Lyons, 2020) as well as related improvements in wellbeing (Lyons, 2020). Future neuroimaging analyses of ours will assess how acute brain complexity relates to functional proxies of LTP as well as long-term anatomical changes in the human brain. It is tempting to speculate that the acute brain changes seen with psychedelics, including increased complexity or entropy, are reflective of a hyper-plastic state that mediates subsequent functional and potentially structural brain changes that correlate with long-term psychological changes.
The adaptive function of pivotal mental states
How could you rise anew if you have not first become ashes. (Nietzsche, Thus Spoke Zarathustra)
As discussed in previous sections, certain types of stress upregulate 5-HT2AR expression and activity (see Table 1 for a review of the relevant literature). Moreover, evidence that acute stress causes the release of 5-HT is compelling (Adell et al., 1997; Amat et al., 1998, 2005; Bastani et al., 2017; Beitia et al., 2005; Bland et al., 2003; Cohen et al., 2015; Ferres-Coy et al., 2013; Fuenmayor and Garcia, 1984; Fujino et al., 2002; Gardner et al., 2005; Hale et al., 2011; Higuchi et al., 2019; Ishida et al., 1997; Ishiwata and Greenwood, 2018; Johnson et al., 2005; Keeney et al., 2006; Kelly et al., 2011; Li et al., 2015; Myers and Beleslin, 1971; Nakajima et al., 2009; Neugebauer, 2020; Paul et al., 2011; Rex et al., 2005; Yoshioka et al., 1995) and it is logical to surmise that increased endogenous 5-HT release will engage the 5-HT2AR system, an assumption backed up by evidence of increased 5-HT2AR associated responses with drug-induced 5-HT release (Kuypers et al., 2018; Liechti et. al., 2000; Pitts et al., 2017; van Wel et al., 2012). These factors have led us (Carhart-Harris and Nutt, 2017) and others (Murnane, 2019) to speculate that the 5-HT2AR system is a stress response system that services adaptability.
Natural questions that follow from this model include why does this function exist and what evolutionary purpose does it serve? Our proposal is that PiMSs have evolved to allow the experiencer a psychological ‘fresh start’, akin to a psychological ‘rebirth’ (Brodersen and Glock, 2016; White, 2004) or allostatic recalibration (Ellis and Del Giudice, 2019). Rather than simply referring to adaptive responses to stress and adversity, which could include many of the symptoms of psychiatric disorders, the PiMS model is intended to invoke the idea that such adaptation can be radical, rapid and discrete, rather than moderate, slow and continuous. These ideas are at least partially consistent with the notion of allostasis (McEwen, 2019; Sterling and Eyer, 1988) and the adaptive calibration model of stress response (Ellis and Del Giudice, 2019), which, like the PiMS model, refuses to characterize adaptive responses to stress as necessarily ‘toxic’ or pathological.
Thus, according to the PiMS model, the outcome of an allostatic recalibration process can be ‘positive’ (e.g. in terms of a therapeutic or spiritual breakthrough) but the same mechanisms could just as easily result in a new or reinforced maladaptive strategy, perhaps best exemplified by a psychotic ‘flight from reality’ (Broome et al., 2007; Freeman et al., 2014; Lyon et al., 1994) or progressive reinforcement of other psychological defence mechanisms. Consistent with previous work (Carhart-Harris, 2018a; Carhart-Harris and Nutt, 2017), it was recently hypothesized that 5-HT2AR signaling forms part of a ‘stress response system’ that serves the adaptive function of reconfiguring responses to environmental stimuli co-occurring with or preceding major perceived existential threats or crises (Murnane, 2019). Thus, the principle is that accelerating learning (Harvey, 2003) and extinction learning or ‘unlearning’ (Cameron et al., 2018) in such extreme situations should aid the individual in the future by causing a domain-general ‘overhaul’ in their outlook.
Evidence of aberrant neuroplasticity in psychosis (Stephan et al., 2009), including both increases (Yee et al., 2018) and decreases (Javitt et al., 1996; Umbricht and Krljes, 2005) in neuroplasticity markers, for example, particularly during critical developmental windows, may bear relevance to the reliable increases in plasticity linked to 5-HT2AR agonism (Ly et al., 2018; Olson, 2018), which are linked here with the PiMS construct. As discussed above, one way in which apparent inconsistencies in the literature may be reconciled is to consider process relevant dynamics, where, during a process, a phase of upregulation in a particular parameter (e.g. 5-HT2AR signaling and associated plasticity) can be followed by a phase of adaptive downregulation is the same parameter (e.g. decreased 5-HT2AR expression). Indeed, agonist-induced downregulation of the 5-HT2AR is entirely consistent with this principle (Bull et al., 2004; Erritzoe et al., 2011; Muguruza et al., 2014; Reneman et al., 2002; also see Madsen et al., 2020). From a neurodevelopmental perspective, we must also acknowledge that hyperactivation of 5-HT2ARs coinciding with childhood stress or chronic drug-use could negatively affect neurodevelopmental processes. Again, however, the influence of context here must be considered.
Near-death experiences are relevant to the theory that the 5-HT2AR system has evolved to aid psychological transformation for adaptive ends. Altered perceptual processing, heightened emotional tone, time dilation and enhanced episodic memory encoding are all features of NDEs. The major shifts in perspective following NDEs may be the product of a period of hyper-plasticity, potentially mediated by either the massive release of 5-HT or indeed endogenous 5-HT2AR agonists such as DMT during the dying process (Dean et al., 2019). One view is that NDE, and associated hyper-plasticity, may function to aid the individual if such life-or-death-type scenarios were to be met with again in the future. However, another, not incompatible view, is that the dramatic cognitive set-shifts seen after NDEs reflect a more fundamental ‘opening-up’ of high-level mental schemata to revision, consistent with a generic recalibration process and the recent REBUS model (Carhart-Harris and Friston, 2019). Major cognitive revisions may enable an individual to ‘start afresh’ in both the behavioural and cognitive sense, with a refined and recalibrated (‘judgement light’) perspective (Carhart-Harris and Friston, 2019). The same functional explanation could also be used in reference to ‘post-traumatic growth’, a construct that, at least by name, is only two decades old (Tedeschi, 1999). The notions of allostasis (McEwen, 2019; Sterling and Eyer, 1988) and ‘adaptive plasticity’ and ‘adaptive calibration’ (see Ellis and Del Giudice, 2019 for review) are also relevant here.
A relevant recent perspective on the evolutionary origin and potential treatment of PTSD (Murnane, 2019) is largely consistent with an earlier perspective piece on 5-HT2AR-mediated ‘active coping’ and radical adaptation proposed by Carhart-Harris and Nutt (2017), as well as animal work linking serotonergic functioning to faster learning rates (Harvey, 2003; Iigaya et al., 2018), plasticity (Iigaya et al., 2018) and adaptability (Matias et al., 2017). It is also consistent with a growing literature base supporting the value of classic (5-HT2AR agonist) psychedelic therapy in the treatment of a broad range of psychiatric disorders (Carhart-Harris and Goodwin, 2017; Nutt et al., 2020; Rucker et al., 2018). A crucial component of the 5-HT2AR-mediated active coping model (Carhart-Harris and Nutt, 2017) was that the relationship between stress and upregulated 5-HT2AR signaling is non-linear, in the sense that once stress crosses a critical threshold (the specifics of which are presently not clear) – in terms of its severity or chronicity – there is an abrupt shift in a system’s functioning (e.g. global brain function) into a radically different mode (also consistent with bifurcation theory). The PiMS model maintains that the emergence of this mode is mediated by increased 5-HT2AR activity and associated plasticity (Olson, 2018) and its evolutionary and adaptive function is to aid radical psychological change when its need is perceived (Carhart-Harris and Nutt, 2017). Evidence for such nonlinearities in the functioning of the 5-HT2AR system do indeed exist (Dwivedi et al., 2005; Madsen et al., 2019) and should be the focus of future PiMS-relevant hypothesis testing.
Hierarchical predictive processing is an increasingly influential model of global mind and brain function that essentially posits that brains evolve and develop into predictive models of the environments they inhabit. Much of brain anatomy and function appears to be organized hierarchically (Felleman and Van, 1991; Margulies et al., 2016) and it is proposed that relevant predictive mechanisms are encoded in the brain’s multi-level functional and anatomical hierarchies (Clark, 2013; Friston, 2018; Rao and Ballard, 1999; Seth, 2013). The hierarchical predictive processing perspective has recently been applied to the action of psychedelics (i.e. in the so-called ‘REBUS’ model). This model, which is receiving growing empirical support (Alamia et al., 2020; Girn et al., 2020), argues that the de-weighting (decreased ‘precision’) of internal predictive models (‘prior probability distributions’) under psychedelics opens a window for their subsequent revision (Carhart-Harris and Friston, 2019). Consistently, it can be hypothesized here that in individuals undergoing intense PiMSs, the brain inhabits a mode of functioning that is conducive to the modulation or recalibration of internal predictive models, including how they are weighted.
Major recalibrations to fundamental beliefs or outlooks have been referred to in the psychology literature as psychological ‘transformations’ or ‘rebirths’ (White, 2004) and have received particular attention under the construct of ‘quantum change’ (C’De Baca and Wilbourne, 2004; Miller and C’de Baca, 2001). We believe the present PiMS model extends this previous (largely phenomenological) work by proposing a plausible mechanistic account for these phenomena. One may draw parallels between the so-called REBUS model (Carhart-Harris and Friston, 2019), the therapeutic application of psychedelics (Watts et al., 2017; Watts and Luoma, 2019) and the Acceptance and Commitment Therapy (ACT) psychotherapeutic model (Walsh and Thiessen, 2018), where the ACT approach endeavours to promote ‘psychological flexibility’, that is, flexible acceptance and integration of emotionally challenging memories and emotions, and seeks to do this via techniques such as ‘cognitive de-fusion’, which are exercises intended to foster an open, tolerant, inquisitive and accepting stance on psychological suffering and its causes. The possible combination of ACT or related mindfulness-based techniques with psychedelic therapy has been discussed in recent review papers (Sloshower et al., 2020; Walsh and Thiessen, 2018; Watts and Luoma, 2019) and is now being supported by empirical findings (Close et al., 2020; Davis et al., 2019).
Therapeutic implications
The primary purpose of this review is to introduce a new psychological and neurobiological construct, the PiMS, and examine its potential causes, mechanisms and functions. PiMSs are defined by three key criteria: elevated cortical plasticity, enhanced associative learning (including extinction learning) and a special capacity to mediate psychologically transformative change. The psychedelic state can be considered a prototypical PiMSs and there is clear evidence this particular state meets all three of its defining properties (Carhart-Harris et al., 2018c; Carhart-Harris and Friston, 2019; Griffiths et al., 2006; Harvey, 1996; Ly et al., 2018). Importantly, however, it is our view that PiMSs represent a more fundamental state. We have argued that the same molecular or proteomic gateway (i.e. 5-HT2AR signalling) is implicated in non-drug, ‘naturally’ occurring PiMSs, as in the psychedelic state. Moreover, we argue that the psychological properties of psychedelics depend on their ability to hijack this natural system, which has evolved for mediating rapid, major and potentially lasting adaptive change.
In this section, we address the therapeutic implications of the PiMSs model. Clinically supervised psychedelic experiences are showing promise for the alleviation of a large number of psychiatric symptoms and unhealthy lifestyle habits (Bogenschutz and Johnson, 2016; Carhart-Harris et al., 2018a; Carhart-Harris and Goodwin, 2017; Johnson, 2018). Psychedelic therapy appears to mediate positive mental health change in a customizable and structured way, achieving a remarkable degree and reliability of therapeutic change, often via a small number of isolated therapeutic sessions, supported by subsequent psychological integration work, designed to sustain the relevant positive outcomes (Richards, 2015; Walsh and Thiessen, 2018). In contrast to the often-distressing conditions that naturally agitate the manifestation of a PiMS, psychedelic treatment primes positive transformation via the structuring of ‘set and setting’ (Carhart-Harris et al., 2018c; Hartogsohn, 2016) as part of a broader notion of ‘context’ (Carhart-Harris et al., 2018c).
In addition to the treatment of diagnosed psychiatric illness, present findings indicate that psychedelic therapy offers an opportunity for the de-weighting of a plethora of maladaptive cognitive/perceptual schemas or ‘sets’ about self, others and the world (Carhart-Harris and Friston, 2019; Hinton and Kimayer, 2017). This realization implies a broad, ‘transdiagnostic’ therapeutic value (Figure 6). Relatedly, data indicate the deployment of this intervention may yield benefits beyond diagnosed mental illness. For example, given observed improvements in psychological wellbeing (Jungaberle et al., 2018; Walsh, 1982) including resilience-related changes (Close et al., 2020; Davis et al., 2019), it is conceivable that psychedelic therapy could be used as a prophylactic or preventative intervention, for example, promoting adaptability in adversity (Close et al., 2020; Davis et al., 2020; Murphy Beiner and Soar, 2020). We are aware that similar inferences have been made about ACT and psychological flexibility, namely that ACT can be transdiagnostically effective and ‘psychological flexibility’ is universally relevant to mental health (Hayes, 2019).

A conceptual schematic illustrating the putative transdiagnostic relevance of over-weighted internal predictive models or ‘priors’ and how they may be effectively targeted by carefully engineered pivotal mental states (PiMSs). This is represented metaphorically by a dart (serotonin 2A receptor (5-HT2AR) signaling inducing a PiMS), targeting the central space where different psychopathologies overlap. The shared characteristic of these psychopathologies is hypothesized to be over-weighted predictive models. Such over-weighted priors may be thought of as aberrant habits or biases (e.g. negative cognitive biases in depression, aberrant self-image in eating disorders or compulsive behaviours in OCD or addiction). Effective PiMS-based therapy should aim to remediate this problematic over-weighting by: (a) acutely ‘relaxing’ problematic priors and then (b) working on promoting a more permanent recalibration of beliefs or bias, for example via a commitment to a healthy behavioural change. Note: the schematic should not be (mis)interpreted as implying psychedelic therapy is a ‘cure all’.
There are (emotional) pros and cons to recognizing PiMSs as important – perhaps universally accessible – states of mind and brain and we welcome future critique of the idea. For example, it may be (emotionally) preferable to maintain strong distinctions between spiritual experiences and psychotic episodes and reject or ignore some of the similarities (DeHoff, 2015; Parnas and Henriksen, 2016). It is easy to appreciate the sensitivity of this debate: the negative stigma, prognoses and interventions that come with a diagnosis of a psychotic disorder (Wong et al., 2009) are a heavy burden, whereas evidence that spiritual experiences and religious conversions are often associated with positive mental health outcomes (Griffiths et al., 2011) argues against them being linked with pathology.
Relatedly, we anticipate some pushback to what is, essentially, a secular, naturalistic/scientific approach to phenomena others might consider ‘supernatural’, particularly given the aforementioned emotional function that these beliefs may serve (Kornfield, 2001). As discussed above with reference to predictive coding, in advocating a particular framework of belief that is meant to be held with the highest level of certainty, believers are promised an escape from an existential uncertainty (Hillen et al., 2017). Given the emotional appeal of such a prospect, it may be unsurprising that a readiness to endorse metaphysical or supernatural beliefs after a psychedelic experience is associated with (at least short-term) improvements in psychological wellbeing (Timmermann et al., 2019). However, important questions need to be asked here about the sustainability of any such effect. One should be mindful of how recourse to supernatural belief may offer the believer a means of emotional escape, consistent with the phenomenon known as ‘spiritual bypassing’ (Masters, 2010), which was also highlighted above. One must also be mindful, however, how an excessive recourse to the rationalism of science could promote a ‘cognitive fusion’ working against rather than for psychological flexibility and associated wellbeing (Hayes, 2019). The key here may be to retain a healthy and appropriate scepticism and uncertainty (e.g. of both specific scientific principles and religious dogma), balanced with respect for assumptions that have been found (e.g. via the scientific method) to be reliable and robust to scrutiny (Rovelli, 2018).
We have placed significant emphasis on the 5-HT2AR system throughout this paper, but it should be acknowledged that some prefer to extend the definition of psychedelics beyond action at this particular receptor. MDMA (Siegel, 1986) and ketamine (Bowdle et al., 1998) are two relevant examples in this regard (Kuypers et al., 2018; Lin et al., 2018; Schmid et al., 2014, Szlachta et al., 2017). Like the classic psychedelics, MDMA and ketamine have both been used as adjuncts to psychotherapy, with notable success (McGirr et al., 2017; Sessa et al., 2019; Wilkinson et al., 2017). However, the duration of the therapeutic response associated with classic (direct 5-HT2AR agonist) psychedelics (Carhart-Harris and Goodwin, 2017) appears to exceed that associated with a single exposure to ketamine and although MDMA causes an appreciable increase in activity at the 5-HT2AR through its potent release of 5-HT (van Wel et al., 2012), the non-specific nature of MDMA’s pro-serotonergic, and indeed noradrenergic and DA effects, complicates our understanding of its (pharmaco)therapeutic mechanisms (Liechti et al., 2000).
It is relevant to ask whether we feel it possible for PiMSs to occur without the involvement of increased 5-HT2AR activity. Although it is plausible to imagine this possibility, for example via a non-5-HT2AR-induced increase in cortical excitation, it also seems reasonable to assume that increased 5-HT2AR signaling may be a particularly robust and reliable inducer. One implication of this is that blocking the 5-HT2AR, as is done routinely in psychiatry via a broad range of psychiatric medications, would make PiMSs, and thus, transformative psychological change, less likely. If this is the case, it might support the fear of some that chronic medication with conventional psychiatric medication diminishes the chances of spontaneous remission and renders the medicated person vulnerable to relapse if the stabilizing medication is withdrawn (Moncrieff, 2008; Omachi and Sumiyoshi, 2018; Whitaker, 2011). Accordingly, remaining unmedicated might provide greater opportunity for self-development (Whitaker, 2011).
The obvious counterargument to this perspective, however, is that untreated psychoses render an individual more susceptible to relapse and that recurrent psychotic episodes serve to further reinforce illness chronicity (Emsley et al., 2013; Omachi and Sumiyoshi, 2018). It is also debatable whether the incipient phase of a recurrent psychotic episode that emerges inadvertently, with few warning signs (Emsley et al., 2013) and features severe paranoia and cognitive and behavioural disturbances, really represents a viable situation for (psycho)therapeutic intervention (Omachi and Sumiyoshi, 2018). Certain remote contextual factors contributing to the emergence of psychotic features, such as polygenics and or early-life trauma, might make the shepherding of a PiMS in a positive direction particularly challenging and, indeed, risky to attempt.
This debate speaks to an important cautionary point about presenting PiMSs as inherently therapeutic. Although PiMSs, by definition, represent opportunities for major change, change is not inevitable and neither are ‘positive’ outcomes necessarily more likely than negative ones. In keeping with the so-called ‘plastic paradox’ (Doidge, 2007), in the same way that states of hyper-plasticity can aid unlearning in the service of new perspectives and behaviour, they can also serve to reinforce old beliefs and biases if the relevant entraining conditions exist.
Limitations
This paper is a theoretical piece with a narrative style. Such approaches inevitably sacrifice self-critique and counterargument in favour of placing the spotlight on supportive evidence. There are obvious limitations to this style, with a ‘cherry-picking’ confirmation bias being one notable problem. We therefore strongly encourage critiques of this work. Moreover, although we have tried to be thorough in our coverage of the literature, it not feasible to be entirely exhaustive and it is inevitable that some truncation and oversight of relevant material has occurred. For example, the perspective that psychedelics model aspects of both (early and acute) psychosis and spiritual experience is treated as largely consensual (Barrett and Griffiths, 2017; Carhart-Harris et al., 2016b; Gonzalez-Maeso and Sealfon, 2009) but either of these positions could be challenged.
This paper’s focus on a small number of example PiMSs could also be questioned. One might argue that the chosen examples are not necessarily ideal and, more specifically, that too much emphasis has been placed on psychosis in particular, ahead of other compelling alternatives, such as trauma or panic attacks. Our preferential focus on psychosis stems from: (a) the wealth of research that has been conducted on psychosis, particularly in relation to the role of the 5-HT2AR system, and (b) that it has a history of having been compared and contrasted with spiritual experiences, which is a conflict that the PiMS model bears special relevance to. Other states we could have looked at in relation to PiMSs include REM sleep (Carhart-Harris, 2007; Carhart-Harris and Nutt, 2014; Hobson et al., 2014; McNamara, 2016; Sanz et al., 2018), particularly given its phenomenological qualities (McNamara, 2019), sensitivity to 5-HT2AR manipulation (Monti et al., 2018; Monti and Jantos, 2006), evidence for its role in processing stressful scenarios (Suchecki et al., 2012) and similarities between its neural correlates and those of the psychedelic state (Carhart-Harris, 2007; Carhart-Harris and Nutt, 2014; Kraehenmann, 2017). Other candidates include the ‘dreamy-state’ aura of temporal lobe epilepsy (Carhart-Harris, 2007), meditative states (Millière, 2017) and certain states observed in dissociative identity disorder (Laddis et al., 2017).
Our emphasis on the importance of contextual factors may seem neglectful of possible genetic and neurodevelopmental predispositions to psychosis and the heterogeneous aetiologies of psychotic disorders more generally. We do accept that not all psychoses have their origins in easily identifiable PiMSs and that genetic and neurodevelopmental factors can influence both the occurrence and outcome of a PiMS. Thus, contrary to dismissing remote aetiological factors, it is our hope that this review will increase interest in translational research and a biopsychosocial perspective that appreciates the relevance of conditional and relational dependencies.
Our selective focus on the 5-HT2AR might also be critiqued but we feel this is justified by the wealth of supportive evidence for its association with PiMS-relevant phenomena and that this may have been overlooked in the past. The present upsurge in interest in classic psychedelics should be welcomed as it has historically been unreasonably difficult to conduct research with these compounds (Nutt et al., 2013). It is inevitable that progress in our understanding of the 5-HT2AR system will have suffered as a result. We acknowledge that other neurotransmitters such as noradrenaline and acetylcholine (Angela and Dayan, 2005), glutamate (Corlett et al., 2016), dopamine (Howes and Kapur, 2009) as well as the endocannabinoid (Ibarra-Lecue et al., 2018) and opiate systems (Butelman and Kreek, 2015) will likely play a role in shaping the quality of PiMSs and their outcome but we maintain that the 5-HT2AR plays a particularly central role, deserving of special attention.
We might also consider whether PiMSs are always transient states or whether they can occur as more protracted episodes or phases, perhaps with more attenuated characteristics in each of the three defining criteria. Infancy, and childhood more generally, might be considered a protracted PiMS or pivotal mental phase. Indeed, the quality of consciousness in infancy has been likened to psychedelic (Carhart-Harris et al., 2018b; Carhart-Harris and Nutt, 2014) and spiritual states (Woodsworth, 1884) and is a period during which we are particularly sensitive to contextual influences that may have an exaggerated influence on our psychological development (Gopnik et al., 1999; Rapoport and Gogtay, 2008; Weimer et al., 2013). Perhaps a period of ‘microdosing’ with a psychedelic might be considered another valid example of a pivotal mental phase rather than a singular state per se (Kuypers et al., 2019) and, similarly, the at-risk mental state may express more as a protracted phase or process, rather than an entirely discrete state.
The PiMS model could also be applied to processes of ideological radicalization and deradicalization. According to one constructivist model of radicalization, self-invalidation and uncertainty lead to a creative reconstruing of the self, followed by a tightening of beliefs around a newfound identity (Feixas and Winter, 2019). This theory is consistent with the ‘Decentring’ model of religious experience (McNamara, 2009), the social defeat hypothesis of schizophrenia (Selten et al., 2016) and our proposal that psychological flexibility often gives way to psychological rigidity (e.g. delusional ideation) in the psychotic process, as in many psychopathologies. Drastic recalibrations of beliefs in response to interpersonal stresses and uncertainties are common features of psychotic, religious and radical conversion experiences.
Throughout this review, we have placed emphasis on the role of stress in increasing 5-HT2AR expression and signaling and cited evidence linking increased 5-HT2AR signaling with aspects of neuroplasticity. However, there is also evidence that stress can negatively impact neuronal integrity, particularly in the hippocampus (Conrad et al., 2017) and may also cause certain functional impairments (McEwen, 1999). This apparent inconsistency needs to be addressed, if not reconciled. Attempting this, one might propose that plasticity is not a singular homogenous phenomenon (Gray et al., 2013) and (like the PiMS itself) neither is it intrinsically salutogenic, that is, promoting of health (Doidge, 2007). Moreover, there is good evidence that stress can facilitate aspects of learning (Joëls et al., 2006) as is the case with trauma, for example (Deppermann et al., 2014; Joëls et al., 2006). Thus, it would be hasty to cite evidence of the negative impact of chronic stress on regionally specific neuronal integrity (e.g. McEwen, 1999) or aberrant synaptic plasticity in schizophrenia (Stephan et al., 2009) as evidence against the model being presented here. One should also note that we have specifically identified cortical plasticity as a key criterion for PiMSs.
One other point of critique is that our central construct, the PiMS, merely repackages previously introduced psychological constructs such as ‘quantum change’ (C’De Baca and Wilbourne, 2004; Miller and C’de Baca, 2001) and ‘transformative experiences’ (Paul, 2014). There is, however, surprisingly little scientific literature on the latter phenomenon and both constructs lay emphasis on change or outcome rather than the mediating state itself. Moreover, there is little-to-no discussion of the underlying neurobiology of quantum change and an arguably one-sided emphasis on positive outcomes (Miller and C’de Baca, 2001). Thus, we feel there is a great deal of scientific and pragmatic value in focusing more on the psychological and neurobiological nature and function of an outcome agnostic, yet (outcome) pivotal, mediating state. We do, however, encourage the reading of literature on quantum change in particular, as it provides a rich coverage of PiMS-relevant phenomenology, including the prevalence with which individuals report psychological crises prior to the onset of a PiMS/quantum change experience (Miller and C’de Baca, 2001). Some other potentially relevant constructs include: (a) the ‘healing crisis’ or Jarisch-Herxheimer reaction (Bryceson, 1976), namely, the principle that a period of heightened pain/suffering can be part of a healing process; (b) the ‘hero’s journey’, which is the notion that a process of overcoming existential struggle and suffering can be positively (trans)formative (Campbell, 2008); and (c) the notions of allostasis (McEwen, 2019; Sterling and Eyer, 1988) and adaptive plasticity and calibration (Ellis and Del Giudice, 2019).
Finally, we are mindful that a significant amount of validation work must now be done to better define the PiMSs construct as well its usefulness as a model. This process should include demonstrating what the PiMS is not – discriminate validity. Relatedly, although good evidence has been provided that the psychedelic state meets all three defining properties of a PiMS, the same evidence has not been collated for other candidate PiMSs. This must be addressed if the case for the fundamental nature of PiMSs is to be strengthened. One way we could model the natural occurrence of PiMS in humans would be to use stress-induction paradigms that are proven to reliably upregulate the 5-HT2AR system in non-human animals, such as sleep deprivation (Elmenhorst et al., 2012; Maple et al., 2015; Zhao et al., 2019) or CO2 inhalation-induced hypercapnia (Buchanan et al., 2015). A 5-HT2AR antagonist pre-treatment or combined EEG and positron emission tomography (PET)-fMRI technology could be utilized to examine the neurochemistry, neurophysiology and phenomenology of the relevant state. Thus, this review should be regarded as a first step in what may be a lengthy but ultimately fruitful validation process.
Conclusion
This paper has proposed that certain traumatic, spiritual, psychedelic and psychosis-relevant experiences can be viewed as examples of what we call PiMSs: transient, intense hyper-plastic states of mind and brain that have the potential to mediate rapid, major and potentially enduring psychological change. Various stressors have been found to upregulate the 5-HT2AR system and converging evidence implies that increased 5-HT2AR signaling may be a key molecular gateway through which PiMSs arise. The pharmacology, systems-level neurobiology and phenomenology of the (5-HT2AR agonist) psychedelic drug state is treated as particularly informative in this regard. PiMSs represent special opportunities for psychological transformation and we propose this may occur through the recalibration of mental schemata, consistent with a recent model of the therapeutic action of psychedelics (Carhart-Harris and Friston, 2019). We propose that 5-HT2AR-mediated PiMSs have evolved to aid major perspective shifts, when conditions demand them. The 5-HT2AR-mediated PiMS may represent a ‘last gasp’ attempt to resolve an intolerable situation, for example, after the expiration of mere tolerance (Carhart-Harris and Nutt, 2017). Suicide and psychosis are tragic but not inconsistent ‘solutions’ within this framework but positive, psychological ‘re-birth’ is another.
Before the relatively recent advent of secular psychology, religious traditions provided useful resources to prepare and guide individuals through PiMSs, for example, for purposes of self-transformation and growth. Note, in this regard, religions have excellent narrative and infrastructural resources for dealing with ‘psychological rebirth’ (Rambo, 1993). In modern secular environments, we presently lack the same quality of integrated support, shared values and unified vision. Thinking towards the future, consideration could be given to the value of creating humanistic contexts supportive of positive self-transformation. Consistent with principles of new-wave psychotherapies, including psychedelic therapy, one can envision an ideal future in which psychological crises are seen less as emergencies requiring immediate suppressive intervention and more as opportunities for development and growth, if, of course, appropriately supportive contexts can be provided for this.
Footnotes
Acknowledgements
Ari Brouwer would like to thank Wesley Wildman and Patrick McNamara for advising a master’s thesis in which some of these ideas were developed. Robin Carhart-Harris would like to thank Fernando Rosas for some useful input on an earlier form of the manuscript. Both authors would like to thank the reviewers for helpful critique. We would like to thank Pedro Oliveira, Gustavo Carreiro, and Hugo Raposo of Favo Studio for the illustrations in Figure 3 and Figure 4.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: RCH is supported by the Alexander Mosley Charitable Trust. The Centre for Psychedelic Research was founded by donations from Tim Ferriss, Ad Astra Chandaria, Nikean Foundation, and Alexander and Bohdana Tamas. It has since received additional support from Amanda Eilian, Brian Rose and Eric Grotesfeld.