
Abstract
In this study, we have investigated the immunoexpression of peptide hormones and mediators associated with human islet cell tumors in a group of proliferative islet cell lesions in F344 rats including islet cell hyperplasias, adenomas, and carcinomas, as defined by conventional histopathologic criteria. All proliferative islets expressed synaptophysin, although decreased expression intensity was observed in hyperplasias and adenomas. Most of the proliferative lesions expressed insulin, which generally decreased as lesions progressed toward malignancy. The distribution of glucagon, somatostatin, and gastrin-expressing cells was altered in proliferative islet lesions but did not comprise a large proportion of cells. Islet cell tumors were associated with increased nuclear expression of cyclin-dependent kinase 4 as well as increased proliferating cell nuclear antigen and decreased β-catenin expression. c-Myelocytomatosis oncogene expression was variable. This is the first study to describe the immunophenotype of islet cell tumors in the F344 rat and to show that islet cell tumors in the F344 rat exhibit similarities in protein expression to the human counterpart.
Keywords
Introduction
Islet cell tumors (ICTs) in rats are classified on the basis of tumor size, cellular pleomorphism, invasion, and metastatic behavior. Although the biological behavior of these tumors is difficult to predict histologically, most ICTs in rats are well differentiated, and metastasis is rare. According to the National Toxicology Program (NTP) historical control database, the incidence of spontaneously arising ICTs in F344/N rats is 8.55% and 2.01% for males and females, respectively (NTP 2009a). This is significantly greater than the incidence of ICTs in humans, which has been shown to be <1 per 100,000 worldwide (Halfdanarson, Rubin, et al. 2008).
In humans, ICTs (currently classified as pancreatic neuroendocrine tumors [PNETs]; Klimstra et al. 2010) are categorized on the basis of both functional and morphologic characteristics including hormone status, tumor diameter, cellular differentiation, mitotic index, local invasion, and metastasis (Ehehalt et al. 2009). These tumors may be sporadic or hereditary, and they are often well differentiated, making biologic behavior difficult to predict when metastatic disease is absent (La Rosa et al. 2007). An ICT may be composed of a homogenous population of one islet cell type, such as insulin-producing beta cells, or a mixture of islet cell types.
Although many human ICTs may express one or more peptide hormones immunohistochemically, unless clinical symptoms are present, human ICTs are considered nonfunctional (Halfdanarson, Rabe, et al. 2008). Clinical manifestations of ICTs in humans vary depending on the cellular origin of the tumor, and some tumors may produce more than one hormone (DeLellis and Shin 2006). Insulinomas are the most common functional ICT in humans, and 90% of these tumors are considered to be benign or low risk (Goldin, Aston, and Wahi 2008; Halfdanarson, Rubin, et al. 2008). Insulinomas are composed of β-cells and are associated with increased insulin levels and fasting hypoglycemia (Riley, Boorman, and Hayashi 1990; Schmitt et al. 2007; Stromberg, Wilson, and Capen 1983). Gastrinomas are the second most common functional ICT, and in contrast to insulinomas, these are most often malignant, with metastasis most common to the liver or lymph nodes (Goldin, Aston, and Wahi 2008; Halfdanarson, Rubin, et al. 2008; House and Schulick 2006). Gastrinomas are associated with overproduction of gastrin, a peptide that stimulates the production of gastric acid. Overproduction of gastrin is associated with a clinical syndrome called Zollinger-Ellison syndrome, characterized by gastric and duodenal ulcers and diarrhea (Gavaghan 2002). Glucagonomas, composed of α-cells, produce excess levels of glucagon and are associated with a syndrome of diabetes mellitus, a distinctive skin rash known as necrotizing cutaneous erythema, and anemia (Gavaghan 2002). Other functional ICTs (somatostatinomas, pancreatic polypeptide-secreting tumors, diarrheogenic ICTs [VIPomas], and pancreatic carcinoid tumors) are rare (Halfdanarson, Rubin, et al. 2008).
Seventeen chemicals tested by the NTP in 2-year carcinogenicity studies have been associated with a significant increase in proliferative lesions of the endocrine pancreas (islet cell hyperplasia, adenoma, or carcinoma) in F344 rats. However, the immunophenotype of these lesions has not been previously investigated, and the underlying pathways responsible for tumorigenesis in sporadic rat ICTs, and their relevance to human tumors, are unknown. In the current study, we have characterized these proliferative islet cell lesions on the basis of the expression of several hormones and mediators associated with biologic behavior and prognosis in human sporadic ICTs, and we discuss the potential mechanistic relevance to human disease.
Materials and Methods
Selection of Cases from the NTP Archives
The hematoxylin and eosin–stained tissue sections and the paraffin blocks of formalin-fixed and processed pancreatic tissue were obtained from the NTP 2-year bioassay studies of methylene blue trihydrate (MBT) and bromochloroacetic acid (BCAA) in male and female F344 rats (NTP 2008, 2009b). BCAA and MBT bioassays were selected for this study because they were the most recent NTP studies with a significant number of proliferative islet cell lesions (Table 1). A total of 34 proliferative pancreatic islet cell lesions from 32 rats were selected from the BCAA and MBT studies; 21 rats from the BCAA study (3 male and 3 female vehicle control, 12 male and 3 female treated) and 11 rats from the MBT study (2 male vehicle control and 9 male treated). From these two studies, there were 131 hyperplasias (71 in males and 60 in females), 63 adenomas (52 in males and 11 in females), and 9 carcinomas (8 in males and 1 in females). Of these proliferative islet cell lesions examined, 12 hyperplasias (7 from BCAA, 5 from MBT), 16 adenomas (8 each from BCAA and MBT), and 6 carcinomas (BCAA) were selected based on clear histopathologic criteria (Riley et al. 1990), lesion size, and location (within the parenchyma rather than edge of section) to ensure acquisition of the most appropriate samples for immunohistochemistry. The remainder of the lesions were excluded from evaluation either because of small size or lack of adequate tissue for immunohistochemistry and to keep the group sizes reasonably manageable. Two male rats from the MBT study were used for both islet cell hyperplasia and adenoma since both diagnoses were present in the same tissue section.
Incidences of proliferative islet cell lesions in MBT and BCAA 2-year bioassays in F344 rats. a
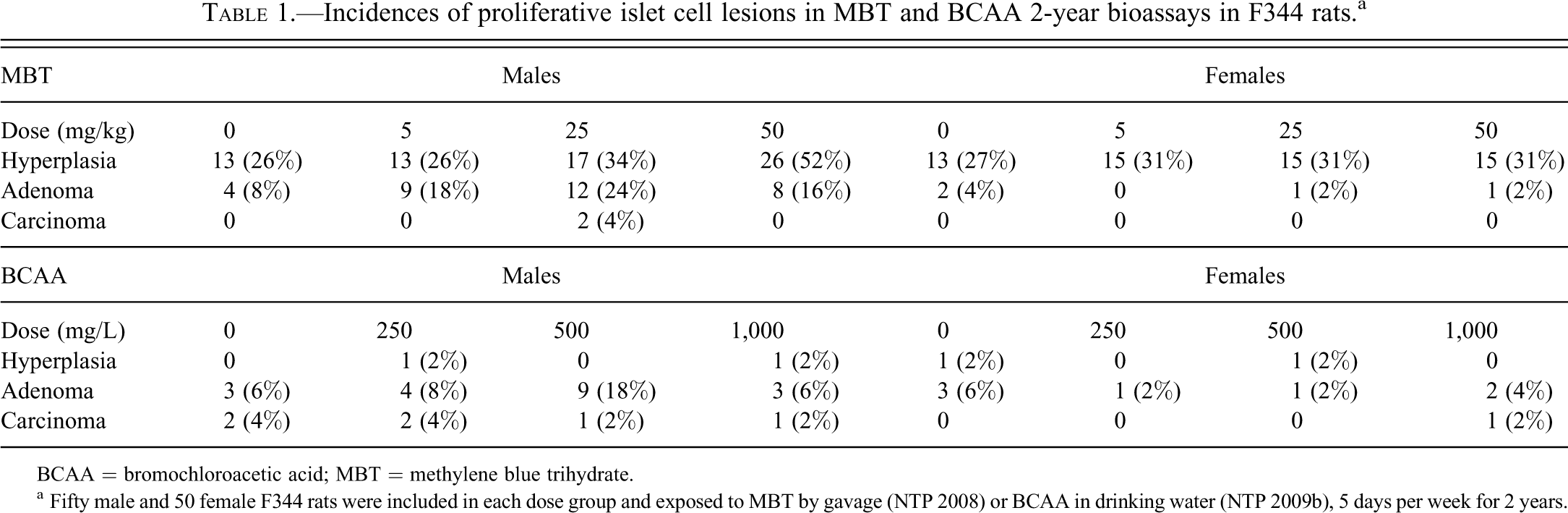
BCAA = bromochloroacetic acid; MBT = methylene blue trihydrate.
a Fifty male and 50 female F344 rats were included in each dose group and exposed to MBT by gavage (NTP 2008) or BCAA in drinking water (NTP 2009b), 5 days per week for 2 years.
Immunohistochemistry
Five-micrometer-thick sections were cut from each paraffin block and placed onto positively-charged glass slides, deparaffinized in xylene, and rehydrated through graded alcohols. To characterize the hormone status of hyperplastic and neoplastic islet cell lesions, we evaluated the expression of the most common peptides produced by functional ICTs in humans: synaptophysin, insulin, gastrin, glucagon, and somatostatin. To evaluate cell proliferation in hyperplasias, adenomas, and carcinomas, we measured protein expression of an important cell-cycle mediator, cyclin-dependent kinase 4 (CDK4), and a protein important in DNA synthesis, proliferating cell nuclear antigen (PCNA). c-myelocytomatosis oncogene (CMYC) and β-catenin, two proteins involved in cellular proliferation and oncogenesis, were also evaluated based on their overexpression in human PNETs and other types of human cancer (Wang, Johnston, and Buchanan 1997). The antibodies were prepared in 1% bovine serum albumin in tris-buffered saline containing Tween 20, pH 7.6 (±0.1), and the concentrations used and manufacturers are as follows: synaptophysin (rabbit polyclonal anti-human antibody, 1:200, Thermo Fisher Scientific, Fremont, CA: RB-1461), insulin (mouse monoclonal anti-human antibody, 1:8,000, Sigma-Aldrich, St. Louis, MO: 12018), glucagon (rabbit polyclonal anti-human antibody, 1:3,000, Dakocytomation, Carpinteria, CA: A0565), somatostatin (rabbit polyclonal anti-human antibody, 1:3,000, Dakocytomation: A0566), gastrin (rabbit polyclonal anti-human antibody, 1:400, Dakocytomation: A0568), CDK4 (rabbit polyclonal anti-mouse antibody, 1:500, Santa Cruz Biotechnology, Inc., Santa Cruz, CA: sc-260), CMYC (rabbit polyclonal anti-human antibody, 1:1,000, Abcam Inc., Cambridge, MA: ab39688), β-catenin (goat polyclonal anti-human antibody, 1:50, Santa Cruz Biotechnology: sc-7199), and PCNA (rabbit polyclonal anti-human antibody, 1:300, Santa Cruz Biotechnology: sc-7907). The β-catenin antibody was detected using the Discovery XT Automated System with the OmniMap Anti-Rabbit Polymer Detection Kit (Ventana Medical Systems, Tucson, AZ). All the other stains were performed manually using the avidin-biotin complex technique. Endogenous peroxidases were blocked by 3% hydrogen peroxide for 15 min. Antigen retrieval was performed using 1X citrate buffer in a Decloaking Chamber (Biocare Medical, Walnut Creek, CA) for 5 min at 120°C. Following washing, the labeled avidin-biotinylated enzyme complex technique was used for amplification of primary antibody binding. See Supplemental Table 1 for details on antibody detection and rationale. Antibody-antigen complexes were visualized using 3,3′-diaminobenzidine chromagen (Dakocytomation) for 6 min at room temperature. Tissues were counterstained with Harris hematoxylin, then dehydrated through graded alcohol, soaked in xylene, and coverslipped with MM24 Mounting Media (Leica Microsystems, Bannockburn, IL). Normalized serum from the host species in which each primary antibody was made was used for negative controls. Tissues normally expressing each protein were used as positive controls (pancreatic islets for synaptophysin, insulin, glucagon, and somatostatin; stomach for gastrin; and small intestinal epithelium for CMYC, CDK4, PCNA, and β-catenin). Each section typically contained normal islets that served as an internal control.
Immunohistochemical Grading and Statistical Analysis
Insulin and synaptophysin expression was evaluated semi-quantitatively based on the overall intensity of cytoplasmic immunoreactivity (1 = weak or no staining, 2 = mild staining, 3 = moderate staining, 4 = marked staining). The expression of β-catenin was evaluated on the basis of overall intensity of membrane and cytoplasmic immunoreactivity (absent = 0, mild = 1, moderate = 2, marked = 3). PCNA scoring was based on the product of the percentage of positive nuclei within each lesion (<50% = 1, 50–74% = 2, 75–90% = 3, >90% = 4) and the intensity of nuclear staining (1 = weak or no staining, 2 = mild staining, 3 = moderate staining, 4 = marked staining). CDK4 scoring was based on the product of the intensity (absent = 0, mild = 1, moderate = 2, marked = 3) of nuclear immunoreactivity and the proportion of positive nuclei (<25% = 1, 25-49% = 2, 50–75% = 3, >75% = 4). CMYC scoring was not pursued since there was no significant difference in expression between normal islets and islet lesions. The expression of glucagon, somatostatin, and gastrin was not graded semi-quantitatively; rather, the approximate proportion of positive cells was noted for each antibody and compared with normal islets, since the intent was only to document hormone status rather than to measure degree of expression. Immunohistochemical staining was evaluated independently by two individuals in a coded (blinded) manner (C.K., M.J.H.), and the trends in immunoreactivity were consistent between each observer. All discrepancies between observers were minimal (within one grade) and infrequent and were resolved by consensus between the two reviewers in a second read. The results for each antibody except glucagon, somatostatin, and gastrin were compared statistically using Kruskal-Wallis analysis of variance due to nonnormality of the scores. Dunn’s test was used to compare scores for each type of proliferative lesion (hyperplasias, adenomas, and carcinomas) to normal islets.
Results
Histopathology
Lesions were diagnosed as within normal limits, hyperplasia, adenoma, or carcinoma based on current histopathologic criteria (Riley, Boorman, and Hayashi 1990; Figure 1A–D). Hyperplasia was diagnosed when islets were enlarged, with lobulated or irregular borders, containing increased numbers of cytologically normal to slightly hypertrophied islet cells (Figure 1B). Adenomas were characterized by well-circumscribed, expansile nodules composed of solid clusters, cords, or ribbons of cells with absent to minimal cellular atypia, which compressed adjacent exocrine pancreatic parenchyma and occasionally contained entrapped exocrine acini (Figure 1C). Carcinomas were diagnosed when one of the following was present: (1) significant cellular atypia and pleomorphism and a heterogeneous growth pattern or (2) local invasion into the surrounding exocrine pancreatic parenchyma or into the peripancreatic adipose tissue (Figure 1D).
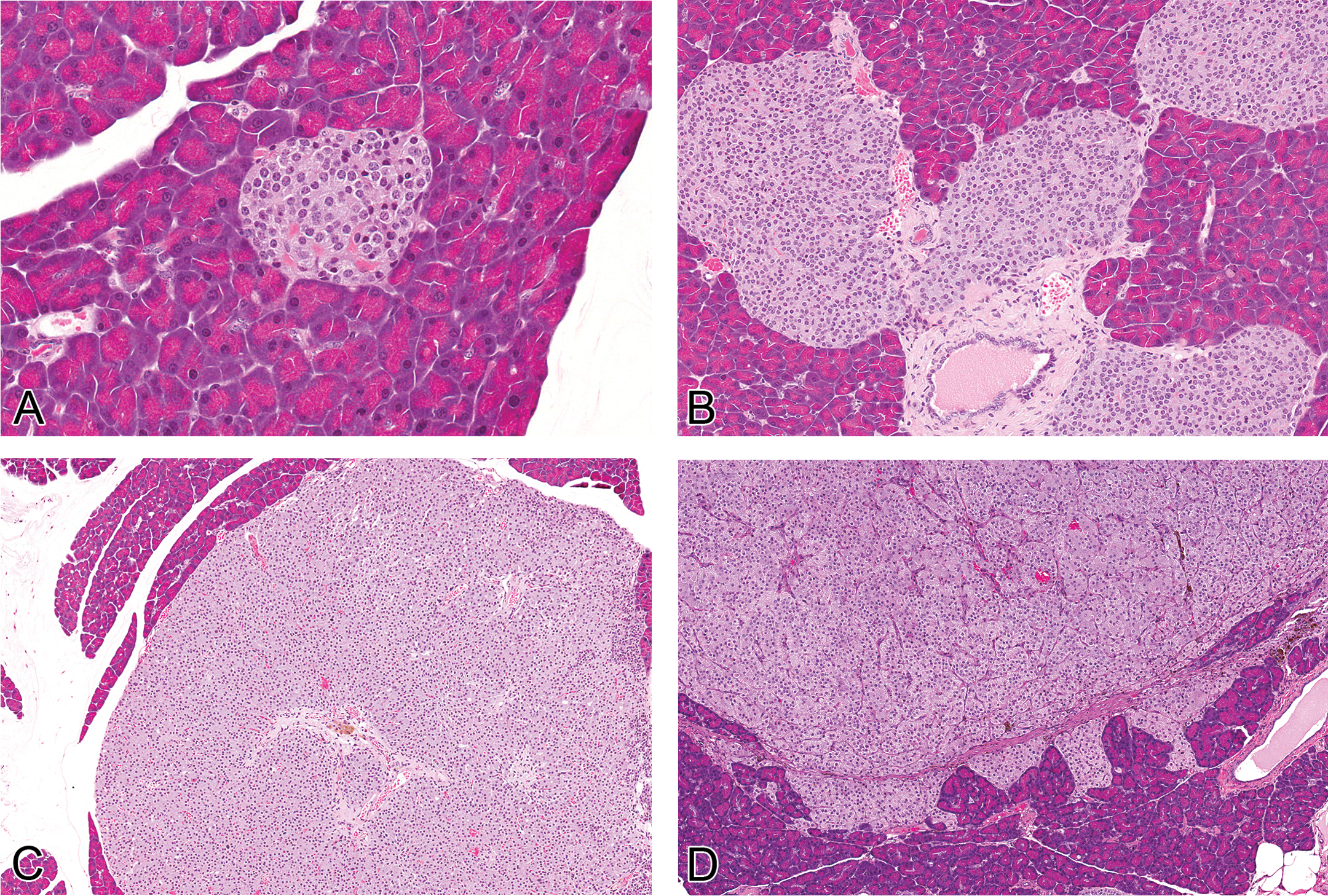
Histopathology of normal islets and proliferative islet cell lesions. (A, 40×) Normal islets are variable in size, with rounded, well-defined borders, composed of nests of well-differentiated neuroendocrine cells. (B, 20×) Hyperplastic islets are increased in size, with irregular or lobulated borders, and composed of nests of well-differentiated to somewhat hypertrophied neuroendocrine cells. (C, 10×) Adenomas are characterized by an expansile, well-circumscribed, unencapsulated proliferation of solid clusters, cords, or ribbons of fairly well-differentiated neuroendocrine cells, and carcinomas (D, 10×) exhibit increased cellular pleomorphism, anaplasia, and local invasion.
Protein expression of synaptophysin and insulin decreases in islet cell lesions as differentiation decreases and proliferation increases
Synaptophysin was diffusely expressed within the cytoplasm of normal islets (Figure 2A) but was significantly decreased in hyperplasias (Figure 2B) and adenomas (Figure 2C; p < 0.05 by Dunn’s test). Carcinomas (Figure 2D) appeared to express similar amounts of synaptophysin as normal islets (Figure 3).
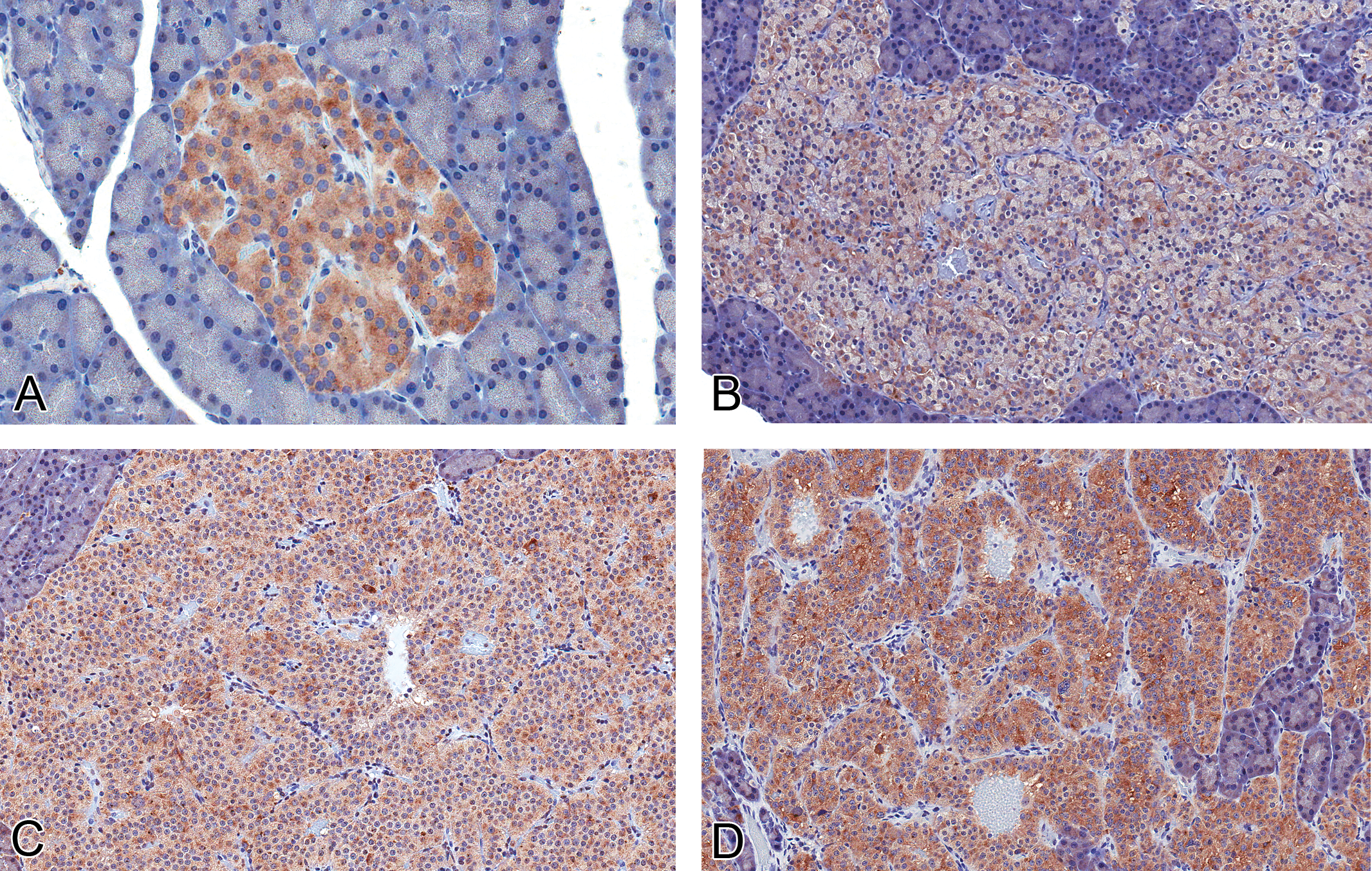
Expression of synaptophysin in normal islets and proliferative islet cell lesions. (A) Normal islets express synaptophysin ubiquitously within the cytoplasm of all islet cells (40×), whereas intensity is decreased in hyperplastic islets (B, 20×) and adenomas (C, 20×). There was similar expression in the carcinomas (D, 20×) in this study compared with normal islets (rabbit polyclonal anti-human synaptophysin antibody).
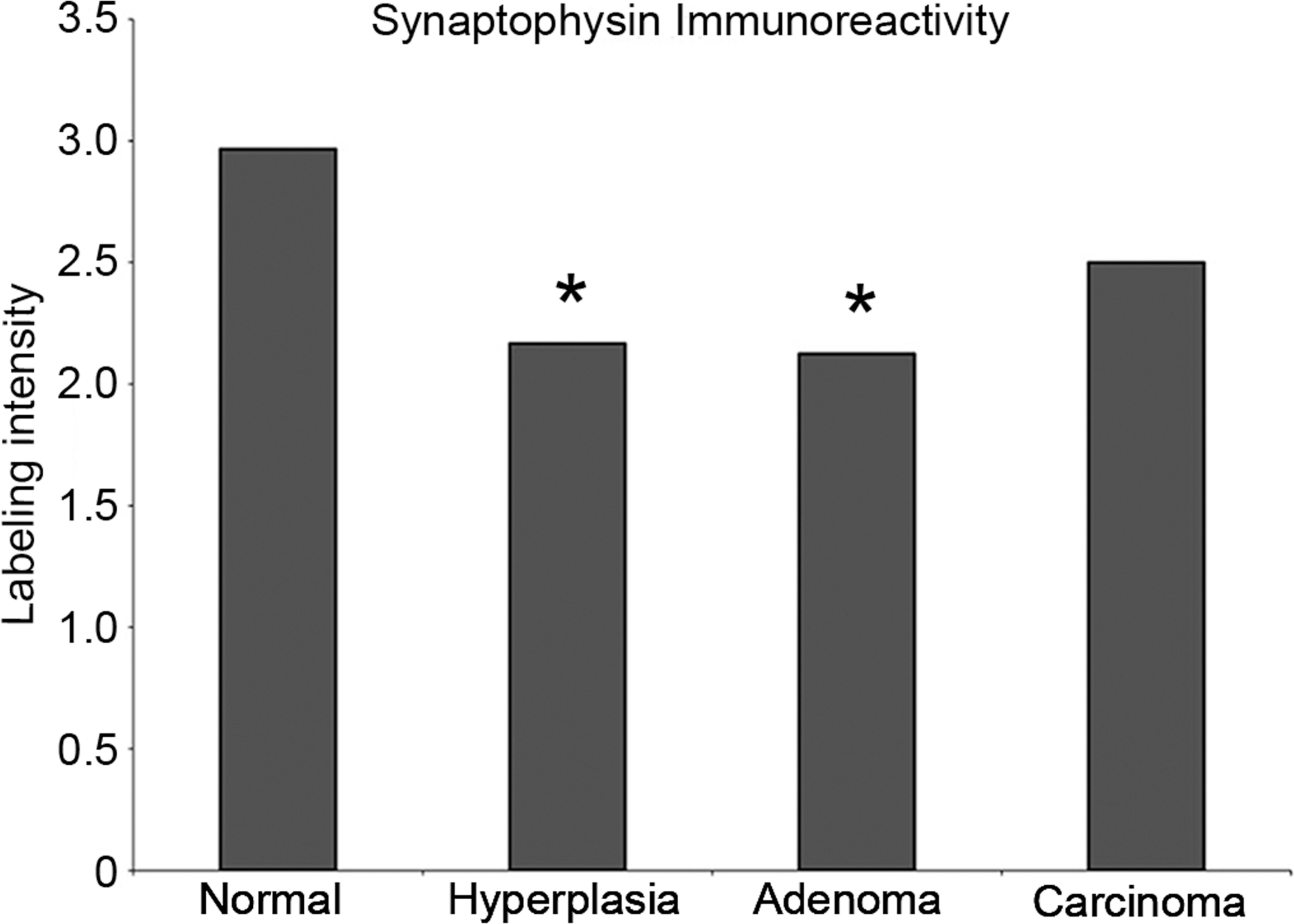
Synaptophysin protein quantification in normal islets and proliferative islet lesions. There is a stastistically signficant decrease in synaptophysin expression in hyperplastic islets (*p < 0.05) and adenomas (**p < 0.01).
In normal islets, insulin expression was present within the cytoplasm of islet cells in a central core surrounded by one to two cell layers of insulin-negative islet cells, consistent with the normal distribution of β-cells in rats (Figure 4A). Islet cell hyperplasias (Figure 4B) and tumors (Figure 4C, D) lost this normal peripheral distribution of insulin-positive cells, such that staining became more multifocal to diffuse within proliferative lesions. However, as lesions progressed toward malignancy (hyperplasia > adenoma > carcinoma), insulin expression decreased, with statistically significant decreases in adenomas (p < 0.05) and in carcinomas (p < 0.01; Figure 5).
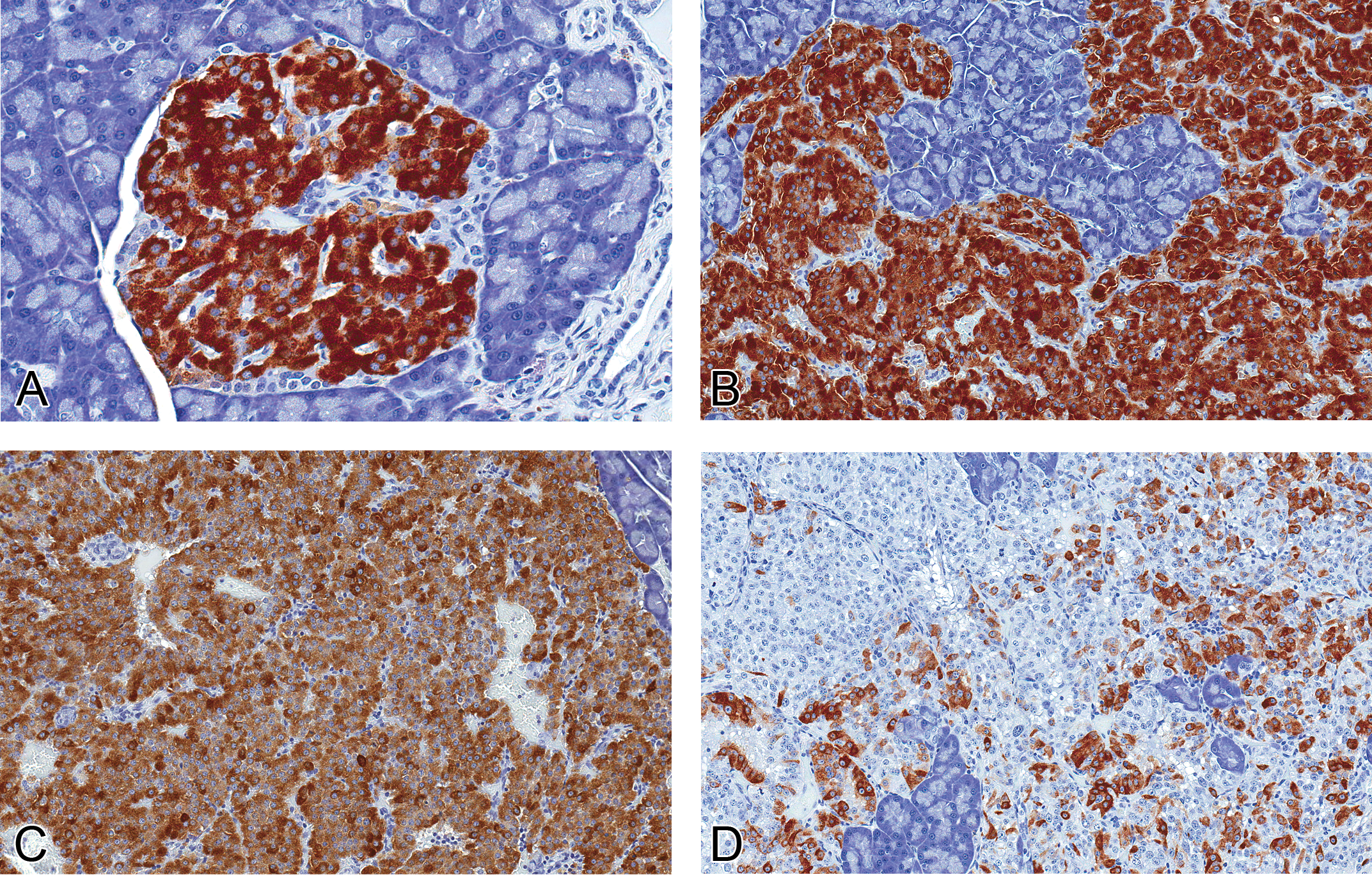
Protein expression of insulin in normal islets and proliferative islet lesions. (A, 40×) Normal and hyperplastic (B, 20×) islets show intense cytoplasmic insulin immunoreactivity. There is decreased insulin expression in islet cell adenomas (C, 20×), and in carcinomas (D, 20×), only sporadic insulin immunoreactivity is present (mouse monoclonal anti-human insulin antibody).
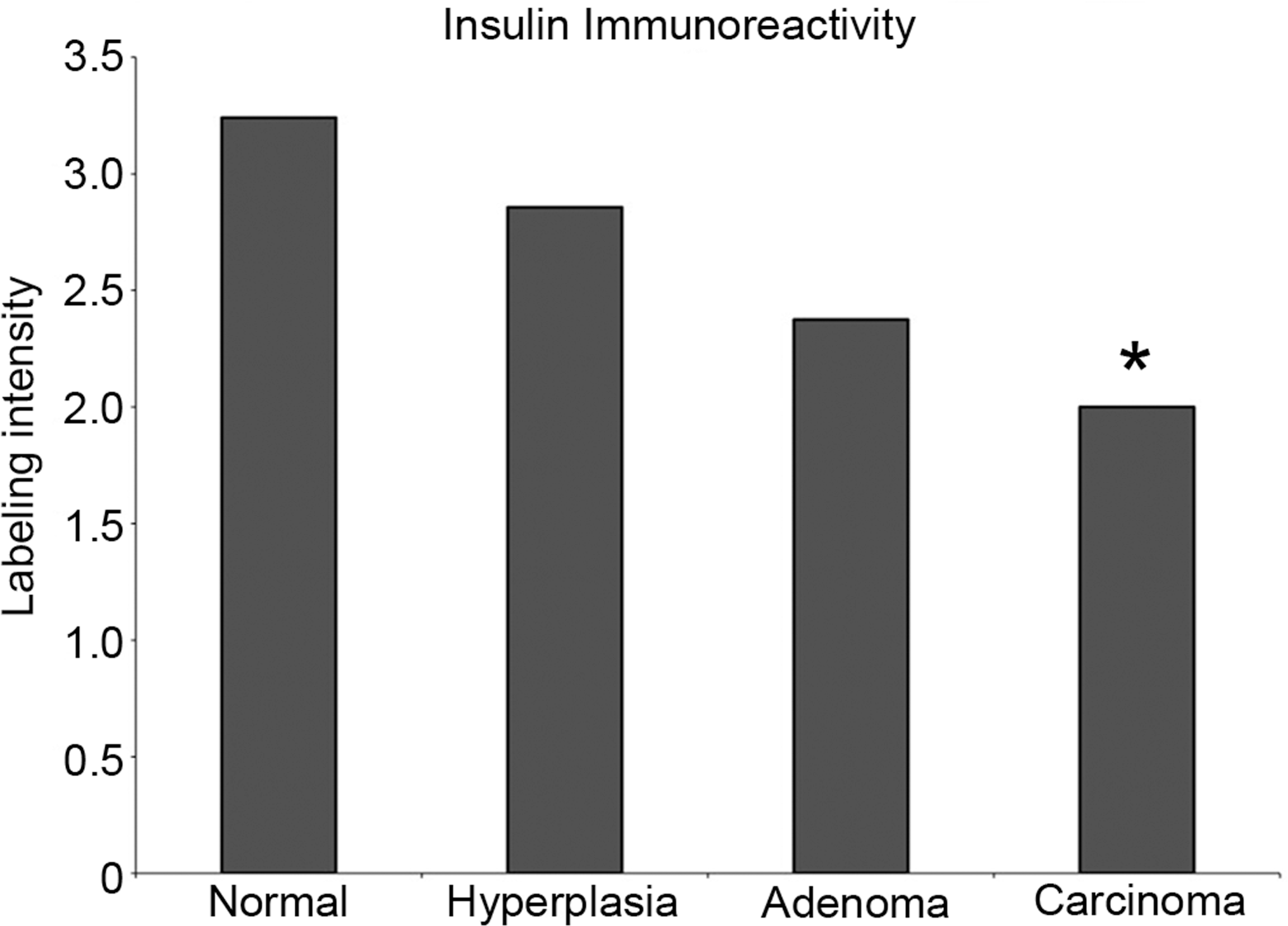
Insulin protein quantification in normal islets and proliferative islet cell lesions. There is a decreasing trend in insulin labeling intensity as lesions progress toward malignancy, with a statistically signficant decrease in adenomas (*p < 0.05) and carcinomas (**p < 0.01).
The expression of the other peptide hormones, glucagon (Figure 6), somatostatin (Figure 7), and gastrin (Figure 8), was variable in islet cell lesions. In normal islets, glucagon and somatostatin expression was confined to small numbers of cells within the periphery, and gastrin expression was absent, except for rare, peripherally located cells. In hyperplasias (four evaluated) and tumors (nine adenomas and five carcinomas evaluated), the distribution of positive cells for glucagon, somatostatin, and gastrin was variable and no longer confined to the periphery of the lesions. In a majority of lesions, less than 5% of cells were immunopositive for any of the non-insulin hormones; however, occasional hyperplasias expressed glucagon in up to 20% of cells, and a single hyperplastic islet expressed glucagon in approximately 75% of cells. Significant (50–95% of cells) expression of somatostatin was observed in two adenomas, and moderate expression (20–30%) was present in one adenoma and one carcinoma. Gastrin expression was minimal (1–5%) to absent in all islet cell lesions but exceeded that seen in normal islets.
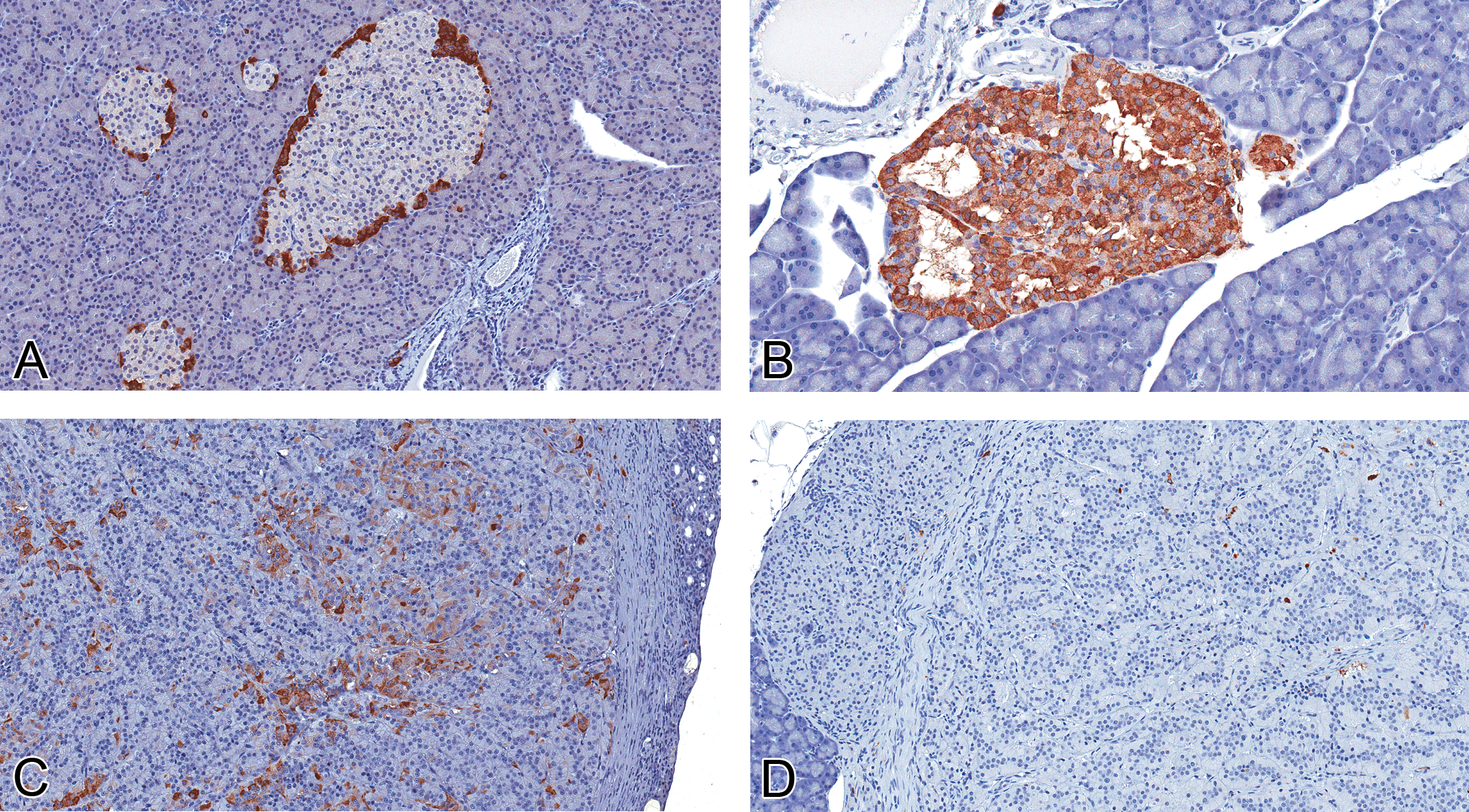
Expression of glucagon in normal islets and proliferative islet cell lesions. (A, 20×) Glucagon expression is limited to the peripheral cells of normal islets. In some proliferative islet lesions, glucagon was no longer confined to the periphery and was variably expressed, from diffusely increased in a hyperplastic lesion (B, 40×), to multifocally expressed in an adenoma (C, 20×). In most islet cell carcinomas, the expression of glucagon was minimal to absent (D, 20×; rabbit polyclonal anti-human glucagon antibody).
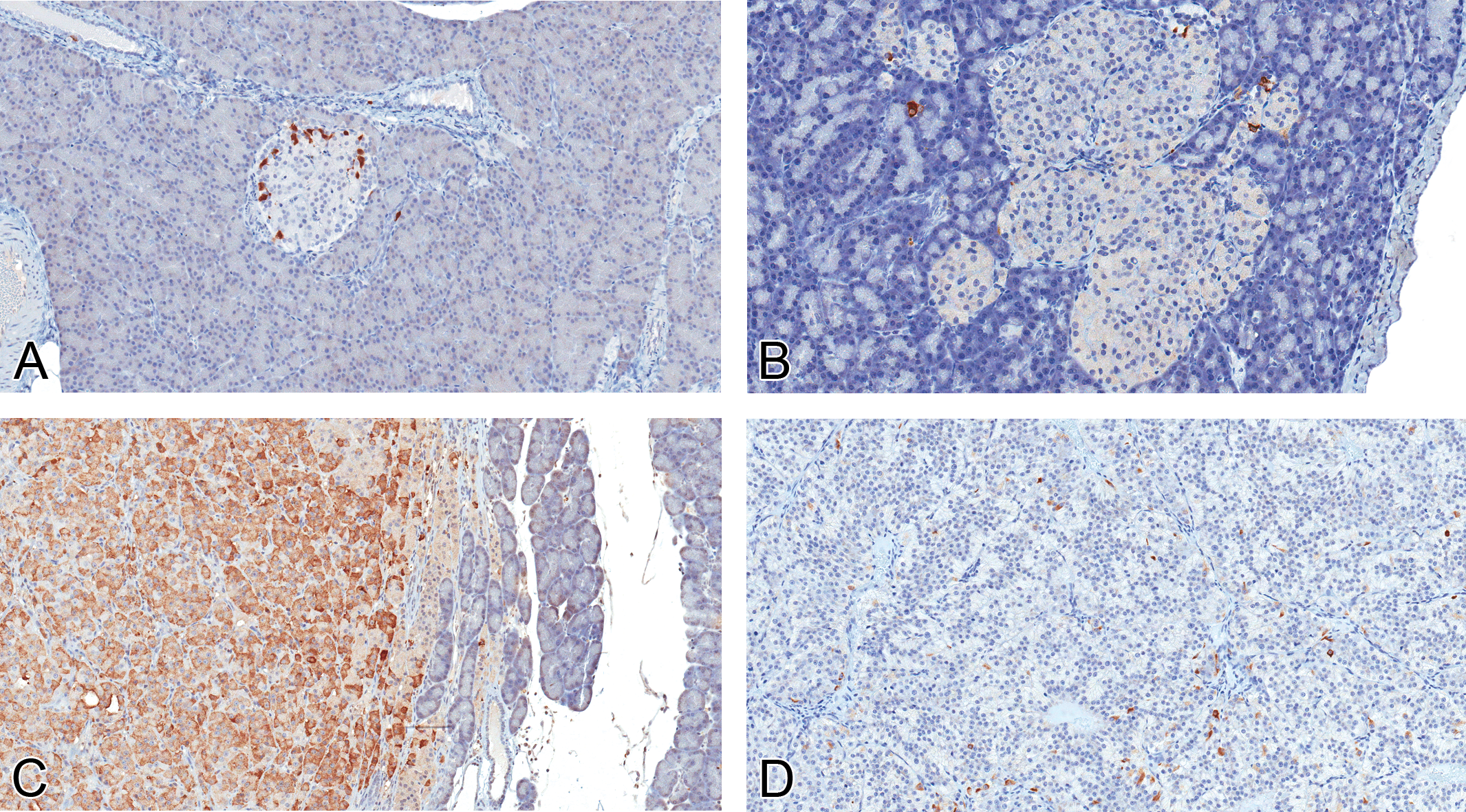
Expression of somatostatin in normal islets and proliferative islet cell lesions. (A, 20×) Somatostatin expression was limited to peripheral cells in normal islets. In proliferative islet lesions including hyperplasias (B, 20×), adenomas, and carcinomas (D, 20×), somatostatin expression was generally minimal. However, in two adenomas (C, 20×), expression of somatostatin was widespread (rabbit polyclonal anti-human somatostatin antibody).
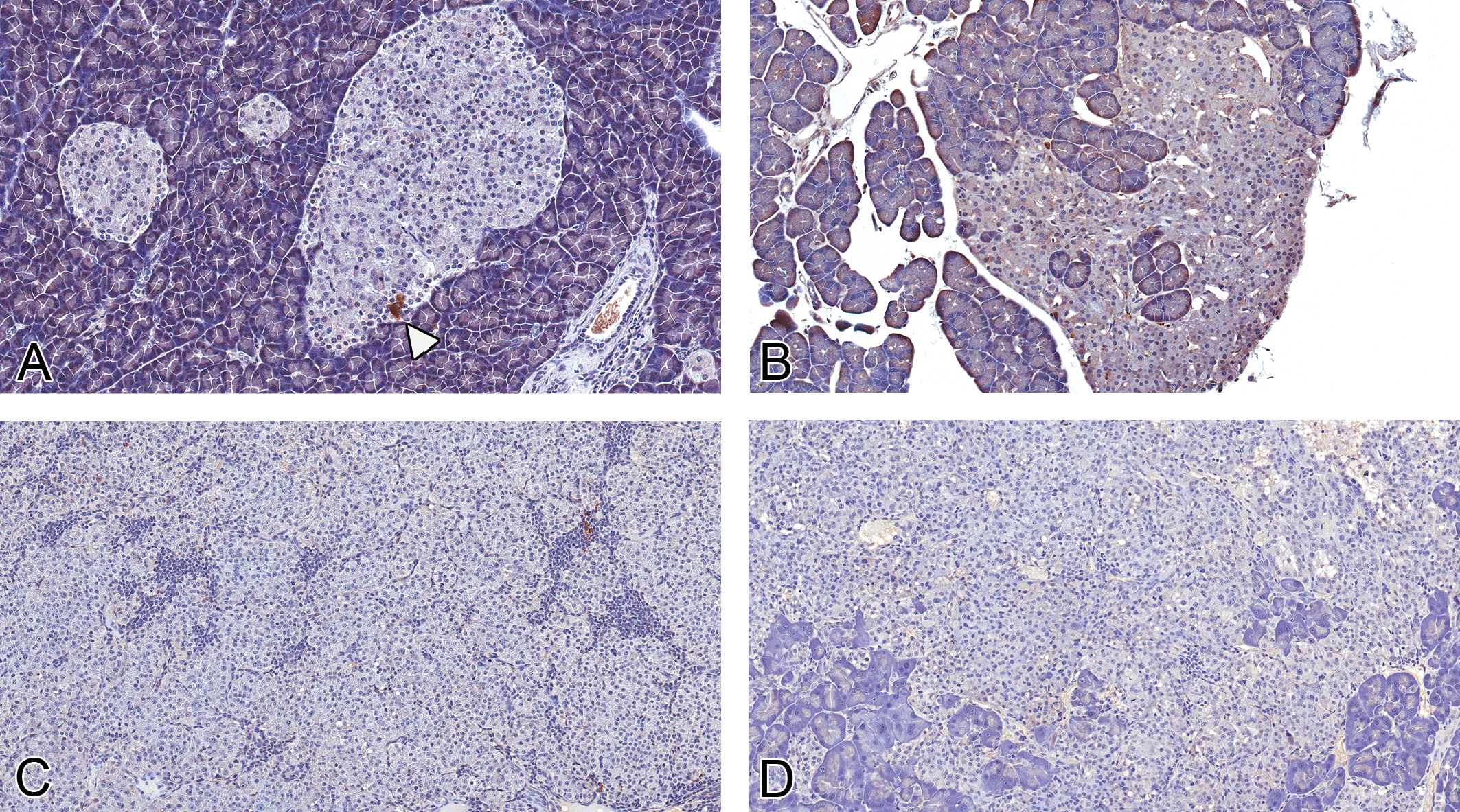
Expression of gastrin in normal islets and proliferative islet cell lesions. (A, 20×) Gastrin expression in normal islets was limited to rare periperally located cells (white arrowhead). In islet cell hyperplasias (B, 20×), adenomas (C, 20×), and carcinomas (D, 20×), the expression of gastrin was minimal to absent (rabbit polyclonal anti-human gastrin antibody).
Proliferative islet cell lesions are associated with increased expression of cell-cycle mediators CDK4 and PCNA
Cytoplasmic CDK4 expression was noted in both normal islet cells and ICTs, with increased nuclear expression in islet cell adenomas and carcinomas (Figure 9). Since nuclear expression of CDK4 is associated with cell-cycle progression (Kim and Diehl 2009), grading of CDK4 expression was based on the proportion of cells with nuclear immunoreactivity and the relative intensity of nuclear expression. There was a statistically significant increase in nuclear CDK4 expression in islet cell adenomas (p < 0.001) and carcinomas (p < 0.05) compared with normal and hyperplastic islets (Figure 10).
There was minimal to mild nuclear PCNA immunoreactivity in approximately 50% of cells within normal islets, and the proportion and intensity of positively labeled cells increased as lesions progressed toward malignancy (normal < hyperplasia and adenoma < carcinoma; Figure 11), with a statistically significant increase in carcinomas (p < 0.01) compared with normal islets (Figure 12).
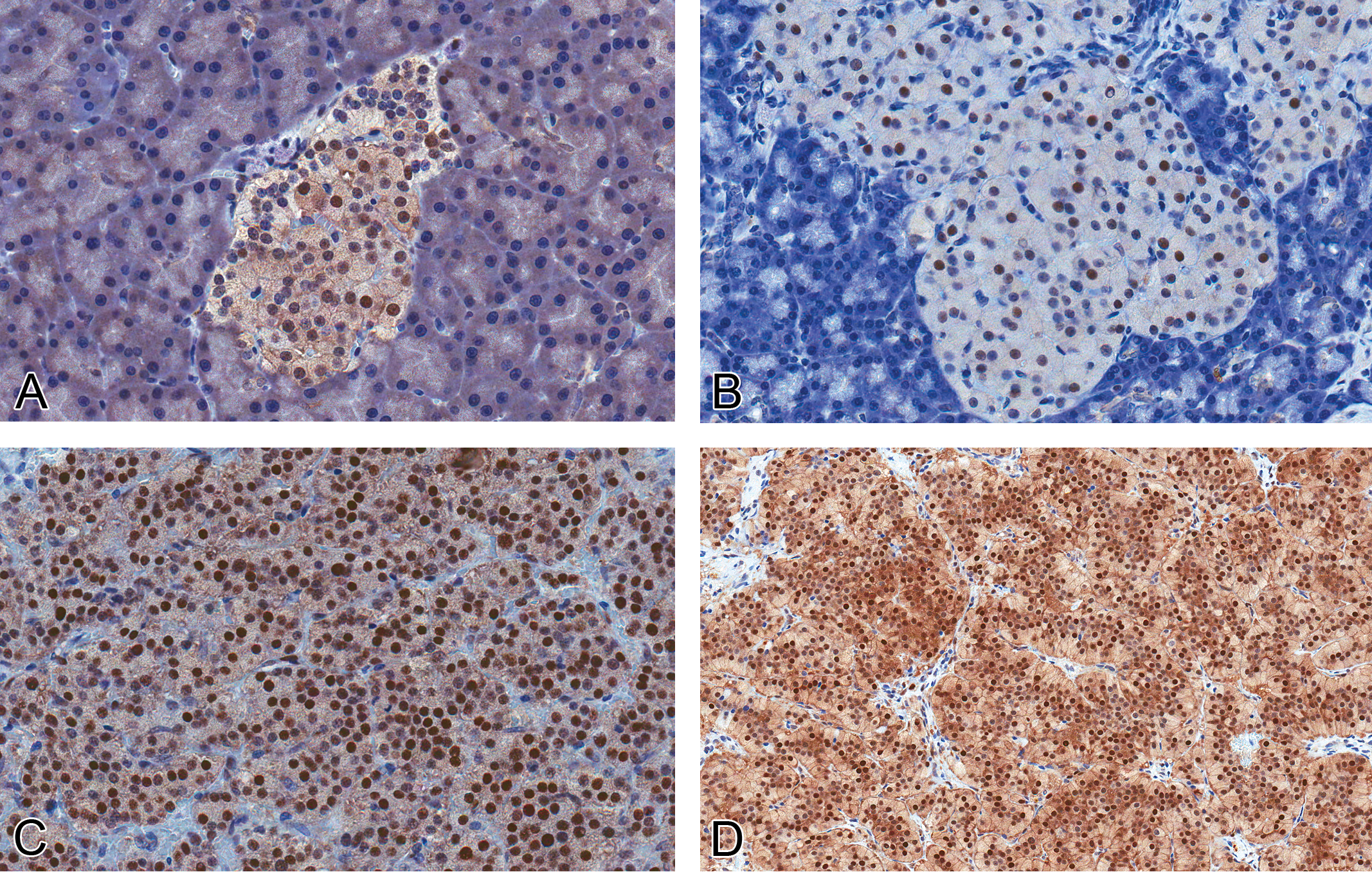
Expression of cyclin-dependent kinase 4 (CDK4) protein in normal islets and proliferative islet cell lesions. (A, 40×) Normal islets and hyperplasias (B, 40×) show significantly fewer and less intensely immunolabeled nuclei compared with islet cell adenomas (C, 40×) and carcinomas (D, 20×; rabbit polyclonal anti-mouse CDK4 antibody).
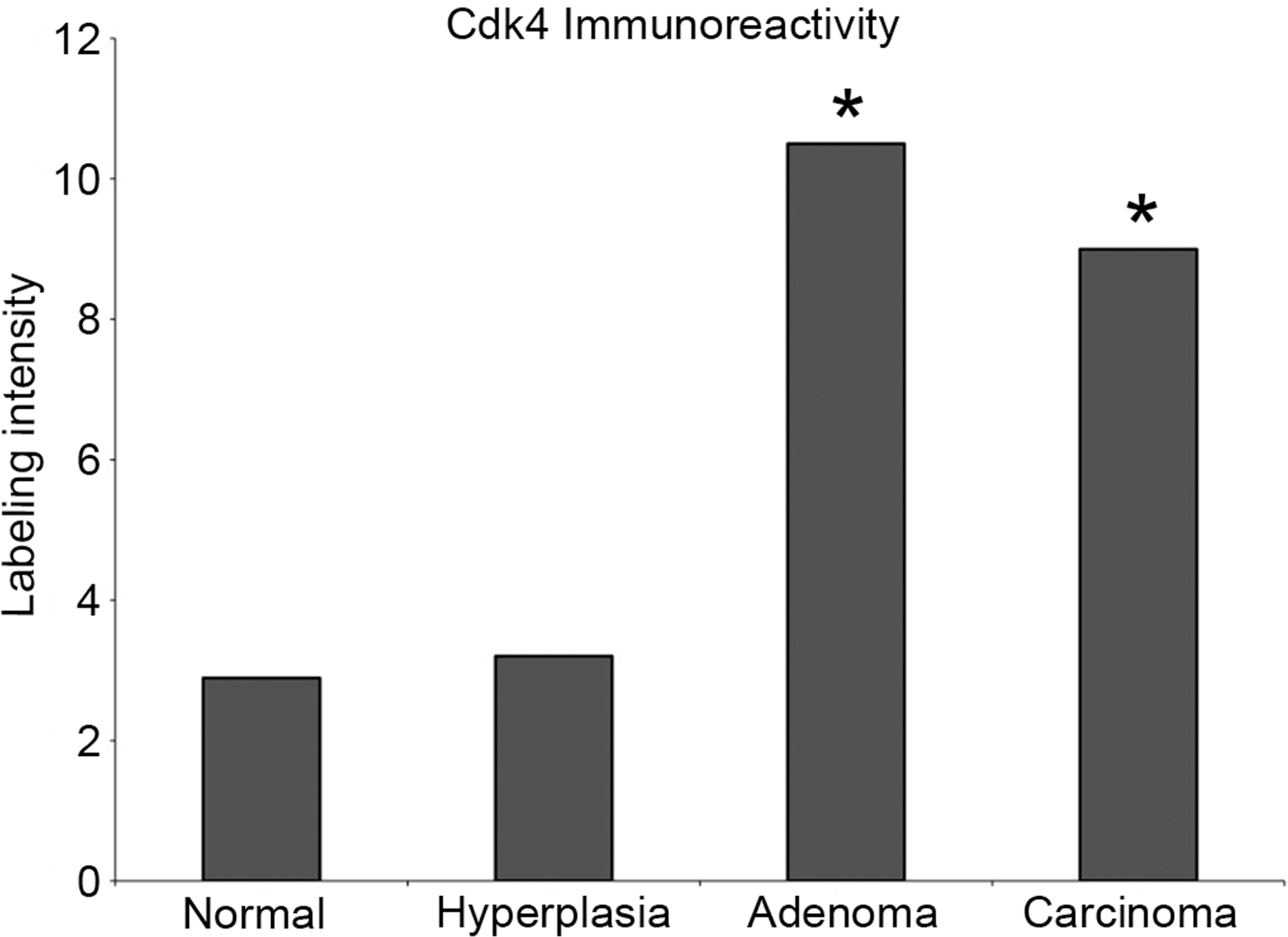
Protein quantification of cyclin-dependent kinase 4 (CDK4) in normal islets and proliferative islet cell lesions. Semi-quantitative analysis demonstrates a stastistically significant increase in number and intensity of CDK4 immunolabeled islet cells in adenomas (***p < 0.001) and carcinomas (*p < 0.05).
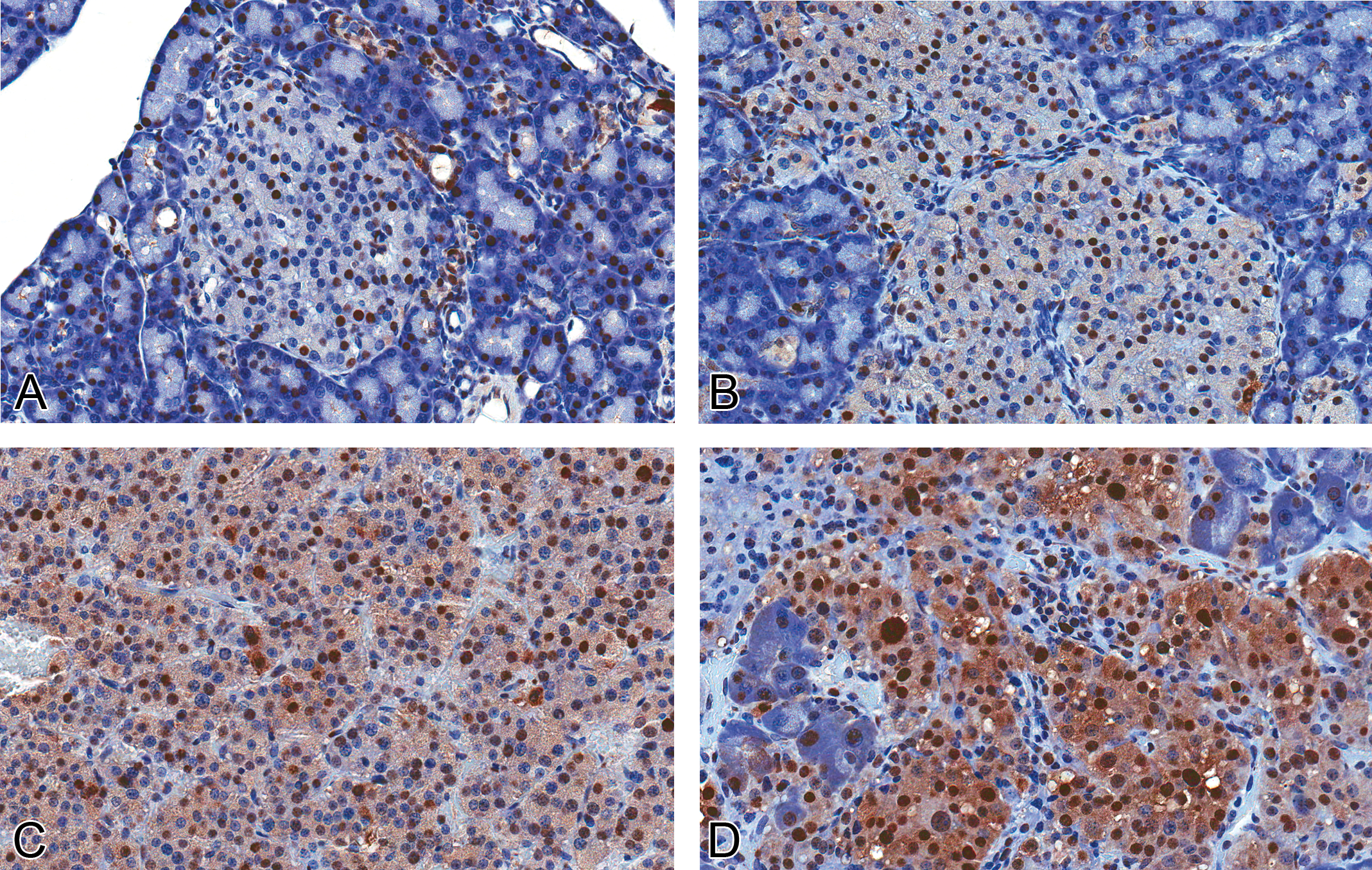
Expression of proliferating cell nuclear antigen (PCNA) protein in normal islets and proliferative islet cell lesions. (A, 40×) There is scattered nuclear PCNA immunolabeling in normal islets, with increased intensity and number of labeled cells in hyperplastic islets (B, 40×) and adenomas (C, 40×). Carcinomas (D, 40×) have a further increase in number and intensity of positively labeled nuclei compared with adenomas (rabbit polycloncal anti-human PCNA antibody).
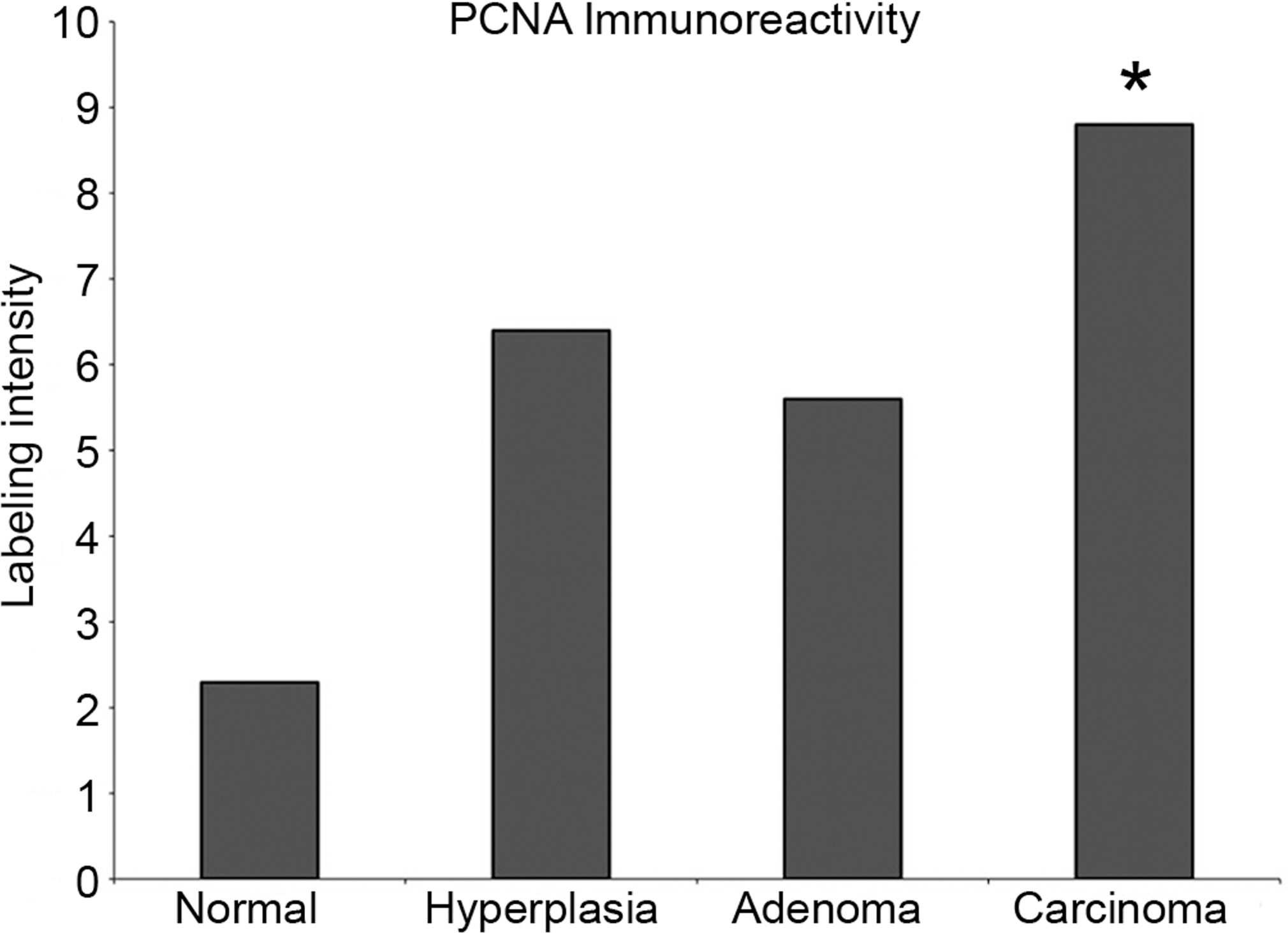
Protein quantification of proliferating cell nuclear antigen (PCNA) expression in normal islets and proliferative islet lesions. Semi-quantitative analysis demonstrates increased expression of PCNA in proliferative islet cell lesions, with a statistically signficant increase in carcinomas (**p < 0.01).
Proliferative islet cell lesions are associated with decreased expression of β-catenin protein expression
In normal islets, there was moderate cytoplasmic and membrane immunoreactivity to β-catenin, with a statistically significant decrease in expression in adenomas (p < 0.001) compared with normal islets (Figures 13 and 14). Nuclear translocation of β-catenin, which is often associated with mutation of the gene and transcription of downstream cell-cycle mediators, was not observed. CMYC protein was variably expressed in the nuclei and cytoplasm of all samples, and there was no significant difference in CMYC localization or intensity between normal islets and proliferative islet cell lesions (data not shown).
Discussion
Molecular characterization of a rodent model is critical in validating its relevance to disease in humans. In this study, we have shown similarities in the protein expression of several hormones and mediators of ICTs in rats common to that of humans. Our results are consistent with a number of previous studies in the human literature, including synaptophysin and insulin expression in normal and neoplastic islet cells (Chejfec et al. 1987; Wiedenmann et al. 1986), decreased insulin expression in islet cell carcinomas (Lam and Lo 1997), increased expression of cell-cycle proteins CDK4 and PCNA (Lindberg et al. 2007; Wang et al. 1997), and lack of nuclear expression of β-catenin (Burford et al. 2009; Comper et al. 2009; Hervieu et al. 2006; Liu et al. 2009). These findings illustrate that there are histologic and molecular features in rat lesions similar to the human disease, which may have bearing on human hazard identification in terms of rat toxicology and carcinogenicity studies in which there is an islet cell tumor response to chemical treatment.
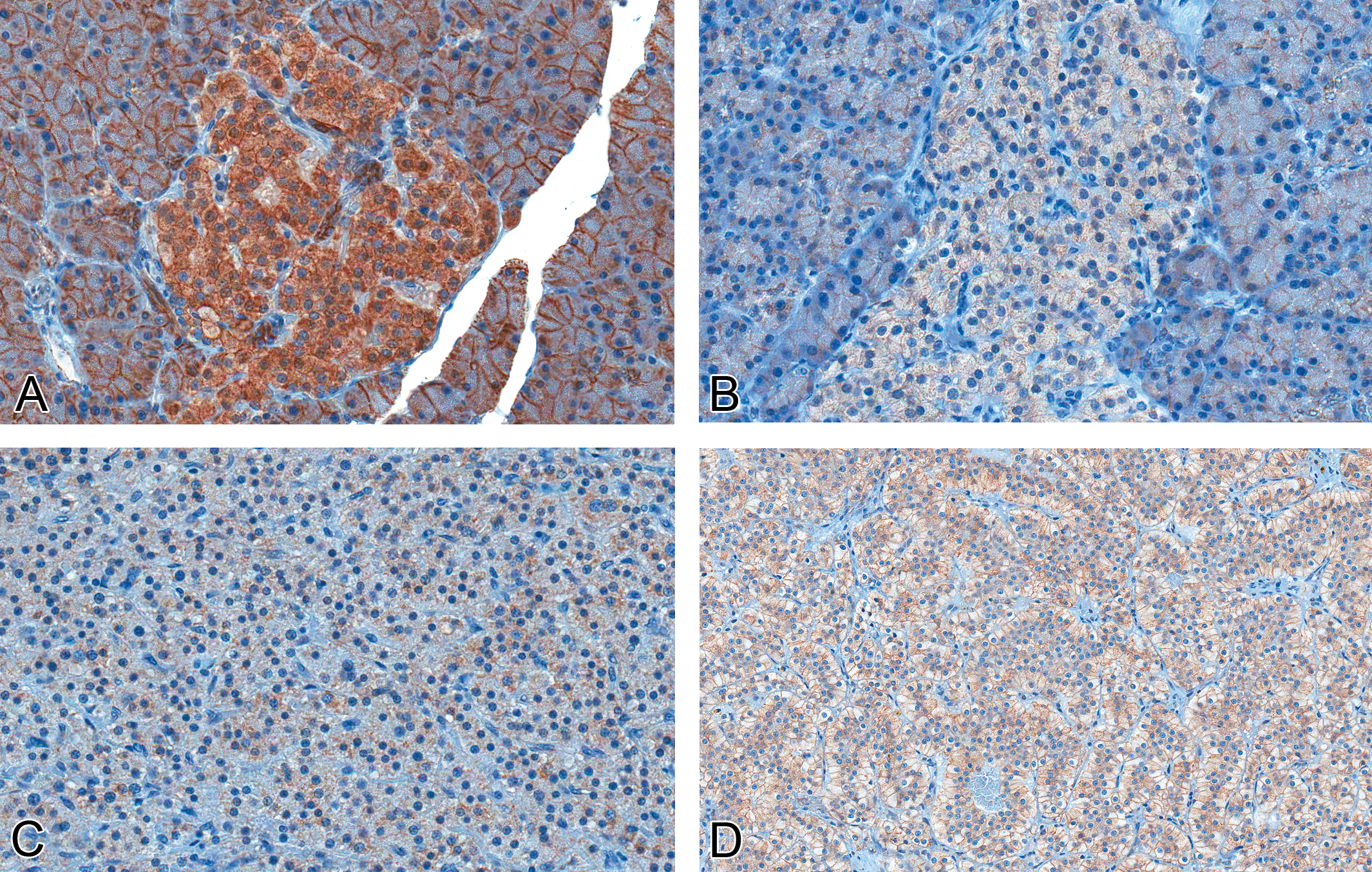
Expression of β-catenin protein in normal islets and islet cell lesions. (A, 40×) There is intense membranous and cytoplasmic expression in normal islets, with significantly decreased expression in hyperplastic islets (B, 40×), adenomas (C, 40×), and carcinomas (D, 20×; polyclonal goat anti-human β-catenin antibody).
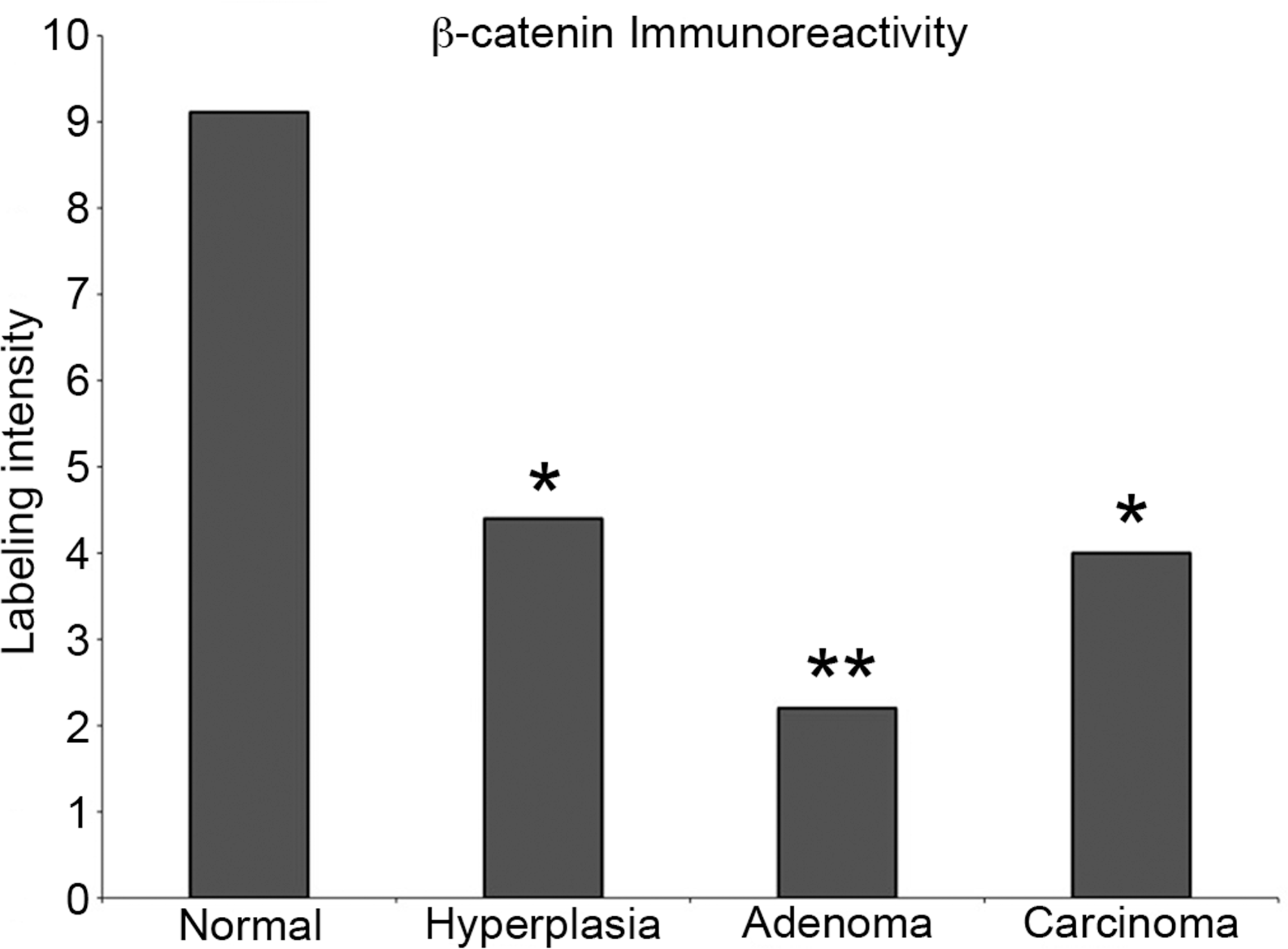
Protein quantification of β-catenin in normal islets and proliferative islet cell lesions. Semi-quantitative analysis of normal and proliferating islets demonstrates a statistically significant decrease in expression of β-catenin in adenomas (***p < 0.001).
The histomorphologic characterization of islet cell lesions in the F344 rat is well defined (Riley, Boorman, and Hayashi 1990), but the underlying changes in peptide hormone distribution and the molecular changes leading to transformation are unknown. As a general marker of neuroendocrine cells, synaptophysin is often used in the diagnosis of human pancreatic neoplasms (Chejfec et al. 1987; Wiedenmann et al. 1986). As expected, all rat islet cell lesions in our study were positive for synaptophysin. Interestingly, there was a decreasing trend in proliferative islet cell lesions, although only statistically significant in hyperplasias and adenomas. The reason for this decreased expression is unknown; however, as cells progress to malignancy and a more poorly differentiated state, they may lose expression of normal cellular proteins, or these proteins may be altered, leading to a change in their antigenicity. However, if this were the case, one would expect a further statistically significant decrease in synaptophysin in islet cell carcinomas; this may be due to small sample size and biologic variability in protein expression between tumors. The inclusion of additional samples of this uncommon tumor may reveal a statistically significant decrease in synaptophysin in carcinomas compared with normal islets.
Similar to islet cell neoplasia in humans (Goldin, Aston, and Wahi 2008; Halfdanarson, Rubin, et al. 2008), a majority of the ICTs in our study (17 of 22 tumors evaluated for insulin and other peptide hormones) were characterized as insulinomas. Variable expression of other islet cell hormones such as glucagon and somatostatin was also noted in some of these tumors. While all proliferative islet lesions in the F344 rat expressed insulin, expression of insulin decreased as these lesions progressed toward malignancy. This may be due to loss of differentiation of the β-cell lineage or loss of insulin expression due to β-cell dedifferentiation. Furthermore, in some human ICTs, peptide hormones may be produced in an abnormal form and thus may not be detectable with a specific antibody, or they may be synthesized and released at such a rapid rate that the concentration of the protein in the tumor is too low to be demonstrated by immunohistochemstry (Portela-Gomes, Hacker, and Weitgasser 2004). Alternatively, mixed ICTs in humans have long been reported (Larsson 1978), with approximately 50% of insulinomas containing cells that are positive for other hormones including glucagon, somatostatin, and gastrin (DeLellis and Shin 2006). Consistent with this, while gastrin expression remained relatively minimal in tumors (1–5%), a few tumors in this study expressed glucagon and somatostatin to fairly moderate to marked levels (10–20% for glucagon in 2 of 14 tumors [14%], 20–95% in 4 of 14 tumors [29%] for somatostatin), possibly representing a diagnosis of mixed ICTs. In a study performed by the Registry of Industrial Toxicology Animal Data (RITA) on spontaneous islet cell carcinomas from control Sprague-Dawley and Wistar rats, it was shown that a majority (93–94%) of islet cell carcinomas were insulin positive with high staining intensity. Gastrin expression was minimal except for one tumor, and coexpression of insulin with somatostatin and glucagon was seen in 16.3% of Wistar rats, while multihormone coexpression was not seen in Sprague Dawley rats (Germann et al. 1999). This illustrates a significant strain difference in peptide hormone expression in these rat tumors. However, in all tumors with marked coexpression of noninsulin peptide hormones, insulin expression still predominated. Similarly, in F344 rats in the present study, insulin expression predominated, gastrin expression was minimal, and approximately 14% to 29% expressed glucagon and somatostatin, respectively, to a significant extent, along the lines of the findings in the RITA study of Wistar rats. Based on the consistently absent to low expression of gastrin, gastrinomas were not a common ICT in this limited group of rat ICTs as compared with humans, in which gastrinomas represent the second most common functional ICT (Goldin, Aston, and Wahi 2008; Halfdanarson, Rubin, et al. 2008; House and Schulick 2006). Brand and Fuller (1988) demonstrated that gastrin mRNA is expressed in the fetal rat pancreas but disappears shortly after birth, and it is absent in the adult rat pancreas but expressed in the gastric antrum. To our knowledge, this is the first comprehensive study of the expression and distribution of noninsulin peptide hormones in the pancreatic islets and proliferative islet cell lesions in F344 rats; furthermore, we have shown that gastrin protein is expressed, albeit rarely, in the adult rat pancreatic islet, which to our knowledge has not been previously demonstrated. However, because of the small size of these lesions in this study of F344 rats, limited available tissue precluded further extensive analysis of the expression of peptide hormones in a larger number of samples. In addition, most of the proliferative lesions in this study were from treatment groups exposed to MBT and BCAA. Although there was not a clear dose-related increase in ICTs in either study from which these samples were acquired, it is nevertheless uncertain how treatment may have influenced the expression of these peptide hormones or the relationship to the development of these hyperplastic and neoplastic lesions. Future studies in which there is an islet cell tumor response will provide additional tissues for more comprehensive evaluation of these proteins in terms of their distribution, localization, and expression level in both spontaneous and treatment-related proliferative islet cell lesions in the F344 rat.
CDK4 expression was significantly increased in islet cell adenomas and carcinomas. CDK4 plays an essential role in β-cell proliferation in both rodents and humans (Cozar-Castellano et al. 2004; Rane et al. 1999) where it complexes with D cyclins, leading to eventual phosphorylation of retinoblastoma protein (Rb), resulting in transition from G1 to the S phase of the cell cycle (Adams 2001). Several mediators in this pathway are dysregulated in various types of human cancer, including ICTs (Hall and Peters 1996; Shapiro 2006). CDK4 is overexpressed in human ICTs (Lindberg et al. 2007), and transgenic mouse models overexpressing the human CDK4 gene develop islet cell hyperplasias and adenomas, whereas knockout models develop islet cell hypoplasia (Rane et al. 1999; Sotillo et al. 2001), illustrating the critical role that CDK4 plays in islet cell regulation and proliferation.
PCNA is a commonly used marker of cell proliferation (Goldsworthy, Butterworth, and Maronpot 1993). It is expressed during the G1-S phase of the cell cycle, plays an important role in DNA repair (Paunesku et al. 2001), and is overexpressed in human islet cell adenomas and carcinomas (Wang, Johnston, and Buchanan 1997). Similarly, rat ICTs overexpressed PCNA, and expression increased in proliferative islet cell lesions with progression toward malignancy, consistent with its role in cell proliferation. Interestingly, in one human study, it was found that ICTs lacking hormone expression had a higher rate of proliferation than those with hormone expression, suggesting loss of endocrine differentiation (von Herbay et al. 1991). This is consistent with the findings in the current study, in which decreased insulin expression was associated with increased expression of PCNA with progression toward malignancy.
Upregulation and nuclear translocation of β-catenin protein is often observed in various human cancers as a result of mutation or constitutive activation of the WNT signaling pathway (MacDonald, Tamai, and He 2009). In human ICTs, membrane and occasional cytoplasmic immunoreactivity is typically observed, with rare nuclear translocation of the protein (Burford et al. 2009; Comper et al. 2009; Hervieu et al. 2006; Liu and Habener 2010). Nuclear translocation was not observed in rat ICTs in this study; instead, a decrease in membrane and cytoplasmic expression was noted. This may indicate that alterations of β-catenin and Wnt signaling do not play a major role in the pathogenesis of ICTs in rats. Alternatively, the diminished expression of β-catenin in these proliferative lesions, although only statistically significant in adenomas, may be indicative of an important phenotypic and biologic transformation. In support of the latter supposition, insulinomas from Men1 knockout mice have reduced membrane expression of β-catenin early in tumor development and progressive loss of expression as tumors progress toward malignancy (Bertolino et al. 2003). The reason for the loss of β-catenin expression is uncertain; however, loss of β-catenin expression has been reported in several types of human cancer, including prostate (Wang et al. 2000), breast (Bukholm et al. 1998), oral cavity and esophagus (Chow et al. 2001; Takayama et al. 1996), stomach (Ebert et al. 2003; Utsunomiya et al. 2000), and thyroid gland (Cerrato et al. 1998). Loss of β-catenin expression in these tumors is associated with decreased cellular adhesion, promotion of cell migration, and a more aggressive phenotype (Foldynova-Trantirkova et al. 2010; Pontes et al. 2010).
In conclusion, we have shown that the immunophenotype of rat ICTs illustrates important similarities to and differences from human ICTs. As in humans, a majority of rat ICTs in this study expressed insulin and were thus characterized as insulinomas. On the other hand, gastrinomas were not observed, and other peptide hormones (glucagon, somatostatin) were not significantly expressed in most of these proliferative lesions. Of particular interest is the potential role of CDK4, PCNA, and β-catenin in human and rat ICTs; although a shared mechanism of tumorigenesis may be present, further studies are necessary to elucidate the role of these mediators in the pathogenesis of ICTs.
Footnotes
Acknowledgments
We wish to thank Ms. Natasha Clayton and Ms. Kimwa Walker of the NIEHS Immunohistochemistry Pathology Support Group for their training and assistance with the immunohistochemical aspects of this study. We also wish to acknowledge the technical expertise of Mr. William Cunningham of the NIEHS Histology Core Laboratory for his assistance in obtaining the highest number of tissue sections possible for these generally small islet cell lesions. This article may be the work product of an employee or group of employees of the National Institute of Environmental Health Sciences (NIEHS), National Institutes of Health (NIH); however, the statements, opinions or conclusions contained therein do not necessarily represent the statements, opinions or conclusions of NIEHS, NIH, or the United States government.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.