
Abstract
Background:
Animal literature suggests an interaction between acetylcholine and serotonin on cognitive functions.
Aims:
The aim of the current study was to assess whether both neurotransmitters interact during memory and novelty processing in humans.
Methods:
We tested the interaction between acetylcholine and serotonin on cognitive functions in healthy volunteers by means of treatment with rivastigmine and citalopram, respectively.
Results:
The main result of the study showed that during the verbal learning task participants significantly recalled fewer words after citalopram treatment than after rivastigmine or placebo during both the immediate and delayed recall tasks. Rivastigmine was not able to reverse the impairing effect of citalopram.
Conclusions:
This finding is in line with previous studies in which we manipulated acetylcholine and serotonin in different manners. Taken together, these studies in humans do not support the notion from animal studies that these two neurotransmitters interact on cognitive functions.
Introduction
In our daily life various aspects of memory are an essential asset for many activities. Declarative memory shapes our mind as it reflects our personal life and experiences. There are many neurochemical processes that underlie declarative memory. These can be characterised by specific neurotransmitters that are involved in memory, such as acetylcholine (ACh), serotonin (5-HT), dopamine, γ-aminobutyric acid (GABA) and glutamate. Neuropsychiatric disorders characterised by memory impairments usually involve multiple neurotransmitter disturbances. Although it is well known that this is the case, most experimental psychopharmacological studies have investigated the function of a single neurotransmitter in memory functions. This may give a limited view on the complexity of the biological functions underlying memory functions.
The cholinergic hypothesis (Bartus et al., 1982) states that an imbalance of ACh underlies the memory loss and related cognitive problems in aging and dementia. Following this hypothesis, cholinesterase inhibitors have shown to be able to improve cognitive function, especially memory performance in Alzheimer’s disease patients (for a review, see Birks, 2006). Alternatively, memory deficits can be induced by blocking ACh in the brains of young healthy participants by means of, for instance, the M1 muscarinic antagonist biperiden (Sambeth et al., 2015) or the nonspecific muscarinic antagonist scopolamine (Broks et al., 1988). Although these findings support the cholinergic hypothesis of memory function, it is presently acknowledged that other neurotransmitters also play a role in memory (e.g. Carreiras and Marco, 2004). The very moderate efficacy of cholinesterase inhibitors on memory performance in Alzheimer’s disease may be related to the fact that it only one transmitter system is targeted. Nevertheless, manipulating the ACh system increases our knowledge about the role of ACh in memory.
Although 5-HT is generally considered as neurotransmitter that is relevant for affective disorders (Maes and Meltzer, 1995), there are also a large number of studies indicating that 5-HT is involved in memory processing. For instance, acute tryptophan depletion (ATD), a method to lower central 5-HT, has been found to impair declarative memory (e.g. Borghans et al., 2017; Riedel et al., 1999; Schmitt et al., 2000). Also, various human studies have shown a positive correlation between 5-HT1a receptors and memory (Glikmann-Johnston et al., 2015; Penttila et al., 2016). In animal studies a clear relationship between serotonergic markers and memory has been described (Meneses, 2017).
Computational models developed to describe memory have indicated the important role of both ACh (Meeter et al., 2004; Sherman et al., 2003) and 5-HT (Meeter et al., 2006) in different stages of memory processing. For instance, the ACh model suggests that a moderate increase in ACh release will facilitate learning and any decrease will impair learning by affecting the encoding process (Meeter et al., 2004). On the other hand, both 5-HT depletion and specific 5-HT agonists lower memory performance, likely by affecting memory consolidation (Meeter et al., 2006). However, previous literature has also reported that citalopram, a 5-HT reuptake inhibitor that increases 5-HT activity, improved memory (Harmer et al., 2002).
Previous research in animals (Cassel and Jeltsch, 1995; Steckler and Sahgal, 1995) and humans (Garcia-Alloza et al., 2005; Little et al., 1995) has suggested that ACh and 5-HT interact in their effect on cognitive functions. However, the exact nature of the ACh and 5-HT interaction is still not clear, nor is the interaction between these two neurotransmitters on declarative memory and novelty processing. Previous data from our group (Borghans et al., 2017; Caldenhove et al., 2017) has shown that a combination of biperiden and ATD has no interaction effects. Thus, the memory impairment was similar to the impairment found after biperiden or ATD alone and does not suggest an interaction between ACh and 5-HT in cognitive functions. This conclusion is in line with another study showing no interaction between ACh and 5-HT after co-administration with biperiden and citalopram (Sambeth et al., 2015). Of note, the manipulations of ACh and 5-HT are of a different nature (e.g. receptor vs global effect), which may explain the lack of interaction.
In the current study, we were interested in whether we could find an interaction between ACh and 5-HT by using two global manipulations that both increase the levels of these two neurotransmitters. Rivastigmine is a cholinesterase inhibitor that increases the levels of ACh in the brain (Farlow, 2003; Jann et al., 2002). It has been shown to be able to improve memory performance in Alzheimer’s disease patients (for a review, see Birks, 2006). Citalopram increases the levels of 5-HT in the brain and has been shown to be able to reverse episodic memory deficits in patients with major depression (Herrera-Guzman et al., 2009). These two compounds could be used to evaluate our hypothesis as to whether increased levels of ACh and 5-HT could have interactive effects on memory.
Imaging methods such as electroencephalography (EEG) provide valuable information about the different memory stages when used in conjunction with controlled acute pharmacological interventions (Blokland et al., 2015). The main aim of the present study was to assess in what way increased levels of ACh and 5-HT interact during memory and novelty processing. To this extent we increased brain levels of both neurotransmitters in healthy volunteers by means of treatment with rivastigmine and citalopram, respectively. Subsequently, participants performed behavioural tasks while EEG signals were recorded.
Experimental procedures
Participants
All experimental procedures were approved by the independent Ethics Committee of Maastricht University and the Academic Hospital Maastricht (The Netherlands). The study was conducted according to the code of ethics on human experimentation established by the Declaration of Helsinki (1964) and amended in Edinburgh (2000) and Seoul (2008) in accordance with the Medical Research Involving Human Subjects Act (WMO [in Dutch]). In total 21 participants (age range 18–30 years) were recruited via flyers and email advertisements at Maastricht University. Participants had to be willing to sign an informed consent form and were paid for their participation. The subjects’ physical and mental health was checked by a physician by means of a standard medical questionnaire, including psychological and psychiatric evaluation, and a medical examination. Subjects were excluded if they suffered from or had a history of cardiac, hepatic, renal, pulmonary, neurological, gastrointestinal, haematological or psychiatric illness. Other exclusion criteria were excessive drinking (>20 glasses of alcohol-containing beverages a week, where glasses of alcohol refer to the typical glasses used for each beverage in the Netherlands: this is for beer (5% alcohol) 250 mL, wine (12% alcohol) 100 mL and hard liquor (35% alcohol) 35 mL, all of which translate to 10 g of pure alcohol per glass), pregnancy or lactation, use of medication other than oral contraceptives, use of recreational drugs from two weeks before until the end of the experiment, and any sensory or motor deficit which could reasonably be expected to affect test performance. Participants who had a first-degree relative with a (history of) psychiatric disorder were excluded as well. The participants could leave the study at any given time without any consequences. All participants provided the written informed consent after receiving a complete description of the study.
Three participants decided to quit participation before the end of the study either due to personal reasons or due to adverse effects after drug intake (i.e. nausea after rivastigmine intake). A total of 18 subjects were included in the final study and analyses. Due to technical failure, the rivastigmine condition was not fully recorded for two participants and omitted from analyses; the other conditions of these participants were included in the analyses. Technical failure occurred halfway through testing for one participant and three-quarters of the way through testing for another participant resulting in n=18, n=17 or n=16 for the different behavioural tests.
Design
The study was conducted according to a double-blind, placebo-controlled, four-way cross-over design. The treatment order (rivastigmine, citalopram, a combination, and a placebo) was balanced over the four test days and separated by a washout period of at least seven days. The balancing of the treatment order was accomplished by counterbalancing. The design used in the current study has been validated in previous studies and has shown to be sensitive to both memory improvements as well as memory impairments of different drug systems (e.g. Borghans et al., 2017; Van Duinen et al., 2017). Rivastigmine and citalopram were distributed over capsules with lactose monohydrate as the principal constituent. The placebo capsules only contained lactose monohydrate in an equivalent amount and the appearance was identical to the treatment capsules. The capsules were manufactured, blinded and labelled by Basic Pharma Technologies BV (Geleen, the Netherlands) according to GMP regulations.
Treatment
Rivastigmine tartrate (Exelon, 3 mg) is a reversible cholinesterase inhibitor, which causes increments in the concentration of ACh, thereby enhancing cholinergic function. Peak plasma concentrations are reached around 50–70 min and the half-life is around 60–90 min. The duration of cholinesterase inhibition is suggested to be around 8–10 h.
Citalopram hydrobromide (Cipramil, 20 mg) is a 5-HT reuptake inhibitor, which increases the extracellular 5-HT levels in the brain. It does so by inhibiting the reuptake of 5-HT into the presynaptic neuron. Citalopram peaks in the plasma about four hours after a single dose and it has a terminal half-life of around 33–35 h.
The doses chosen in this study are within the range of the therapeutically recommended doses for both rivastigmine (6–12 mg) and citalopram (20–30 mg).
Procedures
Participants arrived at the laboratory in the morning. After arrival, the first treatment capsule containing citalopram or placebo was ingested orally with 180 mL of tap water in the presence of the investigator. The second capsule containing rivastigmine or placebo was taken 120 min after the first capsule that contained citalopram or placebo. After treatment administration, volunteers were prepared for the EEG recordings. Subsequently, 185 min after citalopram or placebo intake, they performed the immediate recall of the verbal learning task (VLT), followed by an immediate recognition of the spatial memory task (SMT), and a choice reaction time (CRT) test. After the CRT test, delayed recall and recognition of the VLT was performed (the verbal recognition task (VRT)), which was followed by a delayed recognition of the spatial memory test. Finally, the novelty oddball task was conducted. The total duration of testing was around 1.5 h.
EEG recordings
An EEG cap was used to place a set of 32 EEG electrodes according to the international 10–20 system (Klem et al., 1999). A reference and a ground were placed at the left mastoid and at the forehead, respectively. Eye movements were detected by horizontal and vertical electro-oculogram (EOG) recordings. Before electrode placement, positions were cleaned with alcohol and slightly scrubbed with a gel to provide good measurement. Both EEG and EOG were filtered between 0.01–100 Hz and sampled at 1000 Hz. Offline, the EEG was checked for EOG activity and other artifacts. The EEG that contained artifacts was excluded from analysis. Eye movements were filtered using an ocular correction method. Event-related potentials (ERPs) were extracted by averaging the responses within an epoch of 100 ms before and 1000 ms after stimulus onset.
VLT
The VLT is an adapted version of the Rey auditory VLT (Lezak, 1995), which assesses short- and long-term memory function for verbal information. The test, modified by Riedel and colleagues (Riedel et al., 1999), was developed to maximise the possibility of measuring enhancement rather than impairment only, by means of prolonging the list. The test consists of a list of 30 monosyllabic words (18 nouns and 12 adjectives) in Dutch. The words are shown on a computer screen for one second. Three trials with the same item sequence are presented. Each trial ends with a free recall of the words (immediate recall). Thirty minutes after the third trial, the participant is asked to recall as many words as possible (delayed recall). Subsequently, a recognition test (VRT) is presented, consisting of all former words and 30 new but comparable words (distracters). The words are shown on a computer screen for two seconds and participants are asked to rate whether they were presented in the learning trials by a ‘yes/no’ response. The inter-word interval is two seconds. The dependent behavioural measures are the number of words recalled during each of the immediate recall trials separately. This is a measure of short-term verbal memory learning and retrieval. The number of recalled trials 30 min after learning constitutes the delayed recall score, the long-term verbal memory retrieval. The number of correctly recognised items and reaction times (in ms) were recorded in the recognition phase, which measures the long-term verbal memory encoding.
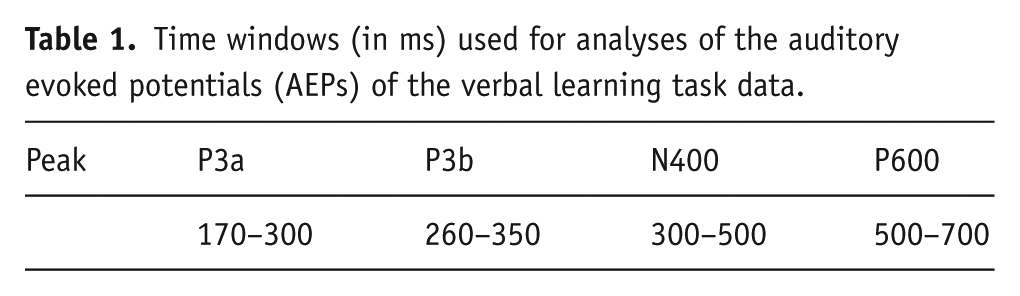
Measures taken from the EEG were as follows: ERPs were calculated during the encoding of the words for the three immediate recall trials separately. The ERP components analysed were P3a, P3b, N400 and P600 (Table 1). The Fz, Cz and Pz electrodes were analysed for the VLT (Borghans et al., 2017; Van Duinen et al., 2017).
Time windows (in ms) used for analyses of the auditory evoked potentials (AEPs) of the verbal learning task data.
SMT
The SMT assesses spatial memory and is based on the object relocation task by Postma and colleagues (e.g. Kessels et al., 1999). It consisted of an immediate and a delayed condition. In the immediate condition, a set of 10 pictures was presented one by one on different locations within a white square on the screen. This was followed by a ‘relocation’ part, which consisted of the presentation of a picture in the middle of the screen, followed by a ‘1’ and a ‘2’ being presented on two different locations. The participants’ task was to decide where the picture was originally presented, in location ‘1’ or location ‘2’. After relocation, which was done by a button press, the next picture was presented followed by the ‘1/2’ choice option. This continued until all 10 pictures had been relocated. After that, the next set of 10 pictures was presented. A total of six sets of 10 pictures were used. All pictures were every day, easy-to-name objects, presented in grayscale (±3.5×5 cm). Each picture was presented for 2000 ms with an interstimulus interval of 1000 ms. The ‘1’ and ‘2’ remained on the screen until the participant made the response. Outcome variables were the number of correct relocations, as well as the reaction time of relocating.
CRT task
The CRT task assesses general alertness, motor speed and attention, and was used to assess whether the drugs administered in the current experiment affected attention/vigilance. Participants were presented with an arrow that was either shown on the left or on the right side of the screen. There were two kinds of arrows: one pointing to the left (left arrow), the other pointing to the right (right arrow). Arrows on the left always pointed to the left, while arrows on the right always pointed to the right (only congruent trials). If the participant saw the left arrow, he/she had to press a button with the right index finger. If the right arrow is presented, the participant had to press a button with the right ring finger. The duration of stimulus presentation was 600 ms, followed by an interstimulus interval of randomly 2000–3000 ms. Outcome measures were number of correct button presses and reaction time (separate for left and right sides).
Novelty oddball paradigm
The novelty oddball task assesses involuntary attention processes which have been described elsewhere (Sambeth et al., 2006). In short, it is a passive paradigm, in which three types of auditory stimuli are presented while the participant watches a silent movie/cartoon and ignores the stimulation. The stimuli consisted of frequent standard, infrequent deviant and infrequent novel stimuli. The standard and deviant stimuli were 500 Hz and 750 Hz tones with two upper harmonic components (1000 and 1500, 1500 and 2250 Hz, respectively). The intensity of the first and second harmonic components was decreased compared to the fundamental by 3 and 6 dB, respectively. The use of those stimuli was counterbalanced between participants, but remained constant for the different measurements within subjects. Novel stimuli consisted of three stimulus categories of 20 different sounds, namely animal, human, and mechanical sounds. The deviant and novel stimuli were each presented in 12.5% of the trials. All sounds had a duration of 300 ms with 10 ms rise and fall times and were presented with a 1000 ms stimulus onset asynchrony and equal intensities to both ears using a headphone.
No behavioural measures were recorded, only ERPs. From the ERPs, the P50, N100 and P200 were analysed (Table 2). Additionally, the response to the standard stimuli was subtracted from the deviant and novel stimuli, which enables the visualization of the mismatch negativity (MMN) and P3a components. The amplitudes and latencies of these components were compared between the deviant-standard and novel-standard responses. The P3a component measures the involuntary switch to novel stimuli, whereas the MMN is a measure of sensory memory. The Fz, FCz and Cz electrodes were analysed for the novelty oddball paradigm because these channels elicit the largest ERP responses in these types of paradigms (Duncan et al., 2009).
Time windows (in ms) used for auditory evoked potential (AEP) analyses of the novelty oddball data.
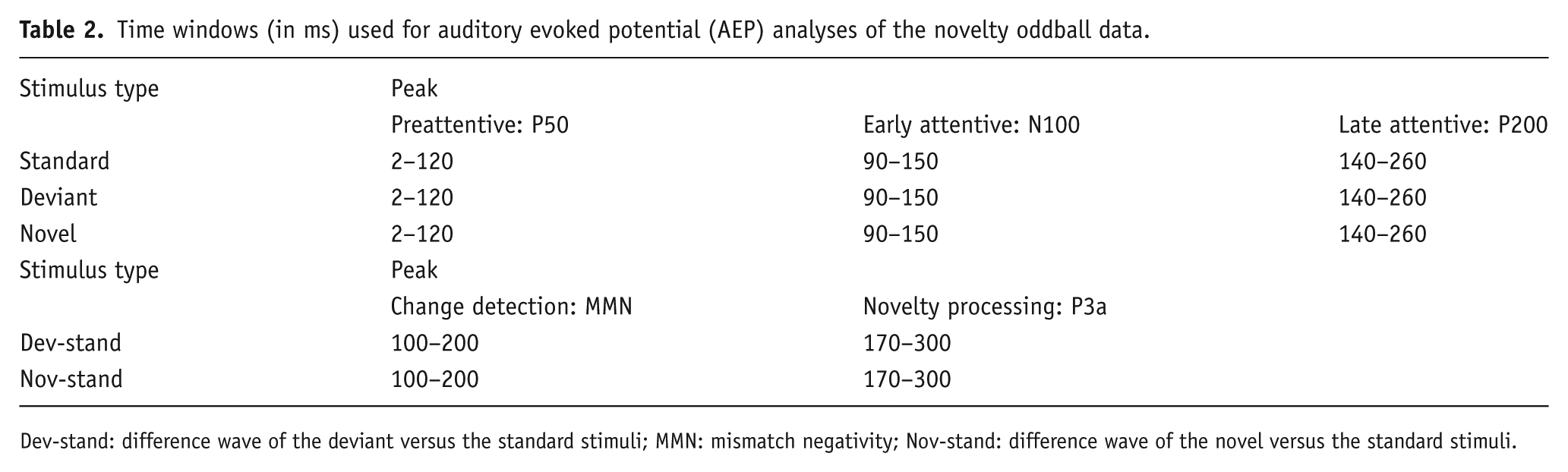
Dev-stand: difference wave of the deviant versus the standard stimuli; MMN: mismatch negativity; Nov-stand: difference wave of the novel versus the standard stimuli.
Statistical analysis
In general, behavioural data were analysed using 2×2 repeated-measures analysis of variance (ANOVA). Rivastigmine and citalopram were used as within-subject factors with both two levels (yes vs no). Separate analyses were performed for accuracy and reaction times. For the VLT immediate recall, a 2×2×3 design was used with rivastigmine (yes vs no), citalopram (yes vs no) and trial (1, 2 or 3) as within-subject factors.
For the EEG analyses of the VLT, a 2×2×3 ANOVA was conducted for amplitude and latency separately with citalopram (yes vs no), rivastigmine (yes vs no) and electrode (Fz, Cz, Pz) as within-subject factors. Of note, we did not include immediate recall trial (1, 2, 3) or peak (P3a, P3b, N400, P600) as within-subject factors as the interpretation of five-way interactions would become unnecessarily complex. Instead, we adjusted the significance level accordingly.
For the SMT, a 2×2×2 ANOVA was performed with citalopram (yes vs no), rivastigmine (yes vs no) and delay (immediate vs delayed) as within-subject factors. For the CRT, a 2×2×2 ANOVA was performed with citalopram (yes vs no), rivastigmine (yes vs no) and side (left vs right) as within-subject factors.
For the novelty oddball paradigm we first checked if our treatment conditions would affect general auditory processing (i.e. P50–N100–P200 complex) in the novelty oddball task. Therefore, we ran repeated measures ANOVAs on the data of the standard stimuli, with rivastigmine (yes vs no), citalopram (yes vs no) and electrode (three levels: Fz, FCz and Cz) as within-subject variables. Next, we sought to determine if our treatment conditions influenced novelty processing (i.e. difference scores of deviant-standard and novel-standard waves). Therefore, we performed several repeated measures ANOVAs with rivastigmine (yes vs no), citalopram (yes vs no), stimulus (two levels: deviant-standard and novel-standard), and electrode (three levels: Fz, FCz and Cz) as within-subject variables. For all data, analyses were done separately for amplitude and latency, and for each of the different peaks (AEP components). Again, significance levels were adjusted accordingly.
Results
VLT (behaviour)
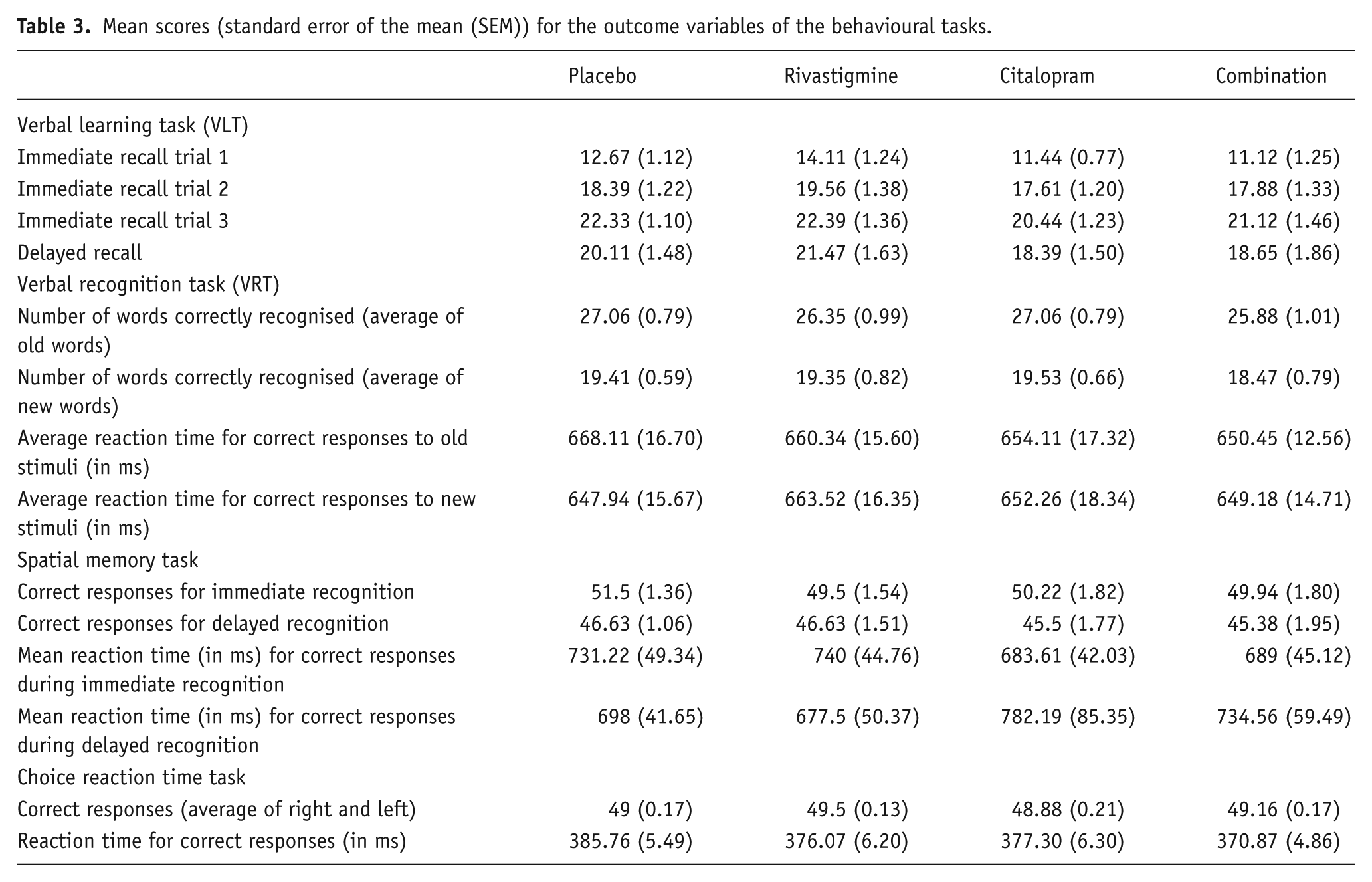
Immediate recall significantly increased from trial 1 to trial 2 to trial 3 during encoding (F(2,34)=128.117, p<0.001; data not shown). No significant interaction effect was found between citalopram and rivastigmine; however we did observe a main drug effect of citalopram on the total immediate recall score ((F(1,17)=7.630, p=0.013); Figure 1). We also found a main effect of citalopram for the delayed recall (F(1,15)=12.022, p=0.003; Figure 2 and Table 3). Participants significantly recalled fewer words after citalopram treatment than after rivastigmine or placebo. Rivastigmine had no effect and was not able to reverse the impairing effect of citalopram.
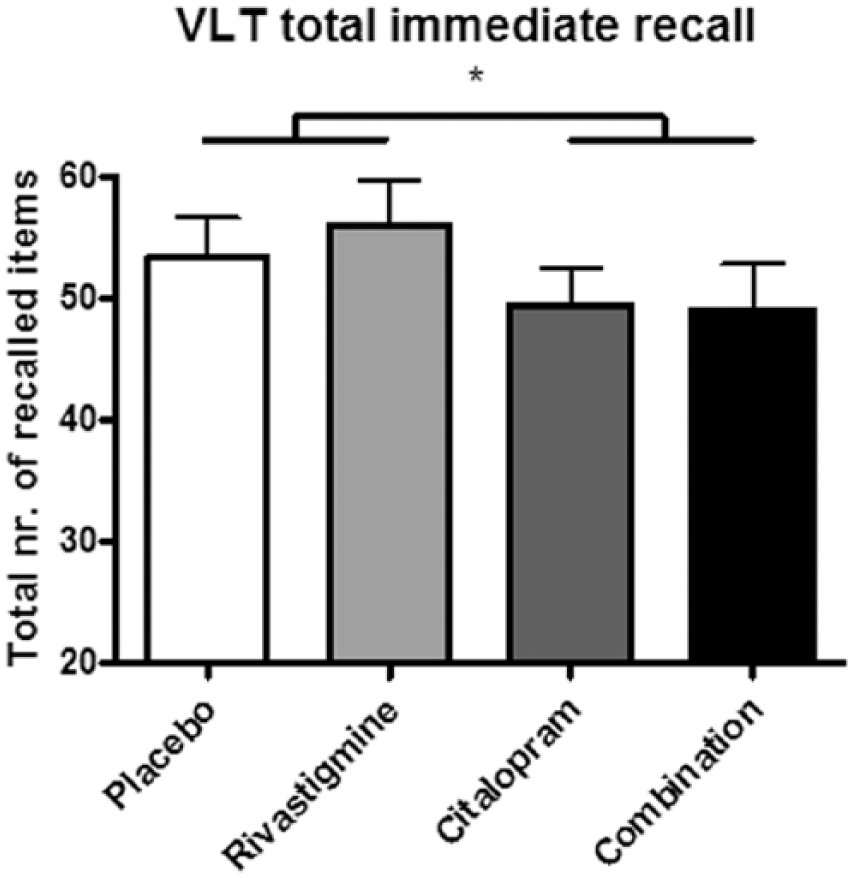
Total number of words recalled during immediate recall after each of the four treatments. Citalopram impaired memory performance, which was not reversed by rivastigmine in the combined treatment condition. VLT: verbal learning task.
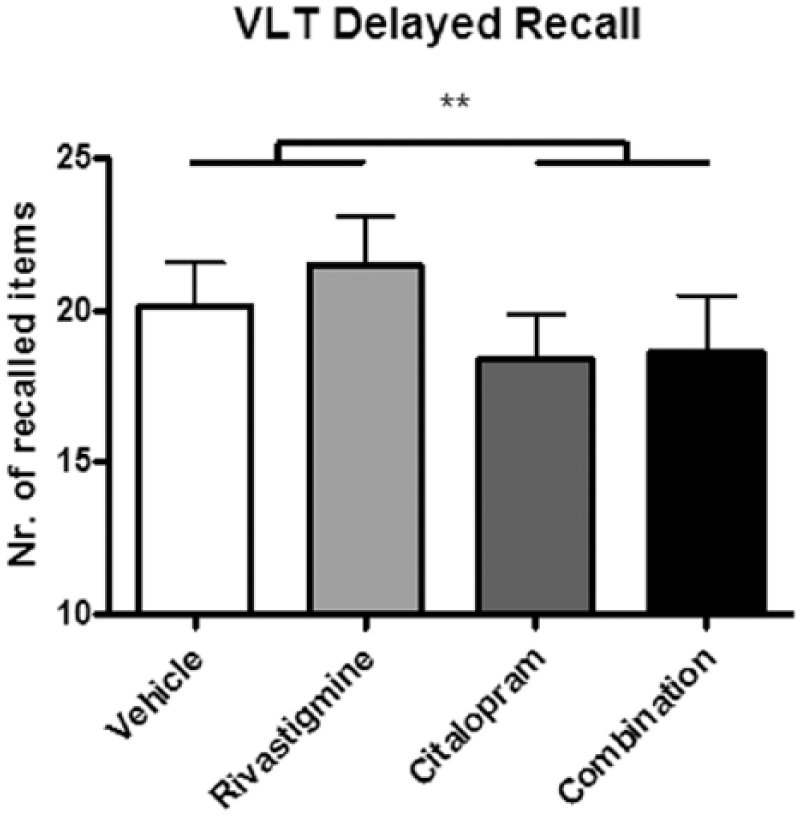
Number of words recalled during delayed recall after each of the four treatments. Citalopram impaired memory performance, which was not reversed by rivastigmine in the combined treatment condition. VLT: verbal learning task.
Mean scores (standard error of the mean (SEM)) for the outcome variables of the behavioural tasks.
VLT (EEG)
P3a
For the P3a, a significant interaction was found between rivastigmine and electrode (F(2,34)=5.619, p=0.008) at trial 2 for the amplitude. Where rivastigmine decreases the P3a amplitude at the Fz and Cz electrodes, it increases the amplitude at the Pz electrode. P3a latency was not affected by any of the treatments for any of the electrodes and trials.
P3b
For the P3b, no effect of any of the treatments at any electrode or trial was found for the amplitude. In contrast, we did observe a main effect of rivastigmine on latency during the first trial (F(1,17)=8.888, p=0.008). Rivastigmine decreased the mean latency of the P3b peak.
N400
Regarding the amplitude and latency of the N400 peak, no statistical differences were found.
P600
For the P600 peak, we observed no effects on amplitude or latency for any of the treatments.
VRT
The mean reaction time in the recognition paradigm did not show a statistical difference after any of the drug treatments, nor did we find any significant interaction. With regard to the recognition of old and new words, again no drug effects were found (Table 3).
SMT
There was a significant effect of delay (immediate vs delayed recognition) (F(1,15)=14.563, p=0.002), indicating that participants correctly recognised more locations during the immediate as compared to the delayed phase. No other significant interaction or main effects were found for the number of correctly recalled locations during the immediate or delayed recall phases of the SMT. The reaction times in this task showed a significant interaction effect between delay and citalopram (F(1,15)=11.061, p=0.005; data not shown). Citalopram appeared to decrease reaction times in the short delay and increase the reaction time at the delayed condition (Table 3). Post-hoc analyses only showed a significant effect of citalopram during the immediate recall (F(1,17)=5.635, p=0.03).
CRT task
Neither treatment showed an effect on the number of correct responses for both left and right side, nor was any significant interaction effect observed. Similarly, we found no effects of any of the treatments on reaction times (Table 3).
Novelty oddball paradigm
Treatment effects on general auditory processing of the standard stimuli
First, effects of the different treatments on general auditory processing were analysed by examining preattentive processing (P50 peak), early attentive processing (N100 peak) and late attentive processing (P200 peak) during presentation of the standard stimuli. No interaction or main effects were observed for any of the peaks at the Fz, FCz or Cz electrode (Figure 3).
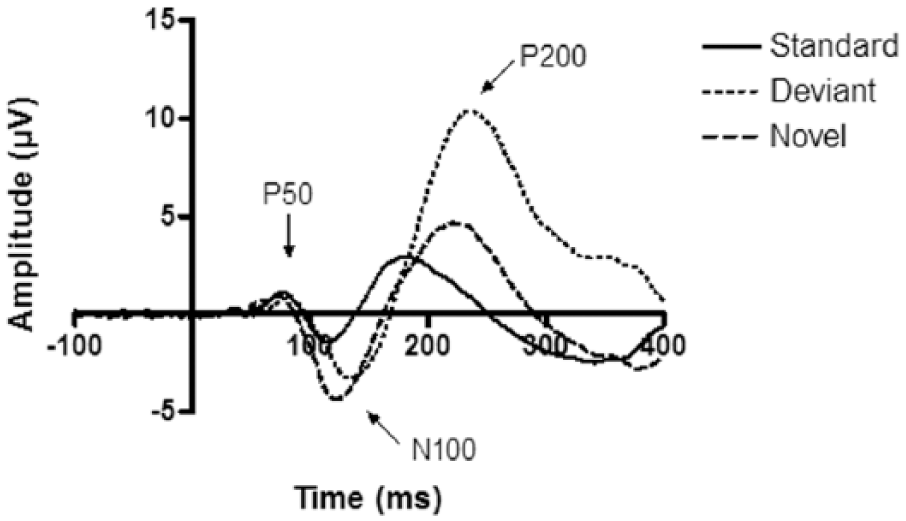
Event-related potential (ERP) responses to the standard, deviant and novel stimuli of the placebo condition for the Cz electrode. In line with normal findings in the novelty oddball paradigm, the deviant and novel sounds elicited larger amplitudes compared to the standard sounds.
Treatment effects on novelty processing: MMN
For the amplitude of the MMN peaks, we observed a significant electrode by stimulus interaction effect (F(2,100)=46.852, p<0.001). Simple effects indicated that only for the Cz electrode there was a significant difference (F(1,50)=12.164, p=0.001) between the two stimulus types showing that the peak of the novel-standard wave had a higher amplitude compared to the peak of the deviant-standard wave. However, no effect of treatment was observed on the amplitude of the waves for any of the stimulus types for any of the electrodes investigated.
For the latency, a significant main effect was observed for stimulus (F(1,50)=28.792, p<0.001) indicating that the peak of the novel-standard wave occurs slightly later then the peak of the deviant-standard wave indicative of the occurrence of the MMN. No effects of rivastigmine or citalopram were observed.
Treatment effects on novelty processing: P3a
The amplitude of the P3a peak showed a significant electrode by stimulus interaction (F(2,100)=140.939, p<0.001). Subsequent simple effect analyses showed that for all three electrodes investigated the amplitude of the P3a peak was significantly larger for the novel-standard wave compared to the deviant-standard wave (all three p<0.001). The interaction effect was caused by the fact that for the Fz electrode the difference in peak amplitude was smaller compared to the other two electrodes. Rivastigmine and citalopram did not show an effect on the amplitude of the P3a.
For the latency of the P3a no interaction or main effects were observed for any of the factors analysed.
Discussion
Considering the interaction between ACh and 5-HT on cognitive function in animal work, the aim of the current study was to assess whether both neurotransmitters interact during memory and novelty processing in humans. To this extent we increased brain levels of ACh and 5-HT in healthy volunteers by means of treatment with rivastigmine and citalopram, respectively. The main result of the study showed that during the VLT participants significantly recalled fewer words after citalopram treatment than after rivastigmine or placebo during both the immediate and delayed recall. Rivastigmine was not able to reverse the impairing effect of citalopram.
The memory-impairing effects of cholinergic muscarinic antagonists, including scopolamine and biperiden, on memory tests in animals and humans are well-established (e.g. Blokland et al., 2016; Borghans et al., 2017; Klinkenberg and Blokland, 2010; Sambeth et al., 2015). Alternatively, beneficial effects of cholinesterase inhibitors, like rivastigmine, on memory performance in animals and humans have been shown by several groups, including our own group, and these inhibitors are currently available as a symptomatic treatment for mild-to-moderate Alzheimer’s disease (e.g. Crowell et al., 2006; Onor et al., 2007; Richter et al., 2018; Theunissen et al., 2015). Together, these types of findings resulted in the cholinergic hypothesis of memory function stating that a moderate increase in ACh release will facilitate, and a decrease will impair, learning and memory. In contrast to this hypothesis, the current study did not show positive effects of rivastigmine on memory performance on the VLT, VRT or SMT. However, it has previously been suggested that cholinesterase inhibitors mainly exhibit their beneficial effects on memory performance in impaired subjects (Giacobini, 2004; Pepeu and Giovannini, 2010). This is in line with previous results of our group (Theunissen et al., 2015) and results of the current study in which healthy young adults were tested.
Regarding the serotonergic hypothesis, we observed a significant main effect of citalopram on VLT immediate and delayed recall performance indicating impaired memory function (Figures 1 and 2). Since citalopram impaired the immediate recall of verbal word memory, this could indicate that citalopram affects information processing during the encoding of words. The impairing effect of citalopram observed at the delayed recall score 30 min after learning could be the result of its interference with encoding of the words or an even earlier process such as sensory memory, or may additionally be induced by interfering with consolidation processes. However, to support the latter, additional studies are necessary. Previous studies testing the effects of 5-HT reuptake inhibitors on memory showed mixed results. For instance, similar to our study, Schmitt et al. (2001) found that the 5-HT reuptake inhibitor paroxetine impaired memory performance on the VLT only for the delayed recall (Schmitt et al., 2001). However, the 5-HT reuptake inhibitor sertraline did not show this affect in the same study. The authors suggested that the memory impairing effects of paroxetine may be induced by its additional anti-cholinergic properties. Based on findings of the current study, the impairing effect of paroxetine could be either serotonergic or cholinergic, but is not caused by an interaction between the two neurotransmitter systems. In contrast, Harmer et al. (2002) showed that citalopram improved delayed recall memory as well as recognition memory in healthy volunteers. It must be noted that in this latter study participants received a lower dose of 10 mg citalopram via an intravenous route of administration. Yet another study found no effect of citalopram on memory performance as measured by several different memory paradigms (Sambeth et al., 2015).
Rivastigmine was not able to reverse the memory deficit induced by citalopram providing support for a lack of interaction between the cholinergic and serotonergic system during verbal word memory processes. This lack of interaction was also reported by previous studies in human participants (Borghans et al., 2017; Sambeth et al., 2015). This is in contrast to what could be expected based on the anatomical level and the animal behavioural data (e.g. Steckler and Sahgal, 1995). Among others, there are serotonergic projections from the dorsal raphe nuclei (the site responsible for 5-HT release) to the basal forebrain (major cholinergic output region of the brain). Also, an important connection lies in the hippocampus, a structure highly important for memory. Here, serotonergic neurons innervate cholinergic neurons (Steckler and Sahgal, 1995). Although there are clear anatomical connections between the ACh and the 5-HT system, our previous studies do not support an interactive effect of these two neurotransmitters in cognitive functions, at least not when both neurotransmitters are manipulated globally rather than on a receptor level.
Next to the behavioural tests, electrophysiological recordings were conducted during the VLT as these have been shown to be more sensitive to drug-induced changes compared to behaviour alone (Blokland et al., 2015). Especially, the P3a and P600 components of the ERP are of interest during memory paradigms. The P3a has traditionally been linked to novelty processing, a prerequisite for memory encoding, a process that is influenced by the cholinergic system (Rangel-Gomez and Meeter, 2016). In the current study, rivastigmine enhanced the amplitude of the P3a peak at the Pz electrode but only for the second trial. A higher P3a amplitude indicates that the stimuli were perceived as more novel. As it only occurred during the second trial, it is difficult to give any meaning to this effect, i.e. it did not manifest itself as increased or decreased verbal word memory performance. Surprisingly, the behavioural effect of citalopram during the immediate recall of the VLT was not accompanied by an effect on the P3a peak. Additionally, neither effect was accompanied by an increase or decrease in P600 amplitude, while this peak has also been related to word learning (Balass et al., 2010; Van Duinen et al., 2017).
A second memory task was included to study spatial memory performance. No signs of memory improvement or impairment were found on the SMT. In contrast to the VLT, during the immediate recall of the SMT, participants all performed at a very high level (upper quartile) resulting in maximum ceiling effects. Also, based on previous studies, the SMT seems to be less sensitive to pharmacological manipulations (e.g. Sambeth et al., 2015; Van Duinen et al., 2017). This may have prevented any effects to be exhibited. Citalopram did show an effect on reaction times, i.e. reaction times decreased after citalopram treatment during the immediate recall. Without any behavioural effects, these results are again difficult to interpret.
The current study also investigated the drug effects on novelty processing using a novelty oddball paradigm. First, treatment effects on general auditory processing were excluded by examining the P50, N100 and P200 peaks of the standard stimuli (Figure 3). In line with normal findings in the novelty oddball paradigm, the deviant and novel sounds elicited larger amplitudes compared to the standard sounds. However, we did not observe any treatment effects. For the N100 and P200 peaks, our results are in line with the general literature (Brown et al., 2015; Meador et al., 1989; 1995; Pekkonen et al., 2001; Klinkenberg et al., 2013). However, for the P50, most studies showed significant cholinergic effects. It must be noted that these studies, in contrast to the current study, investigated a cholinergic muscarinic antagonistic effect of scopolamine or biperiden. In our study we increased global cholinergic levels by rivastigmine which does not lead to opposite findings, i.e. a decrease in the P50 peak.
Subsequently, treatment effects on both measures of novelty processing, i.e. MMN and P3a in the difference waves, were examined. Generally, the ERP components in the novel-standard difference wave have larger amplitudes compared to the deviant-standard difference wave, as was also observed in the current study (Figure 4). Participants exerted therefore a normal novelty response to the novel stimuli. Neither the cholinergic nor the serotonergic treatment used in the current study affected the novelty processing. Like the P50 peak, the P3a novelty peak is generally found to be decreased by scopolamine or biperiden in an oddball task (e.g. Caldenhove et al., 2017). However, Klinkenberg et al. (2013) found biperiden by itself not to affect the P3a. They did observe that biperiden reversed an increase of the P3a amplitude after treatment with rivastigmine. This effect of rivastigmine could not be replicated in the current study. Serotonergic modulation has not previously shown to affect P3a performance when using ATD to lower levels of 5-HT (e.g. Ahveninen et al., 2002). Effects of citalopram on P3a have been observed but were most pronounced in depressive patients when compared to healthy controls (Jaworska et al., 2013).
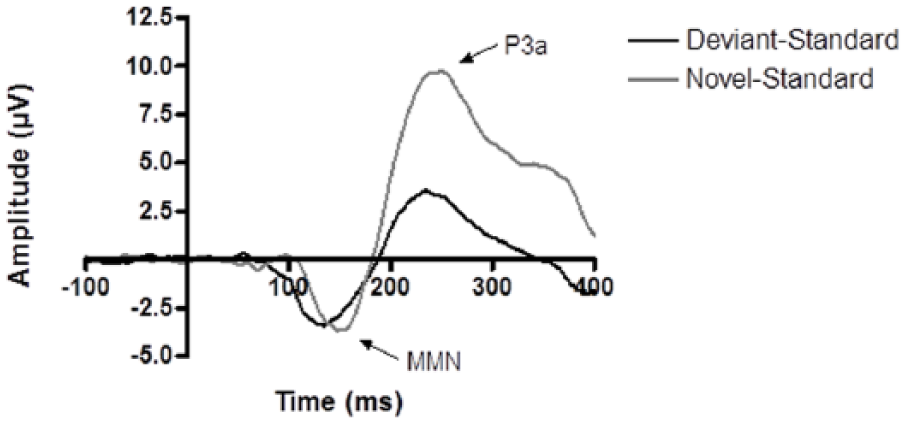
Deviant-standard and novel-standard difference waves of the placebo condition for the Cz electrode. The mismatch negativity (MMN) and P3a components for the deviant-standard wave were smaller in amplitude than the MMN and P3a components of the novel-standard wave.
For the latency of the MMN response, a significant main effect was observed for stimulus indicating that the peak of the novel-standard wave occurs slightly later then the peak of the deviant-standard wave. This observation is indicative of the occurrence of the MMN. The auditory MMN is considered to be an index of sound-discrimination accuracy and auditory sensory memory. In line with previous studies using scopolamine and biperiden, we found no effect of cholinergic modulation. Serotonergic modulation by means of ATD has been found to affect the MMN although both studies report confounding factors (Kahkonen and Ahveninen, 2002; Kahkonen et al., 2005). Citalopram did not affect the MMN in the current study.
Interestingly, the main outcome of this study was the impairing effect of citalopram on the VLT scores. As mentioned before, these effects could have been due to early memory processes like sensory memory or encoding. Given the fact that the MMN and P3a components were not affected by citalopram during the novelty oddball task, it might be suggested that citalopram negatively affects encoding but not sensory memory. This contradicts previous findings suggesting that 5-HT’s role in memory is rather related to memory consolidation (Cowen and Sherwood, 2013; Schmitt et al., 2000). These contrasting findings may relate to different treatment methods since here we used a 5-HT reuptake inhibitor whereas the depletion studies suggest an effect on consolidation.
In conclusion, the current study did not show an interaction between the cholinergic and serotonergic system during memory performance and novelty processing. This finding is in line with previous studies in which we manipulated ACh and 5-HT in different manners (Borghans et al., 2017; Caldenhove et al., 2017; Sambeth et al., 2015). Taken together, these studies in humans do not support the notion from animal studies that these two neurotransmitters interact on cognitive functions.
Footnotes
Declaration of conflicting interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: PRAH is financially supported by the Human Enhancement and Learning (HEaL) initiative of Maastricht University. This study was supported by the Netherlands Organization for Scientific Research (NWO), grant number 451-07-011.