
Abstract
Background:
According to alcohol myopia theory, alcohol reduces cognitive resources and restricts the drinker’s attention to only the more prominent aspects of a visual scene. As human hairstyles are often salient and serve as important facial recognition cues, we consider whether alcohol restricts attention to this region of faces upon initial viewing.
Aims:
Participants with higher breath alcohol concentrations just prior to encoding a series of unfamiliar faces were expected to be poorer than more sober counterparts at recognising the internal but not external features of those faces at test.
Methods:
Drinkers in a nearby bar (n=76) were breathalysed and then shown a sequence of 21 full face photos. After a filled five-minute retention interval they completed a facial recognition task requiring them to identify the full, internal or external region of each of these among a sequence of 21 previously unseen (part or whole) faces.
Results:
As predicted, higher breath concentrations were associated with poorer discrimination of internal but not external face regions.
Conclusions:
Our findings suggest that alcohol restricts unfamiliar face encoding by narrowing the scope of attention to the exterior region of unfamiliar faces. This has important implications for drunk eyewitness accuracy, though further investigation is needed to see if the effect is mediated by gender, hair length and face feature distinctiveness.
Alcohol (ethanol) is an extremely popular recreational drug strongly associated with increased rates of violent crime, including robbery, affray and sexual assault. Intoxicated victims and witnesses of such crimes are therefore widely encountered by law enforcement agencies yet understanding of the drug’s impact on their ability to later recognise perpetrators remains limited (Evans et al., 2009; Palmer et al., 2013). Currently only nine studies directly address the topic (Altman et al., 2018; Colloff and Flowe 2016; Dysart et al., 2002; Flowe et al., 2017; Hagsand et al., 2013; Harvey et al., 2013; Kneller and Harvey, 2016; Read et al., 1992; Yuille and Tollestrup, 1990) two of which report adverse effects of alcohol on performance at a forensically relevant face identification task (Dysart et al., 2002; Read et al., 1992).
Read et al.’s (1992; Experiment 2) participants committed a simulated robbery in a sober or alcohol intoxicated state then attempted to identify each of two individuals from a five-person simultaneous line-up, one a ‘bystander’ seen prior to the mock crime (low arousal condition) and the other an ‘intruder’ encountered during the crime (high arousal condition). Alcohol halved the number of correct ‘bystander’ identifications in the low arousal condition but had no effect on high arousal ‘intruder’ identifications. The authors interpret the low arousal effect as an alcohol-induced face encoding deficit that is negated when the target’s actions induce witness arousal.
In the second report of alcohol-based face memory impairments, Dysart et al. (2002) had participants attempt to identify (from a single photograph) the person who initially recruited them to the study as they were drinking in a local bar. The authors found a positive association between breath alcohol concentration and the likelihood of falsely identifying a foil similar in appearance to the study recruiter. They explain this finding in terms of alcohol myopia theory (AMT), the idea that rising blood alcohol levels reduce cognitive resources and restrict the drinker’s attentional scope to only the most salient or immediate scene features (Steele and Josephs, 1990). The human face is a prominent visual stimulus and encoding of its external features (particularly the hair) is known to play a crucial role in unfamiliar face recognition (Bruce et al., 1999; Ellis et al., 1979; Frowd et al., 2007; Johnston and Edmonds, 2009; Wright and Sladden, 2003; Young et al., 1985). Thus, as Dysart et al. have suggested, alcohol intoxication may narrow the scope of attention to this important facial region, leading to poorer encoding of the target’s internal face features. This was assumed to have made it harder for intoxicated participants to discriminate memories of the recruiter from a facial image of a different female with similarly distinctive hair. Firm evidence of an alcohol-encoding bias to the facial exterior would have important forensic implications as perpetrators often conceal, disguise or distort their hair at the scene (using hats, masks, hair ties, stockings, wigs, etc.) or cut, colour and restyle it after the event in attempts to evade capture. Such alterations significantly impair normal face recognition (Chan and Ryan, 2012; Patterson and Baddeley, 1977) but may pose an even greater challenge to drunken eyewitnesses.
In their use of live, interactive stimulus scenarios the applied studies of Dysart et al. and Read et al. have high ecological validity but reveal little about the face learning processes impaired by alcohol. To gain a deeper understanding of these we turn to more contrived experimental scenarios in which hypotheses concerning specific cognitive mechanisms were tested. A recent example is that of the study of Bayless et al. (2018), which examined the influence of alcohol on the construction of a computerised image (police composite) of a single unfamiliar target face encountered 24 h before. Participants who encoded the face following alcohol consumption rendered a poorer likeness of it the following day than sober controls. Separate likeness judgements for the internal and external region of the composites further revealed that alcohol impaired memory for the external features of shorthaired-male but not longhaired-female faces. This is consistent with the view that alcohol narrows the viewer’s focus of attention on to the external face region but possibly only when the hair is long or distinctively styled (Dysart et al., 2002).
The restricted face encoding view is endorsed by the study of Hilliar et al. (2010) in which intoxicated participants falsely identified more same-race faces than sober controls in the context of an old-new recognition task, which significantly reduced the size of the own-race face-processing bias for the alcohol group. The authors therefore suggest alcohol causes attentional deficits that disrupt the usually expert processing of same-race faces, though we note that Harvey (2014) failed to replicate this effect using a more sensitive within-subjects design.
Converging evidence that alcohol restricts face encoding comes from studies of the drug’s perceptual effects. Normal face learning relies on the extraction of holistic cues regarding the interrelation of features (e.g. distance separating the eyes, their position relative to the nose and mouth, etc.), which may be computed from the length of face-scanning saccades (Henderson et al., 2005). But alcohol is known to slow saccadic motion and restrict eye movements during scene perception, object tracking (smooth pursuit) and face encoding (Buser et al., 1996; Harvey, 2014; Holdstock and de Wit, 1999; Moser et al., 1998; Nawrot et al., 2003; Wilkinson, 1976). It also impairs the detection of bilateral asymmetry in faces, leading to inflated judgments of interpersonal attractiveness (Halsey et al., 2010; Oinonen and Sterniczuk, 2007; Souto et al., 2008). These findings are consistent with the notion that alcohol reduces the amount of configural information drinkers can extract from an unfamiliar face.
In the present study we examined the impact of acute alcohol intoxication on the encoding of a series of unfamiliar faces and their subsequent recognition in either whole or part form. To determine if alcohol narrows the focus of attention to external face regions during encoding, we presented participants with full faces for study but either full, internal or external face regions at test. Participants were required to state whether each whole or part face was one of those presented in the learning phase. We expected increases in breath alcohol concentration to be significantly associated with poorer discrimination accuracy for internal face features, but alcohol was not expected to impair recognition of the external face region as alcohol myopia is presumed to narrow the focus of attention on to this area during encoding. Nor did we expect alcohol to impair recognition of full faces, as hairstyles may be used as effective retrieval cues when attention is biased to this facial region during encoding.
A secondary aim of the study was to explore the relationship between alcohol consumption, part/whole face discrimination accuracy and the confidence participants expressed in their recognition judgements.
Method
Participants
Seventy-six patrons of the host university’s Student Union bar freely volunteered their time to participate (18 female, 58 male). Ages ranged from 18–24 years (M=20.51, standard deviation (SD)=1.63) and all participants reported normal or corrected-to-normal vision.
Design
In the face-learning phase participants were shown a series of 21 young white adult male faces in a single study block. At test different photographs of these same 21 (‘old’) faces were presented among a selection of 21 previously unseen (‘new’) faces of the same broad description. Face presentation order was randomised anew for each participant both at study and test. Dependent variables were the number of old faces correctly identified as old (hits), the number of new faces incorrectly identified as old (false alarms), recognition response time (ms), and self-confidence ratings of accuracy ranging from one (‘guess’) to nine (‘very sure’). Alcohol consumption was treated as both a binary quasi-independent variable (alcohol versus no-alcohol) and a continuous predictor of face recognition performance (blood alcohol concentration (BAC, % by vol.)). The host university’s ethics committee approved the study, which was administered with full adherence to the British Psychological Society Code of Ethics and Conduct.
Apparatus and materials
Measurements of (ethanol) alcohol in participants’ deep lung air were recorded using a Dräger Alcotest 3000 breathalyser (mg/100 mL) and converted to BAC estimates based on a 2300:1 blood-breath partition ratio.
All facial images were drawn from the Psychological Image Collection at Stirling (PICS) retrieved from http://pics.psych.stir.ac.uk/ (1 February 2018). Study faces were comprised of 21 young adult male faces presented in greyscale (neutral expression, frontal view) in the centre of a 15-inch laptop screen against a white background. All images were the same size (H=15 cm, W=12 cm) and faces showed no adornments, eyewear, headwear, scars, tattoos or other distinguishing features. Slightly different photographs of the same 21 encoding faces were presented at test, along with images of 21 new male faces meeting the same criteria. For the recognition test, faces were presented either in full view, or with only the internal or external region visible. This was accomplished by manipulating the face images using the Photoshop software package. Internal features (eyes, nose and mouth) were cut from faces using the elliptical and rectangular selection tools and then smoothed around the edges with the blur tool. External regions (hair, forehead, ears, chin and neck) were simply the inverse selection of the internal region and both parts were presented in the same size as at study (see Figure 1 for an example).
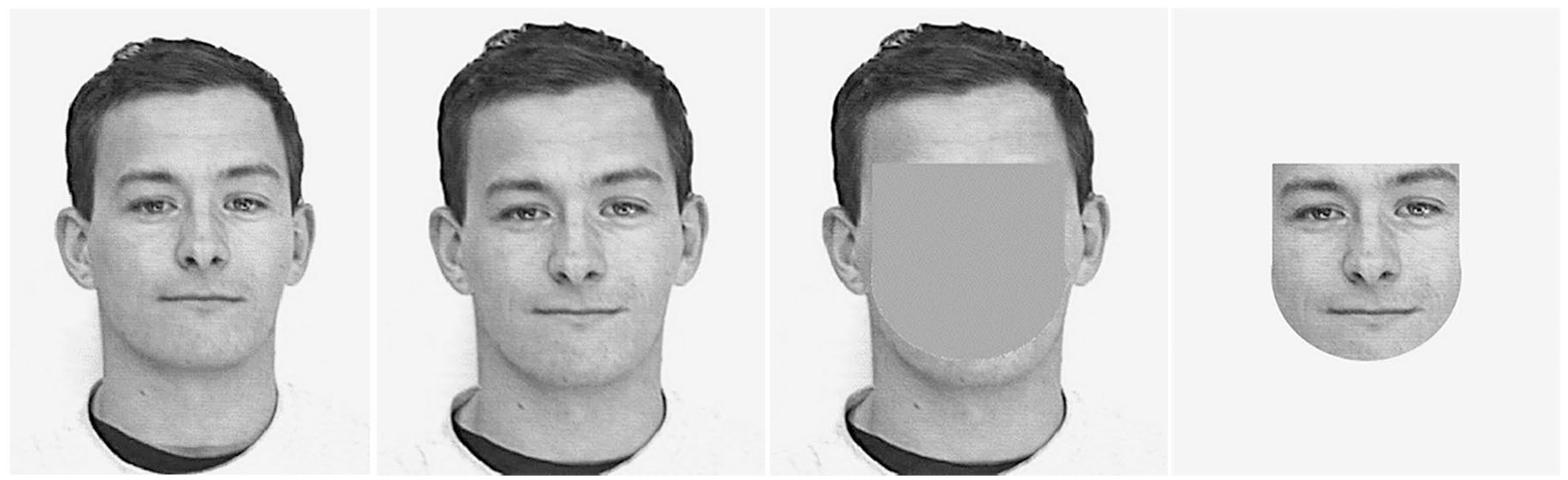
From left to right: example of a stimulus face presented at encoding then a different image of the same individual presented in full, external or internal form at test.
Participants completed a ‘Where’s Wally?’ visual search task for five minutes after face learning. This comprised a series of six illustrations depicting scores of characters engaged in a variety of novel and amusing activities in some themed location (e.g. a beach). Hidden within each of these densely detailed scenes was a character wearing a red and white striped hat and sweater named ‘Wally’, whom participants had to find.
Procedure
Participants were recruited through face-to-face requests in the host institution’s Student Union Bar. The experimenter did not approach drinkers showing obvious signs of extreme intoxication and only those who could clearly understand his request were invited to proceed to the briefing and consent stage. Participants were individually escorted to a quiet pre-booked seminar room one floor above the bar. They were seated at a table, handed an experiment information sheet to read and invited to ask questions about the study. After signing to consent, participants gave a breathalyser reading (not disclosed until debrief) followed by a subjective rating of their alcohol intoxication level, ranging from one (completely sober) to 10 (extremely intoxicated). The researcher then placed a laptop in front of the participant and commenced with the experimental task.
Participants were told they would be presented with a series of faces that they should try to remember for a test later. EPrime software was used to present each face in the centre of a white screen for three seconds separated by a one-second blank screen interval. They then engaged in a ‘Where’s Wally?’ visual search task for five minutes before completing the recognition test. They were told that the faces they had studied previously would be shown again but mixed among a series of new faces all presented in either part or whole form. Participants were instructed to press ‘o’ (for ‘old’) if they believed the face or face part was one shown previously or to press ‘n’ (for ‘new’) if not. They were also asked to use the numeric keypad to indicate confidence in each memory decision on a scale from one (‘guess’) to nine (‘very sure’). Each face remained visible until both a memory decision and confidence rating was recorded, at which point the next face was automatically displayed. The experimental task took around 15 min to complete. After the final response participants were debriefed and told their breath reading. They were then invited to ask questions, escorted back downstairs, thanked and urged not to discuss details of the study with other bar dwellers.
Results
Intoxication levels
Figure 2 shows the distribution of mean breath alcohol measures (averaged across the two breath tests) for participants who had consumed alcohol (n=42). BACs among this group ranged from 0.01–0.16% with a mean of 0.07% (SD=0.04) with all remaining participants recording a BAC of 0.00%. Objective breath alcohol measures were positively and strongly correlated with subjective intoxication ratings for the whole sample, r(76)=0.816, p<0.001, and just the alcohol group, r(42)=0.488, p=0.001.
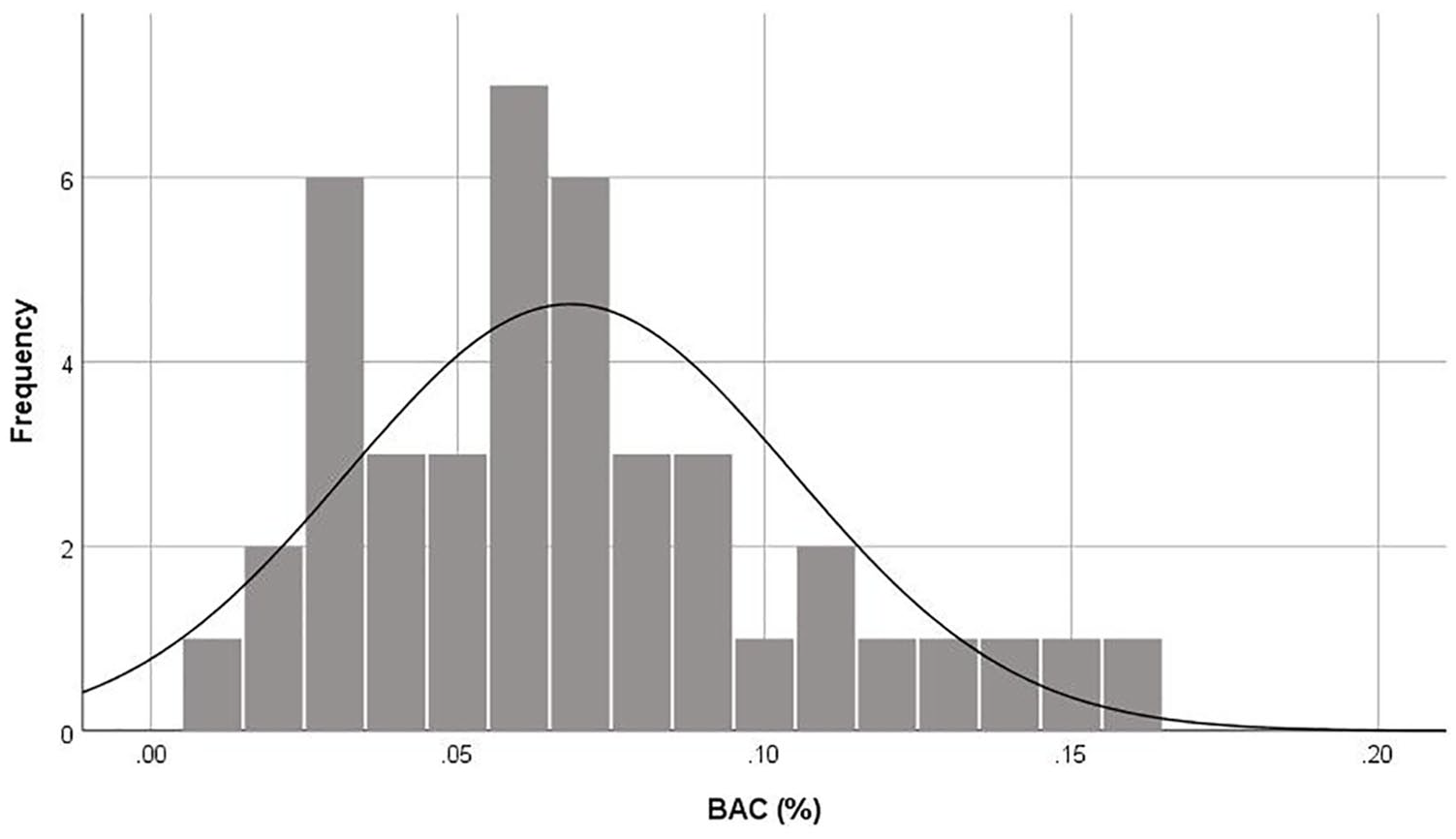
Distribution of blood alcohol concentrations (BACs) for participants who had consumed alcohol (n=42), with normal curve superimposed.
Part versus whole face recognition
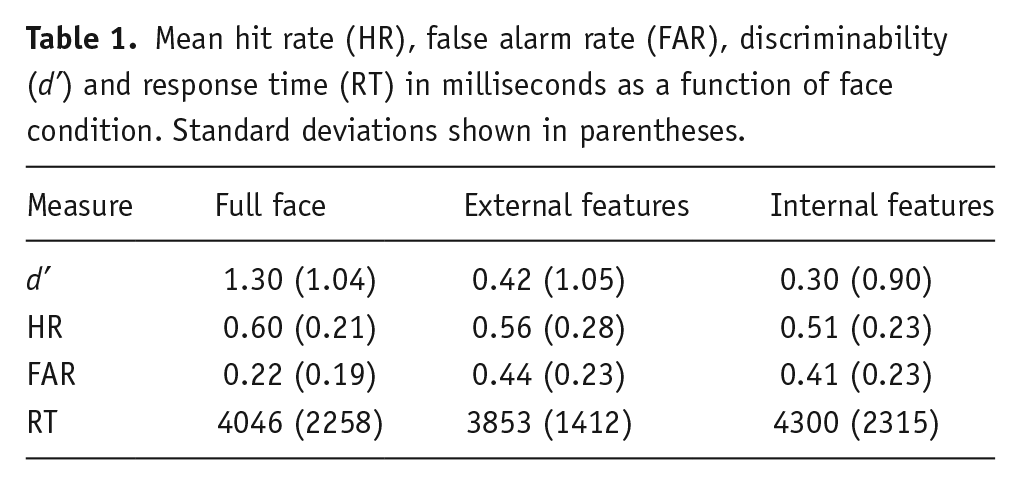
Face recognition performance was evaluated using d’, with higher scores reflecting an increased ability to discriminate old from new faces (for details see Stanislaw and Todorov, 1999). Mean d’ scores for the alcohol and no-alcohol group are shown in Table 1 as a function of internal, external and whole face recognition. Due to technical problems recognition decision and confidence data for eight participants were not recorded, so the following analysis was conducted on the remaining 68 cases.
Mean hit rate (HR), false alarm rate (FAR), discriminability (d’) and response time (RT) in milliseconds as a function of face condition. Standard deviations shown in parentheses.
A 2×3 mixed-design analysis of variance (ANOVA) was conducted on the d’ scores shown in Figure 3, with alcohol consumption (alcohol vs no alcohol) serving as the between-groups, and face region (full vs internal vs external) the within-groups, variable. This revealed a significant main effect of face region, F(2,122)=18.53, mean standard error (MSE)=1.17, p<0.001, ηp2=0.22 (with Huynh-Feldt correction for non-sphericity), confirming that participants were significantly better at discriminating full faces (M=1.29, 95% confidence interval (CI) (1.04–1.55)) than only external (M=0.41, 95% CI (0.16–0.67)) or internal (M=0.30, 95% CI (0.09–0.52)) face regions, with Bonferroni corrected post-hoc comparisons confirming the difference between external and internal conditions to be non-significant (p>0.1). The main effect of alcohol, F(1,66)=1.87, MSE=0.86, p=0.176, ηp2=0.03, and the face region by alcohol interaction were also non-significant, F(2,122)=0.72, MSE=1.17, p=0.48, ηp2=0.01 (Huynh-Feldt corrected).
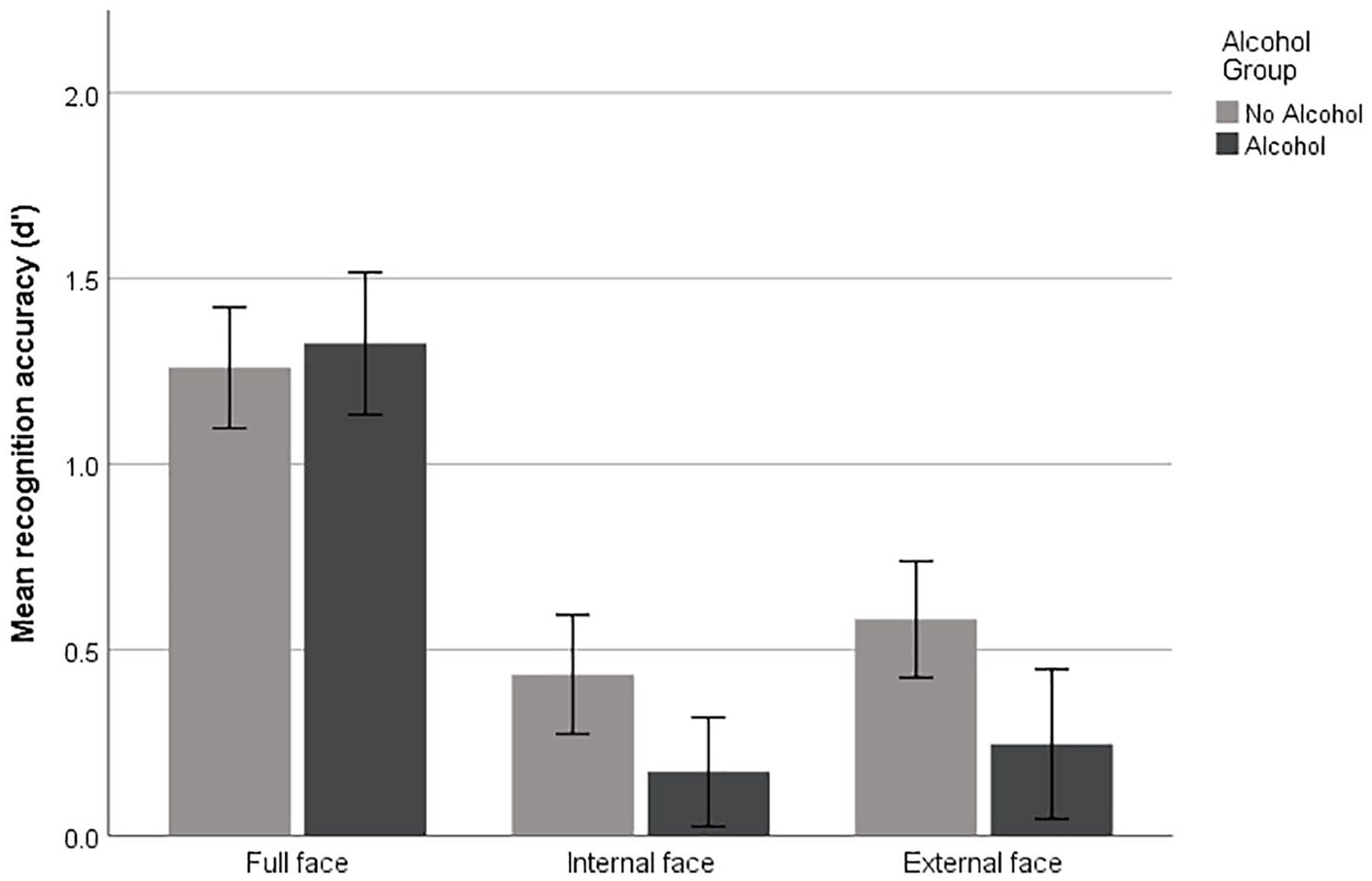
Mean recognition accuracy (d’) as a function of face condition and alcohol group with standard error bars.
A 2×3 ANOVA on the mean response time data revealed alcohol participants (M=3316.63, SD=1265.14) were significantly faster than sober counterparts (M=4389.68, SD=1793.32) at making discrimination decisions, F(1,66)=11.11, MSE=5,280,165.15, p=0.001, ηp2=0.14. However, the main effect of face region on response time (p=0.63) and the interaction between face region and alcohol (p=0.51) were non-significant.
Alcohol (BAC) as a predictor of face recognition performance
As the range of positive breath alcohol measures was wide (0.01–0.16%), with some participants producing readings close to zero (see Figure 2), we used BAC as a continuous predictor of recognition performance (d’), rather than as a quasi-independent categorical variable (alcohol versus no-alcohol). As expected, correlations between BAC and d’ for full face, r(68)=−0.106, p=0.195 (one-tailed) and external face recognition conditions, r(68)=−0.107, p=0.191 (one-tailed) were weak. However, the negative relationship between BAC and d’ for internal face discriminations (displayed in Figure 4) was notably stronger and statistically significant, r(68)=−0.237, p=0.026 (one-tailed).
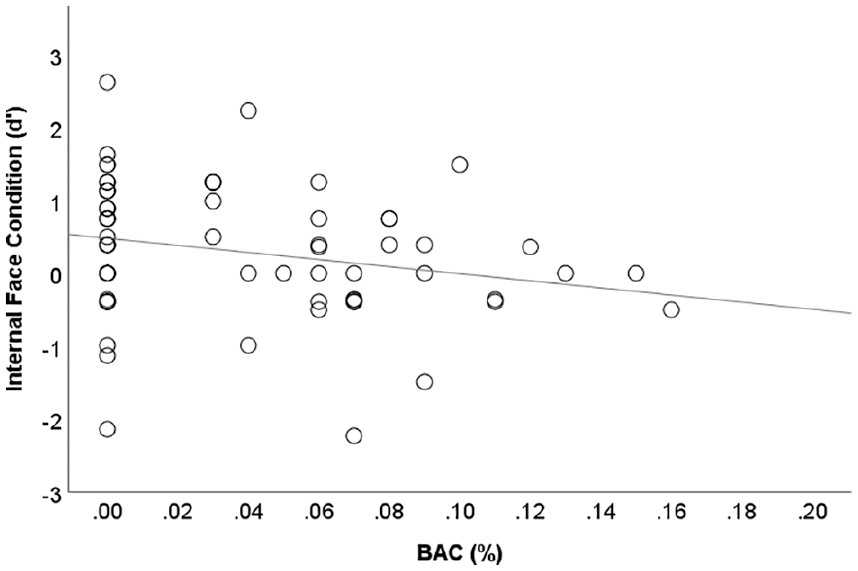
Relationship between breath alcohol concentration (BAC) and internal face discrimination (d’) accuracy (r=−0.24) with best fitting linear function.
Simple regression analyses revealed rising BACs to be significantly predictive of poorer internal face discrimination (d’) across all participants, β=−0.24, F(1,67)=3.93, p=0.05 (one-tailed), r2=0.06, and just those with a positive breath alcohol reading, β=−0.31, F(1,34)=3.58, p=0.03 (one-tailed), r2=0.10.
Face recognition confidence
We ran a 3(Face region)×2(Alcohol group) mixed-design ANOVA to explore changes in face recognition confidence across groups. This revealed a significant main effect of face region, F(2,132)=49.99, MSE=0.42, p<0.001, ηp2=0.43, with post-hoc Bonferroni corrected comparisons confirming that mean confidence ratings for full face recognition decisions (M=6.96, SD=1.12) were significantly higher than confidence ratings for external face recognition decisions (M=6.40, SD=1.15), which in turn were significantly higher than confidence ratings for internal face recognition decisions (M=5.84, SD=1.31) (all p’s<0.001). However, confidence ratings did not vary as a function of alcohol group, F(1,66)=0.21, MSE=3.48, p=0.65, ηp2=0.003, and the interaction between face region and alcohol group was also non-significant, F(2,132)=0.79, MSE=0.42, p=0.46, ηp2=0.012.
Confidence-based receiver-operating characteristic (ROC) curves
In order to explore the effects of alcohol on face discrimination confidence and accuracy we constructed ROC curves for each of the three face recognition conditions. Figure 5 shows the rate of hits (correct identification of ‘old’ faces) and false alarms (‘new’ faces incorrectly identified as ‘old’) for alcohol and non-alcohol drinkers plotted as a function of face condition and response confidence. For each drinking group, the coordinate point in the upper right of each panel represents recognition responses made across all levels of confidence (1–9). The next leftward coordinate represents responses made at confidence levels 2–9, the next point at levels 3–9, and so on, until the final point, which includes responses made with only the highest confidence (level 9). It is important to note that while hit rates at the leftmost positions seem low, the so-called diagnosticity ratios (HR: FAR) they represent are relatively high, as they exclude all lower-confidence recognition decisions (Gronlund et al., 2014). Another useful feature of ROC analysis is that face discriminations at each confidence level may be compared to chance performance, the broken line in Figure 5, representing equality between hit and false alarm rates.
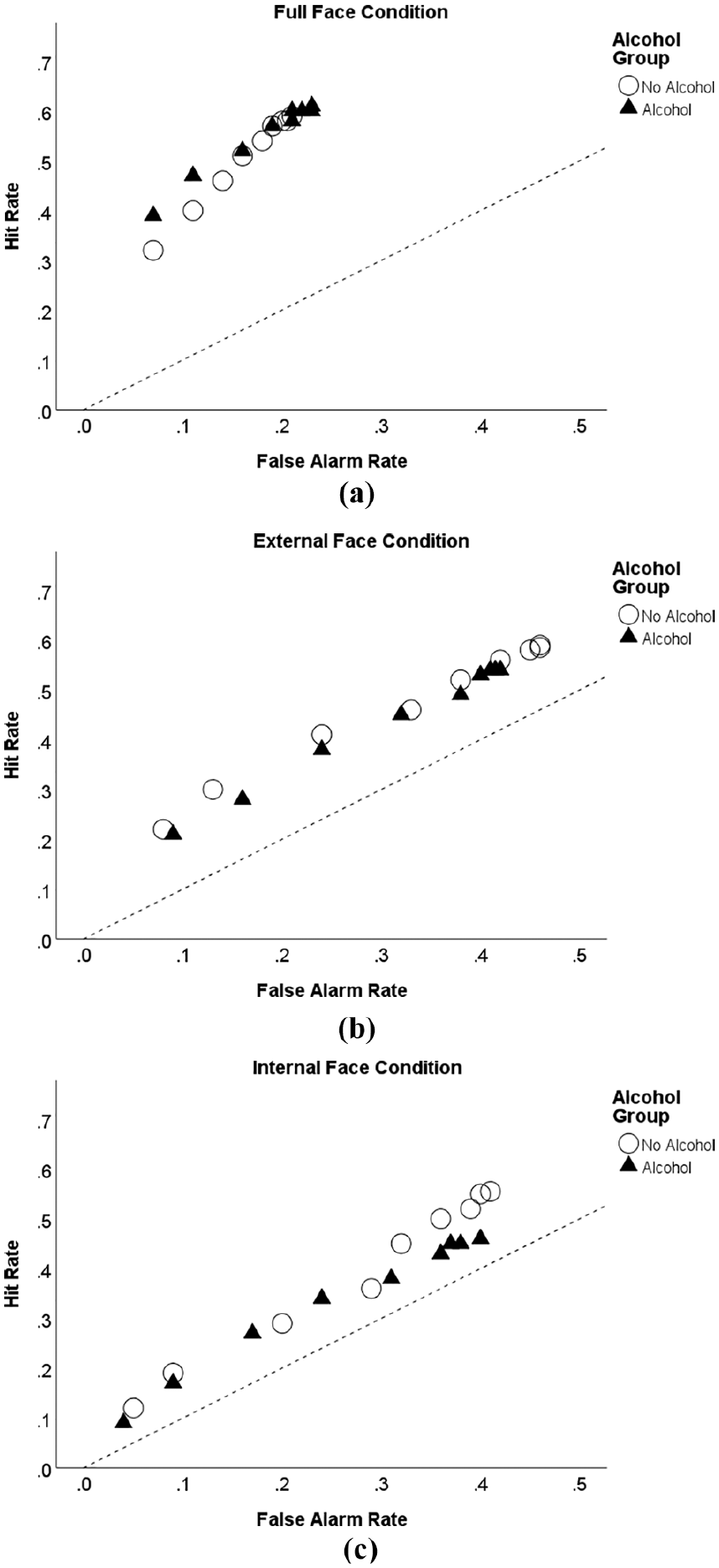
Confidence-based receiver-operating characteristic (ROC) curves for alcohol and no-alcohol groups under (a) full, (b) external and (c) internal face recognition conditions.
It is clear that face discrimination was most accurate for full faces as ROC curves in this condition show low false-alarm rates and sit further above the chance line than curves in the external (mid panel) and internal (lower panel) face conditions (see Figure 5, upper panel). Interestingly, full face discriminations made at the two highest confidence levels were better for the alcohol than no-alcohol group. Discrimination rates were closer to chance performance in the external face condition (mid panel), which shows higher false alarm rates and, importantly, little difference between groups. Accuracy was even closer to chance in the internal face condition (lower panel) though, consistent with predictions, alcohol participants show lower hit rates than no-alcohol counterparts, but only for recognition decisions made with lower confidence.
Discussion
We examined the effects of acute alcohol intoxication on visual attention and face discrimination within the context of an old-new recognition task. Participants studied full faces and then had to recognise either the full, internal or external regions of these faces at test. This design allowed us to examine the claim that alcohol restricts the focus of attention to the external face (contour, hair) region during encoding leading to poorer discrimination of internal face features. As expected, breath alcohol increases were significantly predictive of poorer internal- but not external- or full-face recognition performance. The fact that alcohol did not impair full-face discrimination is consistent with the many previously reported null effects of alcohol on identification performance (Altman et al., 2018; Colloff and Flowe, 2016; Flowe et al., 2017; Hagsand et al., 2013; Harvey et al., 2013; Kneller and Harvey, 2016; Yuille and Tollestrup, 1990). In fact the present findings suggest that some intoxicated viewers with high confidence in their memories of unfamiliar faces may even outperform sober counterparts (see Figure 3, top panel). Alternatively, this effect may reflect more conservatively calibrated confidence among the alcohol group. Our results are also consistent with Dysart et al.’s suggestion that alcohol narrows the focus of attention during encoding to the external region of unfamiliar faces and help explain why the majority of past alcohol challenge studies show no adverse effects on face identification performance.
An encoding bias to the facial exterior should increase the risk of misidentification only when a new face with similarly styled hair happens to interfere at test. If a suspect’s hairstyle remains unchanged when paraded in a line-up, witnesses that were intoxicated at the scene should fare no worse at an identification task than sober counterparts. However, our findings suggest that perpetrators who disguise their hair at the scene or change its style after the event are less likely to be recognised by alcohol intoxicated witnesses as they are too dependent on the external face cues encoded at the scene for an accurate face match. The drunk witness’s poorer memory of the perpetrator’s internal face features therefore increases their risk of falsely identifying an innocent line-up filler whose hairstyle happens to match that of the perpetrator’s at the scene. The extent to which this encoding bias is mediated by distinctiveness of the external face region is unclear. One possibility is that longer or more salient (e.g. unusually coloured or styled) hair increases the distractibility of external face features among alcohol drinkers, meaning the effect might be smaller for faces with shorter less distinctive hair. As Bayless et al. (2018) suggest, when encoding new male faces with typically cropped unremarkable hairstyles, the narrowed attentional focus of alcohol-intoxicated viewers may be more biased to the central face region.
As female hairstyles tend to be longer and more diverse than male hairstyles, especially among younger age groups (Hinsz et al., 2001), we might therefore expect alcohol impairments in unfamiliar face processing tasks to vary along gender lines. Gender-related hair effects in a normal (sober) sample were observed by Kemp et al. (1997), who explored the utility of placing the account holder’s facial image on credit cards to improve financial security. They found experienced cashiers were surprisingly poor at face matching, showing an overall failure rate for detecting fraudulent card use of around 50%, but the detection rate was significantly lower for female than male shoppers. Kemp et al. attribute this difference to a wider range of hairstyles among female shoppers relative to male counterparts, which they suggest distract cashiers causing them to overlook internal face cues crucial for accurate face matching.
The extent to which alcohol might influence face gender effects has not been directly addressed, however, Monds et al. (2019) recently observed a sober face processing advantage for the discrimination of male (Alcohol, d’=0.63; No alcohol, d’=0.81) but not female faces (Alcohol, d’=0.55; Controls, d’=0.57). In a footnote the authors state this interaction was statistically non-significant and of no theoretical relevance, but it may be evidence of alcohol myopia narrowing attention on to a specific facial feature. It is unclear why Monds et al. found an alcohol-linked face recognition deficit for male rather than female faces but the possibility that their findings reflect a more partial form of face encoding under alcohol is intriguing.
A similarly puzzling pattern emerged in the data of an alcohol and face recognition experiment by Harvey (2014), in which the eye-movements of participants were tracked as they encoded a series of male faces. As with the present study, alcohol had no influence on full-face recognition performance but the alcohol group’s gaze fixations were significantly more clustered around the central nose region of study faces than those of sober controls. We can only speculate as to why the nose area rather than the hair region was the focus of the alcohol group’s gaze in this earlier study but the first important point to note is that shifts in eye movements and shifts in attention can dissociate such that eye fixations to a particular stimulus feature do not guarantee the viewer is necessarily attending to that feature (Posner, 1980). But if we assume that eye fixations are a reliable proxy for the focus of overt attention, then one possibility is that Harvey’s (2014) stimulus faces had a less diverse range of hairstyles than those of the present study. This would reduce the likelihood of alcohol participants being distracted by the hairstyle of each stimulus face. To test this suggestion the present study should be replicated with an extended design that includes a hair-distinctiveness manipulation and measures of gaze to internal and external features during face encoding. Future experiments should also explore the effects of alcohol consumption on face recognition accuracy as a function of target face gender.
Under normal (i.e. sober) learning conditions, unfamiliar faces are encoded holistically, with individual features (eyes, nose, mouth, etc.) and their overall configuration being integrated into a unitary memory (Tanaka and Simonyi, 2016). This form of processing is assumed to dominate face learning because previously studied face features are harder to recognise in isolation than when presented in a full-face context, suggesting memories of each feature are embedded in a unitary memory of the whole face (Tanaka and Farah, 1993). But an important question raised by the present study is whether alcohol disrupts holistic processing, making the recognition performance of drinkers more dependent on feature-based face processing. This hypothesis can be tested by comparing sober and intoxicated performance on the unfamiliar face composite task (Hole, 1994; Young et al., 1984). This requires participants to identify matches between the upper halves of a series of face pairs in which the lower half is either from the same face or seamlessly fused with that of a different face. The dominance of holistic processing is evidenced by a weakened ability to perceive matches when upper face halves are paired with lower halves from different faces, but superior matching when the upper and lower face halves are horizontally misaligned, a manipulation presumed to disrupt holistic processing (Le Grand et al., 2004). If alcohol consumption impairs holistic face processing then intoxicated participants ought to outperform sober controls at the face composite task when mixed face halves are aligned, as interference from configural and holistic cues should be reduced under its influence. Experiments along these lines should reveal if intoxication causes a form of local face processing in which visual attention is biased towards the encoding of perceptually salient face features at the expense of global face processing.
Another potentially fruitful avenue for future research is to explore the impact of acute alcohol intoxication on the recognition of familiar faces, the processing of which is known to be more dependent on encoding of internal features (Ellis et al., 1979; Johnston and Edmonds, 2009; Young et al., 1985), particularly the eyes (O’Donnell and Bruce, 2001). If alcohol does disrupt configural processing through restriction of encoding to external face features, then it should also impair recognition of familiar faces.
We accept the present field study is limited by its quasi-experimental design for which participants were not randomly assigned to alcohol and no-alcohol beverage conditions, and its short memory interval, which meant participants both encoded and retrieved faces under the influence of alcohol. We also have no record of participants’ alcohol drinking history or concomitant drug use at or prior to the time of test. Nevertheless, we demonstrate alcohol impairments to mechanisms of face recognition of considerable forensic significance. Our findings provide the first direct evidence that alcohol narrows the scope of attention in face learning to the external hair region of unfamiliar faces. While alcohol did not harm full face recognition performance, consistent with numerous other studies (e.g. Colloff and Flowe, 2016; Flowe et al., 2017; Hagsand et al., 2013; Harvey, 2014; Harvey et al., 2013; Kneller and Harvey, 2016), any weakened ability to identify internal face features places drunk witnesses at greater risk of falsely identifying innocent suspects whose hairstyle, as suggested by Dysart and colleagues, happens to match that of the perpetrator they attempt to identify.
Footnotes
Acknowledgements
The authors are grateful to Paul Waby for programming assistance.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.