
Abstract
Medulloblastomas (MBs) are thought to be derived from granular cell precursors in the external granular layer (EGL) of the developing cerebellum. Heterozygous patched1 (Ptch1) knockout mice develop MBs that resemble those in humans when the sonic hedgehog (Shh) signaling pathway is activated. The present study was conducted to evaluate postnatal effects of a Shh signaling inhibitor, cyclopamine, on the development of MBs in Ptch1 mice. Ptch1 and wild-type mice were treated daily with subcutaneous cyclopamine at 40 mg/kg or vehicle from postnatal day (PND) 1 to PND14, and the subsequent development of MBs and preneoplastic lesions was examined up to week 12 (W12). Proliferative lesions in the cerebellum, MBs, and preneoplastic lesions were only detected in Ptch1 mice. Cyclopamine treatment resulted in a statistically significant reduction in the incidence and/or area of proliferative lesions at PND14 and 21. The trend of decreasing preneoplastic lesions persisted up to W12. At PND7, cyclopamine treatment reduced the width and proliferation of the EGL regardless of genotype. These results indicate that inhibition of Shh signaling during cerebellar development has prolonged inhibitory potential on MB development in Ptch1 mice. This inhibitory potential might be related to inhibition of EGL proliferation, including preneoplastic MB cells.
Keywords
Introduction
Medulloblastoma (MB) is the most common malignant brain tumor in children (Bartlett, Kortmann, and Saran 2013; Dhall 2009; Hatten and Roussel 2011; Jones et al. 2012). Although exposure to environmental compounds and radiation during the developmental period and early life stages has been thought to be critically involved in the causation of tumors, little is known about the etiology of childhood brain tumors (Bunin et al. 2006; Birnbaum and Fenton 2003; Dietrich et al. 2005; Mckean-Cowdin et al. 2003; Norman, Holly, and Preston-Martin 1996; Takahashi et al. 2012).
Molecular analysis of sporadic human MBs revealed activation of the sonic hedgehog (Shh) signaling pathway caused by the loss of patched1 (Ptch1) and mutations in other components of the Shh pathway. Ptch1 encodes a receptor for Shh, Ptch1, and is one of the key genes related to MB formation in humans (Dhall 2009; Raffel 2004). Pathway activation is triggered by binding of Shh to Ptch1; in the absence of Shh, the activity of Smoothened (Smo) is suppressed. Shh binding to Ptch1 or mutational inactivation of Ptch1 relieves the inhibition on Smo culminating in the activation of one or more of the Gli1 transcription factors that regulate the expression of downstream targets (Hahn et al. 1999; Huse and Holland 2010; Roussel and Hatten 2011).
Heterozygous Ptch1 knockout mice (Ptch1 mice) have been used as a valuable model of MB due to the high incidence of MBs (14–30%) and the morphological and molecular similarities to human MBs (Corcoran and Scott 2001; Goodrich et al. 1997; Hahn et al. 1999; Pazzaglia 2006; Raffel 2004; Wetmore, Eberhart, and Curran 2000). Moreover, it has been reported that Shh signaling is activated in MBs of Ptch1 mice (Dyer 2004; Goodrich et al. 1997; Oliver et al. 2005; Wetmore, Eberhart, and Curran 2000).
MBs in humans and Ptch1 mice are thought to be derived from residual granule cell precursors (GCPs) located in the external granule cell or external granular (germinal) layer (EGL) of the cerebellum (Behesti and Marino 2009; Roussel and Hatten 2011). During normal cerebellar development in mice, GCPs proliferate postnatally, and the proliferative period peaks between postnatal days (PND) 4 and 8 (Behesti and Marino 2009; Roussel and Hatten 2011). For GCP proliferation, Shh signaling is required (Lewis et al. 2004; Raffel 2004; Roussel and Hatten 2011; Vaillant and Monard 2009; Wallace 1999).
While Ptch1 mice have been accepted as a useful MB model, it takes an extended amount of time to detect the efficacy of treatments using end points such as the clinical signs of increased intracranial pressure due to MB or death after the long latent period of 9 to over 12 months (Ayrault et al. 2009; Briggs et al. 2008; Ecke et al. 2008; Farioli-Vecchioli et al. 2007; Kimura et al. 2005; Pazzaglia et al. 2006; Pazzaglia et al. 2009; Pogoriler et al. 2006; Uziel et al. 2005; Wetmore, Eberhart, and Curran 2001).
Recently, we found that the earliest signs of MBs and their preneoplastic lesions in Ptch1 mice were morphologically detectable within 2 weeks after birth (Matsuo et al. 2013). Changes in these indicators are thought to be early novel end points for assessment of the modifying effects of chemicals and/or agents on MB development in studies using Ptch1 mice.
Cyclopamine is a naturally occurring alkaloid of the corn lily Veratrum californicum that causes cyclopia in sheep by blocking the Shh/Ptc/Smo signaling pathway (Ecke et al. 2008; Heretsch et al. 2010a and b; Lipinski et al. 2008). The inhibitory effects of cyclopamine on the Shh pathway have been reported in a number of in vitro and in vivo studies (Ecke et al. 2008; Scales and Sauvage 2009; Stecca and Ruiz i Altaba 2002). Furthermore, cyclopamine was shown to inhibit the in vitro growth of human MB cell lines (Berman et al. 2002), and other studies have examined the effect of cyclopamine in vivo on spontaneously developing Hedgehog (Hh)-dependent tumors including MBs (Sanchez and Ruiz i Altaba 2005; Ecke et al. 2008; Fan et al. 2011; Coon et al. 2010). However, the effects of developmental exposure to cyclopamine using Ptch1 mice have not been reported. Here, we used the Ptch1 mouse model of MB to test if cyclopamine treatment from PND 1 to 14 is able to inhibit tumor growth and to affect cerebellar development.
Materials and Methods
Animals
Heterozygous ptch1 knockout mice (Ptch1 mice) maintained on a mixed C57B1/6 × 129Sv background were obtained from the Jackson Laboratory (Bar Harbor, ME). Mice were housed in our facility in polycarbonate cages with wood chip bedding and maintained in an air-conditioned animal room (temperature 24°C ± 1°C, relative humidity 55% ± 5%, 12-hr light–dark cycle) with a basal diet (CRF-1, Oriental Yeast Co., Tokyo, Japan) and tap water available ad libitum. Animals were genotyped by polymerase chain reaction amplification of genomic DNA extracted from tails (Matsuo et al. 2013). The experimental protocol regarding animal use was reviewed and approved by the Animal Care and Use Committee of the National Institute of Health Sciences, Japan.
Cyclopamine Treatment
Cyclopamine (CAS No. 4449-51-8, purity >99%, LC Laboratories, Woburn, MA) was dissolved in ethanol then suspended in triolein (Wako, Osaka, Japan). Animals were weighed just prior to each injection, and cyclopamine (40 mg/kg/day) or vehicle (triolein: ethanol, 4:1 vol/vol) was injected subcutaneously from PND1 to PND14. The dosing volume, dose of cyclopamine and vehicle, and route of administration were selected based on previous studies (Berman et al. 2002; Lipinski et al. 2008) and our preliminary study. The administration period, from PND1 to PND14, was chosen to match the developmental period in the cerebellum in which GCP proliferation is prominent (Behesti and Marino 2009; Haldipur et al. 2012; Vaillant and Monard 2009). This duration also covers the highly susceptible period of Ptch1 mice to X-ray irradiation and carcinogens (Pazzaglia et al. 2002; Takahashi et al. 2012). The number of mice in each group at each time point is listed in Table 1. At least 5 wild-type mice and 9 Ptch1 mice from 3 to 9 dams were allocated to each group.
Body weight, brain weight, and cerebellum weight.
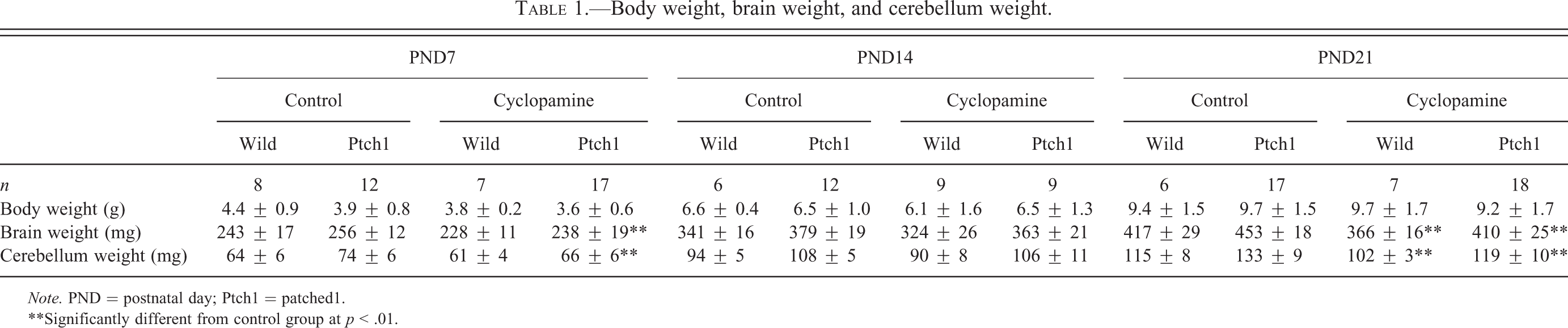
Note. PND = postnatal day; Ptch1 = patched1.
**Significantly different from control group at p < .01.
Necropsy
To examine the effects of cyclopamine on early cerebellar development, Ptch1 and wild-type mice at PND7 were subjected to necropsies. To examine the effect of cyclopamine on MB development, necropsy was performed at PND14 and PND21. In addition, at postnatal week 12 (W12), Ptch1 and wild-type mice were examined to determine whether the effects of cyclopamine treatment on the cerebellum during the developmental period persist after the maturation of the cerebellum. At necropsy, all mice were euthanized under deep anesthesia with isoflurane.
Tissue Processing and Histopathology
After necropsy, brains were removed and weighed before fixation in 10% neutral buffered formalin. Midsagittal (right hemisphere) and cross (left hemisphere) sections of the cerebella were routinely processed for paraffin embedding, sectioned, and stained with hematoxylin and eosin. The prepared histopathological specimens were examined by light microscopy. To examine the effect of cyclopamine on MB development, we counted preneoplastic lesions such as thickenings of the EGL at PND14 and Ki-67-positive foci at PND21 and W12 in addition to MBs. MBs were divided into 2 types according to the previous study (Matsuo et al. 2013): a focal MB occupying 1 to 2 lobules of the cerebellum was defined as a small MB and an advanced MB spreading over 3 or more lobules was defined as a large MB.
Immunohistochemistry
Antibodies used for immunohistochemistry included monoclonal rat anti-mouse Ki-67 (Clone TEC-3, Dako Cytomation, Glostrup, Denmark) as a proliferation marker, monoclonal rabbit anti-cleaved caspase-3 (Clone 5A1E, Cell Signaling, Danvers, MA) as an apoptotic marker, and monoclonal mouse anti-NeuN (Clone A60, Millipore, Billerica, MA) as a mature granule cell marker. A streptavidin–biotin labeling method was performed with the anti-Ki-67 antibody using a polyclonal rabbit anti-rat biotinylated IgG (Dako Cytomation) and streptavidin-conjugated horseradish peroxidase (Dako Cytomation). A polymer labeling method was performed for the anti-cleaved caspase-3 and anti-NeuN antibodies using the Histofine Simple Stain kit (Nichirei Biosciences Inc., Tokyo, Japan). The immunoreactions were visualized by a peroxidase-diaminobenzidine reaction. The sections were then lightly counterstained with hematoxylin.
Morphometric Assessment
For total area of proliferative lesions in the cerebellum, photomicrographs of the cerebellar sections were taken with a digital camera attached to a microscope (DP71, Olympus Corp., Tokyo, Japan), then measurements were made using image analysis software (WinROOF, Version 5.7.1, Mitani Corp., Tokyo, Japan). The total area of proliferating cells in the cerebellum, including Ki-67-positive foci and MBs (Matsuo et al. 2013), was measured as the Ki-67-positive cell area, and the ratio of the total proliferating area to the total area of the cerebellum was calculated for Ptch1 mice at PND21 and W12. For width of the EGL and outer layer of EGL, slides stained with Ki-67 antibody were scanned using Aperio ScanScope (Aperio, Vista, CA), then measurements were made using image analysis software (ImageScope, Version 10.2.1.2315, Aperio, Vista, CA). The width of the EGL and the outer layer of the EGL of each mouse at PND7 was determined by 5 measurements selected at random from the 3rd cerebellar lobule.
Statistical Analysis
For body weight, organ weight, proliferating area of the cerebellum, and width of the EGL and outer layer of the EGL, values of cyclopamine-treated mice at each time point were compared with the corresponding vehicle controls using the Student’s t-test following a test for equal variance. Incidence of histopathological findings was compared using Fisher’s exact probability test.
Results
General Remarks
There were no significant differences in body weight at each time point when control and cyclopamine-treated mice were compared by genotype (Table 1). All wild-type and Ptch1 mice survived the duration of the experimental period. In addition, no clinical signs of tumor progression were detected in either genotype with or without cyclopamine treatment up to 12 weeks of age.
Absolute weights of the whole brain and cerebellum significantly decreased in Ptch1 mice in the cyclopamine group at PND7, and in both genotypes in the cyclopamine group at PND21 compared to the control group (Table 1).
Effect of Cyclopamine on Proliferative Lesions in the Cerebellum of Ptch1 Mice
No proliferative lesions were detected in the cerebellum of wild-type mice at any time point (Figure 1). The incidence of lesions was calculated as the number of animals that had a specific lesion (e.g., Ki-67-positive foci) in the cerebellum relative to the total number of animals examined (Figures 2 and 3). At PND14, the incidence of thickened areas in the EGL was significantly decreased in the cyclopamine group in Ptch1 mice (Figure 3). In addition, the incidence of Ki-67-positive foci and small MBs was significantly decreased in the cyclopamine group in Ptch1 mice at PND21 (Figure 3). The incidence of proliferative lesions in the cerebellum of the cyclopamine group in Ptch1 mice at W12 was decreased but was not statistically significant (Figure 3).
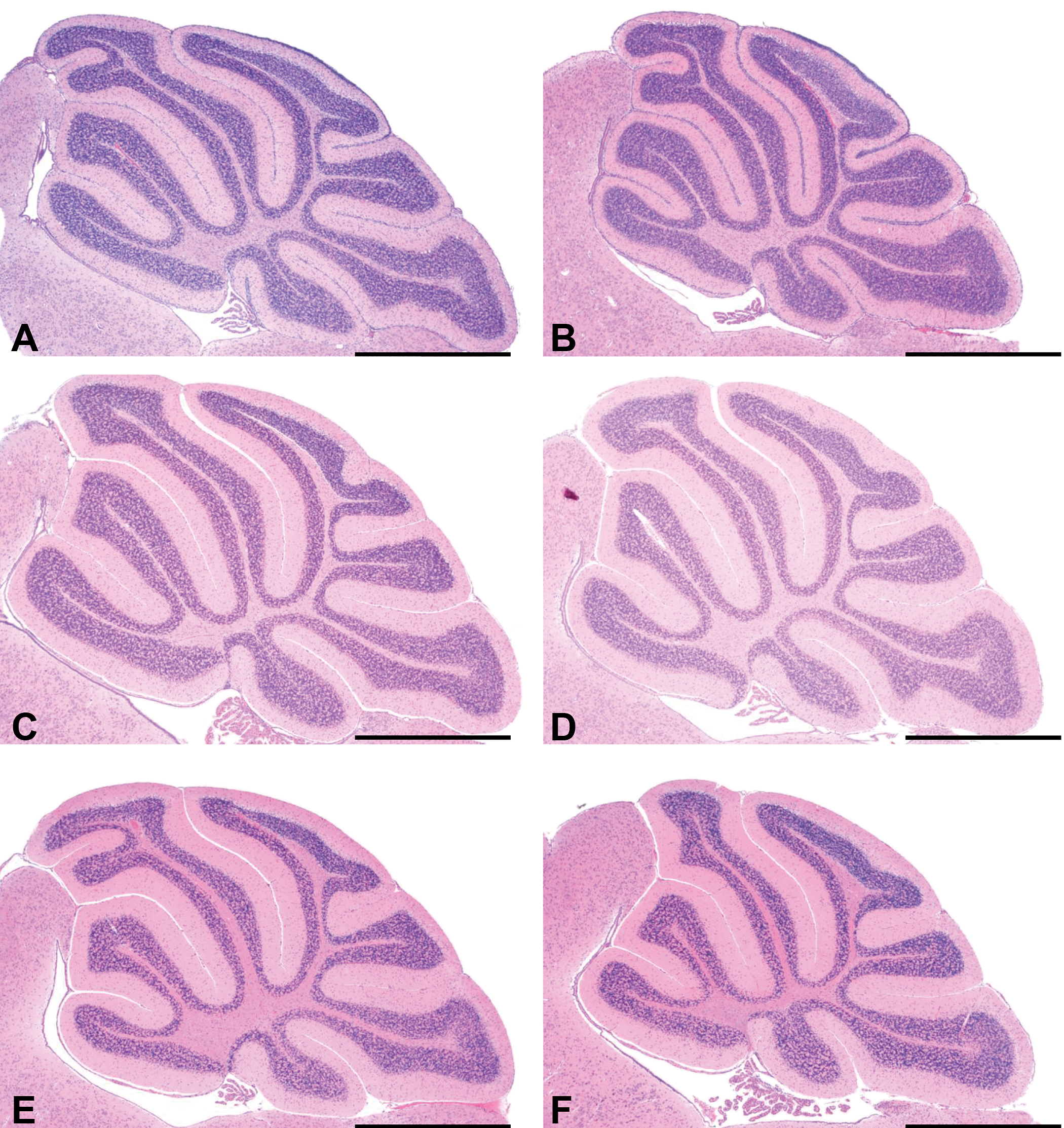
Representative images of the cerebellum from control (left) and cyclopamine-treated (right) wild-type mice at PND14 (A, B), 21 (C, D) and W12 (E, F). No proliferative lesions were detected in the cerebellum of wild-type mice at any time point. Scale bar: 1000 μm. Note. PND = postnatal day; W12 = postnatal week 12.
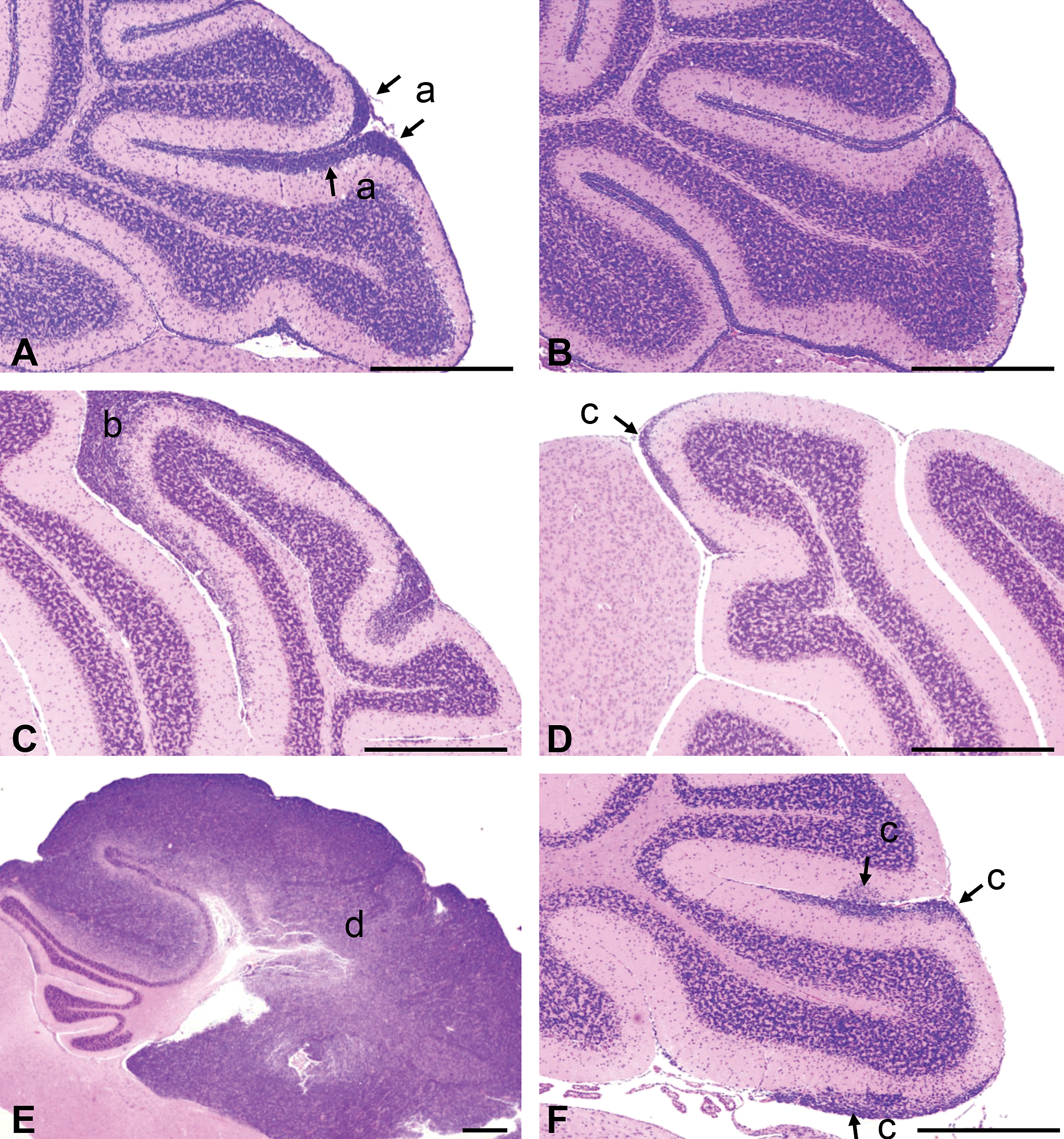
Representative proliferative lesions in the cerebellum of control (left) and cyclopamine-treated (right) Ptch1 mice at PND14 (A, B), 21 (C, D) and W12 (E, F). Proliferative lesions in the cerebellum were classified into the following 4 types: (a) thickened area of the EGL, (b) small MB, (c) Ki-67 positive focus, and (d) large MB. Scale bar: 500 μm. Note. PND = postnatal day; W12 = postnatal week 12; Ptch1 = patched1; EGL = external granular layer; MB = medulloblastoma.
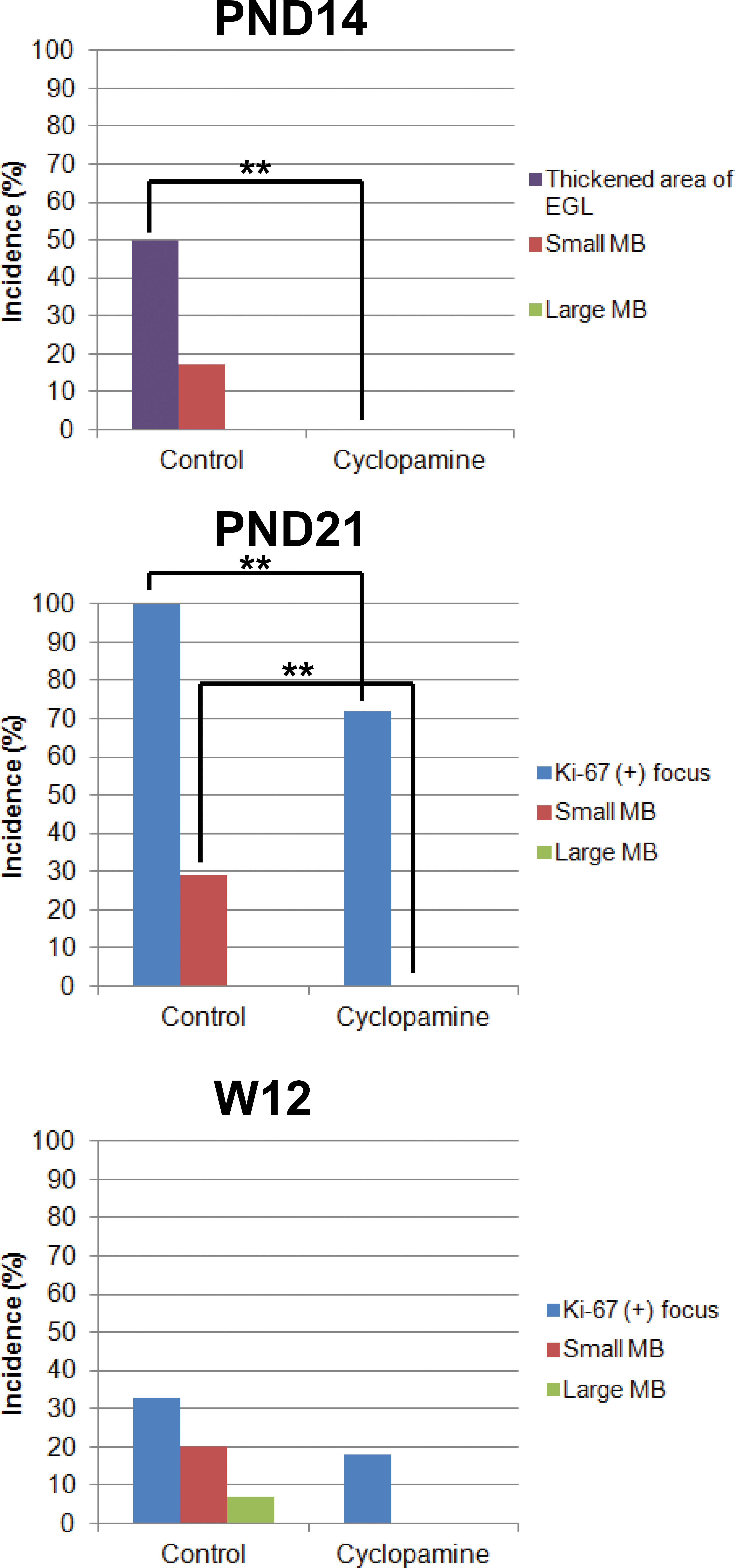
Incidence of proliferative lesions in the cerebellum of control and cyclopamine-treated Ptch1 mice at PND14, 21, and W12. **Significantly different from control group; p < .01. Note. PND = postnatal day; W12 = postnatal week 12; Ptch1 = patched1.
Immunohistochemistry for Ki-67 revealed that the total area of proliferative lesions in the cerebellum was significantly decreased in the cyclopamine group in Ptch1 mice at PND21 (Figure 4). Additionally, a decreasing trend in the total area of proliferative lesions at W12 was observed (Figure 4).
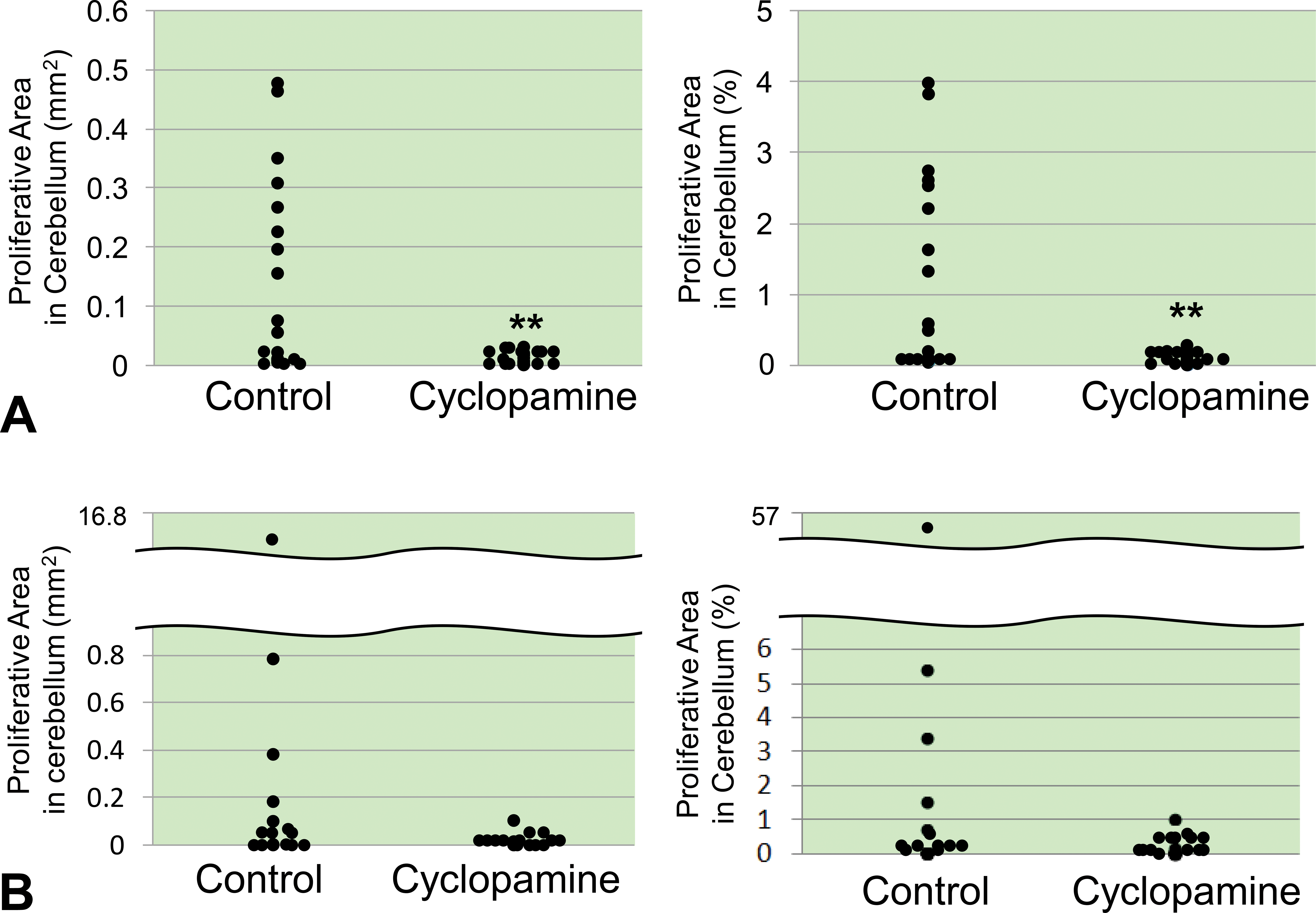
Total area of proliferative lesions in the cerebellum of control and cyclopamine-treated Ptch1 mice at PND21 (A) and W12 (B). The total area of the proliferative lesions was calculated for each animal as the sum of areas of the Ki-67 positive foci and MBs observed in a cerebellum slide stained for Ki-67. **Significantly different from the control group; p < .01. Note. PND = postnatal day; W12 = postnatal week 12; Ptch1 = patched1; MB = medulloblastoma.
Effects of Cyclopamine on Cerebellar Development
To examine the specific inhibitory effects of cyclopamine on the development of MBs and their preneoplastic lesions, we also performed histopathology of the cerebellum of control and cyclopamine-treated mice during the early developmental period of the cerebellum at PND7. The width of the EGL was noticeably thinned in the cyclopamine group compared to the control group regardless of genotype at PND7 (Figures 5A and 6, pictures of wild-type mice not shown). The thinning of the EGL was mainly observed in lobules 2 through 4/5 of the cerebellum and was obscure in lobules 6 through 10. Immunohistochemical staining with the anti-Ki-67 antibody revealed that the outer layer of the EGL, which is composed of proliferating GCPs, was also thinned in Ptch1 mice in the cyclopamine group (Figures 5B and 6, pictures of wild-type mice not shown). Decreasing trend was observed in the outer layer of the EGL in cyclopamine group of wild-type mice (Figure 5B). There was no increase in cleaved caspase-3-positive apoptotic cells in the EGL of the cyclopamine group compared to the control group (Figure 6).
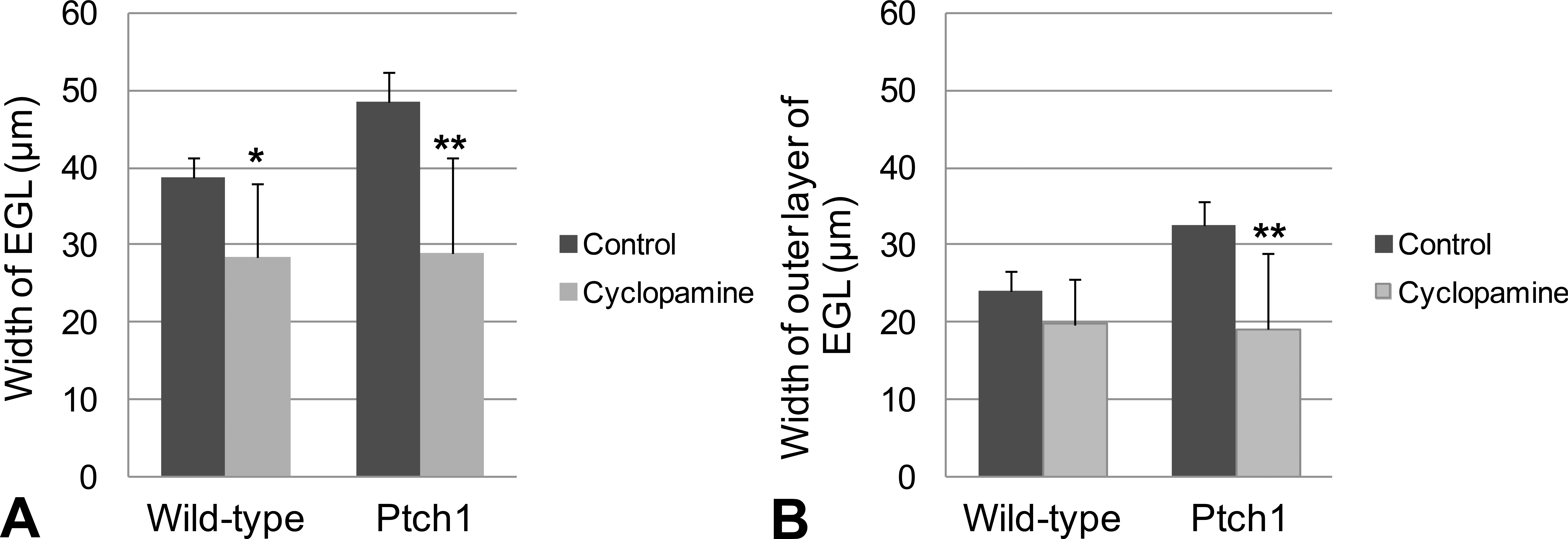
The width of the EGL (A) and the outer layer of the EGL (B) of control and cyclopamine-treated Ptch1 and wild-type mice at PND7. *, **Significantly different from the control group at p < .05 and p < .01, respectively. Note. PND = postnatal day; Ptch1 = patched1; EGL = external granular layer.
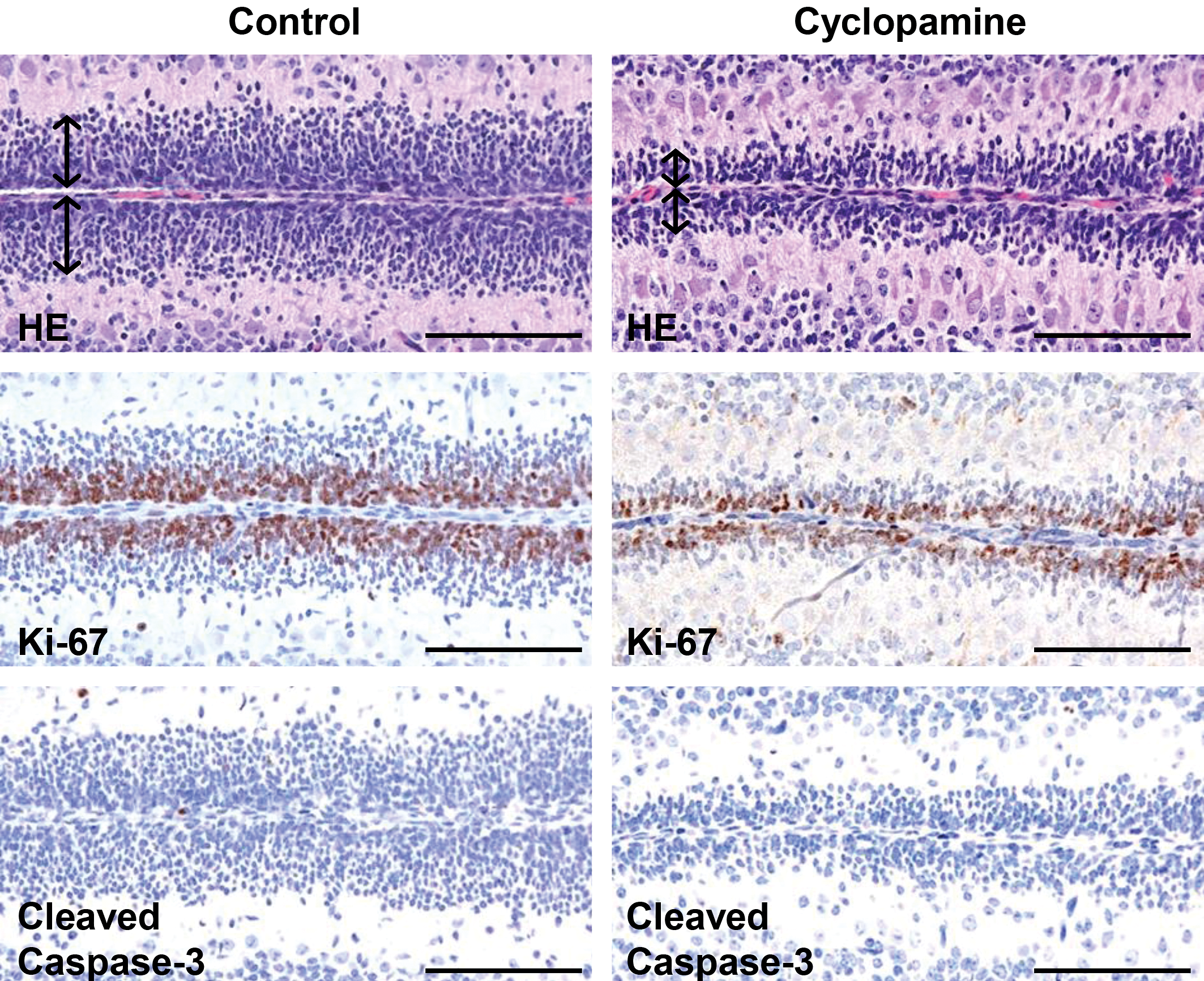
Histopathology of the cerebellum of control (left column) and cyclopamine-treated (right column) Ptch1 mice at PND7. The EGL (arrows) was thinned in the cyclopamine group (HE, top of right column). Thinning of the Ki-67 positive layer in the EGL was also observed in the cyclopamine group (Ki-67 staining, middle of right column). There was no difference in the number of apoptotic cells in the EGL of the cyclopamine group (cleaved caspase-3 staining, bottom of right column) compared to controls. Scale bar: 100 μm. Note. PND = postnatal day; Ptch1 = patched1; EGL = external granular layer; HE = hematoxylin and eosin.
Histopathological examination revealed that cells morphologically resembling nuclei of the internal granular layer were distributed parallel to the Purkinje cell layer in the deep molecular layer of the cyclopamine group at PND14 and 21 regardless of genotype (Figure 7). These nuclei were strongly labeled for NeuN (Figure 8). These findings were also present in the cerebellum of the cyclopamine-treated group at W12 (Figures 7 and 8). Furthermore, misalignment of the Purkinje cells was scattered in the cyclopamine group at PND14 and 21, and this finding was observed up to W12 (Figure 9). These findings were mainly observed in lobules 2 through 4/5 of the cerebellum and were obscure in lobules 6 through 10 in both genotypes (pictures of wild type not shown).
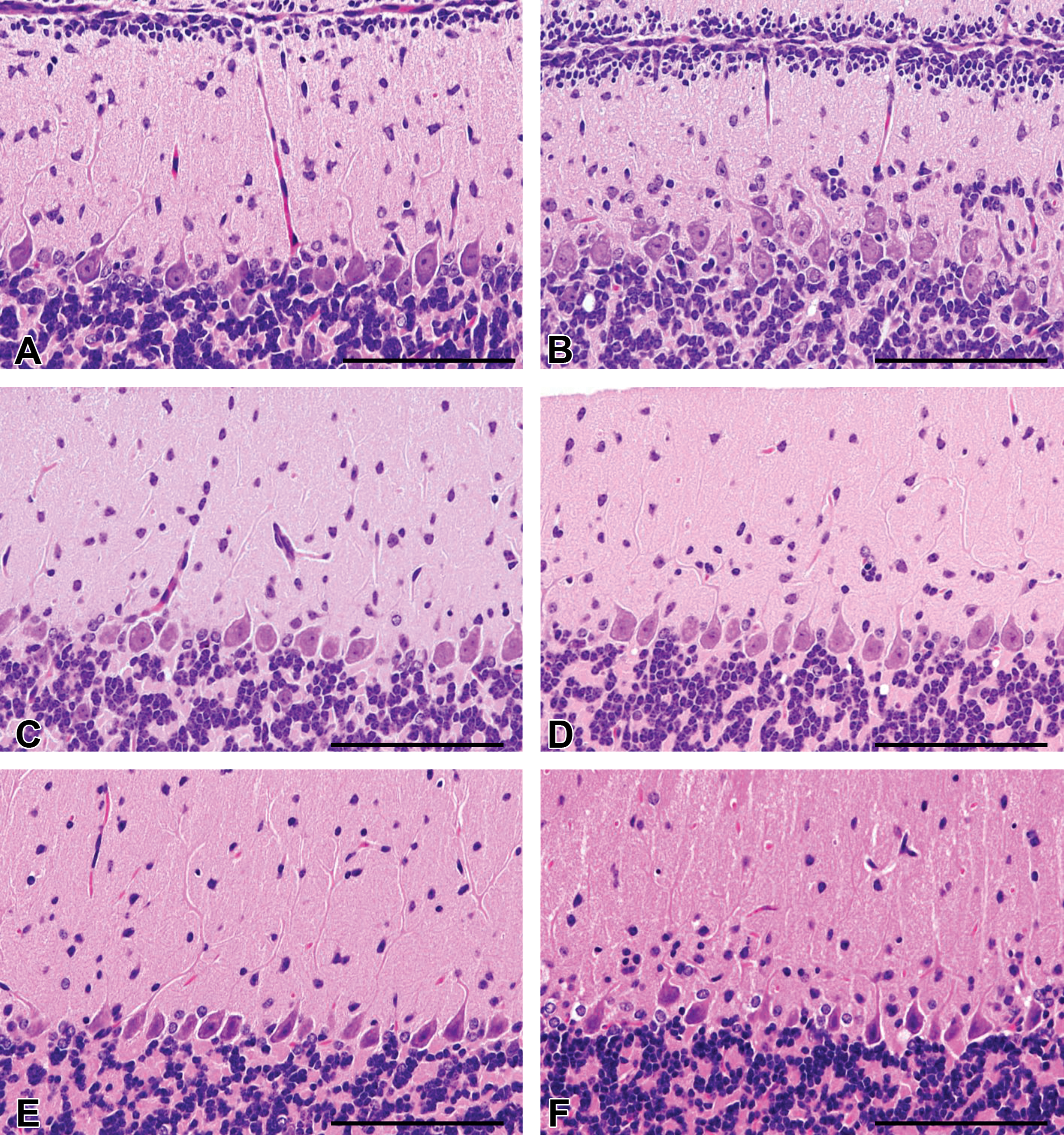
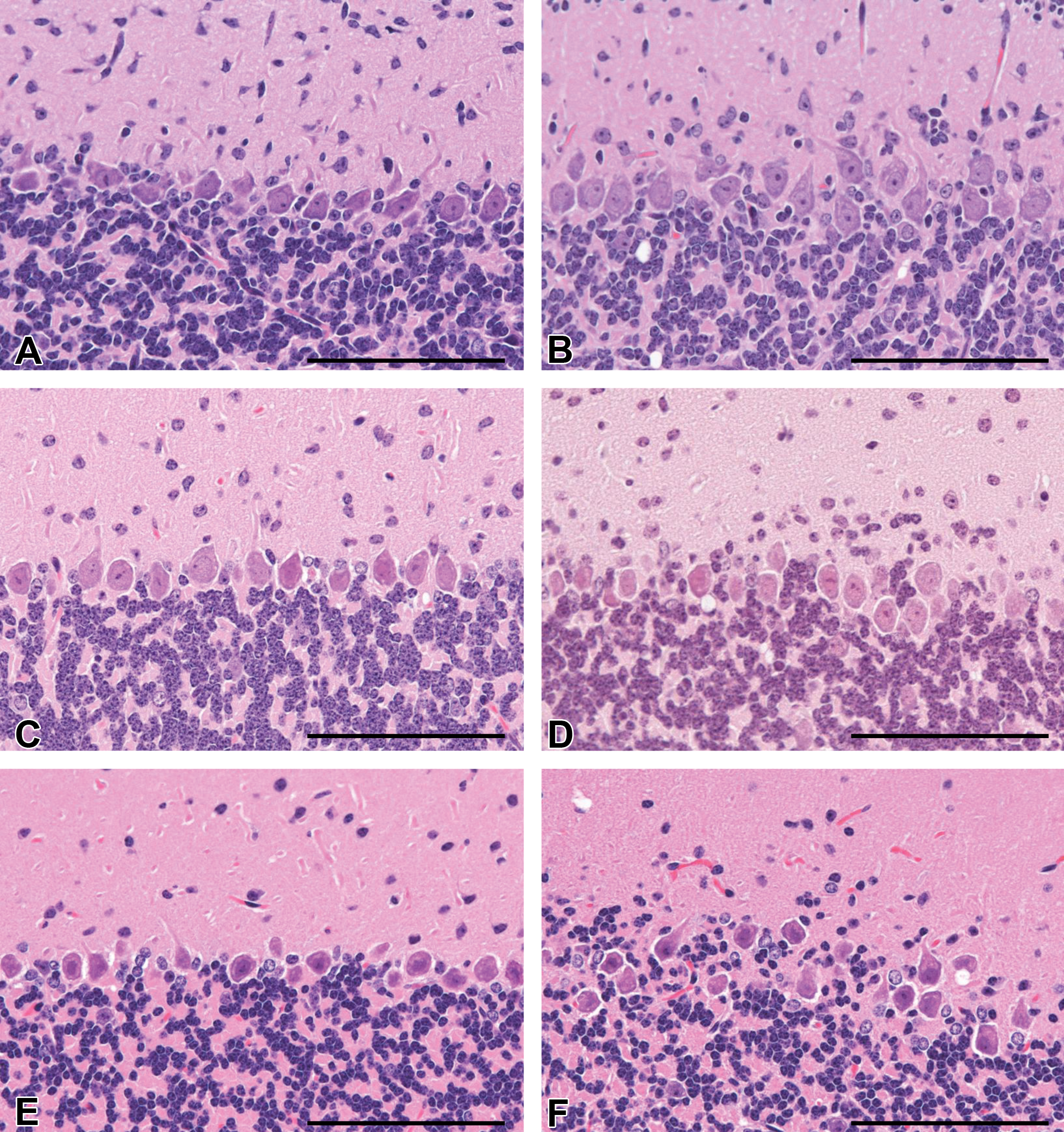
Histopathological findings observed in the molecular layer of the cerebellum from control (left column) and cyclopamine-treated (right column) Ptch1 mice at PND14 (A and B), 21 (C and D), and W12 (E and F) (HE). Granular cells, which resemble cells of the internal granular layer, were distributed parallel to the Purkinje cell layer in the deeper molecular layer of cyclopamine-treated Ptch1 mice at PND14 (B), 21 (D), and W12 (F). Scale bar: 100 μm. Note. PND = postnatal day; W12 = postnatal week 12; Ptch1 = patched1; HE = hematoxylin and eosin.
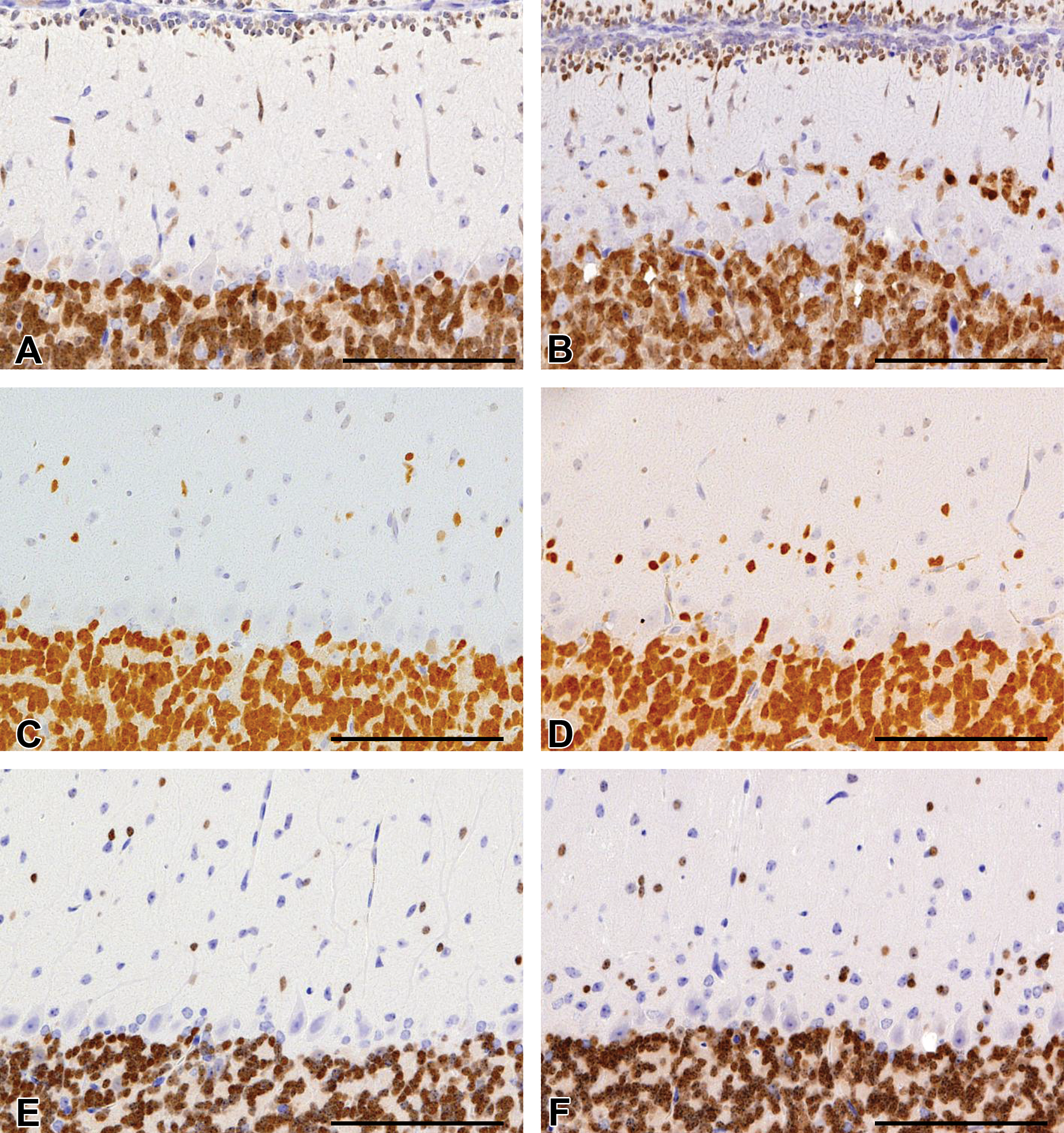
Immunohistochemistry for NeuN of the cerebellum from control (left column) and cyclopamine-treated (right column) Ptch1 mice at PND14 (A and B), 21 (C and D), and W12 (E and F). Granular cells of the internal granular layer were strongly positive for NeuN in all animals. In addition, NeuN positive cells were distributed parallel to the Purkinje cell layer in the deeper molecular layer of cyclopamine-treated Ptch1 mice at PND14 (B), 21 (D), and W12 (F). Scale bar: 100 μm. Note. PND = postnatal day; Ptch1 = patched1; W12 = postnatal week 12.
Histopathological findings observed in the Purkinje cell layer of the cerebellum from control (left column) and cyclopamine-treated (right column) Ptch1 mice at PND14 (A and B), 21 (C and D), and W12 (E and F) (HE). Misalignment of Purkinje cells was observed in the cyclopamine group at PND14 (B), 21 (D), and W12 (F). Scale bar: 100 μm. Note. PND = postnatal day; Ptch1 = patched1; W12 = postnatal week 12; HE = hematoxylin and eosin.
Discussion
Currently, human MBs have been classified into at least 4 distinct subtypes by molecular studies (Ellison et al. 2011; Northcott et al. 2011; Jones et al. 2012; Kool et al. 2012; Kawauchi et al. 2012). MBs of the Shh subgroup result from inactivating mutations of PTCH1 or mutations of other components of the Shh signaling pathway (Kawauchi et al. 2012). It is known that the expression of Gli1 is increased in MBs of Ptch1 mice compared to expression in the normal cerebellum, suggesting activation of the Shh signaling pathway in these tumors (Raffel 2004). Recently, many Shh pathway inhibitors such as cyclopamine, HhAntag, GDC-0449, and IPI-926 have been identified and tested in preclinical and clinical studies targeting MB and other Shh pathway–activated tumors (Gajjar et al. 2013; Gupta, Takebe, and Lorusso 2010; Lee et al. 2012; Low and Sauvage 2010; Scales and Sauvage 2009). The Ptch1 mouse has been used in various studies as an Shh-subtype MB model (Behesti and Marino 2009; Sanchez and Ruiz i Altaba 2005).
During postnatal cerebellar development in mice, proliferation of GCPs occurs throughout the 2 weeks after birth and GCPs migrate into the internal granular layer by PND21 (Behesti and Marino 2009; Haldipur et al. 2012; Vaillant and Monard 2009). During postnatal cerebellar development, Ptch1 mice show a high incidence of preneoplastic MB lesions such as thickened areas in the EGL and Ki-67 positive foci (Matsuo et al. 2013). The present study clearly showed that postnatal exposure to cyclopamine inhibited the occurrence of preneoplastic MB lesions and small MBs in Ptch1 mice up to 1 week after the treatment period. Concurrently, the reduction in the total area of proliferative lesions in the cyclopamine group indicated that cyclopamine treatment inhibited not only the development of preneoplastic lesions but also the expansion of these lesions. Furthermore, the inhibitory potential persisted for 2 months after treatment.
During normal cerebellar development, Shh is an important signal that regulates GCP proliferation in the EGL (Haldipur et al. 2012; Lewis et al. 2004; Vaillant and Monard 2009; Wallace 1999). GCPs express Ptch1, the receptor for Shh, and proliferate in response to Shh secreted by the Purkinje cells (Haldipur et al. 2012; Lewis et al. 2004; Vaillant and Monard 2009). Examination of the cerebellum at the peak period of GCP proliferation, PND7, provided evidence that the inhibitory effect of cyclopamine on GCP proliferation might be related to the thinning of the EGL, a place of occurrence of preneoplastic cells of MBs, in wild-type and Ptch1 mice. Immunohistochemistry indicated that thinning of the EGL was associated with decreased proliferative activity rather than increased apoptosis of GCPs by cyclopamine. In this study, at PND7, the outcome of GCPs treated with cyclopamine was consistent with GCPs treated with neutralizing anti-Shh antibodies in neonatal mice (Wallace 1999). It is known that cyclopamine inhibits the growth of cultured MB cells and allografts by inhibiting Smo (Berman et al. 2002; Dahmane et al. 2001). Furthermore, it has been reported that cyclopamine treatment inhibited MB growth in vivo using Ptc1+/−/p53−/− mice and genetically engineered mice overexpressing hepatocyte growth factor (HGF) and Shh by inducing a potent apoptotic death response in tumor cells and by a suppressive effect on proliferation (Coon et al. 2010; Sanchez and Ruiz i Altaba 2005). In addition, another Smo-binding antagonist, HhAntag, completely eliminated MBs by blocking tumor cell proliferation and stimulating apoptosis (Gupta, Takebe, and Lorusso 2010; Romer et al. 2004). Taken together, the decreased proliferative activity observed in cyclopamine-treated mice in this study is thought to be attributed to the inhibition of Smo and downstream signals. Since Ptch1 haploinsufficiency alone is considered to be insufficient for tumor induction, additional genetic lesions are required for tumorigenesis (Wetmore, Eberhart, and Curran 2000; Corcoran and Scott 2001). Our data suggest that cyclopamine treatment during cerebellar development reduced production of cells that will give rise to MBs and the subsequent chances for additional mutations to occur in those cells, by decreasing proliferation of GCPs and preneoplastic cells.
Cerebellum volume dramatically increases during neonatal development (Lewis et al. 2004). This increase in size is predominantly due to rapid proliferation and expansion of granular cells, which are the most abundant neuronal population in the mature brain (Lewis et al. 2004). In addition, it is known that Shh is required for proliferation of precursor cells of the cerebral neocortex and tectum in postnatal mice in addition to the cerebellum (Dahmane et al. 2001; Stecca and Ruiz i Altaba 2002). Therefore, the decrease in cerebellum weight in the cyclopamine-treated group was considered to be a result of the decreased proliferation of GCPs in the EGL. Furthermore, this suggests that inhibition of Shh signaling by cyclopamine has a similar effect on postnatal cerebral development, as it does on cerebellar development and causes a decrease in overall brain weight.
NeuN-positive granular cells in the deep molecular layer of the cyclopamine-treated group may undergo precocious maturation during their migration to the internal granular layer. Abnormal Purkinje cells similar to those observed in this study have also been reported in Shh-deficient mice and were thought to be a secondary effect due to loss of GCPs (Lewis et al. 2004). In our study, the thinning of the EGL, the presence of ectopic NeuN-positive granular cells, and the misalignment of Purkinje cells are all thought to be caused by cyclopamine treatment and are related to each other because these lesions shared a common location in the cerebellum. Also, the presence of these lesions at W12 indicated that the effects of cyclopamine persisted after the completion of cerebellar development.
Whereas our previous study revealed that more than half of the Ki-67-positive foci and small MBs were located in lobules 6 through 10 of the cerebellum (Matsuo et al. 2013), in the present study the inhibitory effect of cyclopamine was observed mainly in lobules 2 through 4/5 of the cerebellum at PND7. The differences in regions that were highly affected by cyclopamine treatment during the developmental period and the common site of MB formation in the cerebellum of Ptch1 mice might result from incomplete inhibitory effects on the MBs. Another possible explanation for these differences is the potency of cyclopamine, which has recently been shown to be only a moderate inhibitor of hedgehog (Hh) target gene transcription compared to several other more potent synthetic inhibitors of Hh signaling. However, given that cyclopamine was the first Hh signaling inhibitor available, it has been used in many in vitro and in vivo studies (Heretsch et al. 2010a and b; Fan et al. 2011; Gupta, Takebe, and Lorusso 2010; Scales and Sauvage 2009).
In summary, cyclopamine treatment during cerebellar development has inhibitory effect on proliferation of the EGL and development of MB and preneoplastic lesions in Ptch1 mice, and tendency to inhibit these proliferative lesions is prolonged after treatment. This demonstrates that the Ptch1 mouse is a good model for studying the modifying effects of chemical exposure on MB development during the developmental period.
Footnotes
Acknowledgments
We thank Ms. Tomomi Morikawa for technical assistance in conducting the study and genotyping, and Ms. Ayako Saikawa and Yoshimi Komatsu for technical assistance in histopathological preparation.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partly supported by Health and Labor Sciences Research Grants, Research on Risk of Chemical Substances, Ministry of Health, Labor and Welfare [H25-Toxicol-003].