
Abstract
Recent studies have highlighted the need for new research on intergenerational assimilation and how it varies for different origin groups. This article responds by studying the intergenerational assimilation of completed fertility in the United Kingdom. The results provide evidence of assimilation for some origins, in particular for women from Ireland and Jamaica. Yet results also show evidence against assimilation for second-generation Pakistanis and Bangladeshis. The ability of the method used to distinguish between origin groups highlights the importance of a robust statistical approach that takes account of heterogeneity, an approach that can also be used to study outcomes other than fertility.
Introduction
To fully evaluate immigration’s impact, it is important to know the extent to which the descendants of immigrants are integrating or assimilating toward mainstream norms (Portes and Zhou 1993; Zhou 1997; Crul and Vermeulen 2003; Portes, Fernández-Kelly, and Haller 2005; Heath, Rothon, and Kilpi 2008; Vermeulen 2010). This intergenerational perspective has been applied to a range of assimilation outcomes, particularly partnership, fertility, social mobility, segregation, income, and language (Massey 1981; Portes and Rumbaut 2001; Rumbaut and Portes 2001; Alba and Nee 2005; Berry 2005; Waters and Jiménez 2005; Thomson and Crul 2007). The central question for most of this research is whether and to what extent immigrants’ descendants are adopting mainstream behavioral norms (Park and Burgess 1921; Gordon 1964; Yinger 1981; Glazer 1993; Portes and Zhou 1993; Alba and Nee 1997). The answer to this question is important not only because it indicates the impact that immigration has on society but also because it shows the impact that society has on the lives of immigrants and their descendants.
Motivated by these interests, a number of studies have investigated the intergenerational assimilation of fertility (e.g., Young 1991; Parrado and Morgan 2008; Dubuc 2012). According to this literature, assimilation predicts that fertility differentials between immigrants and the native population will become smaller and eventually disappear across migrant generations. The earliest research on fertility assimilation showed evidence in support of this type of convergence (Hill 1913; Goldscheider 1965, 1967; Goldstein and Goldscheider 1968; Rosenwaike 1973; Friedlander and Goldscheider 1978), with only rare exceptions (Uhlenberg 1973). More recently, however, studies have produced conflicting results. For example, although some US studies have found that fertility differentials are smaller for the second generation than the first generation (e.g., Stephen and Bean 1992; Landale and Hauan 1996; Hill and Johnson 2004; Parrado and Morgan 2008), there is also a body of evidence that suggests migrant fertility differentials in the United States do not decline across generations (e.g., Stephen 1989; Bean, Swicegood, and Berg 2000; Carter 2000; Swicegood and Morgan 2002; Frank and Heuveline 2005).
As argued here, this apparent contradiction indicates the need for new research, in particular for research that makes consistent comparisons between different origin groups. At present, the role of origin heterogeneity is unclear in the literature, including its importance for explaining the assimilation of fertility. In addition, recent research on the assimilation of Mexican and Hispanic fertility in the United States highlights the need for further work that uses completed fertility — specifically, the total number of children ever born to a given woman at the end of childbearing — to more accurately evaluate the impact of past immigration on population size (Parrado 2011). Although research has often analyzed the number of children born using period measures (e.g., total fertility rates) based on samples that include women who have yet to complete their childbearing (e.g., women aged 15–45), studies have shown that this approach can overestimate the size of immigrant fertility differentials if women’s pre-migration fertility history is not accounted for (Toulemon 2004; Parrado 2011; Robards and Berrington 2016). This difference may explain contradictory findings, especially as compared with research on completed fertility (Parrado and Morgan 2008).
Only a small number of studies, however, have examined the intergenerational assimilation of completed fertility. Almost all such studies have focused on the United States, finding that completed fertility converges across generations for Jews (Goldscheider 1965; Goldstein and Goldscheider 1968), Italian Americans (Rosenwaike 1973), and Hispanic or Mexican Americans (Parrado and Morgan 2008) and thereby supporting the prediction of (straight-line) assimilation theory (Alba and Nee 1997). However, even though these findings point in a consistent direction, it is difficult to compare them, not least because they use different methods and study different periods. Among other things, this situation suggests the need for research that uses a consistent study design to examine the extent to which the generational convergence of completed fertility varies across origin groups.
Another advantage of comparing the generational convergence of different origin groups is that it helps show the extent to which generalizations about assimilation are appropriate. As already mentioned, much research on the intergenerational assimilation of fertility has focused on the United States, which has a fairly unique immigration history and therefore a fairly unique set of immigrant origin groups as compared with other high-income destination countries (Massey 2005). Moreover, the US literature mostly focuses on Hispanic or Mexican Americans, who, while a large share of immigrants and descendants in the United States, are not found in sizable numbers in other high-income destinations (cf. Haug, Compton, and Courbage 2002; Kulu et al. 2017).
By comparing the generational convergence of different origin groups in a non-US context, it is therefore possible to show the extent to which generalizations about fertility assimilation are appropriate. In addition, such an analysis can lay the foundations for future research, including attempts to explain why assimilation does or does not occur. If particular groups show evidence of fertility convergence, this suggests their fertility is being influenced by the destination or changes in ancestral origin norms. Alternatively, for those groups that show sustained fertility differentials, this suggests evidence of cultural entrenchment (Wilson and Kuha 2017). Just as knowledge about origin heterogeneity can help distinguish between competing explanations for assimilation, it can also demonstrate the long-term impact of different migrant streams on population dynamics. Fertility differentials that persist across migrant generations will influence population growth, which may in turn have implications for a number of policy areas, including those relating to pensions, employment, and welfare (Jonsson and Rendall 2004; Sobotka 2008).
Based on these motivations, this article aims to compare the intergenerational assimilation of completed fertility for different origin groups in the United Kingdom. It departs from previous research on migrant fertility in the United Kingdom (e.g., Dubuc 2012) by examining variations in completed fertility across generations by (ancestral) origin. It also departs from most previous research on the overall topic, which has focused primarily on the United States. In doing so, the aim is to understand the extent to which assimilation is generalizable for origin groups who are different from each other and those that have been studied in the United States. At the same time, by comparing the completed fertility of different generations, this research is also able to generate new evidence about three of the most prominent hypotheses in the literature on immigrant and ethnic minority fertility: childhood socialization, cultural entrenchment, and adaptation. These hypotheses and their relationship to assimilation theory are described in the following section, which also explains how fertility relates to intergenerational assimilation. The rest of the article then includes sections that introduce the context and data, describe the empirical method, present the results, and discuss the conclusions, including recommendations for future research.
Background
The Importance of Assimilation
Since the 1960s, the majority of countries in North America and Western Europe have experienced substantial changes in immigration patterns (Rumbaut and Portes 2001; Haug, Compton, and Courbage 2002; Alba and Nee 2005; Coleman 2009; Edmonston 2010). Not only has the size of first- and second-generation populations grown to unprecedented levels, these populations now reflect a much wider diversity of ancestral origin countries than ever before (Bouvier and Gardner 1986; Gibson 1992; Crul and Vermeulen 2003; Hirschman 2005; Statistics Canada 2006; Edmonston 2010; European Commission 2011). The magnitude of these changes has led to vigorous debate — in public, political, and academic spheres — over immigration’s economic, demographic, and societal impacts (Massey 1999, 2005; Geddes 2003; Massey, Durand, and Malone 2003; Hatton and Williamson 2005; Hirschman 2005, 2006; Picot 2008; Koehler et al. 2010; Livi Bacci 2012). Among other things, immigration has been proposed (and disputed) as a solution to the negative effects of population aging (UN 2000), and it has been debated whether immigrants are a burden on welfare and public services (Nannestad 2007).
To understand immigration’s impacts, research has begun to recognize that its lasting effects — whether social, economic, or demographic — can only be understood by looking beyond the first generation (e.g., Portes and Rumbaut 2001; Crul and Vermeulen 2003; Hirschman 2005; Thomson and Crul 2007). To understand the social outcomes of immigrants’ descendants, many researchers draw on assimilation theory, which offers predictions concerning the convergence of behavior with mainstream norms across generations (Alba and Nee 1997, 2005; Zhou 1997; Yinger 1981). Alongside this growing interest in immigrants’ descendants, researchers have also begun to argue that assimilation varies considerably for different origin groups, especially in European destinations (Crul and Doomernik 2003; Crul and Vermeulen 2003; Simon 2003; Worbs 2003; Thomson and Crul 2007; Heath, Rothon, and Kilpi 2008). Earlier research was more likely to assume that assimilation was universal, but this expectation has been challenged by the notion that assimilation can be segmented, such that it occurs for some groups and not for others (Portes and Zhou 1993; Portes, Fernández-Kelly, and Haller 2005). The fact that assimilation varies by origin makes it increasingly difficult to generalize about the process without stating which migrant groups are being considered. It also means that there is a need for research that describes this variation and allows consistent comparisons between origin groups.
What Does Intergenerational Assimilation Predict for Fertility?
For demographers, these recent trends in assimilation research have not gone unnoticed. The childbearing of immigrants and their descendants is a critical determinant of immigration’s long-term impact on population size, and this has led to increasing interest in the fertility of both immigrants and their descendants (e.g., Feld 2000; Beaujot 2002; Coleman 2002, 2006; Jonsson and Rendall 2004; Sobotka 2008; Parrado 2011; Dubuc 2012, 2016). This attention has been particularly notable in Europe, where the second generation has been found to have distinct patterns of partnership and fertility behavior (Kulu and González-Ferrer 2014). Given that most European countries are receiving immigrants from an increasingly wide range of origins, demographers have also become interested in the heterogeneity of behavior by origin group for both immigrants and their descendants and increasingly acknowledge the lack of research on this topic (Haug, Compton, and Courbage 2002; Sobotka 2008; Kulu and González-Ferrer 2014).
Although researchers have interpreted the concept of assimilation in a variety of ways (e.g., Yinger 1981; Glazer 1993; Portes and Zhou 1993; Zhou 1997; Brubaker 2001; Alba and Nee 2005), for the purposes of this study, it is important to distinguish between individual and intergenerational assimilation. Although for some, assimilation theory predicts the convergence of immigrant fertility over an individual life course (often referred to as adaptation, e.g., Milewski 2010), assimilation also predicts generational convergence, which is the subject of study here (e.g., Goldstein and Goldscheider 1968; Rosenwaike 1973). It has been noted that studies of fertility assimilation have tended to focus on adaptation rather than taking an intergenerational perspective (Bean, Swicegood, and Berg 2000; Haug, Compton, and Courbage 2002; Parrado and Morgan 2008; Sobotka 2008; Milewski 2010). In part, this may be a result of the additional data requirements associated with estimating fertility for immigrants’ descendants.
In general, (straight-line) assimilation predicts a process of convergence, defined as the decline and eventual disappearance of differences between a given migrant or ethnic group and the mainstream destination norm (Alba and Nee 1997). This article refers to these differences as differentials and follows the majority of the literature on fertility assimilation in measuring the mainstream norm using ancestral natives’ average fertility (e.g., Parrado and Morgan 2008). Thus, when intergenerational assimilation is applied to fertility, convergence occurs if there is a smaller fertility differential for the second generation as compared with first-generation migrants. I note, however, that convergence is not necessarily expected to occur over a single generation (between first and second) and that caution is therefore required when interpreting findings that exclude consideration of third or later generations.
When determining what intergenerational assimilation predicts for fertility, one further consideration is how to compare the fertility of different generations. Here, I follow Parrado and Morgan (2008, 651), who argue that studies of intergenerational assimilation should “more closely approximate a comparison of immigrant women with those of their daughters’ and granddaughters’ generation.” In other words, rather than making a cross-sectional comparison of the first and second generations from the same birth cohort, tests of intergenerational assimilation require generations to be compared across lagged birth cohorts (Smith 2003, 2006; Waters and Jiménez 2005). This approach has also been argued for and investigated using period measures of fertility in the United Kingdom (e.g., Dubuc 2016). Generational convergence is taken here to be the prediction of intergenerational assimilation and is evidenced by a narrowing of completed fertility differentials for the descendants of immigrants as compared with immigrants from earlier birth cohorts.
Hypotheses That Are Linked to Fertility Assimilation
The concept of intergenerational assimilation was first applied to research on immigrant fertility over 100 years ago (Hill 1913). Since then, the literature has developed a number of hypotheses related specifically to the fertility of immigrants and their descendants (Goldberg 1959, 1960; Goldscheider and Uhlenberg 1969; Ritchey 1975; Zarate and Zarate 1975; Goldstein and Goldstein 1981, 1983; Hervitz 1985; Coleman 1994; Forste and Tienda 1996; Abbasi-Shavazi and McDonald 2000; Kulu 2005). However, many of these hypotheses relate to the timing or “tempo” of birth rather than completed fertility (Wilson 2013). As such, hypotheses that make predictions relating to completed fertility convergence are childhood socialization, cultural entrenchment, and adaptation, each of which is defined next.
The childhood socialization hypothesis is based on the idea that fertility norms are developed during childhood (Hervitz 1985). For first-generation adult immigrants, the hypothesis predicts that fertility norms are established prior to migration and that this explains why immigrant fertility is different from the native norm. On the other hand, for the second generation, it predicts an absence of fertility differentials because this cohort has been exposed to destination norms throughout childhood. In this research, I therefore conclude that there is evidence of childhood socialization when there is a significant difference between the fertility of ancestral natives and first-generation adult immigrants but no significant difference between the fertility of ancestral natives and the second generation.
In contrast to childhood socialization, cultural entrenchment predicts that assimilation might not occur for some migrant groups because their norms and preferences are “culturally entrenched” (Goldscheider and Uhlenberg 1969; Forste and Tienda 1996). In the context of immigrant fertility, this explanation is sometimes referred to as cultural maintenance or the cultural hypothesis (Abbasi-Shavazi and McDonald 2000; Milewski 2010). There is a long history of research that uses cultural variation to explain immigrant fertility differentials; however, hypotheses based on culture have often been stated in a way that makes them hard to falsify (a point alluded to by Coleman 1994; Forste and Tienda 1996). In response, cultural entrenchment is defined here as a somewhat crude but nevertheless falsifiable hypothesis that predicts that immigrants’ descendants from some origin groups will have different levels of completed fertility than the native norm. This may be due to the influence of subcultural norms or a lack of exposure to mainstream norms.
A third hypothesis often linked to fertility convergence is adaptation. In most research, this hypothesis refers to a prediction that first-generation immigrant fertility will converge toward native norms after arrival (Hervitz 1985; Milewski 2010). Immigrants may differ in their birth timing, for example, if migration disrupts childbearing, but they are expected to rapidly conform to native norms (Goldstein and Goldstein 1983; Hervitz 1985). In this sense, the adaptation hypothesis (as used in fertility research) has considerable overlap with assimilation theory, with some authors suggesting that the two are synonymous (e.g., Stephen and Bean 1992). However, it is important to note that this overlap depends on the way in which assimilation is defined, and evidence of adaptation might also be explained by socioeconomic factors rather than cultural factors like those related to acculturation (Andersson and Scott 2005, 2007). Nevertheless, adaptation predicts a decline in the differences between immigrants and natives over time, and this process may eventually lead to a situation where there is no difference between the fertility of first-generation migrants and natives. In this article, such a situation is referred to as potentially indicative of adaptation, although there are several competing explanations for why this might not be a result of adaptive behavior (including selection and reverse causality, which are discussed below) (Harbison and Weishaar 1981; Toulemon 2006).
All three of the aforementioned hypotheses — socialization, entrenchment, and adaptation — have a link to assimilation theory. In particular, childhood socialization is seemingly synonymous with the prediction of intergenerational assimilation (i.e., generational convergence). Like generational convergence, childhood socialization is evident when first-generation fertility is different from the native norm but second-generation fertility is not. However, the difference between socialization and assimilation (at least in this article) is that the former does not require a direct comparison between generations to be supported. On the other hand, adaptation and entrenchment represent two possible explanations for a lack of convergence. Generational convergence may be lacking because the completed fertility of some second- or later-generation ancestral groups remains different from the native norm, a situation that is evidence of cultural entrenchment. Generational convergence may also be lacking because there is no difference in completed fertility between natives and the first generation, a situation that might be explained by the fact that assimilation or adaptation has already occurred.
It is perhaps worth noting that this study places a priority on establishing whether convergence occurs, with the additional goal of investigating the hypotheses that make predictions for different generations. Causal explanations for convergence therefore fall outside the remit of this study. This is not to say that causality is irrelevant here since it is implicit in the hypotheses and explanations that are investigated. However, if generational convergence (or socialization) and alternative explanations like adaptation and cultural entrenchment are to be distinguished, the first step is to test explicit descriptive predictions. The results of this first step can then be used to guide future research. For example, the comparison of immigrant origin groups undertaken here can be used to highlight the origins that have the most divergent patterns of convergence and therefore the groups that may warrant further investigation.
Another implication of this lack of focus on causality is that reverse causality and selection, two processes that have been used to explain migrant fertility, are not focal concerns for this research (Forste and Tienda 1996; Toulemon 2004, 2006; Sobotka 2008). There is no doubt that selection will be manifest across the various migration processes that determine which immigrants are resident in the United Kingdom. However, the presence of selection does not prevent an assessment of whether convergence occurs. Related to selection is the possibility that differences in the completed fertility of subsequent generations could be explained by differences in their social characteristics (Goldscheider and Uhlenberg 1969; Forste and Tienda 1996; Sobotka 2008). This explanation is briefly considered in the final section of this analysis, with an examination of the influence of education and partnership history.
Context and Data
As a case study for investigating the completed fertility of immigrants and their descendants from different origins, the United Kingdom is advantageous because it has a large, diverse, and well-established immigrant population (Walvin 1984; Rendall and Salt 2005; Zumpe, Dormon, and Jefferies 2012). The history of immigration to the United Kingdom is both nuanced and extensive (e.g., Coleman, Compton, and Salt 2002; Horsfield 2005; Rendall and Salt 2005; Peach 2006), as is the history of UK immigration policy (Home Office 2014). As such, only a brief summary of both is provided here.
Overall, the size and diversity of both the first- and second-generation population in the United Kingdom enable the estimation of completed fertility for a range of origin groups. The easiest origins to compare across generations are those with a history of substantial levels of immigration, which is most clearly applicable to immigrants from Ireland, South Asia, and the Caribbean. Historically, the largest group of immigrants to the United Kingdom have come from Ireland, but that trend began to change in the last decades of the twentieth century (ONS 2013). More recently, Irish immigrants have been replaced by Indians as the largest foreign-born group in England and Wales (ONS 2012). Indian immigration began in earnest in the late 1960s and early 1970s (Walvin 1984), whereas the inflow of immigrants from Pakistan reached significant numbers in the mid-1970s, and immigration from Bangladesh did not gather pace until the late 1970s and early 1980s (Coleman, Compton, and Salt 2002). Nevertheless, by 2011, almost 1.4 million people in England and Wales were lifetime immigrants from one of these three South Asian countries, equivalent to 18 percent of the foreign-born population. Compared with immigration from South Asia, immigration from the Caribbean began earlier in the twentieth century. Peaking in the 1950s and 1960s, it fell significantly after the Commonwealth Immigrants Act introduced restrictions on immigration in 1962 (Foner 2009). Nonetheless, much family reunification occurred after the Act, leading to the continued immigration of Caribbean women throughout the 1960s. In 2011, Jamaica was still the 10th largest foreign country of birth in England and Wales by population size, and a considerable number of first- and second-generation Caribbean women are resident in the United Kingdom (ONS 2013).
There is limited research on fertility in the United Kingdom that distinguishes between first- and second-generation origin groups, in particular with respect to long-term trends and second-generation fertility. Previous research shows that the period total fertility rate (TFR) in the United Kingdom has fallen for many ethnic minority groups since the 1970s, especially for Pakistani and Bangladeshi women (Iliffe 1978; Coleman 1994; Sigle-Rushton 2008; Dubuc 2009, 2012; Coleman and Dubuc 2010; Dubuc and Haskey 2010). In general, there is much less variation in the TFRs of second-generation ethnic groups in the United Kingdom (Coleman and Dubuc 2010; Dubuc 2012, 2016). Furthermore, as Dubuc (2012) has shown, when making a lagged comparison between generations for South-Asian ethnic groups, the period TFR of immigrants’ descendants is closer to the UK average than the TFR of the first generation.
These findings align with research that uses registered births and the own-child method 1 to show that second-generation fertility is more similar, on average, to natives’ fertility in England and Wales (Wilson 2011). Similarly, several UK studies have shown that differences between parity-specific birth rates of natives and immigrants’ descendants are typically smaller than the equivalent differences for immigrants from similar countries of birth (Kulu et al. 2015, 2017; Kulu and Hannemann 2016). Despite the broad alignment between these findings, there is also evidence of persistently high fertility for some origin groups, most notably Pakistanis and Bangladeshis, a finding often attributed to the influence of cultural factors (Kulu et al. 2015, 2017; Kulu and Hannemann 2016; Wilson and Kuha 2017). As of yet, however, there has been no research that compares the completed fertility of immigrants and their descendants in the United Kingdom or in any of its four constituent countries (England, Wales, Scotland, or Northern Ireland).
This investigation makes use of data from the first wave of Understanding Society (UKHLS), which constitutes a representative sample of approximately 40,000 households in the United Kingdom (Buck and McFall 2011). This source is particularly useful because it allows the identification of different migrant generations, estimation of completed fertility, and identification of a range of ancestral origin groups. The first wave of UKHLS data includes around 60,000 adults surveyed between 2009 and 2011. Importantly, approximately 10 percent of this sample is part of an ethnic minority boost, which means that the first and second generations are overrepresented.
The Analytical Sample
The analytical sample examined here is restricted to women born between 1922 and 1971. This provides a suitable range for the sample to be split into two birth cohort groups: 1922–1951 and 1952–1971. These groups are chosen to ensure that all women are aged 40 or above and, because the midpoints of these two groups are 25 years apart, so that a comparison can be made between lagged birth cohorts. However, it is important to note that this is an average and that not all individuals in these two groups were born 25 years apart. The migrant generations analyzed here are defined as follows: First-generation adult immigrants are women who were born outside the United Kingdom between 1922 and 1951 and arrived in the United Kingdom when aged 16 and older. The second generation are women who have one or more foreign-born parent(s) and were born in the United Kingdom between 1952 and 1971.
Ancestral natives are UK-born women with two UK-born parents.
Table 1 shows the cases excluded from the analytical sample because of ineligibility or missing data. To make the results easier to interpret for first-generation migrants, the sample excludes child migrants and foreign-born women with UK-born parents. The inclusion of child migrants as a separate group is not possible with this data source because the sample size is too small to allow child migrants to be disaggregated by country of birth. On the other hand, foreign-born women with UK-born parents are excluded because it is difficult to assess the extent to which these women have been exposed to either foreign or native fertility norms. Cases are also dropped from the sample if they are surveyed by proxy or missing information on parental country of birth, fertility history, or the covariates used in the analysis. This results in a sample size of 14,250 women assumed to have completed their fertility, including 461 first-generation adult migrants born between 1922 and 1951 and 862 second-generation women born between 1952 and 1971.
The Analytical Sample.
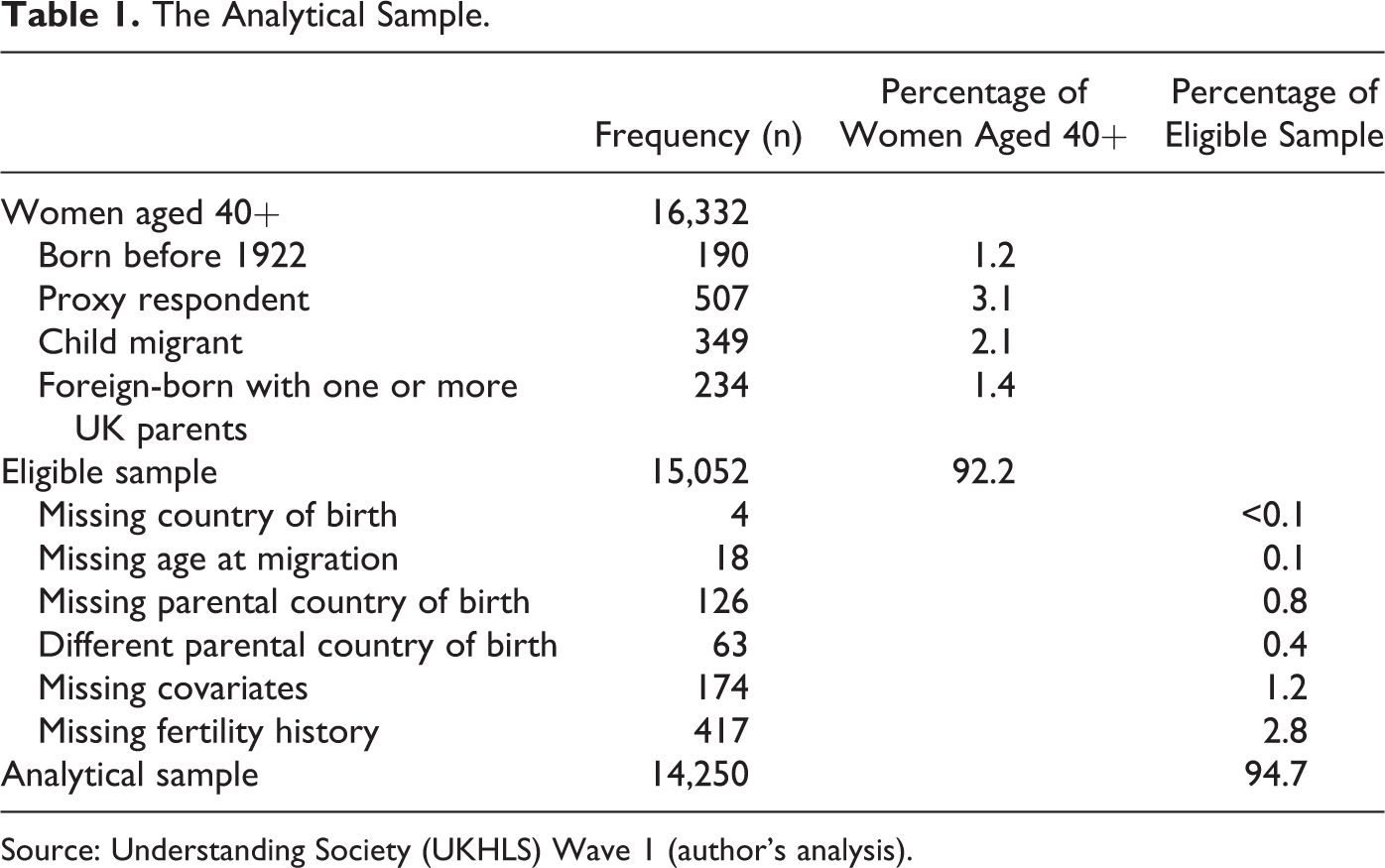
Source: Understanding Society (UKHLS) Wave 1 (author’s analysis).
Method
The method used here builds on US research that analyzes the completed fertility of single ancestral groups (Goldscheider 1965; Goldstein and Goldscheider 1968; Rosenwaike 1973; Parrado and Morgan 2008). In doing so, it also builds on previous approaches that make a lagged comparison of different generations (e.g., Parrado and Morgan 2008; Dubuc 2012). Given the importance of a testing an explicit definition, generational convergence is defined here as occurring when the difference between the completed fertility of first-generation adult immigrants and ancestral natives for a given birth cohort group (G1) is larger than the difference between the completed fertility of the second generation and corresponding ancestral natives born 25 years later (G2). Thus, a comparison is made across generations as though one were comparing first-generation migrants (G1) with their children, but in this case, their children are represented by second-generation women born 25 years later (G2). The use of 25 years as an appropriate gap between generations could be contested, but this value is chosen because it matches that used in previous research (e.g., Parrado and Morgan 2008).
Variables and Model Specification
The analytical sample includes women aged 40 and above, and their fertility is measured using children ever born, which is assumed to represent completed fertility. 2 The UKHLS data measure women’s fertility using information from birth history questions answered by respondents. Country of birth, parental country of birth, and birth cohort are used to define migrant generations and ancestral natives.
The reference group for all migrant/native comparisons is ancestral natives: UK-born women with two UK-born parents. Before comparing migrant generations, each generation is first compared against the average completed fertility for ancestral natives in the same birth cohort as the migrant generation, although this comparison group is altered when control variables are added to the analysis. The differences in fertility between migrant generations and ancestral natives are then compared to discover which generation, if any, is closer to the native norm of their birth cohort. Apart from Table 1 and Figure 1, the rest of the analysis disaggregates migrants by (ancestral) origin.
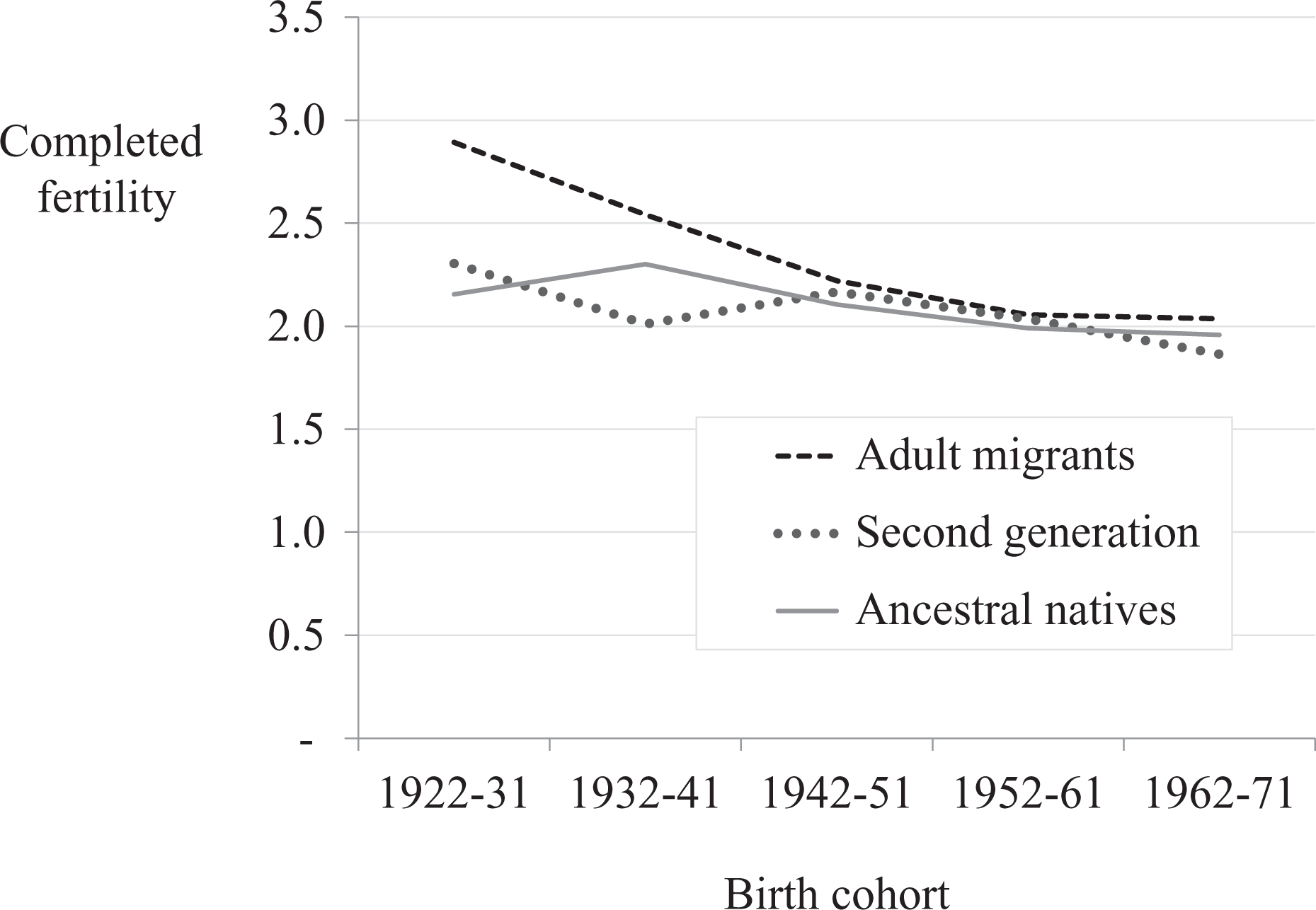
Mean number of children (completed fertility) by birth cohort and generation.
Table 2 shows the different origin groups used throughout alongside their mean completed fertility, unweighted frequency (sample size), and distribution in each generational group. In addition to trying to create ancestral groups with a similar migration history and a reasonable sample size, certain countries have been deliberately separated in this analysis. This decision is largely based on the fact that their total fertility rates are known to be different from the UK-born population (Iliffe 1978; Coleman 1994; Dubuc 2012). Unfortunately, sample size limits this strategy to consideration of the largest populations.
Non-Natives in the Generations and Cohorts of Interest: Completed Fertility, Sample Size, and Distribution by Origin Group.
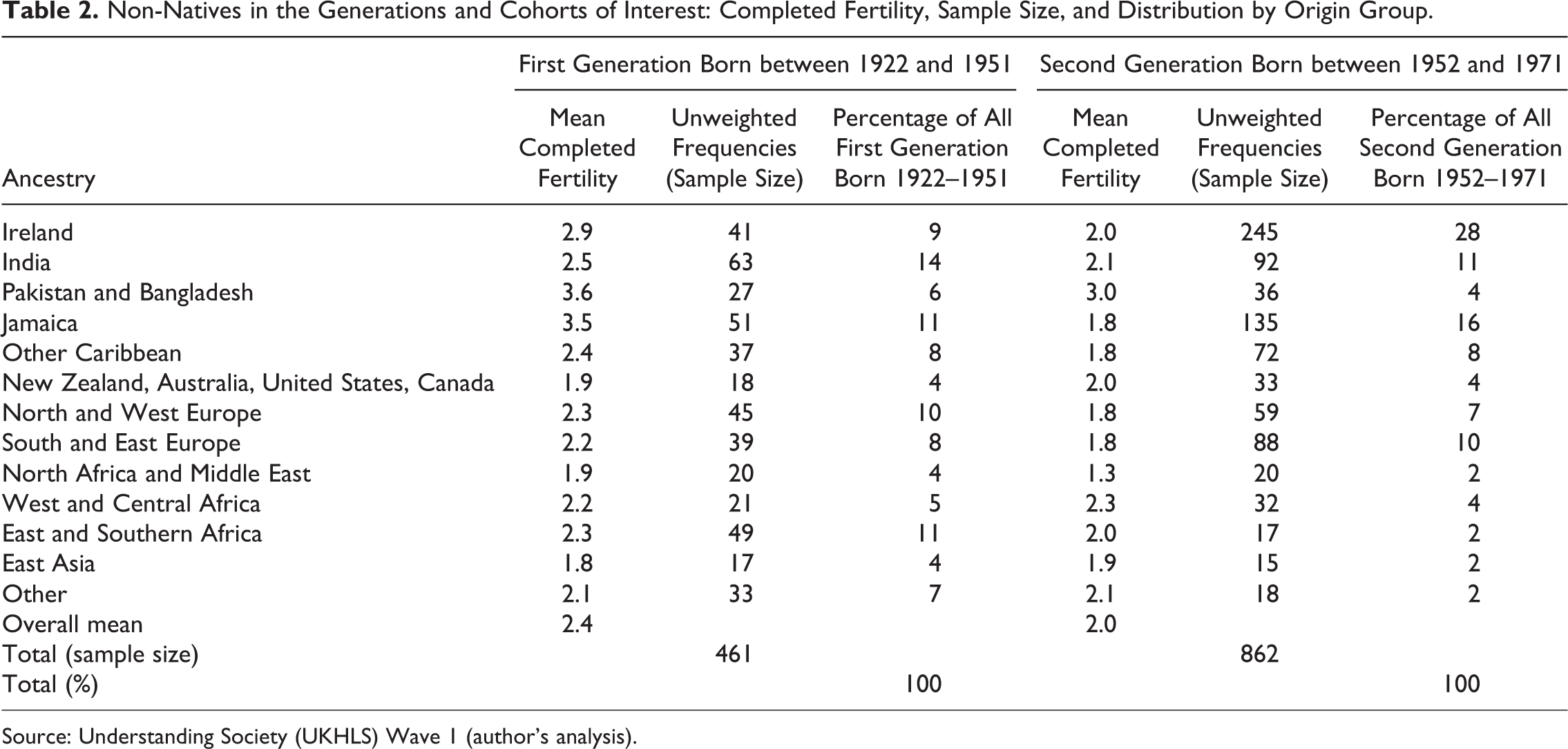
Source: Understanding Society (UKHLS) Wave 1 (author’s analysis).
Except for Figure 1, the rest of the analysis uses count regression models to investigate the effects of origin and ancestry (Agresti 2002). These models have been shown elsewhere to be appropriate for modeling birth counts to evaluate migrant fertility (Mayer and Riphahn 2000; Adserà et al. 2012). All weighted estimates and regression models were estimated using the svy command in Stata version 11 to account for the UKHLS’s complex survey design (StataCorp 2009). Thus, results take into account unit nonresponse and survey design, including the fact that ethnic minorities (and hence immigrants and their descendants) are oversampled, and are representative of the UK population.
The regression models are defined as follows: Let subscript j denote a generation-ancestry group (e.g., first-generation Irish adult migrants, second-generation Irish migrants, etc.). Let subscript k denote birth cohort,
In the second set of regression results, the same three comparisons are made but conditional on covariates. In essence, the main analysis, which estimates completed fertility by origin and birth cohort, is repeated with the addition of controls for education (and partnership history, although these models are discussed rather than shown). The addition of control variables has the effect of changing the comparison between migrants and natives (i.e., the calculation of migrant fertility differentials) to be conditional on these controls, which is viewed here as a form of standardization that helps show how likely it is that the main results might be explained by these social characteristics. However, the results are considered tentative, not least because of the limited number of controls used.
Assessing Generational Convergence
To assess convergence, the IRRs of the first and second generations (as defined previously) are compared across their respective lagged birth cohorts. If the second-generation IRR is closer to 1.0 (i.e., closer to natives) than the first-generation IRR, this suggests evidence of generational convergence. However, a further test is required to establish whether this evidence is significant. To summarize the results for every ancestry group, comparisons are therefore categorized as follows:
For ease of generalization, “yes” is considered to represent groups where there is the strongest evidence of generational convergence, although despite these strictly defined categories, this research does not seek to overinterpret the accuracy of estimates. The 5 percent benchmark is somewhat arbitrary and represents a fairly high type one error. Similarly, results in the other categories may be inaccurate due to uncertainty. As the name suggests, results that are not significant imply some possibility of convergence, which might be detected if a larger sample were available. The strongest evidence against convergence would be a “no” with a statistically significant ratio, which describes a significant movement away from the native fertility norm.
In taking this approach, the aim is to enable a consistent, valid, and reliable comparison of different origin groups. I compare origin groups directly using consistent definitions and statistical tests of convergence that match the conceptual framework. To do so, I make the same comparison for each group, which maximizes the reliability of the approach.
Results
Before examining generational convergence by ancestry, Figure 1 provides an overview of completed fertility trends for the first and second generations alongside ancestral natives. These results give some indication of aggregate patterns of convergence. There is a notable difference in completed fertility between adult immigrants and ancestral natives for the oldest cohorts as compared with the negligible difference between the second generation and ancestral natives for the cohorts born 20 to 30 years later.
Before comparing across generations, each generation can be compared against the ancestral native average for the same birth cohort. An overview of comparative fertility patterns is therefore provided by looking at the estimated IRRs for the two generation/cohort groups (see Table 3). For example, adult immigrant women from Ireland born between 1922 and 1951 have completed fertility that is 30 percent higher than ancestral natives (IRR = 1.3, p = .03). Without any comparison across generations, it is apparent that completed fertility is higher than that of ancestral natives in the United Kingdom for first-generation Irish and Jamaicans and first- and second-generation Pakistanis and Bangladeshis, whereas there is lower completed fertility than natives for second-generation women from North Africa and the Middle East.
Convergence across Generations: By Ancestry.
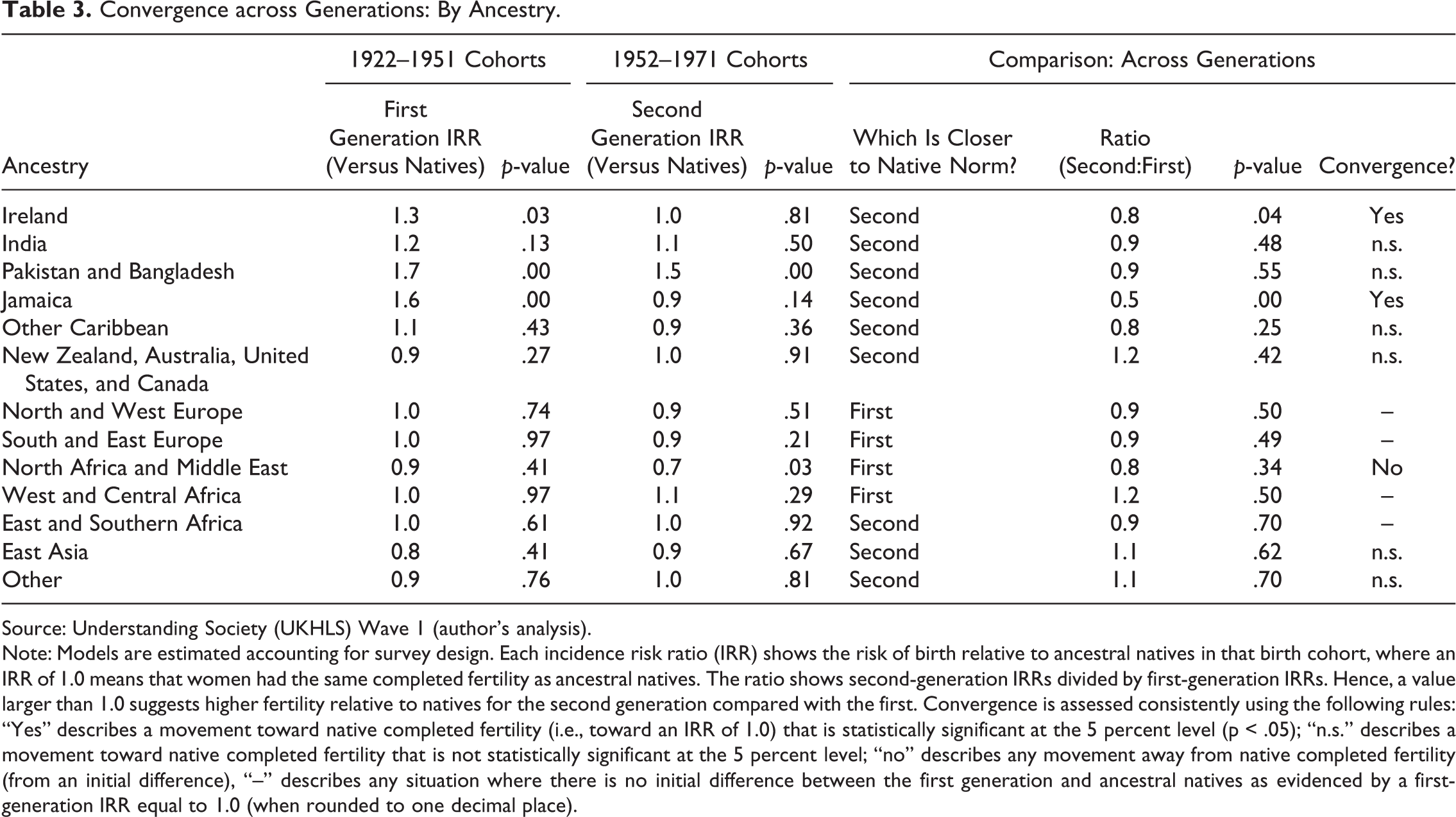
Source: Understanding Society (UKHLS) Wave 1 (author’s analysis).
Note: Models are estimated accounting for survey design. Each incidence risk ratio (IRR) shows the risk of birth relative to ancestral natives in that birth cohort, where an IRR of 1.0 means that women had the same completed fertility as ancestral natives. The ratio shows second-generation IRRs divided by first-generation IRRs. Hence, a value larger than 1.0 suggests higher fertility relative to natives for the second generation compared with the first. Convergence is assessed consistently using the following rules: “Yes” describes a movement toward native completed fertility (i.e., toward an IRR of 1.0) that is statistically significant at the 5 percent level (p < .05); “n.s.” describes a movement toward native completed fertility that is not statistically significant at the 5 percent level; “no” describes any movement away from native completed fertility (from an initial difference), “–” describes any situation where there is no initial difference between the first generation and ancestral natives as evidenced by a first-generation IRR equal to 1.0 (when rounded to one decimal place).
Evidence of Convergence for Different Migrant Groups
Comparing different groups, there is statistically significant evidence of generational convergence for women with Irish and Jamaican ancestries (Table 3). In addition to a significant comparison between generations, these ancestral groups show no significant difference between the second generation and ancestral natives. As such, Irish and Jamaican fertility could be judged to have fully converged. When considered alongside the fact that first-generation fertility is significantly different from the native norm, this evidence also supports the childhood socialization hypothesis (Table 4).
Confirmation of Migrant Fertility Hypotheses with Specific Predictions for Both the First and Second Generation.
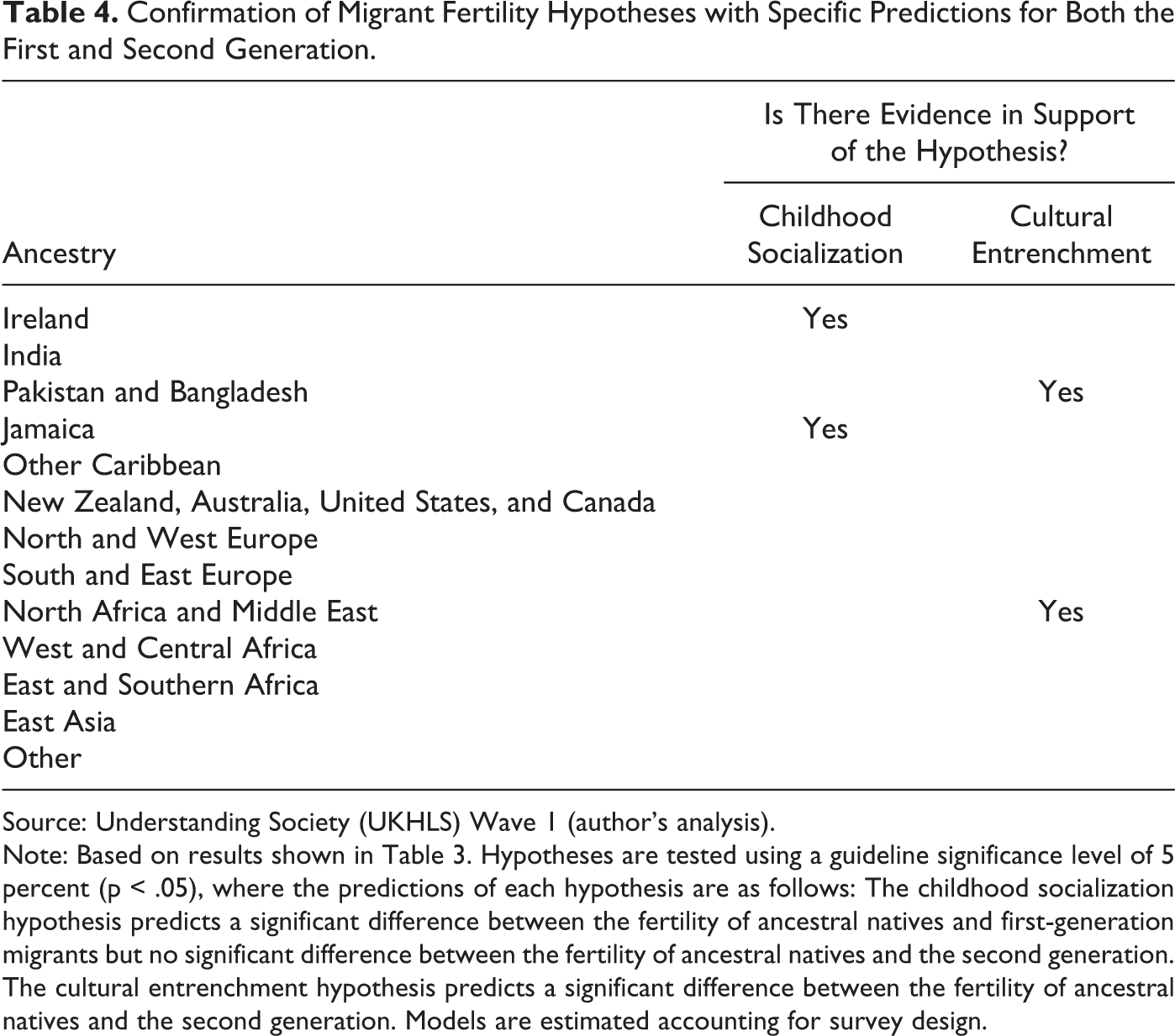
Source: Understanding Society (UKHLS) Wave 1 (author’s analysis).
Note: Based on results shown in Table 3. Hypotheses are tested using a guideline significance level of 5 percent (p < .05), where the predictions of each hypothesis are as follows: The childhood socialization hypothesis predicts a significant difference between the fertility of ancestral natives and first-generation migrants but no significant difference between the fertility of ancestral natives and the second generation. The cultural entrenchment hypothesis predicts a significant difference between the fertility of ancestral natives and the second generation. Models are estimated accounting for survey design.
There is some (nonsignificant) evidence of convergence for all South Asian groups as well, but convergence patterns are very different for Indians compared with Pakistanis and Bangladeshis. Indian women’s completed fertility is not significantly different from natives for either generation. As such, the results for Indian women might be indicative of adaptation. Even if this is not the case, however, it is clear that the IRRs for Indians are much smaller than for other South Asians. On the other hand, there is evidence of cultural entrenchment for Pakistani and Bangladeshi migrants, who have IRRs that are significantly higher than natives for both the first and second generations. At a minimum, these results demonstrate that South Asian migrants in the United Kingdom do not exhibit a homogenous pattern of generational convergence.
The remaining ancestry groups are classed as having no initial difference, nonsignificant evidence, or evidence of no convergence. For those classed as nonsignificant, it is important to differentiate between those groups where first-generation completed fertility is higher than natives (e.g., South Asians) and those for whom it is lower (e.g., East Asians). Only one group — descendants of immigrants from North Africa and the Middle East — shows evidence of no convergence such that second-generation fertility is diverging from the native norm. According to this study’s definitions, there is also evidence of cultural entrenchment for this origin group, although this interpretation assumes that they have a culturally entrenched norm for lower fertility than in the United Kingdom. Given that fertility is on average higher in these regions than in the United Kingdom (UN 2013), this finding suggests that a different explanation for this result may be more plausible.
Social Characteristics
There is neither space nor data to study all the characteristics that may explain convergence. Instead, education and partnership history are investigated as examples to show how this analysis can be extended to explore other social characteristics. It is important to emphasize that the aim is not to try to isolate ancestry’s true effect (net of other characteristics) as this would raise a number of methodological issues, not least those relating to the fact that education, partnership, and fertility are all simultaneous processes. Instead, the aim is to investigate how the results change when the comparison group is changed to individuals with the same characteristics (a somewhat similar approach to that of standardization). In addition to adjusting comparisons of the first and second generations, this change also affects the comparison of migrant generations with natives. For example, the addition of education controls means that Irish adult migrants are compared with natives who have the same education.
Table 5 displays the results after adding education controls to the models shown earlier (in Table 3). For the most part, there are no material changes to the results. For example, the IRR for first-generation Irish women born 1922–1951 is only 2 percent higher after adding controls (although this represents an increase from a rounded IRR of 1.3 to 1.4). Furthermore, the new analysis does not change the qualitative inferences made about convergence for this (or any) group. In fact, the only material change is for women from Western and Central Africa, who change from no initial difference (between the first generation and natives) to showing slight but nonsignificant evidence of an initial difference.
Convergence across Generations: Including Covariates Relating to Education.
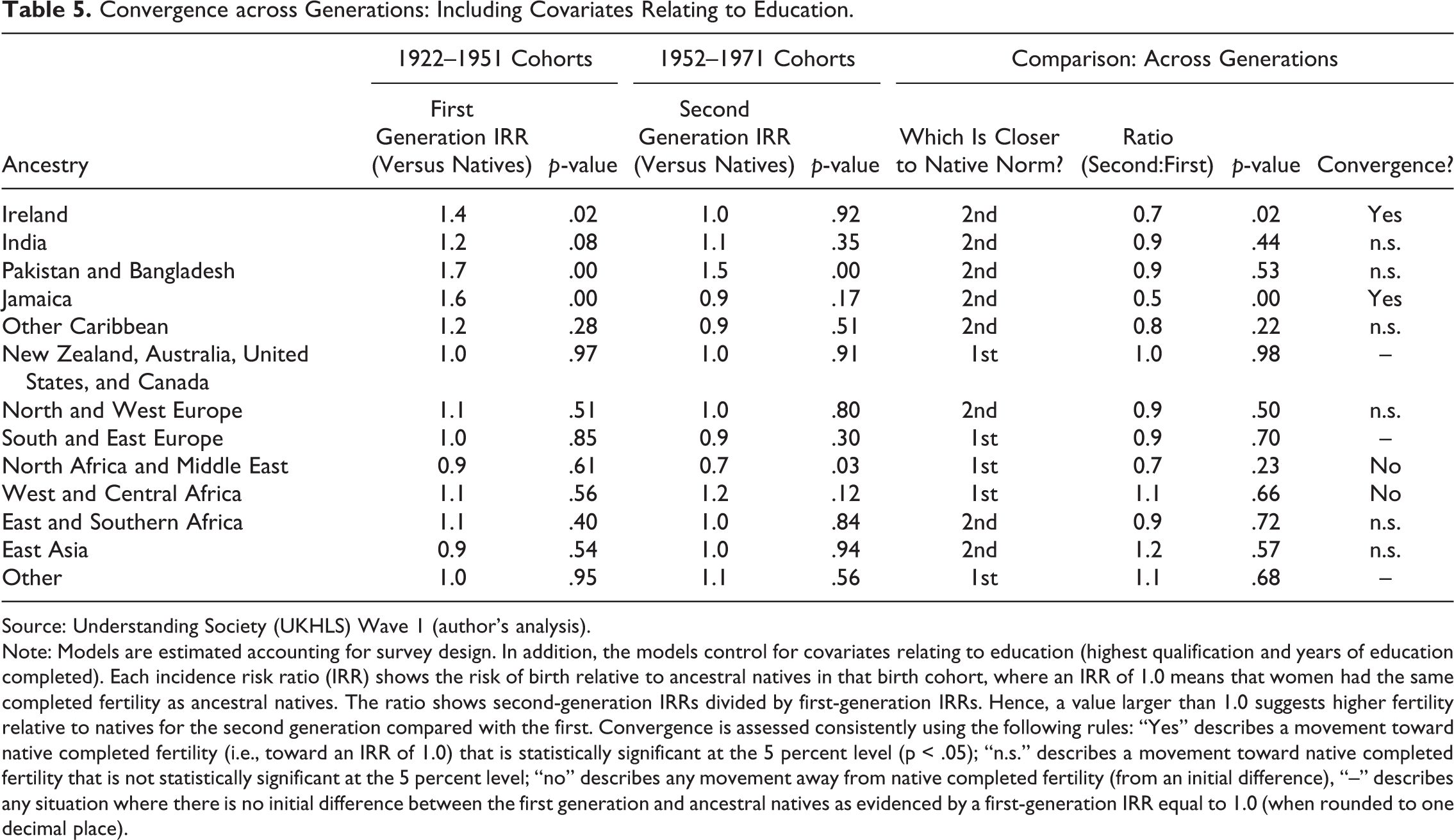
Source: Understanding Society (UKHLS) Wave 1 (author’s analysis).
Note: Models are estimated accounting for survey design. In addition, the models control for covariates relating to education (highest qualification and years of education completed). Each incidence risk ratio (IRR) shows the risk of birth relative to ancestral natives in that birth cohort, where an IRR of 1.0 means that women had the same completed fertility as ancestral natives. The ratio shows second-generation IRRs divided by first-generation IRRs. Hence, a value larger than 1.0 suggests higher fertility relative to natives for the second generation compared with the first. Convergence is assessed consistently using the following rules: “Yes” describes a movement toward native completed fertility (i.e., toward an IRR of 1.0) that is statistically significant at the 5 percent level (p < .05); “n.s.” describes a movement toward native completed fertility that is not statistically significant at the 5 percent level; “no” describes any movement away from native completed fertility (from an initial difference), “–” describes any situation where there is no initial difference between the first generation and ancestral natives as evidenced by a first-generation IRR equal to 1.0 (when rounded to one decimal place).
In addition to Table 5, the same models were run with the inclusion of controls for partnership history. These models are not shown, largely because of concerns relating to endogeneity (i.e., that fertility and partnership are strongly interrelated processes). Nevertheless, it is worth noting that these models also show very few differences from the results shown here. The most material change is that the conclusion for Irish convergence changes from significant to nonsignificant evidence of convergence, although this is entirely driven by a change in the p-value of the comparison of Irish generations from .04 to .06. In addition, there is some evidence of childhood socialization for Indians because first-generation completed fertility becomes significantly higher than the native norm. The only other material difference is that there is some evidence of cultural entrenchment for women from Western and Central Africa due to a significantly higher completed fertility than natives for the second generation. This finding suggests that second-generation women with Western and Central African ancestry may differ markedly, on average, from natives in terms of their education and partnership and that these social characteristics may be important explanations for their fertility.
Conclusion
The findings of this research show that patterns of generational fertility convergence vary considerably between origin groups in the United Kingdom. They show that it is impossible to summarize evidence for and against the intergenerational assimilation of completed fertility without specifying which origin group is being discussed (at least in the case of the United Kingdom). One of the main conclusions is therefore to highlight the importance of accounting for origin heterogeneity not only in appraisals of assimilation but also with respect to discussions about the likely contribution of immigrants and their descendants to long-term population growth. The results clearly show that this contribution varies considerably for different (ancestral) origins.
Although there is some evidence of generational convergence for a number of ancestral groups, there is only significant evidence for two groups — Irish and Jamaicans — which lends support to the childhood socialization hypothesis, thereby suggesting that second-generation Irish and Jamaicans may be adopting mainstream fertility norms because they spend their childhood in the United Kingdom. As indicated by the results from previous research (e.g., Coleman and Dubuc 2010), changing norms in the ancestral origin country may also help explain why convergence is observed for the second generation. Additionally, these findings align with previous results for the United Kingdom that show child migrants have more similar fertility to natives if they arrive in England and Wales at younger ages, thereby increasing their relative exposure to native norms (Adserà et al. 2012). It is interesting to note, however, that while Adserà et al. (2012) obtain results for Irish child migrants that are not significantly different from natives, similar to the results for the second generation obtained here, this is not the case for Jamaican child migrants, who have significantly different fertility from natives (albeit using a different fertility measure). As such, convergence patterns for the children of Jamaican immigrants may differ according to whether they are or are not born in the United Kingdom.
In contrast to this evidence of childhood socialization, there is evidence of cultural entrenchment for some ancestral groups. Most notably, there is evidence that the descendants of immigrants from Pakistan and Bangladesh have higher completed fertility than the native norm (Tables 3 and 5), suggesting that fertility differentials persist across generations for these origins. This result aligns with findings from previous studies that show the period fertility of second-generation Pakistani and Bangladeshi women is higher than that of natives (Dubuc 2012, 2016) and that the same is true for parity-specific birth rates (Kulu et al. 2015, 2017; Kulu and Hannemann 2016). The most common explanation for this general finding is the influence of cultural factors, with recent evidence suggesting that residential segregation may be a determinant of cultural entrenchment for the descendants of immigrants from these two origin countries (Wilson and Kuha 2017). However, it remains to be seen whether generational fertility convergence occurs for third or later generations.
The remaining ancestry groups — other than Pakistanis, Bangladeshis, Irish, and Jamaicans — are classed as having nonsignificant evidence, evidence of no convergence, or no initial difference (for the first generation). In the latter case, this might be due to adaptation or selection. For example, Indian women’s completed fertility is not significantly different from UK natives for either generation, which may be due to the selection of Indian women (into migration) who have similar family size norms to women in the United Kingdom. Alternatively, it may be due to changes in their norms after immigration. Further research would be required to disentangle these two explanations.
Another interesting finding is that only one origin group — North Africa and the Middle East — shows evidence of significantly lower completed fertility than the UK native norm for the second generation. Given the lack of a significant difference for the first generation, this is hard to interpret as evidence of cultural entrenchment (because there is no initial difference that can remain entrenched). One explanation for this result is that it may be due to segmented assimilation, for example, based on social inequality, which in turn may reduce opportunities for childbearing. However, further research would be needed to state this with any certainty. Moreover, one measure of inequality that cannot explain this result is education. As shown in Table 5, there are no material changes to the results after accounting for education (highest qualification and years of education completed), with the exception of a minor change for women from Western and Central Africa.
Beyond those already mentioned, there are several reasons why the results shown here should be treated with caution. The sample includes only those women who were resident in UK households between 2009 and 2011. It therefore excludes women who died or emigrated before these dates, making it difficult to generalize the findings backwards in time, which is important when trying to evaluate the historical impact of immigrant fertility on population size. As noted by previous research, any conclusions and generalizations must be tempered by this consideration (Andersson and Sobolev 2013). Similar caution is required when trying to make inferences about the behavior of future cohorts of immigrants and their descendants, in particular when trying to project the future population or make assumptions about future population change.
Another caveat is that the results are subject to uncertainty in a number of ways, not least the significance thresholds chosen to help communicate the findings. Given that much of the uncertainty in the results can be attributed to sample size, it is recommended that future data collection includes an effort to make available larger samples, including (at minimum) the variables that would enable a similarly detailed analysis. One further limitation of this study relates to its focus on completed fertility, which means that the results cannot be used to infer differences in the timing of births. Immigrants and their descendants from a given origin may share the same completed fertility but have very different profiles of childbearing. For example, recent research has shown that the birth rates and fertility differentials of second-generation Caribbean women in the United Kingdom differ considerably by parity, with evidence of second-birth risks that are lower than those of natives but third-birth risks that are higher (Kulu et al. 2015, 2017). Further research would be required to examine the link between this parity-specific pattern and the completed fertility results presented here.
Despite the aforementioned limitations, it is clear from the findings that assimilation does not occur equally for different ancestral origin groups, at least not with respect to completed fertility in the United Kingdom. For the United Kingdom, the results provide guidance for future research, highlighting the combination of groups and explanations that warrant further investigation. They also provide the first intergenerational comparison of completed fertility for immigrants and their descendants in the United Kingdom, which is important because completed fertility is the only measure that directly indicates the eventual contribution that these groups make to population size.
Beyond the United Kingdom, the results have significant implications, not least because they highlight the importance of origin heterogeneity and the problem with generalizing both within and between countries. On the one hand, this research shows that intergenerational assimilation is not necessarily a “standard” or “straight-line” process, at least not for different origins within the same country. On the other hand, it also shows that evidence concerning intergenerational assimilation in other contexts — such as evidence for Hispanics and Mexican Americans in the United States (Parrado and Morgan 2008) — does not necessarily generalize to other contexts, at least not for all origin groups. This implication has ramifications beyond the study of fertility and applies to any topic in migration research that is dominated by studies of particular origins or destinations.
Finally, this research makes a methodological contribution. It shows how a statistical test of generational convergence can be combined with a comparison of ancestral groups, thereby allowing different patterns of convergence to be distinguished. Within the limits of a descriptive study, this approach generates evidence that can be used to falsify hypotheses (e.g., childhood socialization). The ability of this approach to estimate differences in fertility across generations for different origin groups and including statistical uncertainty could be of material benefit to future research on intergenerational change. It could be used not only to study outcomes other than fertility but also to design research that compares the intergenerational assimilation of multiple outcomes for the same population using the same research design. The value of such an approach is that it would help unify the disparate fields of research on integration and assimilation.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received the following financial support for the research, authorship, and/or publication of this article: Economic and Social Research Council, Grant/Award Number: ESRC Studentship 1014032.