
Abstract
Dimethylarsinic acid (DMAV), the major excreted metabolite of inorganic arsenic, is carcinogenic to the rat urinary bladder. Oxidative stress has been proposed as one possible mechanism of DMAV-induced carcinogenesis. The authors determined whether the antioxidant N-acetylcysteine (NAC) modifies DMAV-induced urinary bladder injury in rats. The treatment solutions—DMAV at 10 mg/kg, NAC at 90 or 1.6 mg/kg (high or low dose, respectively), and their combination—were intravesically instilled into female F344 rats over two hours under pentobarbital anesthesia. The treatment was conducted twice with an interval of three days. All animals were euthanized one day after the second treatment. NAC (low dose) alone did not induce histopathological changes or increase 5-bromo-2′-deoxyuridine (BrdU) labeling index in urothelial cells. Both DMAV and NAC (high dose) induced a weak neutrophil infiltration and an increase in the BrdU labeling index; these pathological changes were enhanced by the combined treatment of DMAV and NAC (high or low dose). Increased oxidative stress and urothelial cell hyperplasia with evidence of activated p44/42 MAPK (ERK1/2) and cyclin D1 were found in the DMAV and NAC (high dose) cotreated group. These results suggest that cotreatment with NAC enhanced DMAV-induced urinary bladder injury and that the effects may be mediated by excess oxidative stress and ERK signaling.
Introduction
Chronic arsenic (As) poisoning, which results from the consumption of drinking water containing high or low amounts of inorganic As (iAs), has become a worldwide public health concern (Yoshida, Yamauchi, and Sun 2004). Accumulated evidence suggests that exposure to iAs, arsenite (AsIII), arsenate (AsV), and its trivalent mono- and di-methylated derivatives monomethylarsonous acid (MMAIII) and dimethylarsinous acid (DMAIII) is associated with cancers of the skin, lung, kidney, liver, and urinary bladder (Tapio and Grosche 2006). However, little is known regarding the mode of action of As, and these investigations have been hampered by the insensitivity of rodents to iAs-induced carcinogenesis relative to humans and a general lack of suitable animal models (Cohen et al. 2007).
iAs is readily absorbed from the intestine and enzymatically methylated in the liver into organic forms of As, such as monomethylarsonic acid (MMAV) and dimethylarsinic acid (DMAV), both of which are excreted in the urine as major metabolites. DMAV has the potential to cause carcinogenicity in the rat urinary bladder (Salnikow and Zhitkovich 2008; Wanibuchi et al. 1996; Yamamoto et al. 1995), and DMAV exposure may be relevant to the carcinogenic risk of As to humans (Wei et al. 2002). Therefore, the rat urinary bladder model has been useful in delineating some of the mechanisms that may be involved in the induction of cancer by arsenicals. In this model, the carcinogenic mode of action involves cytotoxicity due to the formation of reactive metabolites, including DMAIII (Salnikow and Zhitkovich 2008), followed by regenerative hyperplasia and eventually leading to neoplastic transformation (Arnold et al. 1999). Oxidative stress has received a great deal of attention in vitro and in vivo because urinary 8-hydroxydeoxyguanosine (8-OHdG) is a key marker for As exposure in humans (Yamauchi et al. 2004) and rats (Kinoshita et al. 2007). Notably, the antioxidants ascorbic acid and N-acetylcysteine (NAC) rescue DMAV-induced cytotoxicity in MYP3 rat bladder epithelial cells in vitro (Wei et al., 2005).
Intravesical instillation of chemicals to rats has been a suitable and reliable method for elucidating cytotoxicity, cell proliferation, and tumor induction in the urinary bladder (Hasegawa et al. 1990; Lijinsky, Thomas, and Kovatch 1992; Masutomi et al. 2001; Shibata et al. 1990). The local instillation, unlike systemic administration, allows us to evaluate the direct effect of a specific metabolite on the urinary bladder. In this study, we determined whether treatment with NAC modifies cytotoxicity and cell proliferation induced by a major metabolite DMAV using an intravesical instillation method. NAC is a derivative of the sulfur-containing amino acid cysteine and is an intermediary in the conversion of cysteine to glutathione (GSH); NAC can expand natural antioxidant defenses by increasing reduced intracellular GSH concentration (Atmaca 2004). Under our study conditions, however, NAC aggravated DMAV-induced urothelial injury.
Materials and Method
Chemicals and Preparation of Treatment Solutions
DMAV (>98% purity) and NAC (>99% purity) were purchased from Sigma-Aldrich (St. Louis, MO, USA). DMAV and NAC were dissolved in saline shortly before each intravesical instillation. A sodium hydroxide solution was added to adjust the pH of the NAC solution to approximately neutral. For the combined treatment, a mixture of equal amounts of DMAV and NAC was prepared at the appropriate concentration immediately prior to instillation.
Animals
Female F344/DuCrlCrlj rats (nine weeks old) were obtained from Charles River Japan (Kanagawa, Japan) and quarantined for one week before initiation of the experiment. Rats were kept in a barrier-sustained animal room maintained at a suitable temperature (24 ± 2°C), humidity (55 ± 15%), ventilation (continuous circulation of fresh air), and illumination (12 hr light/dark cycle). One rat was housed per cage and offered a pelleted diet (Oriental MF, Oriental Yeast Co., Tokyo, Japan) and tap water ad libitum. All the animals were handled in accordance with the Guidelines for Animal Experimentation issued by the Japanese Association for Laboratory Animal Science (1987). The current study was conducted in accordance with the Code of Ethics for Animal Experimentation of the Institute of Environmental Toxicology.
Experimental Design
After a one-week acclimatization period, all animals were randomly divided into seven groups: one untreated and six treated groups. In the treated groups, rats were subjected to intravesical instillation of NAC 1.6 mg/kg (low dose) (N = 4), NAC 90 mg/kg (high dose) (N = 4), DMAV 10 mg/kg (N = 6), DMAV 10 mg/kg plus NAC (low dose) (N = 6), DMAV 10 mg/kg plus NAC (high dose) (N = 5), or the vehicle (saline) (N = 4). Intravesical instillation was performed as described before (Masutomi et al. 2001). The urine remaining in the urinary bladder was removed by mild compression of the lower abdominal regions under pentobarbital anesthesia (30 mg/kg, i.p.). A catheter tube was introduced into the urinary bladder via the urethra, and the test substances described previously was instilled using a syringe. The dosing volume was set at 1 mL/kg body weight. After two hours, the solution in the urinary bladder was excreted via the urethra in rats still under anesthesia. Intravesical instillation was conducted twice (0 and 72 hr) at an interval of three days. An untreated group, not subjected to any form of intravesical instillation, was used as a control (N = 4). The dose of DMAV was selected based on the results of a preliminary experiment where 0.1, 1, 10, or 100 mg/kg of DMAV was instilled once into the urinary bladder of female rats (N = 1 per group) for two hours. Ten milligrams per kilogram was judged as a suitable dose level in the present study, as it induced mild inflammation in the urinary bladder (data not shown). When sodium arsenite was administered in feed to female F344 rats at a dose level of 100 ppm for five weeks, pathological findings, namely, cytotoxicity, cell proliferation, and hyperplasia of urothelial cells, were observed, and urinary concentration of a metabolite DMAV was found to be 665.2 µM at week 4 (Suzuki et al. 2010). The dose of DMAV selected in this study was approximately 100-fold higher than the level reported by Suzuki et al. (2010). The dose of NAC was selected based on the results of in vitro study previously reported (Wei et al. 2005).
All animals were euthanized one day after the second treatment under diethyl ether anesthesia. One hour before euthanasia, all animals were injected intraperitoneally with 100 mg/kg of 5-bromo-2′-deoxyuridine (BrdU; Sigma-Aldrich) as previously described (Shibata et al. 1990). At necropsy, the urinary bladder was inflated with 10% neutral-buffered formalin and then further fixed in this solution for twenty-four hours. The tissue was cut longitudinally, routinely embedded in paraffin, and stained with hematoxylin and eosin (H&E) for histopathological examination.
Immunohistochemistry and Analysis of Immunopositive Reaction
Immunohistochemistry was performed using the Dako EnVision kit (Dako, Glostrup, Denmark). Additional paraffin sections from all animals were incubated with monoclonal antibodies against BrdU (×1,000; Dako). Other sections from four animals selected in the groups excluding the NAC (low dose) alone treated group and DMAV plus NAC (low dose) treated group were incubated with monoclonal antibodies against p44/42 MAPK (ERK1/2) (×200; Cell Signaling Technology, Danvers, MA, USA), phosphorylated p44/42 MAPK (ERK1/2) (Thr202/Tyr204) (×100; Cell Signaling Technology), cyclin D1 (×100; Cell Signaling Technology), 3-nitrotyrosine (3-NT) (×50; Alexis Biochemicals, San Diego, CA, USA), and 4-hydroxy-2-nonenal (4-HNE) (×20; Japan Institute for the Control of Aging, Shizuoka, Japan). For BrdU, the section was pretreated with proteinase K solution (Dako) for fifteen minutes and soaked in 4N HCl for twenty minutes. For others, the sections were pretreated with microwave irradiation in 0.1 M citrate buffer at pH 6.0 for ten minutes. Immunohistochemical products were visualized with the substrate 3.3′-diaminobenzidine and counterstained with hematoxylin. The duodenum tissues from all animals were used as positive control for BrdU, and positive reactions in all animals were obtained. For negative control, phosphate-buffered saline replaced the primary antibodies. To determine the BrdU-positive urothelial cell ratio, 1,000 urothelial cells were randomly counted manually under a light microscope. BrdU-positive cells in the neck of the urinary bladder were excluded from the estimation because of mechanical injury caused by catheterization. The BrdU labeling index (LI) was calculated by dividing the number of labeled nuclei by the total number of nuclei counted, and the results were expressed as percentages. To analyze immunointensities of 3-NT and 4-HNE, all the sections were stained under the same conditions. The immunointensities of 3-NT and 4-HNE were measured with an image analyzer (Nireco, Tokyo, Japan) as previously described (Yoshida et al. 2001). For each section, five fields were randomly selected and measured.
Statistical Analysis
The data for BrdU LI and immunointensity of 3-NT and 4-HNE are presented as the mean ± standard deviation. Multiple group data were analyzed using Holm’s multiple comparison test. The incidences of each of the histopathological findings were analyzed using the Fisher’s exact probability test (one-tailed analysis); p values below .05 and .01 were judged significant.
Results
Histopathological Examination of the Urinary Bladder
The histopathological findings are summarized in Table 1 . No histopathological changes were observed in the vehicle control (Fig. 1A), NAC (low dose) alone, or untreated control groups. Inflammation, mainly composed of neutrophil infiltration in the urothelial and submucosal tissues, was observed in one of four rats treated with DMAV alone and one of six rats treated with NAC (high dose) alone (Fig. 1B). In contrast, an inflammatory response was observed in four of six rats or three of five rats treated with DMAV and NAC (low or high dose), respectively. Surprisingly, urothelial cell hyperplasia was observed in two of five rats cotreated with DMAV and NAC (high dose); one was nodular hyperplasia (Fig. 1C) and the other was papillary hyperplasia (Fig. 1D). Submucosal edema and hemorrhage was concurrently seen. Diffuse simple hyperplasia was also present in the area surrounding nodular or papillary hyperplasia. The area of the urothelial cell hyperplasia including simple hyperplasia occupied about half in the longitudinal section of the bladder. These hyperplastic lesions observed diffusely were considered to be caused by the treatment with DMAV and NAC (high dose). There were no proliferative lesions, including simple hyperplasia, in any other treated groups.
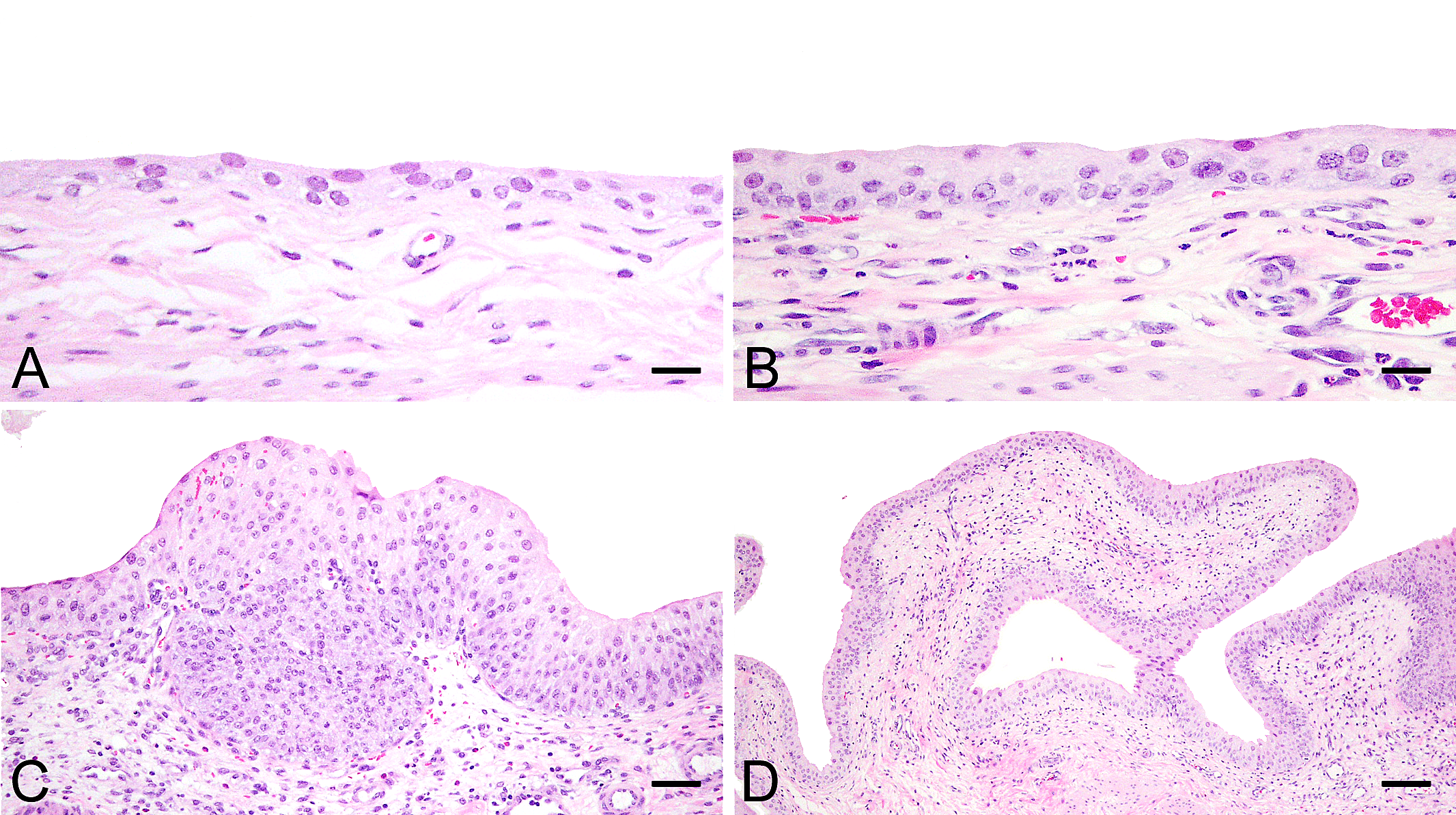
Representative histopathological changes in the urinary bladder, hematoxylin and eosin-stained. (A) Normal urothelial cells in the vehicle control group (scale bar = 18 µm). (B) Submucosal neutrophil infiltration in the N-acetylcysteine (NAC) (high dose) treated group (scale bar = 18 µm). (C, D) Nodular and papillary urothelial cell hyperplasia in the group cotreated with dimethylarsinic acid (DMAV) and NAC (high dose) (scale bar = 100 and 270 µm, respectively).
Histopathological findings in the urinary bladder.
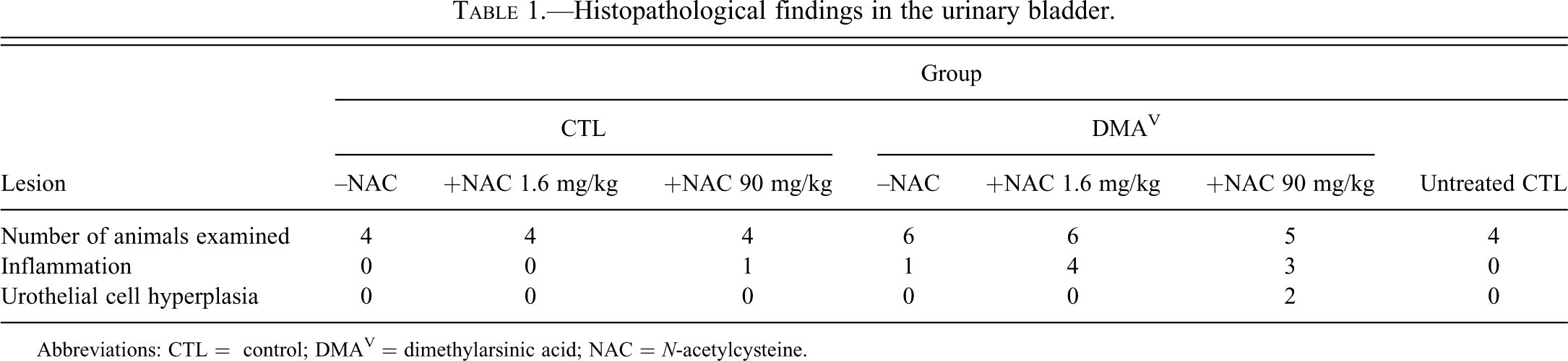
Abbreviations: CTL = control; DMAV = dimethylarsinic acid; NAC = N-acetylcysteine.
Analysis of Cell Proliferation Activity and Immunohistochemical Detection of Cell Proliferation Signaling
To assess cell proliferative activity, we counted the number of BrdU-positive urothelial cells in the urinary bladder. Increased BrdU LI in urothelial cells was not observed in NAC (low dose) alone treated group. The BrdU LI was significantly increased in the DMAV alone or NAC (high dose) alone treated group compared with the vehicle control group (Figs. 2A–D). Combined treatment of DMAV and NAC (high or low dose) markedly increased the BrdU LI, and the increases in these groups were statistically significant compared with the vehicle control or DMAV alone treated group (Figs. 2A and 2E). These results indicated that cotreatment with NAC (high or low dose) enhanced DMAV-induced cell proliferation. The BrdU LI of the vehicle control group (1.6 ± 2.0%) was slightly higher than that of the untreated group (0.3 ± 0.2%), although this apparent difference was not statistically significant. This observed effect was presumably due to nonspecific mechanical injury associated with the intravesical instillation.
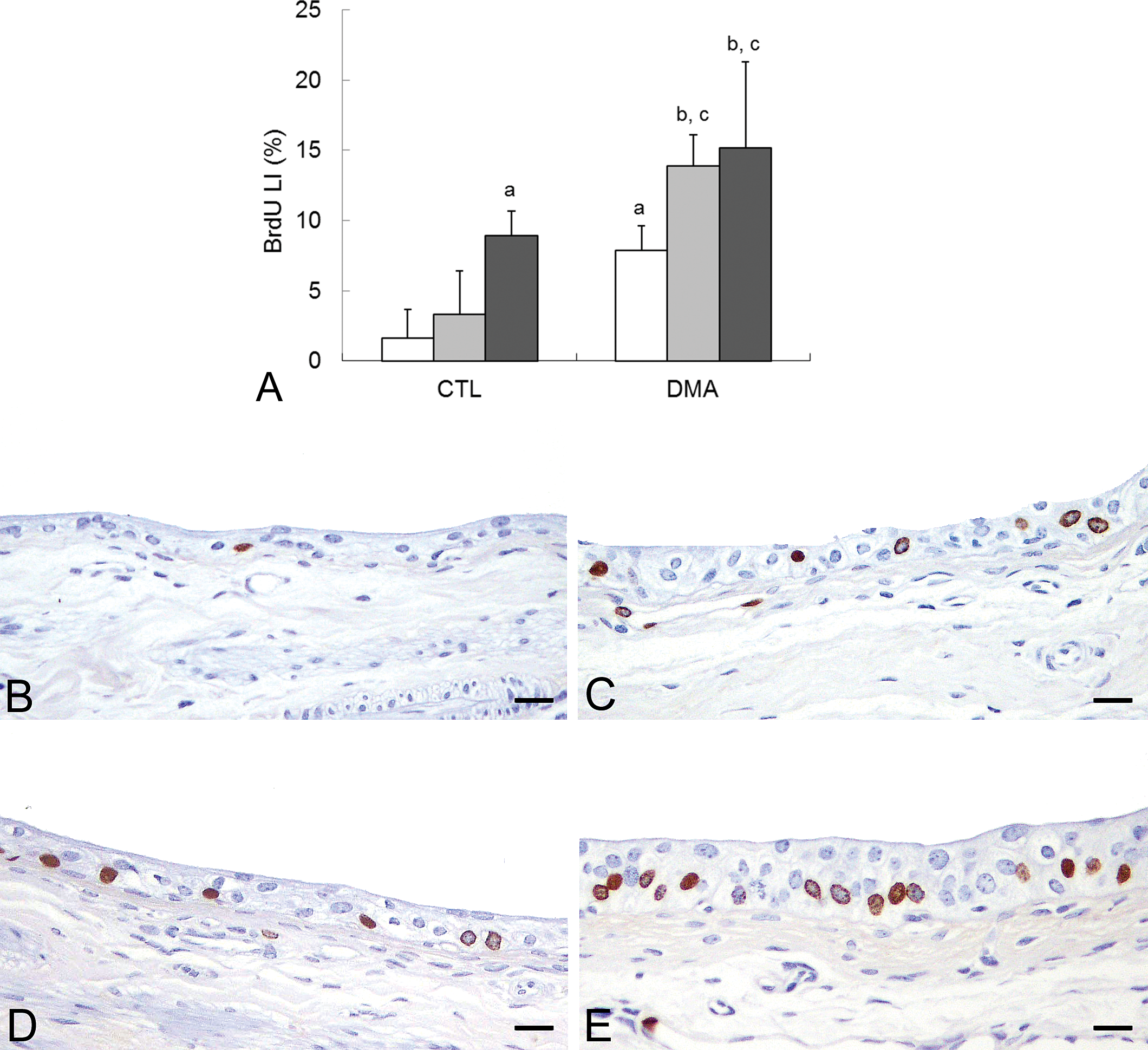
The 5-bromo-2′-deoxyuridine (BrdU) labeling index and representative BrdU-positive urothelial cells in the urinary bladder. (A) The BrdU labeling index. Data are represented as the mean ± standard deviation and were analyzed by the Holm’s multiple comparison test (a, p < .05 vs. vehicle control group; b, p < .01 vs. vehicle control group; c, p < .05 vs. dimethylarsinic acid [DMAV] treated group). (B) Normal urothelial cells in the vehicle control group. (C, D) Scattered BrdU-positive urothelial cells in N-acetylcysteine (NAC) (high dose) or DMAV treated group. (E) Frequent BrdU-positive urothelial cells in the group cotreated with DMAV and NAC (high dose). Scale bar = 18 µm.
Because increased cell proliferation activity was observed in the DMAV and/or NAC treated groups, we determined signal transduction to be related to cell proliferation using immunohistochemical staining. Normal urothelial cells (nonproliferative) of the vehicle control group showed prominent positive reaction for ERK1/2 but slightly focal positive reaction for phosphorylated ERK1/2 (Figs. 3A and 3C). Similar positive reactions were observed in normal urothelial cells of other treated groups including the DMAV and NAC (high dose) treated groups (data not shown). Thus, normal urothelial cells of the aforementioned groups expressed ERK1/2, but mostly as the inactive (underphosphorylated) form. In contrast, immunopositive reactions for both ERK1/2 and phosphorylated ERK1/2 were observed in the area of urothelial cell hyperplasia in the DMAV and NAC (high dose) treated group (Figs. 3B and 3D). Some of the phosphorylated ERK1/2 appeared to have translocated into the nuclei. These results indicated that activation of ERK1/2 is important for formation of hyperplastic lesions. In addition, immunopositive reactions of cyclin D1 were observed only in the area of urothelial cell hyperplasia, not in normal urothelial cells in all groups examined (Figs. 3E and 3F).
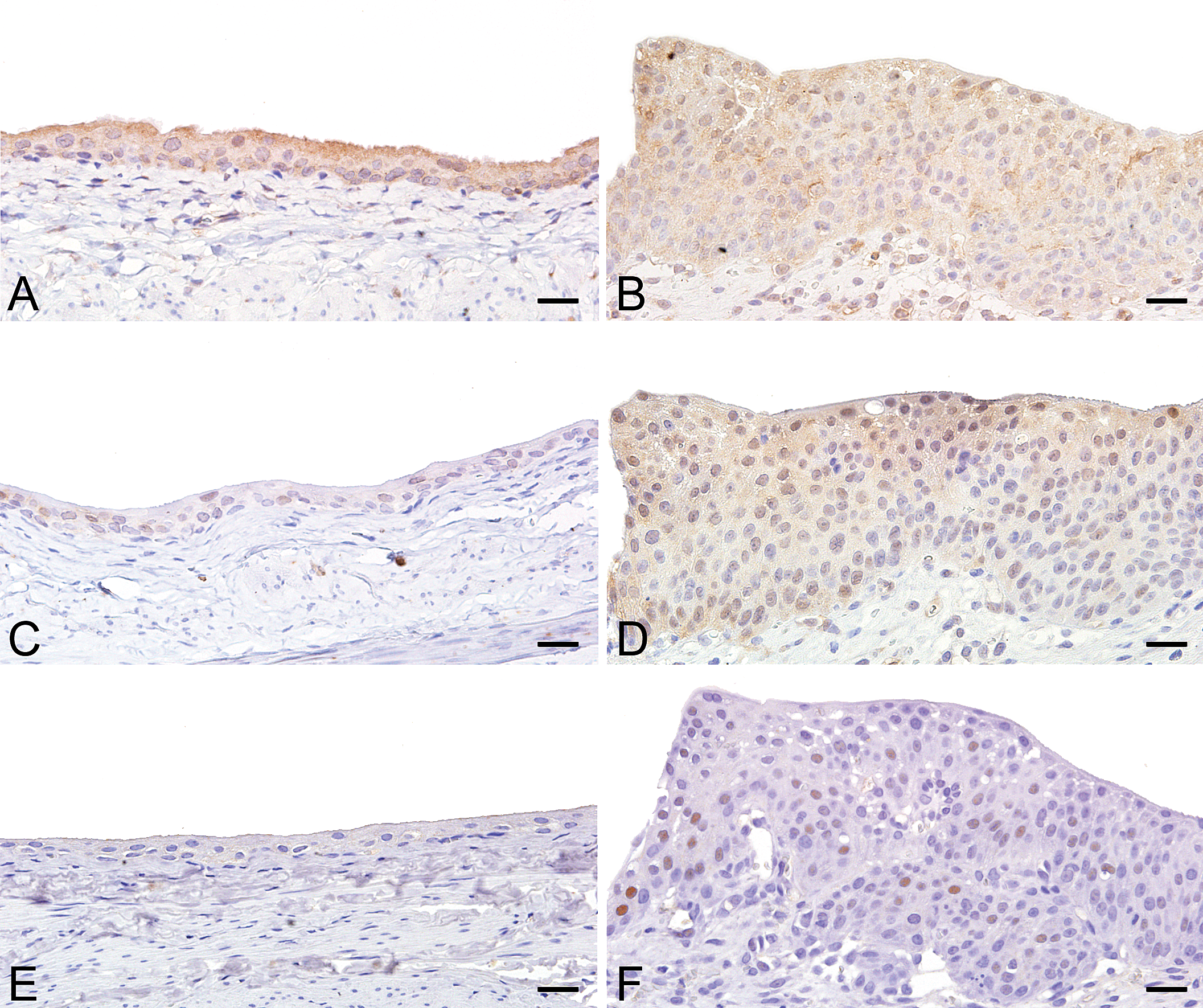
Representative immunostaining images of cell proliferation markers. (A, B) p44/42 MAPK (ERK1/2). Prominent positive reactions in normal urothelial cells in the vehicle control group (A) and in hyperplasia in the group cotreated with dimethylarsinic acid (DMAV) and N-acetylcysteine (NAC) (high dose) (B). (C, D) Phosphorylated p44/42 MAPK (ERK1/2). Slightly focal positive reaction in normal urothelial cells in the vehicle control group (C) and prominent positive reaction in hyperplasia in the group cotreated with DMAV and NAC (high dose) (D). (E, F) Cyclin D1. No positive reaction in normal urothelial cells in the vehicle control group (E) and frequent cyclin D1-positive urothelial cells in hyperplasia in the group cotreated with DMAV and NAC (high dose) (F). Scale bar = 20 µm.
Semi-Quantitative Analysis of the Oxidative Stress Marker
Next, we examined whether oxidative stress was related to cell proliferation. The immunointensity of 3-NT, which is known as a biomarker that indicates oxidative damage to protein, was slightly increased in the DMAV alone or NAC (high dose) alone treated group compared with the vehicle control group (Figs. 4A–D). These immunointensities were enhanced by the combined treatment with DMAV and NAC (high dose), and this was statistically significant compared with that in the vehicle control or NAC (high dose) alone treated group (Figs. 4A and 4E). In contrast, no clear difference was observed in the immunointensity of 4-HNE, a major product of lipid peroxidation, between the DMAV and/or NAC (high dose) treated groups and the vehicle control group (data not shown).
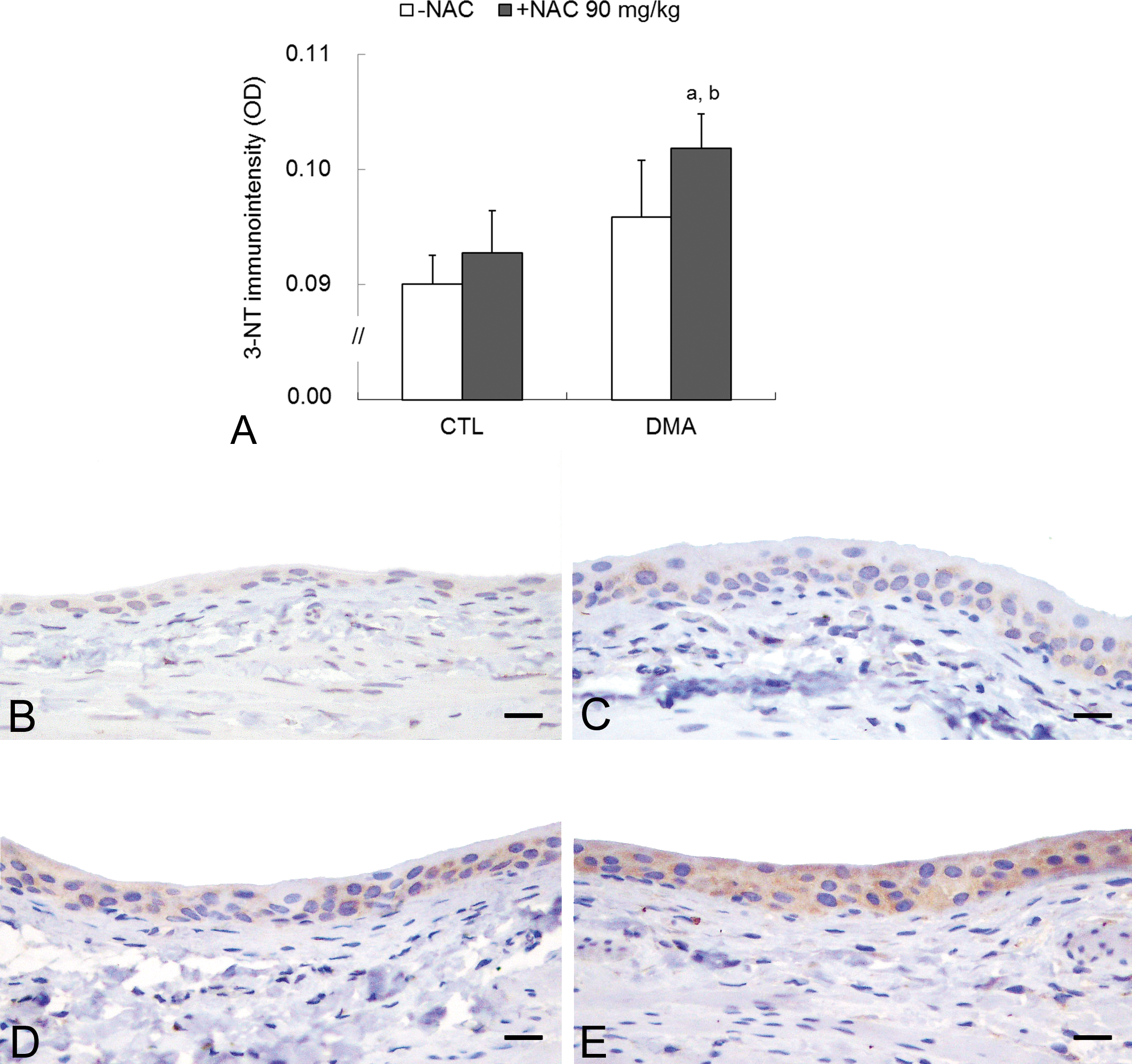
Analyses of oxidative stress marker 3-NT in the urinary bladder. (A) Immunointensity of 3-NT. Data are represented as the mean ± standard deviation and were analyzed by the Holm’s multiple comparison test (a, p < .01 vs vehicle control group; b, p < .05 vs. N-acetylcysteine [NAC] (high dose) treated group). (B–E) Representative images of 3-NT immunostaining. (B) Very weak positive reactions in normal urothelial cells in the vehicle control group. (C, D) Weak positive reactions in the NAC (high dose) or dimethylarsinic acid (DMAV) treated group. (E) Prominent positive reactions in the group cotreated with DMAV and NAC (high dose). Scale bar = 18 µm.
Discussion
Oxidative stress has been proposed as a possible mechanism of DMAV-induced carcinogenic action in the urinary bladder of rats. Thus, we determined whether the antioxidant NAC would modify DMAV-induced urinary bladder injury in rats using an intravesical instillation method. The present study paradoxically showed that cotreatment with NAC enhanced DMAV-induced inflammation and cell proliferation, eventually forming papillary/nodular hyperplasia in the urinary bladder. These changes were probably accompanied by increased oxidative stress and activated ERK1/2 and cyclin D1 as demonstrated by immunohistochemistry. The upregulation of ERK signaling is consistent with an early change in bladder tumorigenesis in humans (Mo et al. 2007) and controls positive regulation of the continued expression of cyclin D1 in human urothelial cell (Huang et al. 2011). ERK signaling is also known to be activated by oxidative stress or mitogenic stimuli (Luster and Simeonova 2004). Therefore, these results suggest that oxidative stress–mediated ERK activation has a crucial role in enhancing effects of NAC on DMAV-induced urothelial injury and proliferation.
Both NAC and GSH are thiol (-SH) compounds, and their direct scavenging of hydroxyl radicals has been suggested as the mechanism by which they protect against oxidative damage. Nevertheless, it has been emphasized that the interaction of thiols with reactive radicals can generate thiyl radicals (Sagristá et al. 2002). Aitio (2006) reviewed the unbeneficial effects of NAC in human disorders supposedly linked to oxidative stress. For example, inhaled NAC promoted hydroxyperoxide generation in patients with chronic obstructive pulmonary disease (Szkudlarek et al. 2004). When 1.2 and 2.4 g/day of NAC were given orally to healthy volunteers, the blood concentration of glutathione disulfide (GSSG; oxidized glutathione) was higher than that in the controls, and lower concentrations of GSH and GSH/GSSG ratio were observed, indicating that NAC likely acts as a pro-oxidant at the dose used (Kleinveld, Demacker, and Stalenhoef 1992). Intriguingly, the postadministration, but not preadministration, of NAC aggravated acute ethanol-induced liver damage because of a high level of hepatic lipid peroxidation (Wang et al. 2006). In the present study, only NAC (high dose) treatment induced inflammation and cell proliferation, suggesting that NAC acts as a pro-oxidant under the conditions of this study; combined treatment with DMAV and NAC induced oxidative stress in an additive manner, resulting in more severe injury to urinary bladder epithelium.
Another possibility in terms of a mechanism of action is that NAC generates reactive metabolites from DMAV as a reducing agent. The metabolism of As is critical for its cytotoxicity and carcinogenicity. Once inside the cell, AsV is reduced enzymatically to AsIII, which is more reactive than AsV. AsIII is oxidatively methylated by the addition of a methyl group (MMAV) and is reduced to trivalency (MMAIII) and then metabolized to DMAV and DMAIII by repeating the similar reaction, which are the most active metabolites (Aposhian et al. 2004; Thomas, Waters, and Styblo 2004). GSH mediates the generation of a DMA-GSH conjugate as well as DMAIII from DMAV, eventually producing dimethylarsenic peroxide, dimethylarsenic radicals, and dimethylarsenic peroxy radicals (Yamanaka et al. 2004). Sakurai et al. (2004) clearly demonstrated that cellular GSH is required for DMAV-induced apoptosis, and they proposed that the activation of caspases after DMAV-GSH conjugation appeared to be of mechanistic significance. NAC is a precursor of GSH, suggesting that it may mediate the reduction of DMAV to generate oxidants and impair the balance between its pro-oxidant and antioxidant properties in intravesical circumstances. In addition, glutathione-S-transferase-omega (GSTO) catalyzes the reduction of MMAV and DMAV in the presence of GSH (Aposhian et al. 2004). This enzyme shows higher distribution in the urinary bladder than in other tissues in hamsters (Sampayo-Reyes et al. 2000) and mice (Chowdhury et al. 2006).
The present study showed unbeneficial effects of NAC on DMAV-induced urothelial injury and proliferation in the rat urinary bladder via the intravesical instillation of both agents. These results raise two possibilities on the exacerbating effect of NAC in response to DMAV. First, NAC acts as a pro-oxidant; second, NAC generates reactive metabolites from DMAV. Further studies are required to determine the role of this antioxidant in the mode of action of DMAV in the urinary bladder.
Footnotes
The author(s) declared no potential conflicts of interests with respect to the authorship and/or publication of this article. The author(s) received no financial support for the research and/or authorship of this article.
Acknowledgments
We thank Yuko Chiba, Mutsumi Kumagai, Takako Kazami, Yukie Jibiki, Satoshi Akema, Haruka Miura, Chizuko Tomiyama, and Kayoko Iijima for their assistance with tissue preparation.