
Abstract
Cell clusters were observed in the seminiferous tubules of C57BL/6J mice as a spontaneous lesion in a 2-week toxicity study, and they were demonstrated to be basically composed of Sertoli cells by immunohistochemistry for claudin-11 and GATA-4 (GATA-binding protein 4), which are both Sertoli cell markers. The clusters were composed of about 5 to 50 cells, which had eosinophilic and occasionally vacuolated cytoplasm with an unclear cell boundary. The cell clusters involved some sperm. No mitotic figures were observed and no immunoreactivity for proliferating cell nuclear antigen (PCNA) was detected in the clusters. In most cases, the cell clusters were observed in seminiferous tubules that also showed degenerative changes. In rare instances, cell aggregates immunohistochemically positive for claudin-11 were observed in the lumen of the epididymis, suggesting that some of the Sertoli cell clusters were sloughed off from the seminiferous epithelium into the epididymal ducts. To our knowledge, this is the first report of Sertoli cell clusters in any animal species except for transgenic or surgically altered animals.
Keywords
Sertoli cells have a pivotal role in supporting spermatogenesis and their injury will rapidly result in germ cell degeneration (Creasy and Foster 2001). While various non-neoplastic, histological abnormalities such as multinucleated giant cells, apoptosis, and exfoliation are noted for germ cells, only a few abnormalities including vacuolation and necrosis are commonly described for Sertoli cells (Creasy et al. 2012). We investigated the presence of cell clusters, believed to be Sertoli cell clusters, observed as a spontaneous lesion in the seminiferous tubules of C57BL/6J mice, using both histological and immunohistochemical approaches.
The treatment and handling of the animals were approved by the Institutional Animal Care and Use Committee of the Toxicology Research Laboratories, Central Pharmaceutical Research Institute, Japan Tobacco Inc.
Six-week-old male C57BL/6J mice were purchased from Charles River Laboratories Japan, Inc. (Kanagawa, Japan) and used in a 2-week oral dose toxicity study, which consisted of 1 vehicle (0.5% methylcellulose aqueous solution [0.5%MC]) control group and 4 treatment groups with different dose levels of a compound (6 males per group). The animals were housed in stainless steel cages in an environmentally controlled room (temperature of 23°C ± 3°C, relative humidity of 52.5% ± 22.5%, and a 12-hr light–dark cycle) and were allowed free access to a commercial diet (CRF-1; Oriental Yeast, Chiba, Japan) and tap water. After the treatment period, the animals were anesthetized with ether, sacrificed by exsanguination via abdominal aortic transection, and subjected to complete necropsy at 8 weeks of age. No macroscopic findings were observed in the testis and no treatment-related findings were observed in any organ at necropsy. The testes and epididymides were collected from all animals, fixed in Davidson’s fixative, paraffin embedded, and then sectioned at 4 μm thickness and stained with hematoxylin and eosin (H&E) or Periodic acid–Schiff (PAS). One transverse section from each testis and one longitudinal section from each epididymis were prepared for staining. Immunohistochemistry using rabbit polyclonal anti-claudin-11 antibody (diluted 1:100; Novus, Littleton, CO), mouse monoclonal anti-GATA-4 (GATA-binding protein 4) antibody (diluted 1:100; Santa Cruz Biotechnology Inc., Santa Cruz, CA), and mouse monoclonal anti-proliferating cell nuclear antigen (PCNA diluted 1:100; Dako, Glostrup, Denmark) were also conducted on paraffin-embedded sections. Claudin-11 is expressed in Sertoli cell tight junctions and is a marker for Sertoli cells in the testis (Morita et al. 1999). GATA-4 is expressed in the nuclei of somatic cells and is a marker for Sertoli cells in the seminiferous tubules (Ketola et al. 1999; Viger et al. 1998). For claudin-11 and PCNA staining, sections were boiled for 15 min in 0.01 M sodium citrate buffer (pH 6.0) for antigen retrieval. Histofine Simple Stain MAX-PO (Nichirei, Japan) was employed for the detection of primary antibodies.
Microscopically, in H&E-stained sections, cell clusters composed of about 5–50 cells each were observed unilaterally or bilaterally in approximately 1–4 seminiferous tubules in the testes of 1–3 mice per group including the vehicle control group (Table 1 and Figure. 1A–D). In total, cell clusters were observed in 9 of 30 mice used in this study, unilaterally in 7 mice and bilaterally in 2 mice. The cell clusters were located in the lumen of the seminiferous tubules or within the seminiferous epithelium. The constituent cells had eosinophilic and occasionally vacuolated cytoplasm with an unclear cell boundary. The nuclei of the cells were oval or bent, containing a prominent nucleolus and condensed chromatin. No mitotic figures were observed in the cell clusters. Mature sperm were occasionally seen associated with the cell clusters (Figure 1C). In addition to the above findings, very slight to slight tubular degeneration was observed in the testis, and in most cases, the cell clusters were observed in seminiferous tubules undergoing degeneration (Table 1 and Figure 1A and B). Seven mice with cell clusters also had tubular degeneration while two mice clusters appeared in tubules with otherwise histologically normal seminiferous epithelium (Figure 1D). In some cases, clusters were also observed in the tubuli recti, these tubules being identifiable due to their flat or incomplete epithelium and their position is directly beneath the capsule and adjacent to the rete. No inflammatory response was observed in any of the affected seminiferous tubules. In the epididymis, cysts, sperm granulomas, and slight cell debris were observed although there was no correlation of the epididymal findings with the presence of cell clusters in the testes (Table 1). As the testicular findings were seen at a similar incidence and severity in both control and treated animals in this study, their presence was considered to be spontaneous (Table 1).
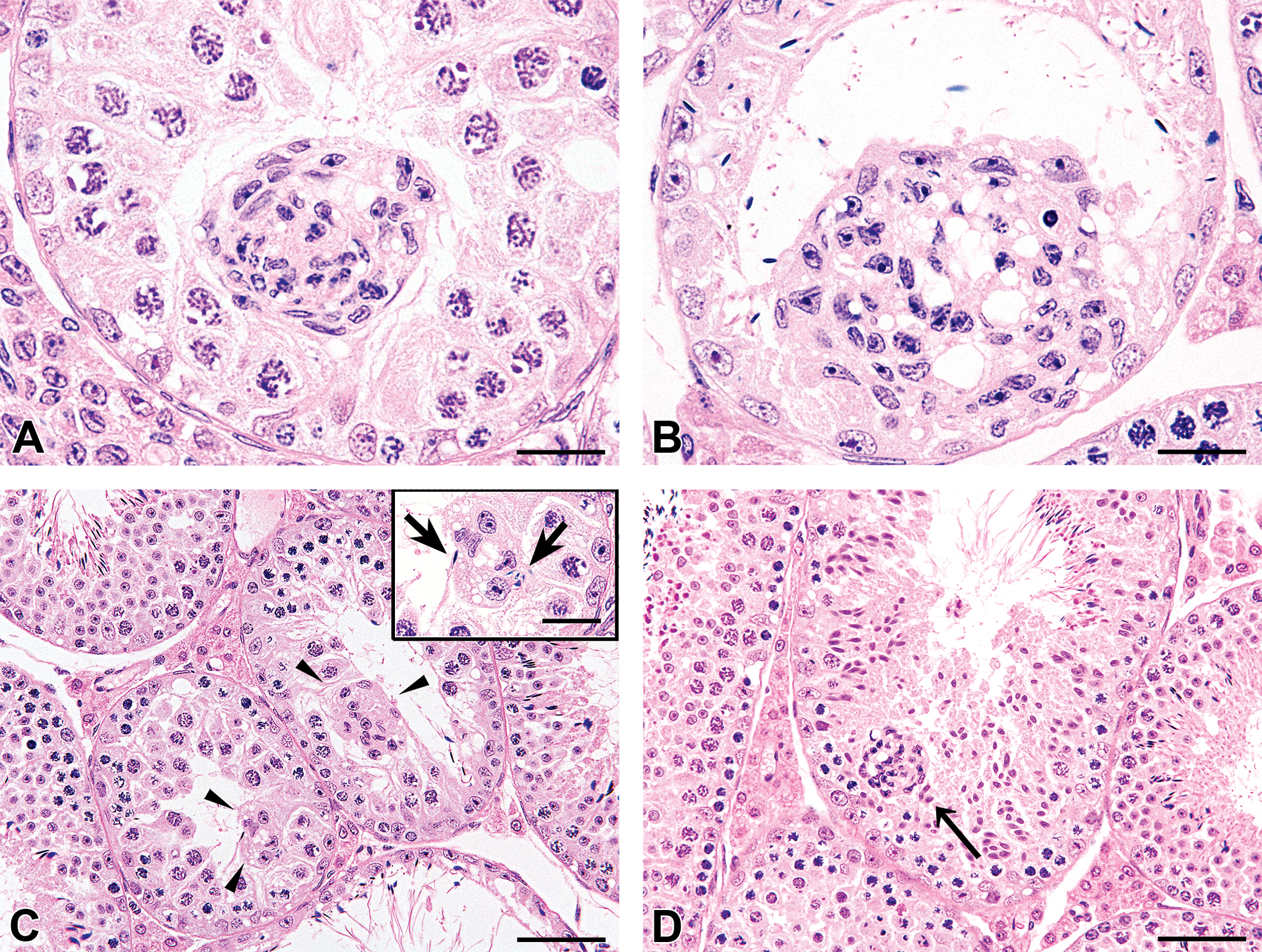
Histological appearance of the lesions, HE staining. A cell cluster observed in the lumen of the seminiferous tubule (bar: 20 μm) (A). A cell cluster present in degenerative seminiferous epithelium, projecting into the tubular lumen. Constituent cells had eosinophilic and occasionally vacuolated cytoplasm with an unclear cell boundary. The nuclei of the cells were oval or bent, containing a prominent nucleolus and condensed chromatin. No mitotic figures were observed in the cell clusters (bar: 20 μm) (B). Cell clusters (arrowheads) composed of several cells (bar: 50 μm). A few sperms were involved in the cluster (arrows; inset bar: 20 μm) (C). A cell cluster (arrow) in the seminiferous epithelium that was histologically normal, except for the presence of the cluster (bar: 50 μm) (D).
Microscopic findings in the testis and epididymis.
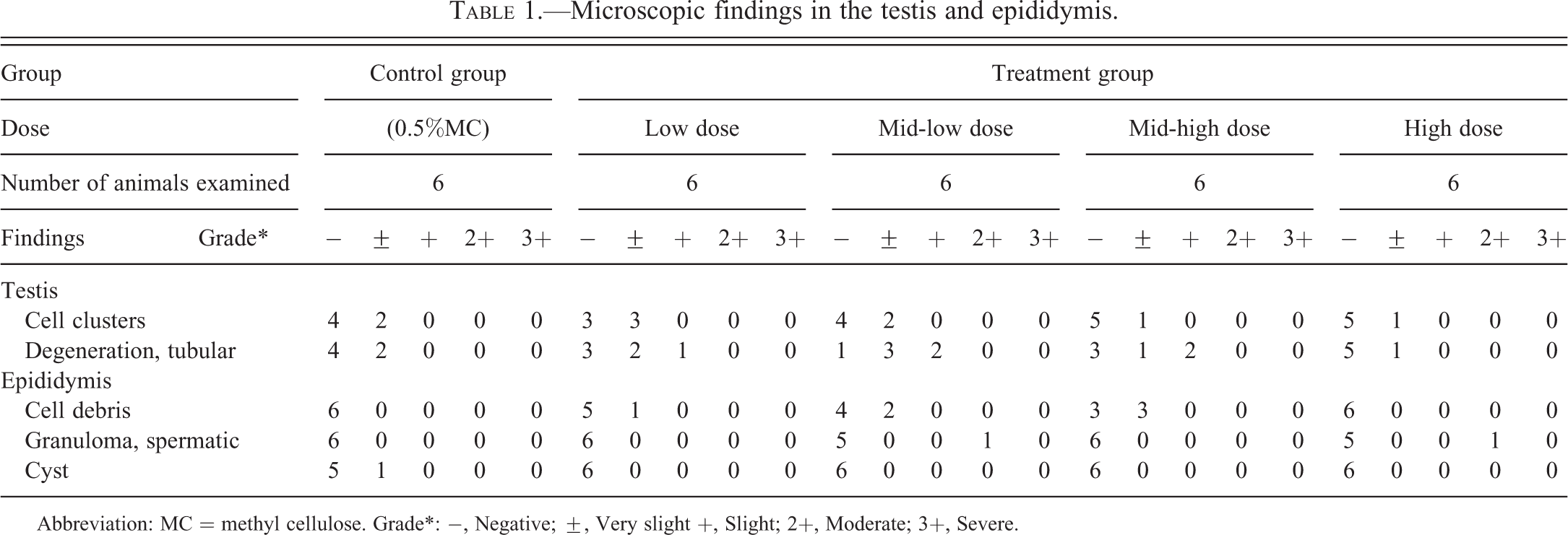
Abbreviation: MC = methyl cellulose. Grade*: −, Negative; ±, Very slight +, Slight; 2+, Moderate; 3+, Severe.
On immunohistochemistry, the cell clusters were positive for claudin-11 (Figure 2A) and GATA-4 (Figure 2B) and were thought to be composed of Sertoli cells. The cell clusters were negative for PCNA (Figure 2C), suggesting no proliferative activity in their constituent cells. In rare cases, cell aggregates (cell debris) that were positive for claudin-11 were observed in the lumen of the epididymis (Figure 2D).
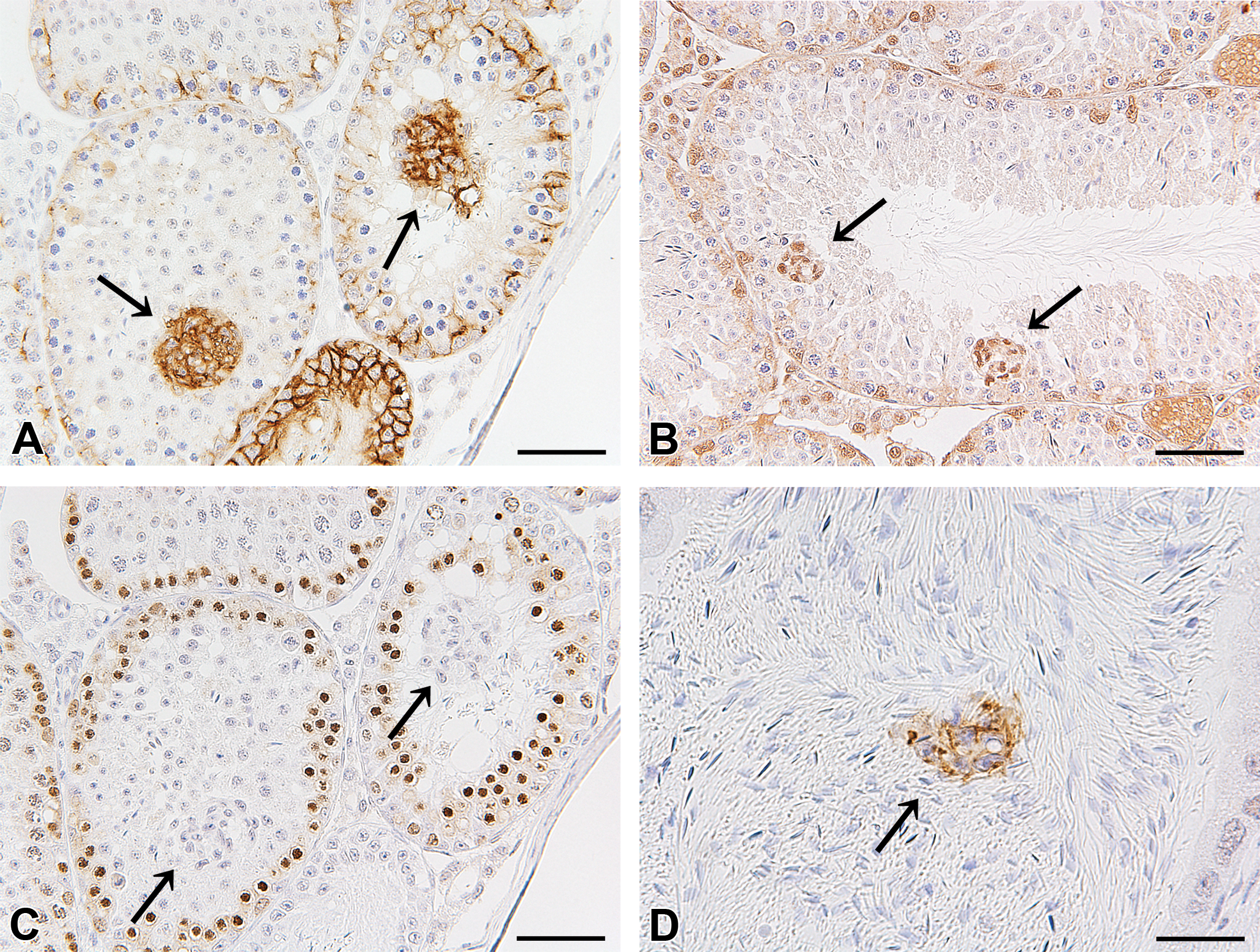
Immunohistochemistry for claudin-11 in the testis (bar: 50 μm) (A), GATA-4 (GATA-binding protein 4) in the testis (bar: 50 μm) (B), proliferating cell nuclear antigen (PCNA) in the testis (bar: 50 μm) (C), and claudin-11 in the epididymis (bar: 20 μm) (D). The cell clusters (arrows) were positive for claudin-11 (A). The nuclei of the cell clusters (arrows) were positive for GATA-4 (B). The cell clusters (arrows) were negative for PCNA (C). Cell aggregates positive for claudin-11 (arrow) were observed in the epididymis in rare cases (D).
The presence of cell aggregates (cell debris) immunoreactive for claudin-11 in the epididymis indicated that some of the Sertoli cell clusters were sloughed from the seminiferous epithelium into the epididymal ducts.
To date, Sertoli cell clusters or similar lesions have only been reported in a few transgenic mice strains such as claudin-11-deficient (Cldn11 −/−) mice (Mazaud-Guittot et al. 2010), Dazl-deficient (Dazl −/−) mice (Schrans-Stassen et al. 2001), Sertoli cell–specific knockout of connexin 43 gene (SCCx43KO) mice (Weider et al. 2011), or in rats rendered cryptorchid (Kopecky, Semecky, and Nachtigal, 2005). Claudin-11 is an obligatory protein for tight junction formation and barrier integrity in the testis and its expression in this organ is Sertoli cell specific. In the absence of claudin-11, spermatogenesis does not proceed beyond meiosis, resulting in male sterility. In Cldn11 −/− mice, GATA-4 immunolabeling demonstrated the presence of abnormal Sertoli cell arrangements in the testes, including groupings of Sertoli cells adjacent to the base of the tubule, round clusters with few cells attached to the basement membrane, and completely detached Sertoli cell clusters filling the tubule lumen. GATA-4-positive Sertoli cell clusters were also observed in the lumen of the epididymis of Cldn11 −/− mice. At the molecular level, the absence of claudin-11 expression is associated with selective changes in several cell junction−related genes, specifically tight junction genes as well as cytoskeleton-related genes. Dazl encodes an RNA-binding protein essential for spermatogenesis. Dazl −/− mice are infertile, lacking any formation of spermatozoa, and the only germ cells present are spermatogonia and a few spermatocytes. Cluster formation of Sertoli cells is a common feature of the testes of these mice, although the reason for the formation of the clusters is not known. In SCCx43KO mice, Sertoli cell−specific knockout of the predominant testicular gap junction protein connexin 43 results in spermatogenic arrest, cell clusters (cell clumps), and still proliferating adult Sertoli cells in the testis. The intratubular cell clusters were mainly formed by abnormal Sertoli cells and their nuclei appeared smaller and more convoluted with numerous heterochromatic patches along the nuclear membrane. The cell clumps were always immunonegative for the proliferation marker Ki-67. Sridharan et al. (2007) hypothesized that the continued proliferation of Sertoli cells in SCCx43 KO mice into adulthood may lead to an excessive buildup of Sertoli cells along the basement membrane of the tubule, and this may result in sloughing and loss of some Sertoli cells as their proliferation continues.
According to the above information from the transgenic mice, some genes associated with cell junctions or cytoskeletons are considered to be related to cluster formation of Sertoli cells and the present changes of the Sertoli cells might also be related to an alteration in genes that are associated with the organization of their cytoskeleton or proteins, which establish intercellular junctions.
To our knowledge, this is the first report of Sertoli cell clusters as a spontaneous lesion in any animal species except for those reported in transgenic or surgically altered animals.
Footnotes
Author Contribution
Authors contributed to conception or design (AA, KK); data acquisition, analysis, or interpretation (AA, KK, KM, YY, YK, KT, ET, AT, TS); drafting the manuscript (AA); and critically revising the manuscript (HA, GA, JW). All authors critically revised the manuscript, gave final approval, and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.