
Abstract
Botanical remains from dated Native American archeological contexts in the greater Upper Delaware Valley region are cataloged. This foundational chronology is necessary for future research related to paleoenvironments, human management of the environment, and the transition to a mixed farming, gathering and hunting subsistence economy. The chronologies of select botanicals and their relative importance for understanding Native American lifeways are highlighted.
Introduction
Native American peoples of the Middle Atlantic and Northeast regions historically used plants for a variety of purposes in addition to subsistence including: medicines, hygiene, domestic activities, ceremonial agents, structures, watercraft, tool and implement production, crafts, clothing, and adornment (cf. Hill and Rementer, 2015; Moeller, 1992: Appendix; Moerman, 1986: 816–818, 1998; Newcomb, 1956; Tantaquidgeon, 1977). Ethnohistoric and ethnographic sources are useful starting points for developing different approaches to the analysis and interpretation of paleobotanical assemblages from archeological sites. Understanding pre-Contact plant use by native peoples includes a consideration of the environmental and archaeological contexts of finds, and the chronology and frequency of artifact types often implicated in plant use (e.g., fire cracked rock, pitted stones, mullers, edge ground cobbles, mortars and pestles, pottery, and stone bowls). The analysis of residues on artifacts, pollen, and phytoliths has, and continues to be significant for illuminating the use history of plants.
The Native American transition from a hunting and gathering way of life to farming domesticated plants like maize, beans, and squash (the “Three Sisters”) is an issue of local, regional, and national significance. The probable management and intensive use of wild plant resources prior to the adoption of cultigens like the Three Sisters may itself represent a form of low level food production (Smith, 2001) and facilitated the adoption of cultigens. Environmental management and the adoption of farming would have had an impact on the character of the environment (e.g., Stinchcomb et al., 2013), essentially making it one that humans had “domesticated.” These are some of the issues that need to be explored in depth with existing data and additional research, and are part of the grand challenges for archaeology (Kintigh et al., 2014: 15–18).
An alternative mitigation project sponsored by the New Jersey Historic Preservation Office (Stewart, 2018) provided the opportunity to gather botanical data for the Upper Delaware Valley dispersed through formal publications and the large number of cultural resource management studies on file for the area and assess its potential value for furthering an understanding of Native American lifeways. For the purposes of this endeavor the Upper Delaware Valley is defined by portions of the drainage basin that exist in the following states and counties: Warren and Sussex counties, New Jersey; Orange, Sullivan, Delaware, and Broome counties, New York; and Northampton, Monroe, Pike, and Wayne counties, Pennsylvania (Figure 1). The defined area is much larger than what has often been considered as the Upper Delaware by archaeologists in the past. For example, past definitions of the Upper Delaware have bounded it by the Delaware Water Gap on the south and Port Jervis to the north (cf. Custer, 1996, Kinsey, 1972; Kraft, 2001). The Pennsylvania State Historic Preservation Office includes Northampton County in what it considers to be the Upper Delaware Valley, which extends the geographic boundary well south of the Water Gap. I have included Northampton County here to complement the downriver extent of Warren County in New Jersey.
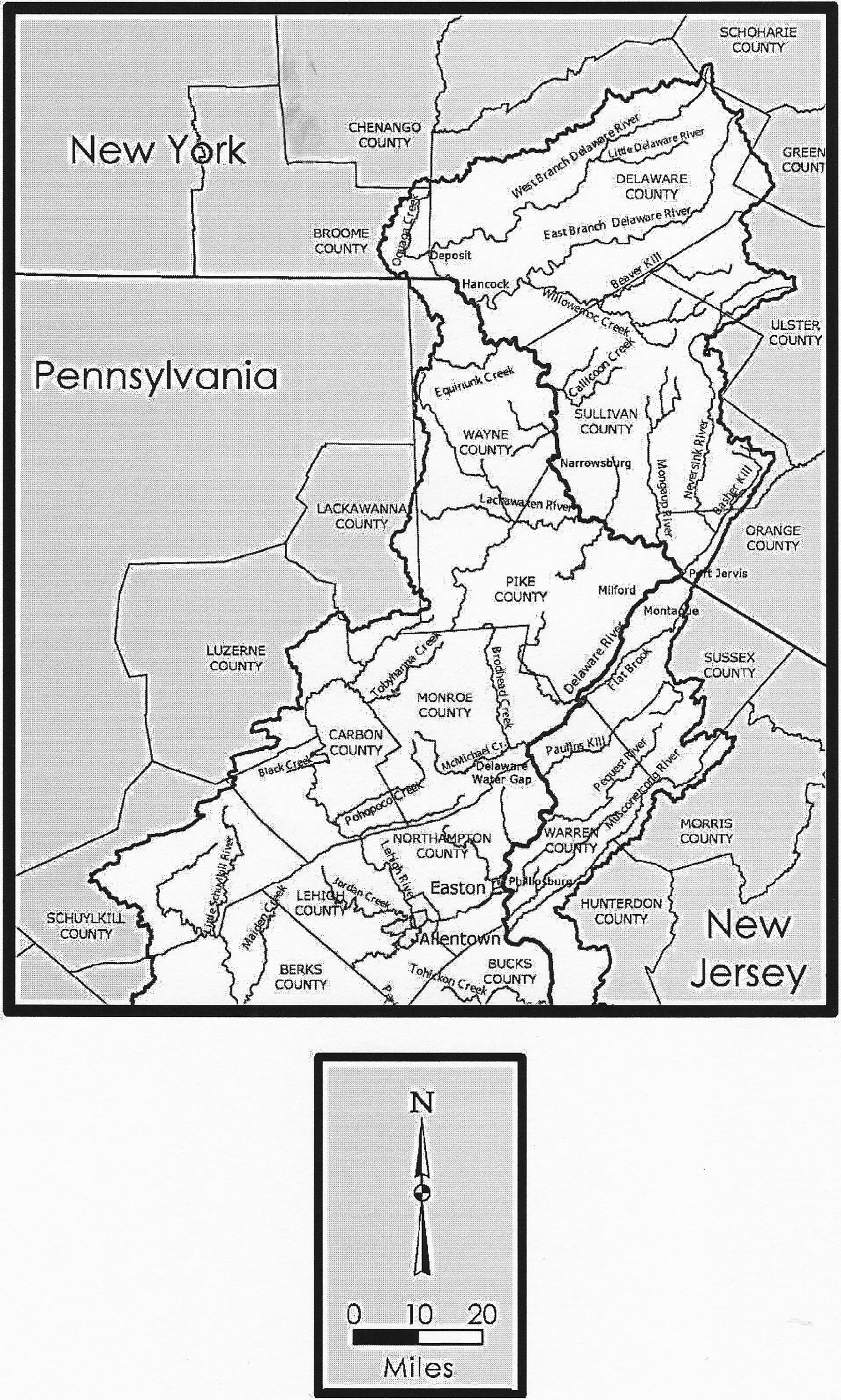
Upper Delaware Valley project area showing relevant countries of New Jersey, Pennsylvania, New York. Not labelled is Otsego County situated to the north of Delaware County, New York. Areas within the Delaware River Basin are shown in white. Modified from a map available from the Delaware River Basin Commission at http://www.state.nj.us/drbc/library/documents/maps/countries.pdf.
The study area encompasses portions of a number of physiographic provinces and related sections: the Reading Prong section of the New England Province, corresponding with the Highlands in New Jersey; the Ridge and Valley which includes the Great Valley and Appalachian Mountain (Blue Mountain, PA and Kittatinny Mountain, NJ) sections; and the Appalachian/Allegheny Plateau which includes the glaciated Low Plateau and Pocono Plateau sections in Pennsylvania, and the glaciated Low/Allegheny Plateau and Catskill sections in New York (Briggs 1999; New Jersey Geological Survey, 2018; Perles et al., 2007; Sevon, 2000; Wolfe, 1977: 204–243). The degree to which all, or portions of the larger area used in this project correspond with Pre-Contact cultural or group territories remains an open question contingent on a variety of diachronic analyses. Evaluating data from a broader geographic context is necessary to examine existing assumptions about the correspondence of natural and cultural area, sand to better understand patterns of social interaction and cultural change.
The list of botanical remains from dated contexts in the study area is surprisingly substantial (Table 1). This compilation builds on previous syntheses for the Upper Delaware and broader region (e.g., McConaughy, 2008; Messner, 2008, 2011; Messner et al., 2008; Stewart, 2015a; Stewart et al., 2015; Stinchcomb et al., 2011). The previous work of Stewart (2015a) and colleagues (Stewart et al., 2015) incorporates data and insights generated as part of the current project. The chronologies of select botanicals and their relative importance for understanding Native American lifeways are highlighted below.
Radiocarbon dates associated with botanical remains.

aCalibrated with Intcal13.14c data (Reimer et al., 2013) using Calib 7.10 (Stuiver et al., 2017; cf. Stuiver and Reimer, 1993). Calibrated 2 sigma age ranges represent the greatest relative area under the probability distribution curve, generally areas between .90 and 1.00.
bItalicized and bold indicate direct AMS date of botanical.
Common and taxonomic names employed in Table 1 follow those used in the reports in which they were described. Where family, genus, or species was not designated in a reference they were assigned where feasible by the current author. The radiocarbon foundation for the occurrence of the Three Sisters in the study area has been addressed elsewhere (Stewart, 2021).
Botanical remains that were directly dated using AMS are shown in bold in Table 1. Unidentified nuts are listed in entries as they are useful information regarding subsistence and the seasonality of occupations, even in the absence of an assignment to genus or species. Unidentified seeds are not included in entries because the information that they provide is equivocal, other than as a sign of the preservation potential of a deposit. Genus/species represented by wood are distinguished in the listings from other elements of the same genus/species. For example, the listing of hickory indicates the occurrence of nutshell. The appearance of hickory in the “wood” portion of an entry refers only to the presence of charred wood.
Messner (2008: Tables 1, 3, 11; 2011: Tables 2.2-2.4) previously compiled lists of macrobotanical remains from archaeological and ethnohistoric contexts for the Delaware River watershed. Table 1 represents a more comprehensive catalog. Dent (1985), Goldman (1975), and Moeller (1992) provide lists of current plant species in the Upper Delaware for comparative purposes. Table 2 is a summary of paleoenvironmental changes for the study area and serves as general environmental background for some of the discussion that follows.
Summary of paleoenvironmental changes, Upper Delaware Valley. a
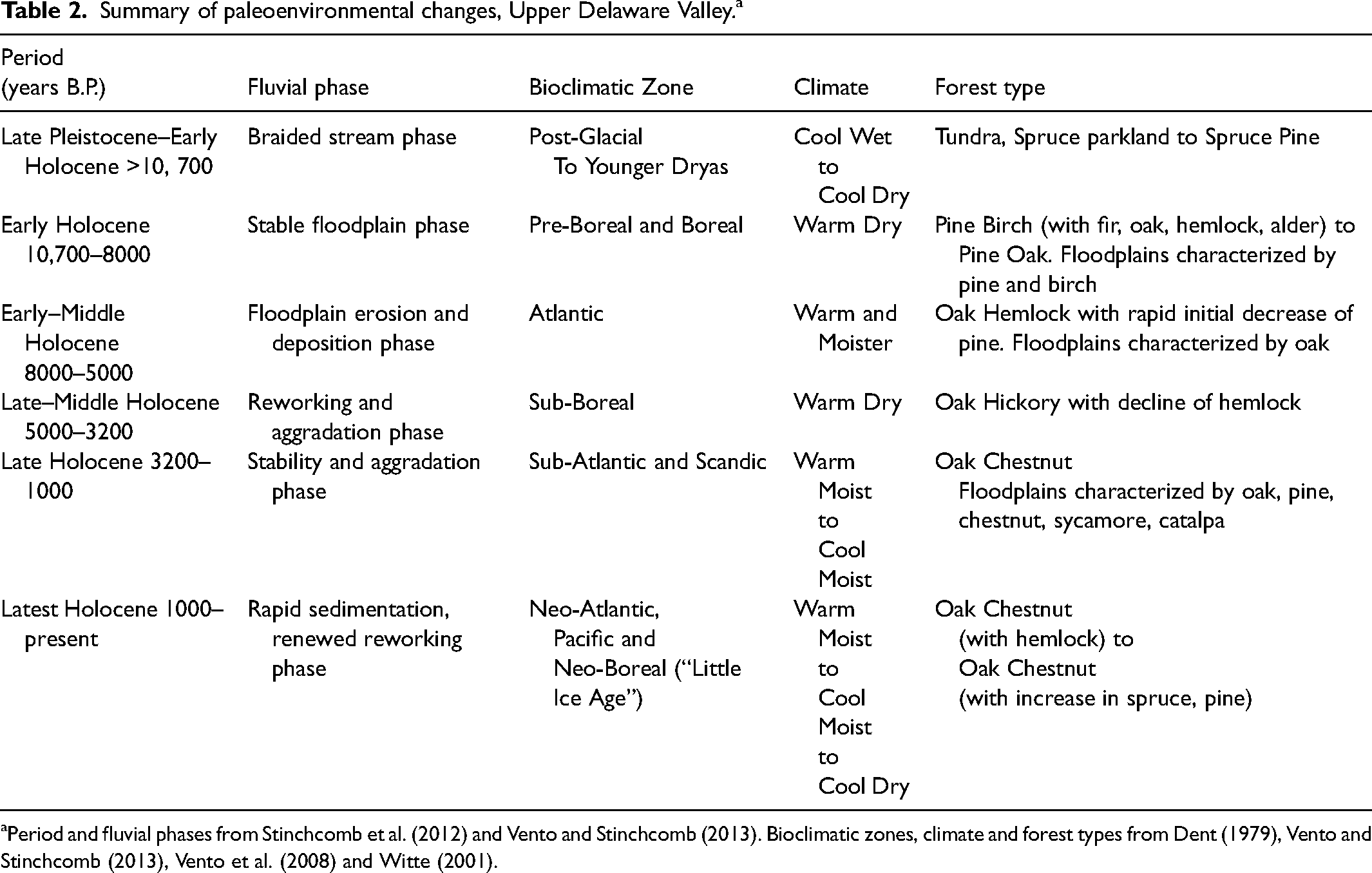
aPeriod and fluvial phases from Stinchcomb et al. (2012) and Vento and Stinchcomb (2013). Bioclimatic zones, climate and forest types from Dent (1979), Vento and Stinchcomb (2013), Vento et al. (2008) and Witte (2001).
Chronological trends and issues
Botanical remains from the deeply stratified Shawnee Minisink Site not only anchor the diverse use of plants in Paleoindian times, but reveal the consistent use of a variety of plants through time (Dent and Kaufman, 1985). Recoveries from other dated contexts in the Upper Delaware support this trend.
What becomes important for future research is determining the relative importance of individual items to various cultural practices over this same span of time.
Hickory has the longest dated use history of the nut- and acorn-bearing trees represented in the archaeological record, first appearing in the Paleoindian deposits at Shawnee Minisink. However, its next dated occurrence isn’t until 4710±40 BP (3495 BC calibrated median), a time during which hickory has become well represented in area forests (see Table 2). As yet unidentified nutshell has been found in Paleoindian contexts at the Snyder Complex (Rankin and Stewart, 2016; Stewart and Rankin, 2018). Hazelnut and butternut are more consistently represented in assemblages of Early and Middle Archaic age. Chestnut first appears in a dated context at 3495 BC and acorn at 3046 BC (calibrated medians). Of course any “trends” seen in the small number of dated contexts with mast for Paleoindian, Early and Middle Archaic times frames could easily be overturned with the addition of a handful of new finds. Black walnut (Juglans nigra) is not specifically identified until 884/513 BC (calibrated medians) although it could be represented by a number of earlier finds that only could be identified to the level of genus (Juglans sp.).
Given the longevity of oak in regional environments, and the use of acorns documented in historical sources, it is interesting that acorns appear later than other sources of mast in the archaeological deposits of the Upper Delaware. An AMS date of 9530 ± 60 BP (Beta 81355) on carbonized acorn from the Steele site in southern New Jersey is more in line with expectations (Stanzeski, 1996: 44, 1998: 45). The wood of white and red oaks is much more well represented in dated deposits and might indirectly reflect the consistent use of acorns beginning during the Late Archaic. The under-representation of acorns in the archaeological record of the broader Eastern Woodlands has been noted by many authors (Messner, 2011: 17).
The more frequent appearance during the Early, Middle, and Late Archaic periods of tools and fire cracked rock that could be implicated in the processing of mast suggests that mast had gained in importance. Truncer (1999: 265–279) proposes that archaeologists test the hypothesis that steatite bowls were used in the processing of acorns, based on his analysis of residues and the spatial distribution of bowls and mast-producing forests. Dates that can be assigned to steatite vessels predominantly fall within a Late Archaic to Early Woodland time frame (Truncer 2004: Table 2).
It is during a Late Archaic to Early Woodland time frame that regional prehistories characteristically acknowledge the fundamental importance of mast and plant foods to native lifeways (e.g., Carr, 2015: 68–69; Custer, 1996: 213–214; Kraft, 2001: 111–115; Kraft and Mounier 1982: 68; Messner, 2008: Table 11, 2011: 115–123; Stewart, 2015b:13; Williams and Thomas, 1982: 107). The intensive use of plant resources during this time corresponds with inferred reductions in settlement movements and territory size and a trend in seasonal, semi-sedentary types of settlements focused on riverine settings. The systemic relationship of these behaviors is complex (Stewart, 2015b:14): Whatever is driving the emphasis on riverine settings and the creation of semi-sedentary settlements, the ecology of the situation could reinforce economic behaviors already in place, that is, the use of mast and other plant resources. Prolonged habitation at a fixed location would eventually lead to a decline in the availability of game within the foraging radius of the settlement. This in turn might lead to an increase in the importance of more sustainable plant resources, an intensification of their use, and a greater investment in storage facilities to offset subsistence risk. The importance of fish and small game also might be enhanced as would relevant capture and processing technologies.
Wood charcoal representing mast-producing trees, common in archaeological deposits dating circa 3000 BC and thereafter, could be an indirect reflection of the use of mast in addition to the preparation of medicines. The use of the bark of walnut and oak to make medicines to treat a number of ailments has been noted for the Delaware (Hill and Rementer, 2015: 12; Newcomb, 1956: 71).
Macrobotanical assemblages from dated contexts become more frequent during a Late/Transitional Archaic time frame and intensify thereafter, an observation in-line with cultural historical narratives regarding the growing importance of plants in Native American lifeways. However, it is difficult to ignore the probable impact that taphonomic and site formation processes have had on the preservation and patterning of this evidence. The botanical assemblage from the deeply buried Paleoindian deposits at the Shawnee Minisink site show what is possible when conditions are favorable.
Caveats aside, data from the contexts listed in Table 1, as well as botanical assemblages from contexts dated by association with diagnostic artifacts, support the interpretation of the consistent use of seeds, fruits, and greens during a Late/Transitional Archaic time frame and continuing throughout the Woodland era. Assemblages dating to the Late Woodland period are the most extensive with those from 36Pi4, 36Pi136, and BRO-117 (Broome County, New York) being exceptional examples.
Of special note is the limited occurrence of hog peanut. Hog peanut is found in a dated late Middle Woodland context at the Deposit Airport I site (Delaware County, NY) and in a dated Late Woodland context at the Manna site (36Pi4). Historically, the seeds and tubers of hog peanut were subsistence items in the region (Messner, 2008: 42, Appendix A) but also figure into the production of native medicines (Moerman, 1986: 32–33). The sole occurrence of giant ragweed (Ambrosia trifida) in a Late Woodland context at the Manna site is also of interest. Giant ragweed was once thought to be a deliberately cultivated species in the Eastern Woodlands (Cowan, 1985: 214–217) and its continued occurrence in contexts with other domesticates “argues for its candidacy as a crop” (Smith and Cowan, 2003: 112).
Messner (2008, 2011) and colleagues (Messner and Dickau, 2005; Messner et al., 2008) have recovered starch grains from a variety of tools and implements that complement macrobotanical inventories for the Early, Middle, and Late Woodland periods of the Delaware River watershed. For the Upper Delaware starch representing maize, acorn, possible little barley, the legume or bean family, and the true grasses family has been recovered from implements found in contexts dated to the Late Woodland period (see Table 27). Maize starch also has been found on implements attributed to undated Late Woodland contexts (Messner, 2008: Figure 129), as has acorn and possibly little barley (Messner et al., 2008; Messner, 2008: 314). In a context likely Middle Woodland in age at the Manna site, starch grains from two different taxa were isolated from a triangular biface consisting of wild potato/yam (Dioscorea villosa) and maize (Messner, 2008: 316; Stewart et al., 2015: 172–173, 294, Figure 62).
In a summary of early plant domestication in eastern North America, Smith (2011: Table 1, Figure 1) lists the earliest dates for a series of indigenous domesticated seed crops. They derive from sites in the Mississippi river basin of Arkansas, Illinois, Kentucky, Missouri, and Tennessee, and fall within what would be a Late Archaic time frame: -Squash (Cucurbita pepo ssp. ovifera) at 4440 ± 70 BP; -Sunflower (Helianthus annuus) at 4265 ± 60 BP; -Marshelder (Iva annua) at 3920 ± 40 BP; and -Chenopod (Chenopodium berlandieri) with dates from 3490 ± 40 to 3400 ± 150 BP.
Plants whose domestication is in doubt but appear to have been subject to deliberate planting, harvesting and storage of seed stock include: erect knotweed (Polygonum erectum), little barley (Hordeum pusillum), and maygrass (Phalaris caroliniana; Smith, 2011: 472). Together with the listed domesticates these plants comprise what has been termed the Eastern Agricultural Complex.
None of the indigenous domesticates noted by Smith have been found in comparably early contexts in the Upper Delaware or the broader river valley as Messner (2008: 290) points out in his basin-wide synthesis. In the Susquehanna Valley of Pennsylvania potentially domesticated squash (Cucurbita pepo) has been AMS dated to 5404 ± 552 BP and 2625 ± 45 BP (Hart and Asch-Sidell, 1997: 525, 527, 531; Hart et al., 2007: 579). McConaughy (2008: Table 2–1; 2015: 35) notes the connection of cucurbit specimens with dates of 3065 ± 80 BP, 2820 ± 75 BP, and 2815 ± 80 BP at the Meadowcroft Rockshelter in southwestern Pennsylvania, but expresses caution in accepting the validity of the associations. In western New York squash phytoliths are dated to 2905 ± 35 BP at the Scaccia site (Hart et al., 2007: 579, Tables 1, 6).
A seed of potentially domesticated chenopodium from the Calver Island site (36Da89) in the Susquehanna Valley of Pennsylvania is AMS dated to 3980 ± 40 BP (Miller et al., 2007: 75–76). Other seeds from the same context have attributes that are suggestive of a cultivated form. The first domesticated chenopodium in western Pennsylvania occurs during a Middle Woodland time frame (McConaughy, 2008: Table 2-3; 2015: 36). Residue on a steatite bowl sherd from a Transitional Archaic occupation dated to ca. 3080 BP at Calver Island produced Hordeum/Elymus-type starches indicative of little barley or wild rye grass seeds (Miller et al., 2007: 73). Hordeum/Elymus-type starches also were identified on Transitional Archaic steatite vessel sherds from 36Pe16 in the Susquehanna Valley (Miller, 2015: 93; Miller et al., 2007: 76).
Maygrass dates to 2950 ± 100 BP at Oberly Island (36Nm140) in the southern portion of the study area in Northampton County, Pennsylvania. New Jersey, Pennsylvania, and New York are not part of the plant's current range (Messner, 2008: 290–291; 2011: 115, Figure 6.1; United States Department of Agriculture, Natural Resources Conservation Service017a, hereafter USDANRCS). Erect knotweed has not been found in the project area or on archaeological sites in the greater Delaware River basin (Messner, 2008, 2011). Erect knotweed and maygrass have been found in western Pennsylvania in dated contexts that fall within a Transitional Archaic time frame (McConaughy, 2008: Tables 2–4, 2–6).
Wild marshelder/sumpweed (Iva annua) is associated with an occupation dating ca. 2900 BP to 2150 BP at the Broome Tech site to the west of the New York portion of the Upper Delaware basin (Asch Sidell, 2008: 39, Tables 3.2, 3.3; 2011: 1057; Messner, 2011: 115). It has been found in a context dated to 2460 ± 130 BP at a site (7S-F-68) in the Lower Delaware Valley, and is believed to be a domesticated form (LeeDecker et al., 1996: 34, 136–138; Messner, 2008: 310). Additional finds of marshelder are from two later contexts (1020 ± 70 BP, 310 ± 80 BP) at the site. The site is located in an area where marshelder is not a part of modern plant communities (Figure 2; Messner, 2008: 290, Figure 128; Messner, 2011: 115).
Archaeological distribution of marshelder in the Delaware Valley and vicinity. The northernmost star represents the location of the Broome Tech, Chenango Point, and Chenango Point South sites the New York. Modified from Messner (2008: Figure 128).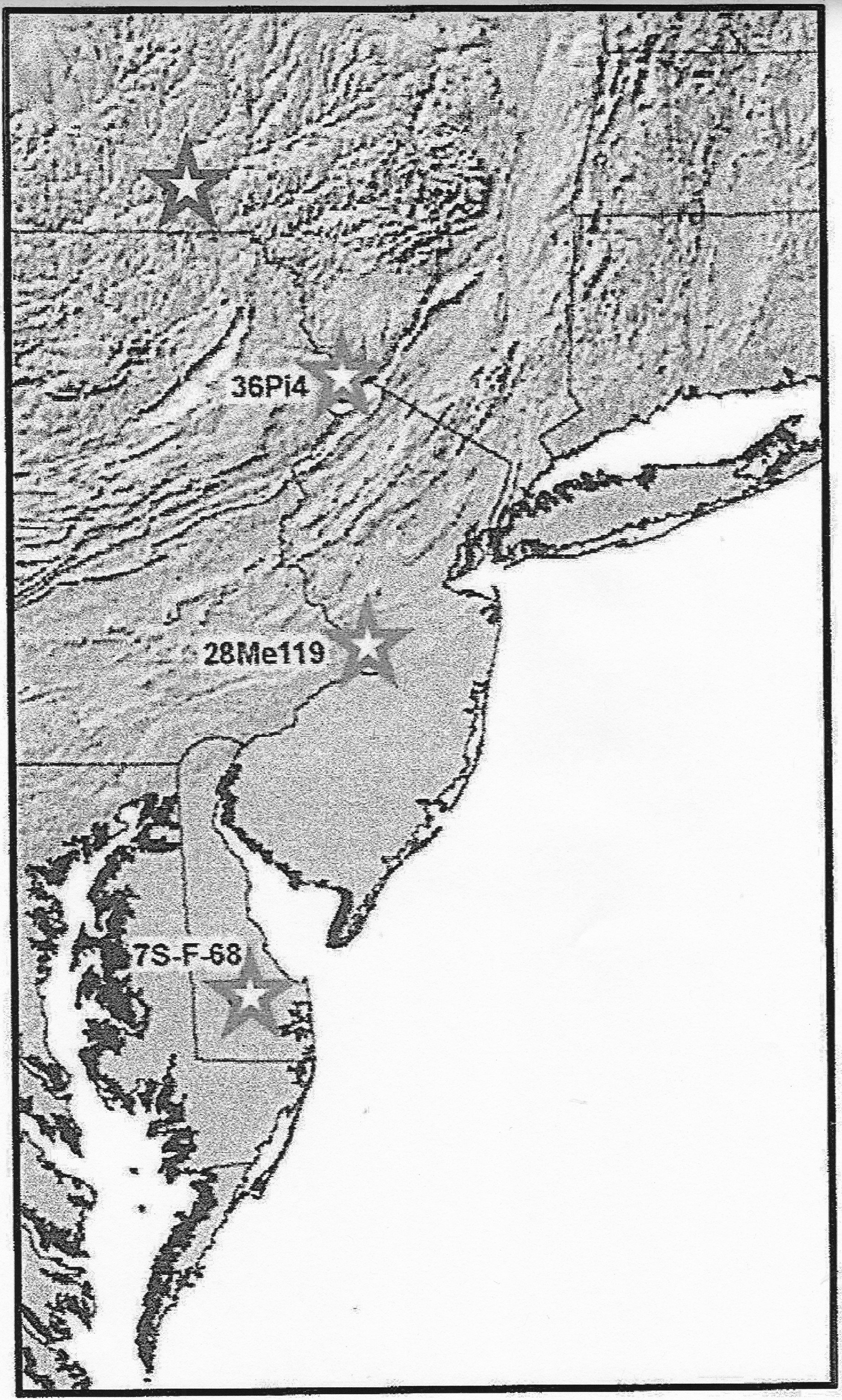
Domesticated plants associated with the Eastern Agricultural Complex are found in Late Woodland contexts in the study area. In the Upper Delaware Valley wild forms of Chenopodium (sp.) first occur in the Paleoindian deposits of the Shawnee Minisink site but don’t reappear in a dated context until circa 4460 ± 130 BP and 4370 ± 140 BP at the Shohola Flats site (36Pi169). After this time recoveries are much more frequent and continue into the era of contact with Europeans. A likely domesticated form of Chenopodium berlandieri is found at the Deposit Airport 1 site dated to 920 ± 40 BP and represents the easternmost occurrence of the cultigen (Asch Sidell, 2002d; 2008: 47, Table 3.3; Messner, 2011: 107). No other domestic forms of Chenopodium berlandieri have been identified in the Upper Delaware.
To the west of the project area dates for domesticated Chenopodium are slightly earlier but still relate to a Late Woodland time frame. In northcentral Pennsylvania at the Mansfield Bridge site (36Ti116) domesticated seeds of Chenopodium berlandieri were recovered from what is defined as an early Owasco occupation with dates ranging from 1360 ± 80 BP to 830 ± 40 BP (cf. Asch Sidell, 2003: Table 9; 2008: 46–47; 2012: 714; Wall et al., 2003: 13, 120, 129, Table 2.1). To the north of Mansfield Bridge in southcentral New York a component dated ca. 950 to 650 BP at the Scudder site also contained this domesticate (Asch Sidell, 2008: 46–47). Domesticated chenopodium is known from two components at the Memorial Park site in central Pennsylvania, the earliest of which has dates ranging from 1190 ± 40 BP to 1120 ± 60 BP (cf. Asch Sidell, 2008: 46–47; Hart and Asch Sidell, 1996: 17, Table 1).
Marshelder or sumpweed is found only at the Manna site (36Pi4) in the Upper Delaware Valley and was recovered from a feature dated to 840 ± 70 BP. Given its size it might represent a domesticated seed, but it is not possible to be certain on the basis of the single carbonized kernel found (Asch Sidell 2015: 374; Stewart et al., 2015: 197–198, 295). Marshelder was also found in an undated feature assigned to a Contact period time frame given the presence of colonial pipe fragments in the feature fill (Stewart et al., 2015: Table 115). The natural range of the plant does not include the Upper Delaware Valley, New Jersey or New York; the plant's distribution is limited in Pennsylvania (USDA NRCS, 2017b; Messner, 2008: 310; 2011: 108, Figure 5.12; Rhoads and Klein, 1993). The presence of marshelder in the Upper Delaware raises the possibility that native peoples were intentionally transplanting it to new areas and nurturing it in gardens or habitats adjacent to residential areas.
Approximately 30 miles west of the Delaware Basin in New York marshelder is found at the Chenango Point, Chenango Point South, and Broome Tech sites (see Table 1 and Figure 2; Asch Sidell, 2008: 39; Messner, 2011: 115). The date from Chenango Point, 920 ± 40 BP, is earlier than that for the Manna site specimen, while the others relate to a time frame similar to that indicated by the finds at Manna. The marshelder from Chenango Point and Chenango Point South appear to be domesticated forms (Asch Sidell 2011: 1060; 2012: 719–720; Miroff, 2014: 24). Even farther west, marshelder is associated at Mansfield Bridge with Features 13 and 17 dated to 1130 ± 80 BP and 1360 ± 80 BP, respectively (Asch Sidell, 2003: Table 9; Wall et al., 2003: 13, 120, 129, Table 2.1).
No confirmed finds of domesticated sunflower have been made in the Upper Delaware Valley although wild sunflower has been dated at 36Pi136 to 780+-/110 BP and 740 ± 80 BP. A sunflower or gourd seed is part of a dated feature (1300 ± 110 BP) at 36Nm15. Just to the west of the New York section of the Upper Delaware drainage, wild sunflower is associated with Late Woodland contexts at the Otsiningo Market, Park Creek II, and Chenango Point South sites (see Table 1). In the basin of the Upper Susquehanna River domesticated sunflower is dated to 940 ± 40 BP (Feature 2) at Mansfield Bridge (Asch Sidell, 2003: Table 9; 2012: 714; Wall et al., 2003: 13, 120, Table 2.1; contra Messner, 2008: 309; 2011: 107) and ca. 650–500 BP at the Thomas/Luckey site, in addition to specimens of wild sunflower (Asch Sidell, 2008: Table 3–6; Knapp, 2002: 171, 189, Tables 9.1, 9.4, End Note 5; Miroff, 2009: 77).
Little barley is another seed plant of the Eastern Agricultural Complex only found in dated Late Woodland contexts in the Upper Delaware Valley at the Manna site and Ventura Tract, and to the west in New York at Chenango Point (see Table 1). Not far from Manna, little barley starch was detected on an artifact from an undated context at the Loch Lomond site. Two stemmed points were associated with the same deposit and could potentially represent an occupation pre-dating the Late Woodland (Messner, 2011: 93, Table 5.1, Figure 6.1; Messner et al., 2006). Little barley occurs in a variety of Late Woodland contexts at the Memorial Park site on the West Branch of the Susquehanna River (Asch Sidell, 2008: 46–47; Hart and Asch Sidell, 1996: Table 2). Of significance is the fact that little barley does not occur naturally in New Jersey, Pennsylvania, or adjacent portions of New York (USDA NRCS, 2017c).
Discussion and prospects for further analysis
Determining the relative abundance of hickory, hazelnut and butternut (Juglans cinerea) in comparison with black walnut (Juglans nigra), chestnut and acorn-bearing species in early and middle Holocene environments would be informative. If the representations are fairly equitable one might conclude that hickory, hazelnut and butternut are intentionally selected as a matter of cultural preference. The habitat requirements and productive cycles of the trees also would need to be considered, as would the changing nature of climate and environment in terms of the representation of useful trees. For example, hazelnut trees can’t survive as an understory tree and must mature in relatively open areas. Walnuts don’t occur in true stands. Chestnut trees would be more concentrated in upland areas but also found in lowland settings. Hickory occurs in both lowland and upland settings. Settlement patterns and plant processing technologies must be considered in conjunction with the habitats of mast-producing species in order to explain the use history of a particular genus/species.
In speaking of the prehistory of plant use by the native peoples of the Eastern Woodlands Smith (1978: 114) notes:
Of all the wild plants, nuts as a group were certainly the most important food. They are more abundant than any grains or fruits, are easier to harvest, and contain more calories and protein per gram of food. However, nut trees have one major drawback: they do not produce a consistent crop every year.
The productivity of mast-producing species is a key variable in understanding the intensity with which this resource was exploited by native peoples. Limited control over mast productivity may be one reason why seed-bearing plants figure more prominently in discussions of the evolution of food producing systems. White oak acorns mature in a single year while those of red oaks require two years. Chestnut trees produce a reliable mast every year, hickory productivity varies on a two year cycle, and beech productivity varies widely over 2–3 year cycles (cf. Messner, 2011: 16; Petruso and Wickens, 1984; Scarry, 2003: 57–67, Table 3.2; Talalay et al., 1984).
Of the mast species represented in the Upper Delaware and broader region, butternut and black walnut have the greatest nutritional value in terms of calories, protein and fat, while acorns and chestnut are outstanding sources of carbohydrates (Messner, 2011: Table 2.1; Scarry, 2003: 64, Table 3.3). Nut/acorn meats as well as oils that can be rendered from them are potential foods requiring varying investments of labor depending upon the type of processing and storage involved (Briggs 2015: 321; Messner, 2011: 12–18; Scarry, 2003: 57–67). The ease of getting at nut/acorn meat is probably greatest for white oak acorns and hazelnuts, which have relatively thin shells and can be eaten or used in cooking without further processing (Scarry, 2003: 57–67). Given the time and labor investment, the growing emphasis on mast and other plant resources may have stimulated the trend in settlements of longer duration. Chestnuts and acorns are typically dried before being further processed or stored. Red oak acorns require more extensive processing than white oak acorns because of higher levels of tannic acid. The difficulty in separating the nut meat of hickory from its shell makes rendering its oil with water after crushing the nut a more productive endeavor, or using the crushed product to make stock for soup or stews. Most forms of mast could be rendered for oil which generally involves the use of hot or boiling water. (cf. Briggs 2015; Messner, 2008, 2011; Ortiz, 1991). All of these approaches to the use of mast or its storage have implications for occupation spans of days, weeks, or more at a given location.
A number of botanicals representing potential foods and medicines rarely occur on more than one site in study area: black cohosh, bracken fern, buckbean, bunchberry, burclover, carrot or parsley family, chickweed, cinquefoil, clover, coneflower, false pennyroyal, hackberry, horned pondweed family, ironwood, mint, morning glory, mulberry, plantain, poppy family, spike rush, toadflax, tupelo, tupelo/black gum, violet, winter cress, and wormwood.
There is tantalizing evidence that along with wild local species, native peoples in the region were cultivating wild economic species brought in from elsewhere. It is probable that this activity took place prior to Late Woodland times; however, evidence is best associated with the Late Woodland when an indigenous complex of economic plants is being used in conjunction with squash, maize, and eventually beans (Asch Sidell, 2008: 46–47; Messner, 2011: 115–131). This is certainly the case in the Upper Delaware Valley. What may have changed with the consistent use of maize is how and where the other species were planted (cf. Doolittle, 2001; Keeley, 1995; Messner, 2011: 125; Smith, 2001; Smith and Cowan, 2003). Squash, maize and beans, the “Three Sisters,” have traditionally been considered the base of Late Woodland farming in the region. It is now clear, however, that the use history of each of these cultigens is not identical, and that wild and likely cultivated indigenous plants continued in use from an earlier time (e.g., Hart, 2011: 104; Messner, 2011: 115–131).
When considering the long period of time over which Native peoples in the Upper Delaware and broader region were using plants, it is difficult to see the adoption of maize, beans, and squash as revolutionary. Pre-existing subsistence systems were already geared for including new items into an already extensive plant inventory. The variable degree to which new items would come to be relied upon likely results from how readily they can be accommodated with existing gardening and landscape management practices, and the existing productivity of the environments in which a group has long resided and exploited (cf. Doolittle, 2001; Messner, 2011: 27–39, 123–127; Wagner, 2003). Maize and the plants of the Eastern Agricultural Complex have different life cycles with the cultivation of maize requiring more attention (Smith and Cowan, 2003: 118–121). Once farming is in place shifting gardens/fields serves to revitalize soil but also allows other useful indigenous plants to grow in the plot left fallow while domesticates thrive in the newly cultivated field (Wymer, 2015).
We have yet to explore the degree to which foodways reflect unique cultural preferences, rather than a strictly mechanistic or functional interaction of people with their environment. Introduced foodstuffs need to make cultural “sense” in terms of the existing cuisine and approaches to cooking (Wetterstrom, 1978). We have the ability to reconstruct prehistoric environments and identify things within it that are potentially useful to a people with a given level of technology. By comparing this baseline data with what the archaeological record indicates that people are selecting from the environment, we should be able to define cultural preferences regarding subsistence. Stable isotopy and trace element analysis of human bone will be critical in resolving outstanding issues regarding the importance of specific subsistence items and their impact on other behaviors.
Documenting and refining the chronology of plant use in accordance with the latest approaches in paleoethnobotany (e.g., Van Derwarker et al., 2016) remains an important endeavor for ongoing research. Direct AMS dating of botanical remains is essential for chronological clarity and should be linked with precise or renewed identification of remains. The reanalysis of maize from the Holding site in Illinois (Simon, 2017) provides an object lesson. What were initially identified as a fragment of a maize kernel and a cob fragment were AMS dated to 2107 ± 50 BP and 2077 ± 70 BP respectively (Simon 2017: 140). Long thought to be the oldest directly dated maize in the Eastern Woodlands, analysis of the stable carbon isotopes of these and other specimens from the site were shown to not be maize (Simon, 2017: 142–144). Simon (2017: 146) recommends that evaluating carbon isotope ratios using stable isotope ratio mass spectrometry prior to AMS dating should become standard protocol.
What useful plants are attested historically (e.g., Hill and Rementer, 2015; Moerman, 1986, 1998; Tantaquidgeon, 1977) but not represented in the archaeological record of any period? For example, given the chronology of pipe use in the Delaware Valley and its likely importance in social interactions and ritual, there is no paleobotanical evidence of tobacco. Tobacco is known from pre-Contact contexts in the broader region (e.g., Hart and Asch Sidell, 1996; McConaughy, 2008: 12–13, Table 2–2; Miroff, 2014).
Programs of starch and phytolith analysis of residues on pottery and other artifacts should continue as exemplified by the work of Messner and colleagues, and Hart and colleagues. Macrobotanical remains are infrequently encountered on sites, even in the context of large cultural resource management projects, emphasizing the importance of making residue analysis of artifacts a standard procedure. Carbon isotope analysis of the sediments comprising feature fill can also be informative. Lattanzi and Stinchcomb (2015) used this method as an independent means of assessing what is currently known about the use history of maize in the downriver area of the Abbott Farm National Historic Landmark.
Related to mortuary features we need to ask how their botanical contents compare with those of non-mortuary features. Can the argument for graveside feasting, associated with historic and ethnographic practices, be supported (cf. Cushman, 2007; Dean, 1984; Newcomb, 1956: 39–43; Obermeyer, 2016, 2017; Obermeyer et al., 2015; Sieg, 2008: Table 5)?
Prior to the dated appearance of domesticates in the Upper Delaware Valley the intensive use of plants likely paved the way for the integration of domesticates into the subsistence system. The analysis of dental caries in skeletal populations pre-dating the first appearance of maize could offer support of this contention. AN example of this is provided by research conducted at the Island Field cemetery dated circa AD 400–1150 and AD 1200–1400 (Custer et al., 1990; Messner, 2008: 319–320). Island Field is located in northern Delaware (Lower Delaware Valley) and has no evidence of maize, beans or squash. Island Field burials exhibit high frequencies of dental caries interpreted as indicating reliance upon starchy, carbohydrate-rich seeds and tubers, a trait also typical of ancient maize farmers. In fact, when compared to a large number of skeletal populations representing hunter/gatherers, mixed subsistence practices, and agriculturalists, the Island Field burials have higher frequencies of caries than known agriculturalists. The inference is that the Island Field folk are intensively using wild plant foods to a degree mimicking the diet of farmers subsisting upon maize and other domesticates.
Plant remains recovered from on-site contexts can cautiously be employed in the reconstruction of local environments in conjunction with off-site sampling of macro-remains, pollen and phytoliths. Site and near-site environments may be anthropogenic in nature, intentionally or unintentionally created. This emphasizes the need for the collection and interpretation of paleoenvironmental data representing ever-broadening geospatial scales with sites representing central focal points. This approach will enhance our understanding of the environment in general and potentially make the impact of human activities on the landscape more visible (e.g., Abrams and Nowacki, 2008; Asch Sidell, 2008: 47–48; Fulton and Yansa, 2016; Messner, 2011: 127–128; Russell, 1983; Scully and Arnold, 1981; Stinchcomb et al., 2011; Trachtenberg et al., 2008: 136–137). People adapt to local environmental conditions; we have reconstructions of paleoenvironments at broad geographic scales; we need reconstructions for increasingly smaller geospatial contexts.
Research agendas that address landscape management and the manipulation of plant resources should be emphasized. This will contribute to a broader systemic understanding of subsistence, medicinal practices, a variety of technologies, settlement patterns, notions of territory, and social interactions (e.g., Stewart, 2015b: 14–15). The ethnohistoric/ethnographic record of the region and the Eastern Woodlands in general reveals a variety of practices whose antiquity could be explored (e.g., Cronon, 1983; Holmes, 1896; Keeley, 1995; Mounier, 2003: 151; Russell, 1979, 1980, 1981, 1983; Scarry, 2003; Smith, 2001; Wagner, 2003). These data must be used with caution (Lightfoot et al., 2013). Investigating landscape management must recognize the broad and patchy geospatial scales that might be involved, as well as the fact that altered environments or niches may be the cumulative result of incremental changes over extended periods (Lightfoot et al., 2013: 290; Wagner, 2003: 129–130). This suggests that efforts should focus on micro-geographic areas that include one or more settlements with a long history or re-occupation. It is critical that such research be interdisciplinary employing multiple lines of evidence.
The use of fire is one of the most frequently cited techniques used by native peoples to modify the environment, although opinions are divided about the frequency and extent to which settings were burned over (cf. Abrams and Nowacki, 2008; Buell et al., 1954; Patterson and Sassaman 1988; Russell, 1983). Fire can be used to open woods for better hunting, increase ease of travel, clear trails, stimulate or improve browse to attract game, encircle game, aid in spotting or tracking game, improving the yield of useful plants, clear away leaf litter for easier nut collecting, create a fire free zone around habitations, reduce vermin, influence the onset of spring growth, clear fields for planting, maintain old fields, stimulate woody shoots, drive away or escape from enemies, and create open woods around habitations to prevent ambush (Wagner, 2003: 133–134). A variety of methods have been employed in the investigation of the anthropogenic use of fire that could serve as models for future research in the Delaware Valley (e.g., Abrams and Nowacki, 2008; Fesenmyer and Christensen, 2010; Lightfoot et al., 2013: 292–294; Russell, 1983; Wagner, 2003: 134–142).
Recognizing vegetation patterns that are at odds with expectations stemming from an understanding of climate, landscape setting, influence of wild fires, and the habits and presumed natural ranges of particular species and their mutualist partners have been used to implicate or question the role of Native Americans in landscape modification and plant dispersal (e.g., Abrams and Nowacki, 2008; Asch Sidell, 2008: 47–48; Fulton and Yansa, 2016; Keener and Kuhns, 1997; MacDougall, 2003; Murphy, 2001; Russell, 1981; Trachtenberg et al., 2008: 136–137; Tulowiecki and Larsen, 2015; Warren, 2016). For example, given the use of nuts throughout prehistory in the Eastern Woodlands it is reasonable to ask if arboriculture or silviculture was practiced (Cowan, 1985: 218). This observation and related question certainly apply to the archaeological record of the Upper and broader Delaware Valley. On the basis of macro-regional studies in Eastern Woodlands (Abrams and Nowacki, 2008) the answer to the question would be, yes. Following Abrams and Nowacki (2008: 1133):
Current and future paleobotanical research in the Upper Delaware and wider region offers more than “food for thought.” Information from dated contexts is already substantial. Additional data exists from contexts that have been assigned a relative age on the basis of typological and stratigraphic associations that could be refined with radiocarbon assays of archived material.
Possible research projects would bring together archaeologists and a variety of environmental scientists addressing issues that have relevance for current concerns over climate change and environmental management.
Footnotes
Acknowledgements
Funding for the research reported here was provided by the New Jersey Historic Preservation Office of the state's Department of Environmental Protection. My heartfelt thanks go to Kate Marcopul, State Historic Preservation Officer and Administrator. She initially envisioned the overall project of which this article is a part and made it a reality. Thanks also to Vincent Maresca, Senior Historic Preservation Specialist at the New Jersey Historic Preservation Office, for providing information that I missed during my initial review of cultural resource management (CRM) reports. An extensive list of colleagues provided assistance in numerous ways to the overall project. An extensive list of colleagues provided assistance in numerous ways to the overall project. They include: Keith Bastianini, William Barse, Joe Blondino, Kurt Carr, Jay Custer, Richard Hunter, Gregory Lattanzi, Timothy Messner, Roger Moeller, Dave Orr, Alison Oskam, Mike Owens, Philip Perazio, Lori Rohrer, Nina Versaggi, Elizabeth Wagner, and John Wright. I am grateful to all.
Declaration of conflicting interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the New Jersey Office of Historic Preservation.
Author Biography
![]() .
.