
Abstract
The handling-induced dark neuron is a histological artifact observed in brain samples handled before fixation with aldehydes. To explore associations between dark neurons and immunohistochemical alterations in mouse brains, we examined protein products encoded by Cav3 (neuronal perikarya/neurites), Rbbp4 (neuronal nuclei), Gfap (astroglia), and Aif1 (microglia) genes in adjacent tissue sections. Here, dark neurons were incidental findings from our prior project, studying the effects of age and high-fat diet on metabolic homeostasis in male C57BL/6N mice. Available were brains from 4 study groups: middle-aged/control diet, middle-aged/high-fat diet, old/control diet, and old/high-fat diet. Young/control diet mice were used as baseline. The hemibrains were immersion-fixed with paraformaldehyde and paraffin-embedded. In the hippocampal formation, we found negative correlations between dark neuron hyperbasophilia and immunoreactivity for CAV3, RBBP4, and glial fibrillary acidic protein (GFAP) using quantitative image analysis. There was no significant difference in dark neuron hyperbasophilia or immunoreactivity for any protein examined among all groups. In contrast, in the hippocampal fimbria, old age seemed to be associated with higher immunoreactivity for GFAP and allograft inflammatory factor-1. Our findings suggest that loss of immunohistochemical reactivity for CAV3, RBBP4, and GFAP in the hippocampal formation is an artifact associated with the occurrence of dark neurons. The unawareness of dark neurons may lead to misinterpretation of immunohistochemical reactivity alterations.
Introduction
Artifacts associated with handling of animals can occur in any step of experimental neuroscience research, including behavioral testing, 1 neuroimaging, 2 and tissue collection and processing. 3 The handling-induced dark neuron is a common feature among a variety of histological artifacts observed in brain samples. 4 With hematoxylin and eosin (H&E) or cresyl violet staining, dark neurons are characterized by the monotonous morphological pattern of contracted hyperchromatic (hyperbasophilic on H&E staining) cell bodies and dendrites. The formation of dark neurons is associated with mechanical handling of fresh brain samples prior to fixation with aldehydes. 4 –6
There have been a number of published research articles in which handling-induced dark neurons were misinterpreted as evidence of neuronal degeneration. 4,5 Therefore, the recognition of dark neurons, as contrasted with degenerating neurons, is critical for the valid interpretation of neuropathologic changes that are identified on the basis of histopathological assessment. 4,5 Of note, it is plausible that the presence of dark neurons could give rise to alterations in immunohistochemical reactivity. Nonetheless, to our knowledge, there are no reports in the literature describing immunohistochemical reactivity changes in association with handling-induced dark neurons.
In the current study, to explore associations between the occurrence of handling-induced dark neurons and the alterations of immunohistochemical reactivity in the mouse hippocampal formation, we chose to examine 4 protein markers: caveolin-3 (CAV3, neuronal perikarya/neurites), retinoblastoma binding protein-4 chromatin remodeling factor (RBBP4, also known as RbAp48, neuronal nuclei), 7 glial fibrillary acidic protein (GFAP, astroglia), and allograft inflammatory factor-1 (AIF1, microglia). We used mouse brains obtained from our previous project originally aimed to study the effects of age and long-term high-fat diet exposure on metabolic homeostasis, in which the presence of dark neurons was an incidental finding. We hypothesized that dark neuron hyperbasophilia density would negatively correlate with the immunoreactivity density of protein markers, particularly neuronal markers.
Materials and Methods
Animals
Male C57BL/6N mice (Taconic Biosciences, Taconic Farms, Hudson, New York) were used. All the experiments were conducted in accordance with the guidelines of the American Association for the Accreditation of Laboratory Animal Care and National Research Council’s Guide for the Care and Use of Laboratory Animals and approved by the University of California San Diego Institutional Animal Care and Use Committee. Upon the completion of experiments (4-month exposure to either high-fat diet or control diet), mice were divided into 4 harvest batches for full necropsy and tissue collection within a period of 3 weeks with order counterbalanced for age/diet groups. Immediately after being removed from the skull, the fresh brains were bisected midsagittally. The left hemibrains were immersion-fixed with 4% paraformaldehyde/phosphate-buffered saline (PBS) at 4°C for 72 hours and then stored in 0.4% paraformaldehyde/PBS at 4°C until being processed for paraffin embedment.
For neuropathologic examination, hemibrains were available from 4 age/diet study groups of mice: middle-aged (14-15 months)/control diet (n = 11), middle-aged/high-fat diet (n = 6), old (24-25 months)/control diet (n = 9), and old/high-fat diet (n = 7). To acquire baseline data from mice of younger age, we used young (3 months)/control diet male C57BL/6N mice (n = 9). All fresh brains in the baseline group were harvested in 1 batch and the left hemibrains were immersion-fixed and processed for paraffin embedment with the same protocols as those used in the study groups.
For all the left hemibrains (n = 33 totally in the study groups and n = 9 in the baseline group), the paraffin-embedded tissue blocks were trimmed 500 µm off the medial surface. The first 5-μm-thick section taken was stained with H&E. The following 5-μm-thick sections were immunostained for GFAP, AIF1, RBBP4, and CAV3 in order. All the parasagittal hemibrain sections revealed the entire dorsal hippocampal formation and fimbria, consistent with the levels between 500 and 700 µm from the midline. 8 For each hemibrain, 1 tissue section was evaluated for each stain.
Chromogenic Immunohistochemistry
Primary antibodies used were directed against CAV3 (mouse monoclonal clone A-3 specific for an epitope mapping amino acid residues 3–40 at the N-terminus of mouse CAV3, sc-5310, 1:10 000 dilution; Santa Cruz Biotechnology, Santa Cruz, California), RBBP4 (mouse monoclonal clone 11G10, GTX70232, 1:40 000 dilution; GeneTex, Irvine, California), 7 GFAP (rabbit polyclonal, Z0334, 1:1000 dilution; Dako, Carpinteria, California), and AIF1 (rabbit polyclonal, 019-19741, 1:1000 dilution; Wako, Richmond, Virginia).
Tissue sections were deparaffinized with xylene and rehydrated through graded ethanol series and water. For RBBP4, GFAP, and AIF1, antigen retrieval was performed by autoclaving the tissue sections at 121°C for 20 minutes with 10 mM Tris/1 mM EDTA-2Na/0.05% Tween-20 buffer (pH 9) and then cooling down at room temperature for 20 minutes. The tissue sections were treated for 30 minutes with 0.3% hydrogen peroxide/PBS to quench endogenous peroxidase activity, rinsed in PBS (2 times, 5 minutes each), and incubated for 30 minutes with 2.5% normal horse serum (Vector Laboratories, Burlingame, California). Following 24-hour incubation with primary antibodies at 4°C, the tissue sections were rinsed in 0.1% Tween-20/PBS (2 times, 5 minutes each) and PBS (5 minutes) and then incubated for 40 minutes at room temperature with horse anti-mouse or -rabbit immunoglobulin G (IgG) secondary antibody (ImmPRESS HRP anti-IgG [peroxidase] polymer detection kits, MP-7402 and MP-7401; Vector Laboratories). Following washing with 0.1% Tween-20/PBS (2 times, 5 minutes each) and PBS (5 minutes), the signals were developed with 3,3′-diaminobenzidine (ImmPACT DAB peroxidase [HRP] substrate, SK-4105; Vector Laboratories) for 5 minutes at room temperature. After washing with water for 5 minutes, the tissue sections were dehydrated through graded ethanol series, cleared in xylene, and mounted with Cytoseal 60 (Richard-Allan Scientific, Waltham, Massachusetts). For the negative reagent control, the primary antibody was omitted, as previously described. 9
Quantifications of Dark Neuron Hyperbasophilia and Immunohistochemical Reactivity
By means of 2-dimensional computer-assisted image analysis, hyperbasophilia of dark neurons on H&E-stained tissue slides and immunoreactivity for CAV3, RBBP4, GFAP, and AIF1 on DAB tissue slides were quantified, as described previously. 9,10 In brief, the hemibrain sections were digitally scanned using a microscope slide scanner (Aperio ScanScope GL; Leica Biosystems, Buffalo Grove, Illinois) equipped with a 20× objective lens. Using Aperio ImageScope software (version 10.2.2.2319), a square of 3000 × 3000 μm2 covering the dorsal hippocampal formation and fimbria was extracted from each hemibrain.
Using Image-Pro Analyzer software (version 6.3; Media Cybernetics, Bethesda, Maryland), the hippocampal formation (excluding the subiculum) and fimbria separately were digitally drawn on each of the extracted images having the same size and resolution. Because dark neurons were distinctly hyperbasophilic in comparison with well-preserved neurons, hyperbasophilia of dark neurons was quantified within the hippocampal formation by setting histogram-based RGB color segmentation to select the specific hyperbasophilia. This color segmentation setting was consistently applied to all the hemibrains. The hyperbasophilia intensity per unit area (ie, hyperbasophilia density of dark neurons) was calculated, as described previously. 9
For each of the protein markers examined, histogram-based RGB color segmentation was set to select the specific immunoreactivity signal and consistently applied to all the hemibrains. The signal intensity per unit area (ie, signal density) was calculated, as described previously. 9 To adjust for the between-batch variation, 1 brain section from the same positive tissue control block was included in each immunostaining batch. The control signal density value (in a specified neuroanatomic area) was used to normalize all the signal density values of studied samples in the same batch, generating the immunoreactivity density values.
Statistical Analysis
The assumption of normality was tested using the Shapiro-Wilk test for the hyperbasophilia density of dark neurons (W = 0.750, P < .0001), the immunoreactivity density of CAV3, RBBP4, GFAP, and AIF1 (W = 0.924, 0.865, 0.940, and 0.977; P = .008, .0002, .029, and .536, respectively) in the hippocampal formation, and the immunoreactivity density of GFAP and AIF1 (W = 0.946 and 0.887, P = .045 and .0006, respectively) in the hippocampal fimbria. Because the normality assumption could not be confirmed for all variables except AIF1 in the hippocampal formation, nonparametric methods were used for all analyses. Spearman rho was used to test correlations between the hyperbasophilia density of dark neurons and the immunoreactivity density of each protein marker. The Kruskal-Wallis test was used to compare the hyperbasophilia density of dark neurons and the immunoreactivity density of each protein marker across all 5 age/diet mouse groups (4 study and 1 baseline groups). Post hoc comparison procedures were conducted using the Dunn multiple comparison test. Two-sided P values of less than .05 were considered statistically significant. The statistical analyses were performed using GraphPad Prism (version 6.0h; GraphPad Software, San Diego, California).
Results
Presence of Dark Neurons on H&E Histopathology
On H&E-stained histopathologic examination, we observed dark neurons of varying density as an incidental finding in a high proportion of mouse hemibrains in the study and baseline groups. In the hippocampal formation (Figure 1), dark neurons were more conspicuous in the stratum pyramidale than in the stratum granulosum. There was no consistent difference in the distribution of dark neurons across the hippocampal subfields. No definite changes in the number or morphology of astroglia or microglia were observed in spatial relation to the presence of dark neurons in the hippocampal formation of any hemibrains examined. In the cerebral cortex of all mice in the study and baseline groups, dark neurons were frequently observed at least in the superficial layers. The hippocampal fimbria white matter showed no significant histopathologic changes. Thalamic mineral deposits 11 were observed in one of the old/control diet mice.
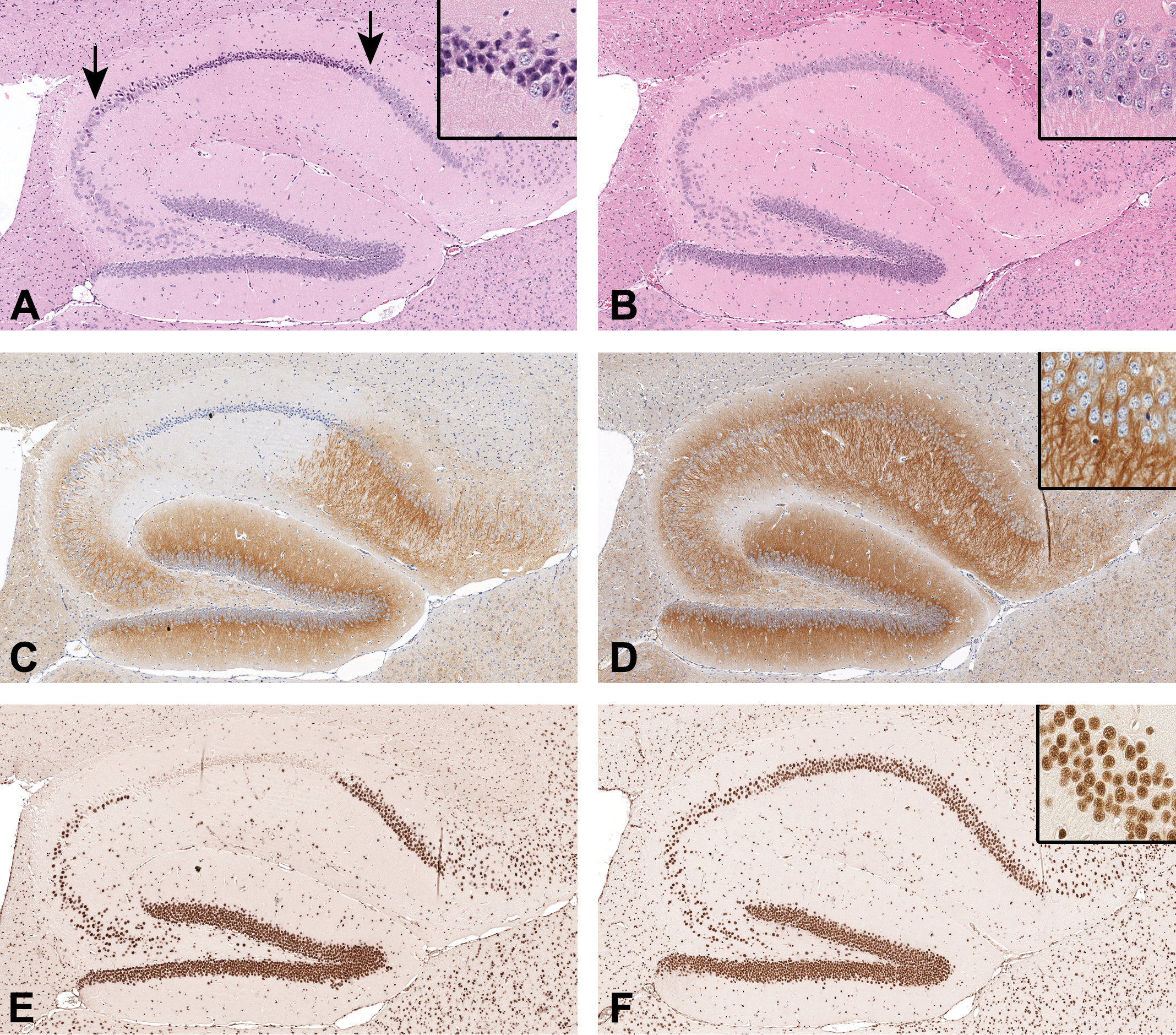
Representative images of the left dorsal hippocampal formation from 2 middle-aged (14-15 months) male C57BL/6N mice (in columns of adjacent tissue sections, original scan 20×). On hematoxylin and eosin staining, (A) dark neurons having contracted hyperbasophilic cell bodies and dendrites are shown in the stratum pyramidale (between arrows), in comparison with (B) well-preserved neurons. On diaminobenzidine immunohistochemistry for (C, D) caveolin-3 (CAV3, neuronal perikarya/neurites, with hematoxylin counterstaining), (E, F) retinoblastoma binding protein-4 chromatin remodeling factor (RBBP4, neuronal nuclei), (G, H) glial fibrillary acidic protein (GFAP, astroglia), and (I, J) allograft inflammatory factor-1 (AIF1, microglia), focal loss of immunoreactivity for CAV3, RBBP4, and GFAP is observed in a pattern that spatially matches the presence of dark neurons, whereas the variation of AIF1 immunoreactivity is not conspicuous. Insets = high magnification (original scan ×40).
(continued).
Focal Loss of Immunohistochemical Reactivity in the Hippocampal Formation
Based on qualitative assessments in the hippocampal formation, we found focal loss of immunoreactivity for CAV3, RBBP4, and GFAP in a pattern that spatially matched the presence of dark neurons observed in the H&E-stained adjacent hemibrain sections (Figure 1). The spatial variation of AIF1 immunoreactivity was not conspicuous in relation to the presence of dark neurons in the hippocampal formation (Figure 1).
Negative Correlations Between Dark Neuron Hyperbasophilia and Immunohistochemical Reactivity in the Hippocampal Formation
Based on quantitative image analyses in the hippocampal formation, we found negative correlations between the hyperbasophilia density of dark neurons and the immunoreactivity density of CAV3, RBBP4, and GFAP to be statistically significant (r = −0.444, −0.614, and −0.527; P = .003, P < .0001, and P = .0003, respectively, n = 42, Spearman rho; Figure 2A-C). For AIF1 immunoreactivity, the correlation was not statistically significant (r = −0.246, P = .116, n = 42, Spearman rho; Figure 2D).
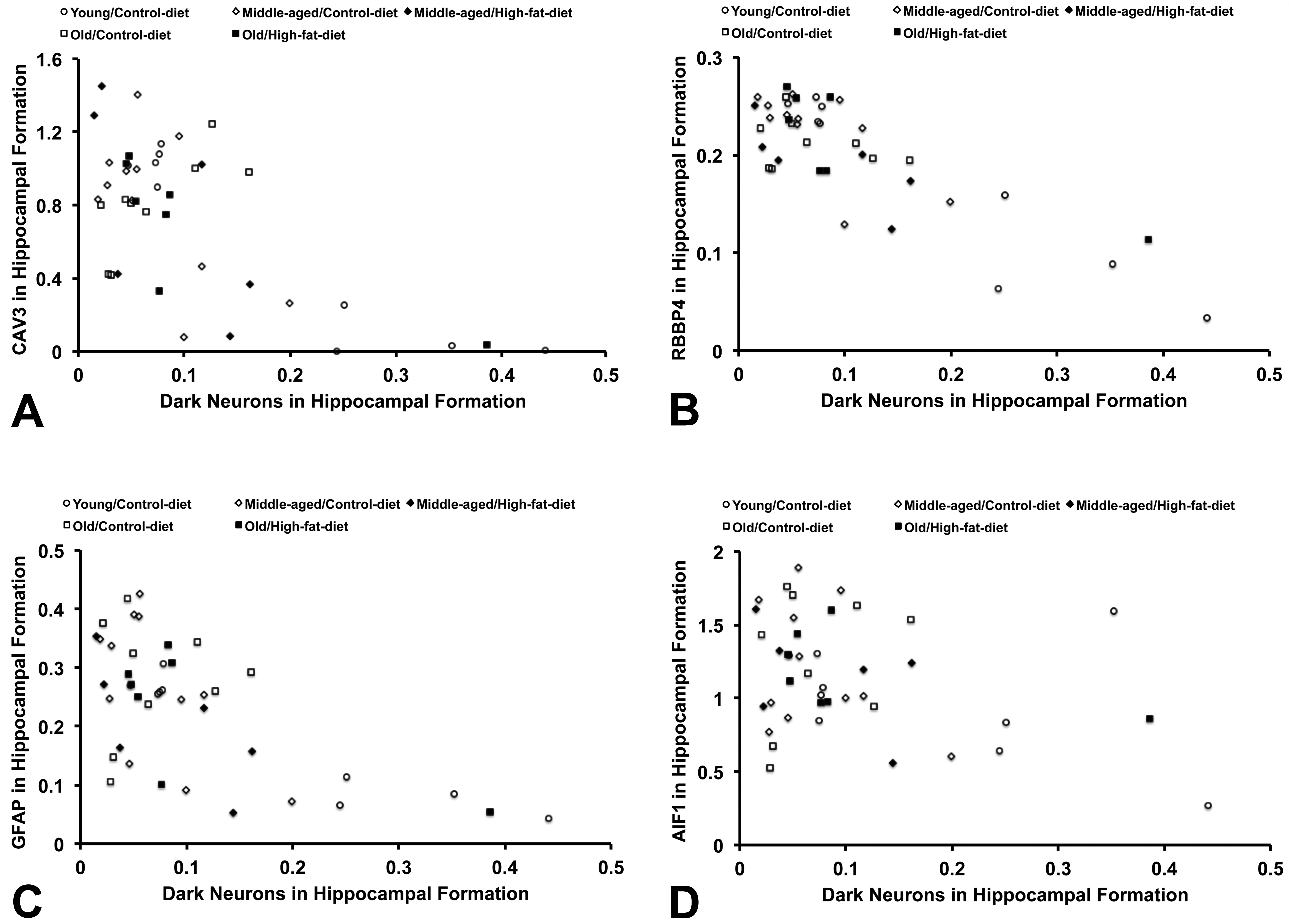
Scatter plots showing the relationship between the hyperbasophilia density of dark neurons and the immunoreactivity density of each protein marker in the left dorsal hippocampal formation. The negative correlation is observed for (A) caveolin-3 (CAV3; r = −0.444, P = .0032), (B) retinoblastoma binding protein-4 chromatin remodeling factor (RBBP4; r = −0.614, P < .0001), and (C) glial fibrillary acidic protein (GFAP; r = −0.527, P = .0003) but not statistically significant for (D) allograft inflammatory factor-1 (AIF1; r = −0.246, P = .116; n = 42, Spearman rho).
No Difference in Dark Neuron Hyperbasophilia or Immunohistochemical Reactivity in the Hippocampal Formation Across Age/Diet Mouse Groups
In the hippocampal formation, we found no significant difference in median values of the hyperbasophilia density of dark neurons or the immunoreactivity density of CAV3, RBBP4, GFAP, or AIF1 across all 5 age/diet mouse groups (n = 42, Kruskal-Wallis test; Table 1).
The Hyperbasophilia Density of Dark Neurons and the Immunoreactivity Density of Protein Markers in the Hippocampal Formation by Age/Diet Mouse Groups.a
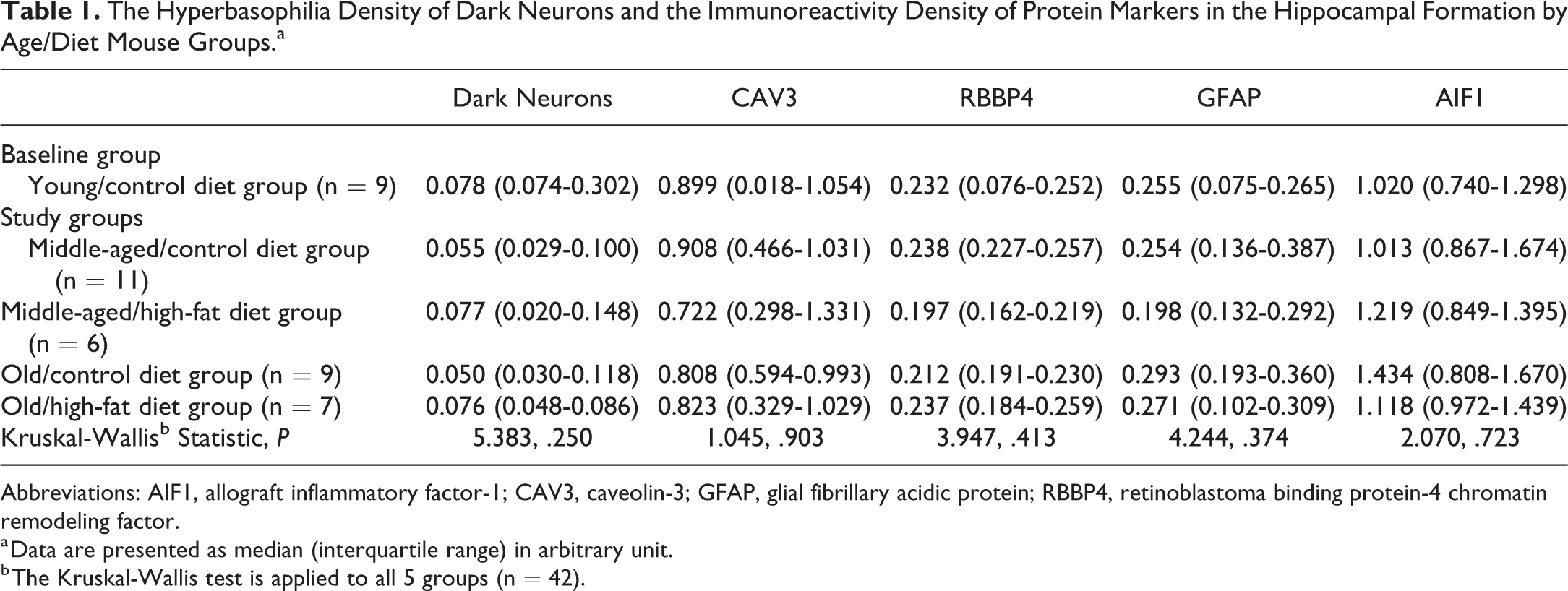
Abbreviations: AIF1, allograft inflammatory factor-1; CAV3, caveolin-3; GFAP, glial fibrillary acidic protein; RBBP4, retinoblastoma binding protein-4 chromatin remodeling factor.
a Data are presented as median (interquartile range) in arbitrary unit.
b The Kruskal-Wallis test is applied to all 5 groups (n = 42).
Differences in Immunohistochemical Reactivity for GFAP and AIF1 in the Hippocampal Fimbria Across Age/Diet Mouse Groups
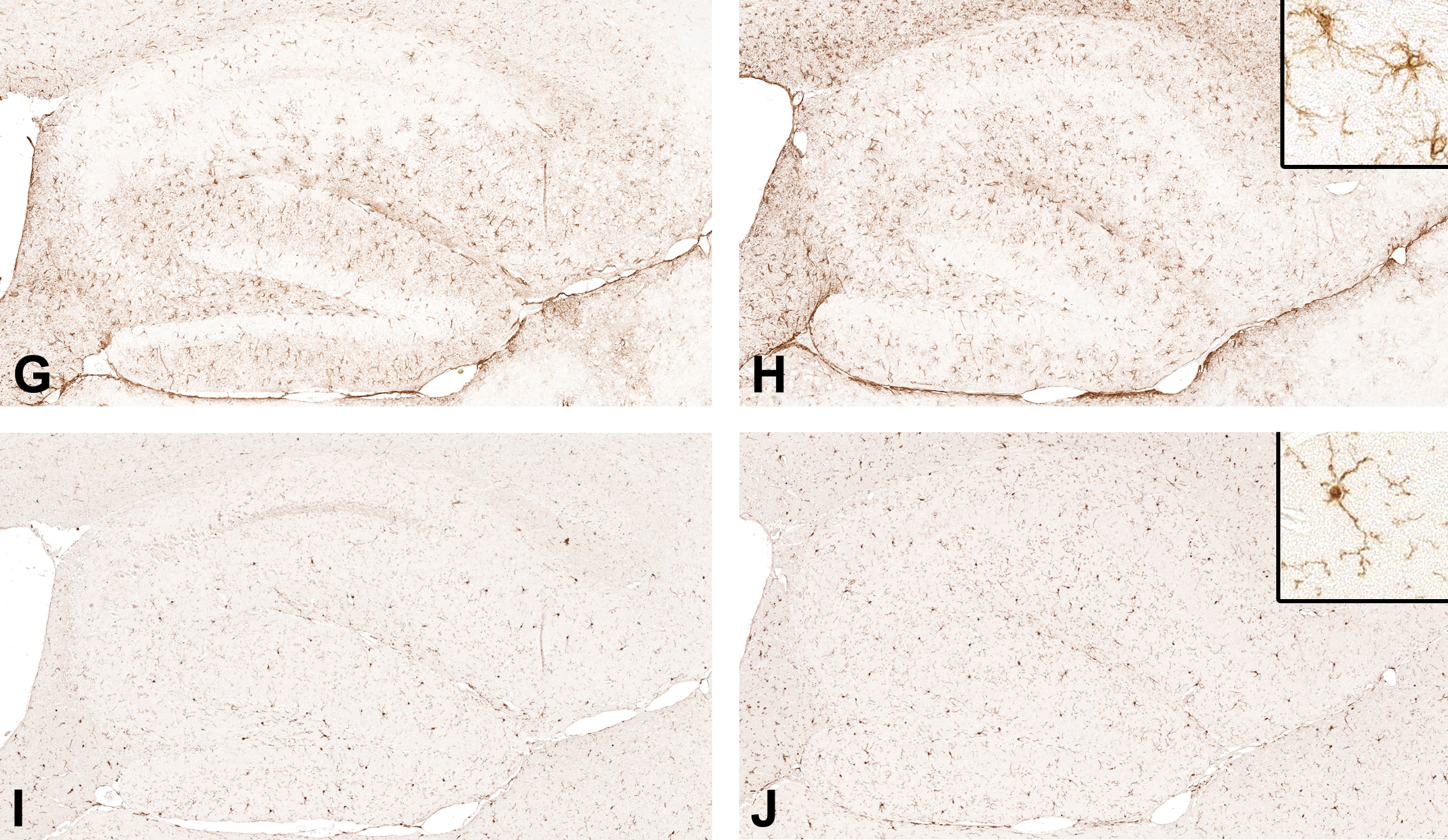
To compare the hippocampal formation affected by the occurrence of dark neurons with a neighboring white matter region in regard to the immunohistochemical reactivity, we examined GFAP and AIF1 immunoreactivity in the hippocampal fimbria (Figure 3).
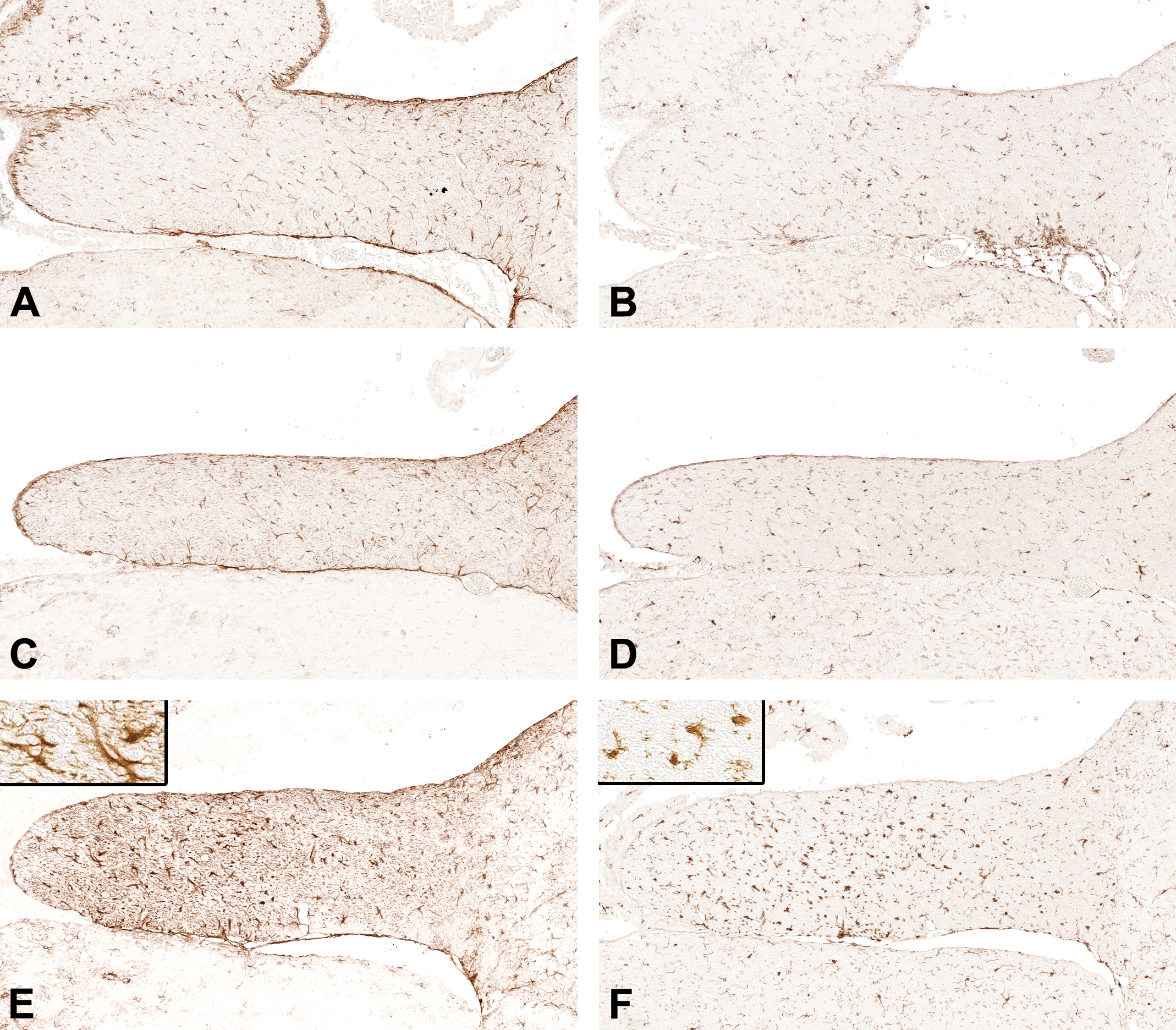
Representative images of the left hippocampal fimbria from young (3 months), middle-aged (14-15 months), and old (24-25 months) male C57BL/6N mice (in rows of adjacent tissue sections). The diaminobenzidine immunohistochemistry for (A, C, and E) glial fibrillary acidic protein (GFAP, astroglia) and (B, D, and F) allograft inflammatory factor-1 (AIF1, microglia) is depicted (original scan 20×). Insets = high magnification (original scan ×40).
Across all 5 age/diet mouse groups, we found differences in median values of the immunoreactivity density of GFAP and AIF1 (P = .020 and P < .0001, respectively, n = 42, Kruskal-Wallis test; Table 2). Compared with the baseline young/control diet group, the GFAP immunoreactivity density was higher in the old/high-fat diet group (P < .05) but not significantly higher in the other 3 study groups (Dunn multiple comparison test; Figure 4A). Compared with the baseline young/control diet group, the AIF1 immunoreactivity density was higher in the old/control diet (P < .05) and old/high-fat diet (P < .01) groups but not significantly higher in the other 2 study groups (Dunn multiple comparison test; Figure 4B).
The Immunoreactivity Density of GFAP and AIF1 in the Hippocampal Fimbria White Matter by Age/Diet Mouse Groups.a

Abbreviations: AIF1, allograft inflammatory factor-1; GFAP, glial fibrillary acidic protein.
a Data are presented as median (interquartile range) in arbitrary unit.
b The Kruskal-Wallis test is applied to all 5 groups (n = 42).
cP < .05.
dP < .0001.
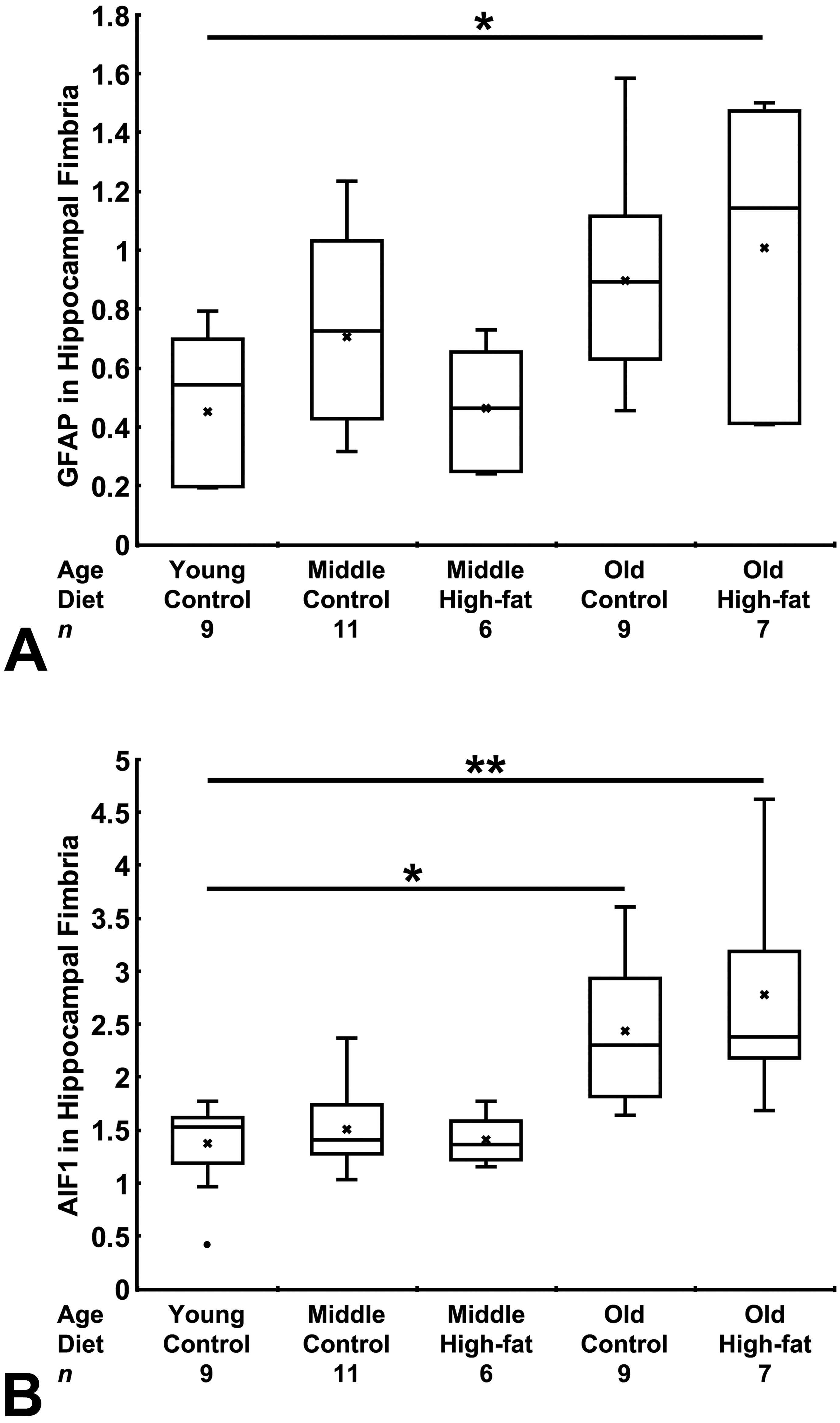
Box plots showing differences in the immunoreactivity density of glial fibrillary acidic protein (GFAP) and allograft inflammatory factor-1 (AIF1) in the left hippocampal fimbria among 5 age/diet mouse groups. Data in each group are presented in arbitrary unit as a box and whisker plot; X = mean, • = outlier. Compared with the young/control diet group, the immunoreactivity density of GFAP (A) was higher in the old/high-fat diet group and that of AIF1 (B) was higher in the old/control diet and old/high-fat diet groups (Dunn multiple comparison test). *P < .05, **P < .01.
Discussion
As an incidental finding in our previous project studying the effects of age and long-term high-fat diet exposure on metabolic homeostasis in male C57BL/6N mice, we observed dark neurons of varying density in the dorsal hippocampal formation in a high proportion of mouse hemibrains from both study and baseline groups. Based on quantitative image analysis, the hyperbasophilia density of dark neurons (ie, dark neuron burden) did not significantly differ among 5 age/diet mouse groups. Accordingly, we interpreted that dark neurons in the present study occurred in association with the mechanical handling of fresh mouse brains prior to immersion fixation with paraformaldehyde. 4 –6
Of note, we found focal loss of immunohistochemical reactivity for CAV3, RBBP4, and GFAP in spatial association with the presence of dark neurons in the hippocampal formation. These qualitatively observed associations were confirmed by our findings of negative correlations between the hyperbasophilia density of dark neurons and the immunoreactivity density of CAV3, RBBP4, and GFAP in the quantitative image analysis. Furthermore, for each of protein markers examined, we found no significant difference in the immunoreactivity density in the hippocampal formation among 5 age/diet mouse groups. On the other hand, in the hippocampal fimbria (a white matter continuation of the hippocampal formation), there were differences in the immunoreactivity density of GFAP and AIF1 among 5 age/diet mouse groups. That is, old age seemed to be associated with the higher immunoreactivity density, in agreement with previous studies showing aging-related increases in GFAP and AIF1 expression in C57BL/6 mice. 12,13 One potential explanation for these observations is the difference in texture of these 2 neuroanatomic structures with regard to the handling-induced histological artifact, where the fimbria white matter is less vulnerable to mechanical pressure applied to the brain surface. Collectively, loss of immunohistochemical reactivity for CAV3, RBBP4, and GFAP in the hippocampal formation was likely a histological artifact associated with the formation of handling-induced dark neurons.
Accumulating evidence suggests that handling-induced dark neurons are formed as a result of pressure applied to living fresh brains that are subsequently fixed with aldehydes. 4 –6 In an experiment with rat cerebral cortex biopsy, the formation of dark neurons could be prevented by pharmacologic blockage of glutamate receptors. 14 In addition, pathologically induced dark neurons (ie, either reversibly or irreversibly injured neurons), morphologically indistinguishable from handling-induced dark neurons, were well documented in rodent brains that were adequately perfusion-fixed with aldehydes following the experimental induction of certain acute insults, 5,6 such as status epilepticus, 15 hypoglycemia, and ischemic reperfusion injury. 16 Taken together, it is possible that the formation of dark neurons of either handling or pathological origin represents an early-stage neuronal response to complete energy deprivation and glutamate release, the perturbation that leads to the cellular contraction at the time of fixation with aldehydes. 6
With regard to loss of immunohistochemical reactivity in association with handling-induced dark neurons, it remains to be explored in future studies as to how conformational changes in protein molecules inherent to dark neurons 16 could interfere with the antigen–antibody interaction on immunohistochemistry. Note that our present study of immunohistochemical reactivity alterations in association with handling-induced dark neurons was based solely on 4 antibodies. This histological artifact may or may not occur with other antibodies not tested in our study. Moreover, our present finding may not be applied to the context of dark neurons of pathological origin. 17
In the current study, in addition to neuronal markers examined (CAV3 and RBBP4), the association between the presence of dark neurons and loss of immunohistochemical reactivity for an astroglial marker (GFAP) was unexpectedly observed. Our findings suggest that the handling-induced histological artifact resulting from pressure applied to fresh brains before being fixed with aldehydes can interfere with the antigen–antibody interaction on immunohistochemistry in astroglia as well.
In conclusion, our findings suggest that loss of immunohistochemical reactivity can occur as an artifact associated with the formation of handling-induced dark neurons. In experimental neuroscience research, if both histopathologic assessment and molecular analysis are necessary, brain samples are to be mechanically handled prior to fixation. In this scenario, the brain should be handled gently and carefully to avoid the formation of dark neurons. 3 The unawareness of handling-induced dark neurons, which are identifiable on bright-field H&E microscopy, may lead to misinterpretation of immunohistochemical reactivity alterations. 5 In particular, this misinterpretation may occur in the assessment of protein expression on dark-field immunofluorescence microscopy of brain samples that are not optimally handled during harvest and subsequently fixed with aldehydes and in which histopathologic changes are not concurrently evaluated on bright-field microscopy.
Footnotes
Authors’ Note
The data are available upon request. Please direct all requests to the corresponding author at
Author Contributions
Authors contributed to conception, design, and supervision of the study (D.V.J., H.H.S., and V.S.); animal experiments (J.M.S.); neuropathologic examination, immunohistochemistry, microscope slide scanning, quantitative image analysis, or statistical analysis (V.S., R.C.C., and B.S.); the first manuscript draft (V.S., R.C.C., B.S., and J.M.S.); and the final manuscript (V.S., H.H.S., and D.V.J.). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was support by Sam and Rose Stein Institute for Research on Aging at University of California San Diego and by the US National Institutes of Health (NIH) under Award Number R21MH115825. The funding sources had no involvement in study design; in the collection, analysis, and interpretation of data; in the writing of the report; and in the decision to submit the article for publication.