
Abstract
The metabolic syndrome (MetS), a cluster of dyslipidemia, hypertension, and diabetes and an important contributor to cardiovascular morbidity and mortality, occurs in nearly 35% of adults and 50% of the aging population in the United States. However, the underlying mechanisms by which MetS orchestrates and amplifies cardiovascular events remain elusive. Furthermore, traditional therapeutic strategies addressing lifestyle modifications and individual components of MetS are often unsuccessful in decreasing morbidity due to MetS. The availability of an adequate experimental platform that mimics the complexity of MetS may allow development of novel management techniques. Swine models, including domestic pigs and minipigs, have made important contributions to our understanding of many aspects of MetS. Given their similarity to human anatomy and physiology, those models may have significant predictive power for elucidating the pathophysiology of MetS in a manner applicable to humans. Moreover, experimental maneuvers and drugs can be tested in these preclinical models before application in patients with MetS. This review highlights the utility of the pig as an animal model for metabolic disorders, which may play a crucial role in novel drug development to optimize management of MetS.
Overview
The prevalence of obesity and metabolic syndrome (MetS) is increasing in the developed world (Eckel, Grundy, and Zimmet 2005). MetS is defined by the constellation of obesity, dyslipidemia, hypertension, insulin resistance (IR), and increased levels of serum proinflammatory markers (Grundy et al. 2004). According to the American Heart Association, individuals are diagnosed with MetS when they show ≥3 of following components: (1) central or abdominal obesity (measured by waist circumference), (2) elevated triglyceride levels, (3) low high-density lipoproteins (HDL), (4) hypertension, and (5) elevated fasting glucose. The International Diabetes Federation criteria are similar but more restrictive for central obesity (Parikh and Mohan 2012). MetS is an important contributor to cardiovascular morbidity and mortality, which can be observed in nearly 35% of adults in the United States and 50% of the aging population (Aguilar et al. 2015). MetS patients are also at increased risk of developing microalbuminuria (Hoehner et al. 2002) and renal dysfunction (Chen et al. 2004).
Investigation into the pathogenesis of MetS and its adverse outcomes has been facilitated by the development of animal models of MetS. Various species have been used as models of MetS, particularly rats and mice, whereas studies in swine models to date account for no more than 10% of the total number of publications in this area as identified in a search of the PubMed database (http://www.ncbi.nlm.nih.gov/pubmed; http://www.ncbi.nlm.nih.gov/pubmed?term=(%22Metabolic+Syndrome+X%22%5BMesh%5D)+AND+%22Swine%22%5BMesh%5D; year 1960–2015). Although rodents are small and thus useful for multivariable experiments, they show several important differences in metabolism and physiology from humans (Davis et al. 2013; Arner 2005). For example, normal mouse lipoprotein profiles have primarily atheroprotective HDL, whereas normal human lipoprotein profiles contain primarily atherogenic low-density lipoprotein (LDL; Kennedy et al. 2010). Adipsin, an adipokine that is increased in human MetS, is lower in rodents with MetS (Rosen et al. 1989; Napolitano et al. 1994). Unlike swine (Table 1), rodents may not always develop hypertension (Mark et al. 1999). As rodents do not readily exhibit all the clinical signs of MetS observed in humans (Spurlock and Gabler 2008), translation of rodent data has been hindered. In contrast, pigs possess many anatomical and physiological similarities to humans, as well as a high sequence and chromosome structure homology (Vamathevan et al. 2013; Groenen et al. 2012). Comparisons of 317 known human drug target genes revealed less variation from minipig (19 genes) than from beagle (41 genes; Vamathevan et al. 2013). However, pigs do show some anatomical differences from humans. In the kidney, the avascular plane is transverse in swine, rather than longitudinal as in humans, and it has 2 large venous trunks (humans have 3) and no large veins on the dorsal surface of the pelvis (Bagetti Filho et al. 2008). In the heart, innervation of the atrioventricular node and ventricular conduction tissues also differs from humans (Crick et al. 1999). Nonetheless, the many similarities to humans support the use of the pig model for investigating MetS (Spurlock and Gabler 2008).
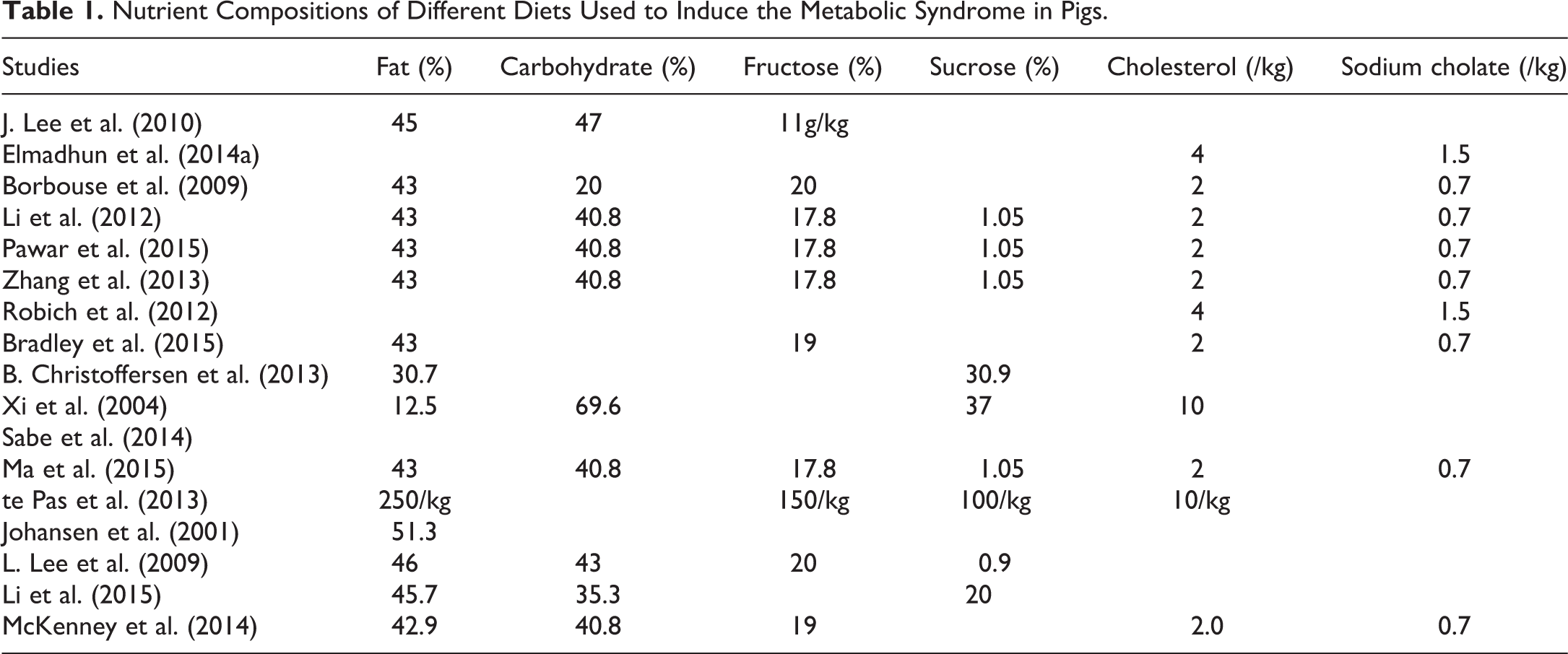
Nutrient Compositions of Different Diets Used to Induce the Metabolic Syndrome in Pigs.
In recent years, the impact of MetS on the heart and kidney has been characterized in swine models, and this will be our focus in this article.
Inducing MetS in Swine
MetS has been induced in different strains of pigs by high-caloric diets (Table 1), mostly including 15 to 25% (by weight) fatty acids (mainly lard supplemented with hydrogenated soya bean and coconut oil), 1 to 2% cholesterol, 40% refined sugars (commonly 20% fructose and 20% sucrose), 17% protein, and 15% other carbohydrates like starches and fibers. In most MetS models, features of this disorder appear after 3 to 6 months on the diet, but in some can emerge as early as 2 to 5 weeks on a high-fat/high-energy diet (B. Christoffersen et al. 2013; Johansen et al. 2001). At 12 weeks after high-fat/high-sucrose diet, there are usually apparent manifestations of MetS (Xi et al. 2004), which progressively increase over time (Pawar et al. 2015). After 6 months on a high-fat/high-fructose diet, some atherosclerotic plaque formation in the coronary artery may be evident (McKenney et al. 2014; Borbouse et al. 2009; Table 2).
Pig Characteristics and Manifested Metabolic Syndrome (MetS) Components Induced by MetS Diets.
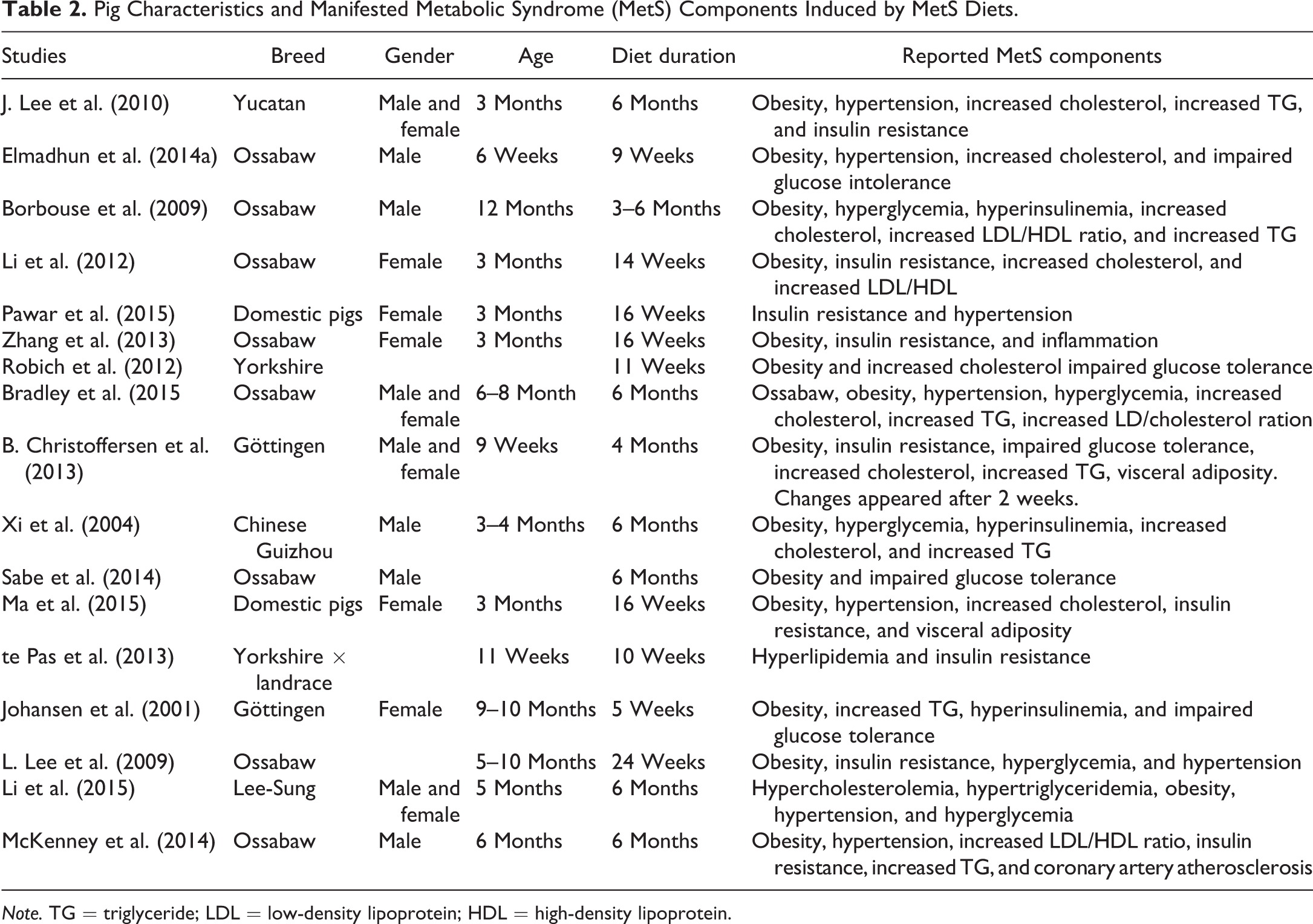
Note. TG = triglyceride; LDL = low-density lipoprotein; HDL = high-density lipoprotein.
Minipig breeds frequently used to simulate MetS include Yucatan (J. Lee et al. 2010), developed in the Yorkshire area of northern England and imported into the United States in 1890s; Sinclair, originally developed at the Hormel Institute of the University of Minnesota in the 1950s; Göttingen (Johansen et al. 2001), developed in the 1960s at the Institute of Animal Breeding and Genetics of the University of Göttingen, Germany, by crossbreeding the Vietnamese, Hormel, and German improved Landrace swine; and the Chinese Guizhou (Xi et al. 2004) and Taiwan Lee-Sung (S. J. Li et al. 2015) strains, as well as Ossabaw pigs.
The Ossabaw strain encompasses descendants of minipigs brought from Spain and subsequently isolated on the Ossabaw Islands off the coast of Georgia (Brisbin et al. 1977). They develop MetS upon high-fat/high-sucrose feeding (Clark et al. 2011; L. Lee et al. 2009; Zhang et al. 2013) and readily exhibit obesity, IR, and hypertension, not often seen in other breeds (Litten-Brown, Corson, and Clarke 2010). This unique strain has lived in relative genetic isolation for centuries, surviving on abundant food in the fall and famine conditions in the winter, a scenario that may have selected for a “thrifty genotype” (Speakman 2008). Hypothetically, in early evolutionary history, genes promoting efficient fat deposition might have been advantageous by allowing survival during periods of famine. In the modern society, such genes are disadvantageous because famine is not necessarily experienced, resulting in facilitated evolution of metabolic disorders. Ossabaw pigs have low insulin binding affinity for liver microenzymes and thereby are relatively insensitive to insulin (Meserole and Etherton 1984) and susceptible to MetS (Elmadhun et al. 2014a; Zhang et al. 2013). This strain, however, is costly and does not grow to the full body size of domestic pigs, possibly due to lower plasma levels of growth hormone (Kasser et al. 1981; Wangsness, Martin, and Gahagan 1977), resulting in more limited sample collection capacity.
Recently, MetS was successfully induced in large domestic pigs such as American Yorkshire (Sus scrofa domesticus) using high-saturated fat/cholesterol/sugar (te Pas et al. 2013) or high-fat/high-fructose diets (Pawar et al. 2015; Ma et al. 2015). In domestic pigs, a 2% high-cholesterol diet causes only endothelial dysfunction and early changes of atherosclerosis but not obesity, IR, or hypertension (Eirin et al. 2014; Urbieta-Caceres et al. 2010). Contrarily, a 3- to 4-month ad libitum high-fat/high-fructose diet (Table 1) induced significant manifestations of MetS resembling those observed in Ossabaw pigs. After 12 to 16 weeks on this diet, a 20% increase in blood pressure, 2-fold increase in IR (Figure 1A), a 4-fold increase in serum cholesterol, and doubling of triglyceride levels were observed. In addition, plasma tumor necrosis factor (TNF)-α levels were also elevated 3-fold (Z. L. Li et al. 2012; Pawar et al. 2015; Zhang et al. 2013). These findings suggest that the MetS model in domestic pigs is effective and may offer some advantages in cost and genetic heterogeneity.
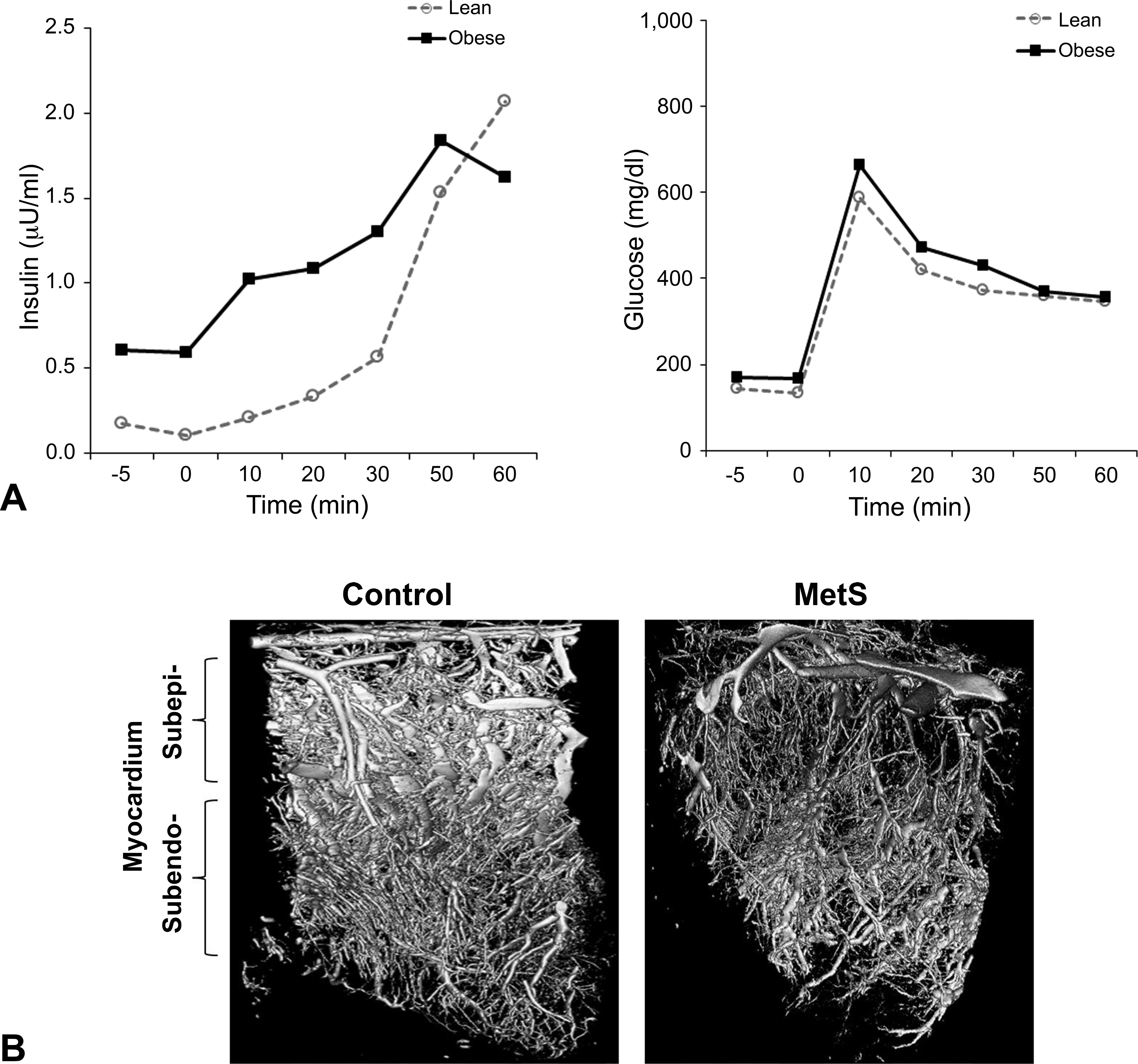
(A) Plasma glucose and insulin levels during an intravenous glucose tolerance test in Ossabaw pigs fed with MetS (Obese) or control (Lean) diets. (B) Metabolic syndrome in Ossabaw pigs show decreased microvascular density in myocardium compared with control using 3-dimensional microcomputed tomography.
Interestingly, female Göttingen minipigs seem to develop MetS models more readily than their counterpart intact male pigs. While male pigs also become obese and increase fasting blood glucose and insulin levels following a MetS diet (Larsen et al. 2001), female pigs develop a larger abdominal circumference and higher concentrations of plasma insulin, triglyceride, total cholesterol, and leptin. Compared with male pigs, females are more insulin-resistant and might therefore constitute better models for MetS (B. Christoffersen et al. 2013; B. O. Christoffersen et al. 2007).
In addition to diet-induced MetS, a genetically modified MetS model has been induced. This model leads to the increased activity of proprotein convertase subtilisin/kexin type (PCSK) 9, which results in lower liver LDL-receptor levels, which in turn reduces LDL uptake from the circulation, and thereby increases systemic LDL levels. In human subjects, a PCSK9 gain of function mutation (D374Y) can lead to increased cholesterol levels (Abifadel et al. 2003). Yucatan minipigs induced with liver-specific expression of human D374Y-PCSK9 displayed reduced hepatic LDL-receptor levels, severe hypercholesterolemia, and spontaneous development of progressive aortic atherosclerotic lesions with human-like histopathological features (Al-Mashhadi et al. 2013). This model may be useful for translational research in atherosclerosis, although it requires a relatively long induction period, often 12 months. Additional studies are also required to determine whether this model exhibits indices of MetS such as IR and hypertension.
MetS pig models develop distinct injuries in many vital organs and tissues. Swine MetS has provided insight regarding obesity-induced alterations in the central nervous system (Karmi et al. 2010), digestive system (Liang et al. 2015), pancreas (Fullenkamp et al. 2011), gut microbiome (Pedersen et al. 2013), heart (Neeb et al. 2010), kidney (Z. Li et al. 2011), reproductive (Newell-Fugate et al. 2014), and musculoskeletal (Karmi et al. 2010) systems, as well as metabolic health in the offspring (Arentson-Lantz et al. 2014). On the other hand, the higher cost of large compared to small animals may limit the experimental sample size, and assessment of central obesity in swine is limited by lack of defined standards for waist circumference or body mass index. Nevertheless, this model holds promise for exploring clinically relevant pathways of metabolic disorders.
MetS and the Swine Heart
A recent study showed that normal weight patients with MetS had a 1.6-fold and obese patients with MetS had a 2.5-fold increased risk for death from cardiovascular events as compared with normal weight patients without MetS (Arnlov et al. 2010). Pigs are useful models for the study of cardiovascular disease, owing to a 90% similarity of morphology and physical function of the cardiovascular system to human (Smith and Swindle 2006; Hughes 1986).
We have shown in MetS Ossabaw pigs that a 16-week high-fat/high-fructose diet induced a 30% increase in the rate–pressure product, impaired myocardial perfusion, and blunted response to vasoactive challenge by adenosine (Z. L. Li et al. 2012). Myocardial oxygenation was greatly reduced in both Ossabaw and domestic MetS models on this diet (Pawar et al. 2015; Z. L. Li et al. 2012). MetS diet also accelerated formation of atherosclerotic plaque and aggravated in-stent stenosis in both Yucatan and Ossabaw pigs but to a greater extent in the latter breed (Neeb et al. 2010). Ossabaw MetS pigs also showed decreased microvascular density in the subepicardial myocardium, as shown by microcomputed tomography (Z. L. Li et al. 2014; Figure 1B), indicating adverse effects of MetS on both the cardiac macro- and microcirculation. Altered myocardial vascular function in Ossabaw pigs fed an atherogenic diet has been linked to decreased myocardial endothelial nitric oxide (NO) synthase functionality and NO bioavailability (Bradley et al. 2015), and in Yucatan pigs to impaired function of large conductance Ca2+-activated K+ (BKCa) channels (Mokelke et al. 2005). MetS in Ossabaw pigs induced by a high-cholesterol diet also impairs signaling of cardiac angiogenesis (Elmadhun et al. 2014a), thereby restricting the compensatory capacity of the myocardium for recruiting adequate blood and oxygen supplies.
In Ossabaw MetS models, myocardial oxidative stress was found to be increased and antioxidant enzymes reduced (Bradley et al. 2015). Moreover, MetS attenuated myocardial autophagy in response to overwhelming nutrient supplies which were observed at both early (3 months; Z. L. Li et al. 2012) and late (9 months; Sabe et al. 2014) stages of MetS. As autophagy is a housekeeping process to maintain cellular energy homeostasis, its attenuation may have profound influence on myocardial viability and cardiac adaptability in MetS. Additionally, in Ossabaw pigs with superimposed renovascular hypertension, we found that MetS magnified downregulation of mitochondrial proteins and activity (Zhang et al. 2015) and synergistically exacerbated the diastolic dysfunction and myocardial fibrosis induced by hypertension (Zhang et al. 2015). Importantly, in Yucatan pigs, chronic (7 months) MetS may lead to cardiac IR, along with impaired insulin signaling such as PI3-kinase activation, Akt phosphorylation, and abnormal phosphorylation of insulin receptor substrate 1 (J. Lee et al. 2010). These effects may further inhibit proper cardiac energy utilization and function. These findings suggest multiple potential pathways that mediate the myocardial injury in MetS, but their roles in directing pharmacological exploration need to be carefully examined.
MetS and the Swine Kidney
Patients with 1 to 2 and those with ≥3 traits of MetS are, respectively, 80% and 130% more likely to have microalbuminuria than those without the syndrome (Hoehner et al. 2002). The incidence of chronic kidney disease in patients with MetS increases progressively with the individual number of the MetS components (Chen et al. 2004). These studies suggest a close link between MetS and renal dysfunction, but their mechanistic relationship is incompletely understood, partly because of the need for appropriate experimental platforms. One of the advantages of pigs as experimental models for kidney research, as compared with rats and dogs, is that their kidneys are more similar in anatomy and physiology to the human kidney (Yokota, Benyajati, and Dantzler 1985; Schook and Tumbleson 1996). The porcine kidney is multipyramidal with an undivided cortex and has several different medullary structures. Each medullary pyramid forms a separate papilla, and fusion results in the formation of some compound papillae. Pigs also possess maximal urine concentration, glomerular filtration rate (GFR), and renal blood flow (RBF) similar to humans (Sachs 1994). These important properties render swine a suitable model to study renal disease, including effects of MetS on the kidney.
Both MetS Ossabaw and domestic pig models almost double their RBF and GFR upon high-fat/high-fructose diet feeding (Z. Li et al. 2011; Ma et al. 2015). In Ossabaw pigs, the augmented renal hemodynamic indices are associated with increased renal cortical volume and microvascular density (Z. L. Li et al. 2012), linking augmented hemodynamics and microcirculatory remodeling in the development of kidney hypertrophy (Z. Li et al. 2011). Interestingly, neither circulating nor renal inflammatory and oxidative stress markers in the MetS Ossabaw pigs were different from the control group at 10 weeks (Z. Li et al. 2011), but increased renal deposit of triglycerides was positively associated with increased GFR (Z. Li et al. 2011), suggesting a possible role for excessive lipid deposition in kidney injury in early MetS.
Nevertheless, the enhanced renal hemodynamic state does not seem to be renoprotective. Instead, the proximal and Henle’s tubular flow in the MetS kidneys was found to be slower than in healthy pig kidneys and associated with marked proximal tubular vacuolization, suggesting tubular degenerative changes (Z. Li et al. 2011). Furthermore, tubulointerstitial fibrosis becomes evident in Ossabaw pigs after 16 weeks of the same diet, although RBF and GFR remain elevated (Zhang et al. 2013). By 16 weeks, MetS Ossabaw pigs exhibited increased renal macrophages and plasma oxidized-LDL levels (Zhang et al. 2013), indicating that inflammation may play a role at later stages of renal injury. Both MetS Ossabaw and domestic pig kidneys are also encased in greater amounts of perirenal fat, compared with control kidneys, enriched in inflammatory (M1) macrophages and cytokines, including TNF-α (Zhang et al. 2013; Ma et al. 2015). At least in the domestic pig MetS model, the perirenal fat may not only serve as an inflammatory depot but may also impair renal artery endothelial function (Ma et al. 2015). These findings may have implications in therapeutic management of MetS-induced kidney disease.
MetS and the Swine Adipose Tissue
Visceral obesity is a major component of MetS. High-energy diet induced MetS impairments in glucose and insulin metabolism are associated with an increase in accumulation of body fat (B. Christoffersen et al. 2013). We have recently shown that in domestic pigs both abdominal fat tissue volume and adipocyte size progressively increased over 16 weeks of MetS diet, accompanied by a parallel increase in intraadipose capillary count and fibrosis (Pawar et al. 2015). The adipokines, adiponectin and leptin, are reciprocally regulated by obesity (Spurlock et al. 1998; Jacobi et al. 2004; Pawar et al. 2015). Indeed, MetS upregulates the release of abundant inflammatory adipokines, such as TNF-α, interleukin 6, and monocyte chemotactic protein 1, from the adipose tissue (Pawar et al. 2015), and stimulates macrophage infiltration (Ma et al. 2015; Zhang et al. 2013). Furthermore, MetS upregulates the expression of toll-like receptors in porcine adipose tissue (Gabler et al. 2008), which is closely associated with development of IR (Shi et al. 2006). Importantly, in the domestic MetS model, the inflamed adipose tissue may affect the organ it encapsulates, at least partly by disrupting the vascular endothelial function, which can be restored by TNF-α inhibition (Ma et al. 2015). Similarly, in Ossabaw MetS, perivascular fat impairs coronary endothelial function (Payne et al. 2010) and is linked to atherosclerosis formation in the coronary artery wall (McKenney et al. 2014). Whether these changes are similar to human events is unclear, and further investigations would be useful to provide more insight in this area.
Experimental Treatment in the MetS Swine Model
Lifestyle changes are the essential and fundamental management strategies in MetS. In a swine model, pigs fed a Paleolithic diet consistent with the hunter–gatherer lifestyle of our ancestors are leaner, more sensitive to insulin, and have lower circulating concentrations of C-reactive protein than their counterparts fed a cereal diet reflective of modern-day habits (Jonsson et al. 2006), signifying the importance of dietary modification in alleviating the progression of MetS.
Additionally, medications have been shown to modulate cellular turnover and improve tissue viability. The antidiabetic drug metformin confers a survival advantage in patients with cardiovascular disease and was recently found to selectively downregulate the apoptosis pathway in MetS pigs and upregulate cardioprotective proteins including mitogen-activated kinase proteins p38 and extracellular signal-regulated protein kinases 1 and 2 (Elmadhun et al. 2014b). Atorvastatin elicits a net decrease in apoptosis as well (Sabe et al. 2015), prevents myocardial autophagic dysfunction produced by MetS, which may in part account for its cardioprotective effect (Sabe et al. 2014), and attenuates adiposity in pigs on a high-fat diet (Amuzie et al. 2016; in this issue). On the other hand, cholesterol levels in domestic pigs are less responsive to statins than humans (Hasler-Rapacz et al. 1996), possibly due to differences in lipid metabolism. Like rats and dogs, pigs have low plasma activity of cholesteryl ester transport protein and manifest a high HDL and low LDL distribution (Yin et al. 2012). Many traditional models, including rabbit, Zucker diabetic fatty rat, and mice, do not show overall similarity to dyslipidemic humans. Nonhuman primates exhibit the most similar lipid profile compared to humans with respect to basal plasma total cholesterol, LDL/HDL ratio, and lipoprotein traces, but ethical issues restrict their widespread use as experimental models. Furthermore, interspecies differences in cholesterol handling might also be related to the primary site for fatty acid synthesis, which is the liver in humans, the adipose tissue in pigs, and both the liver and adipose tissue in rodents and rabbits (Nafikov and Beitz 2007).
Treatment with sustained-release nitrite reduces myocardial oxidative stress while increasing myocardial antioxidant capacity and NO bioavailability and is associated with marked improvement in vasoreactivity of coronary arteries (Bradley et al. 2015). In addition, resveratrol polyphenol, which is often found in high concentrations in red wine and considered cardioprotective, improves regional ejection fraction and myocardial perfusion in the high-cholesterol diet-induced MetS heart, with or without chronic ischemia (Robich et al. 2012; Sabe et al. 2013), as well as body mass index (Sabe et al. 2014). Such beneficial effects are thought to be mediated through binding to its primary target protein sirtuin 1, a key regulator of energy metabolism (X. Li 2013), and activating NO synthase (Robich et al. 2012). In a Chinese Guizhou pig MetS model, a lipoprotein lipase activator (ibrolipim) decreased ectopic lipid deposition, improved IR, and alleviated the β cell damage (Yin et al. 2004).
Clearly, these studies have markedly advanced the understanding and identification of new pathways contributing to MetS and their interaction with targeted pharmacological agents. Nevertheless, drug metabolism in MetS pigs requires cautious assessment. More studies are therefore needed to gauge optimal dosages of anti-MetS drugs and to decrease the potential of MetS subjects experiencing life-threatening toxicity or lack of effects.
Conclusion
It is becoming increasingly apparent that both minipigs and larger pig strains are valuable models to mimic human MetS and test new treatment strategies. Additional advantages of the pig are the ability to use standard diagnostic and treatment technologies similar to humans and collect body fluid and tissues in adequate quantity, especially in larger swine, thereby accelerating clinical translation. Future studies need to continue exploring the molecular basis of MetS and its comorbidities, as well as pathogenesis of MetS in other vital organs such as liver (steatohepatitis). Studies that focus on validation of the MetS models by controlling the severity of symptoms of MetS and organ-specific complications through developing and optimizing novel therapeutics would be of great significance.
Footnotes
Authors’ Contribution
All authors (XZ and LL) contributed to conception or design; data acquisition, analysis, or interpretation; drafting the manuscript; and critically revising the manuscript. All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Partly supported by NIH Grants DK104273, DK102325, DK73608, and HL123160.