
Abstract
7,12-Dimethylbenz[a]anthracene (DMBA) presents a pollutant implicated in various toxicological effects. The aim of this experiment was to study the effects of DMBA administration on oxidative stress, histopathological signs, and 18 kDa translocator protein (TSPO) binding characteristics in rat liver. We also studied the effects of dose stoichiometry, dose frequency, and duration of protocol of DMBA administration. In this study, rats surviving eighteen weeks after DMBA exposure showed mild to moderate histopathological changes in the liver, mainly characterized by glossy appearance of hepatocytes, heterochromatic nuclei, and glycogen overload in the midzonal region of the hepatic lobe. These changes were accompanied by significant rises in oxidant levels, along with declines in nonenzymic antioxidants, indicating that DMBA induced oxidative stress in the liver. This finding correlated well with decreases in TSPO binding capacity in the liver of the rats in our study. Other studies have shown that TSPO can be affected by oxidative stress, as well as contribute to oxidative stress at mitochondrial levels. Further studies are needed to assay whether the decreases in TSPO density in the liver are part of the damaging effects caused by DMBA or a compensatory response to the oxidative stress induced by DMBA.
Keywords
Introduction
The polycyclic aromatic hydrocarbon (PAH) 7,12-dimethylbenz[a]anthracene (DMBA), which is abundantly present in cigarette smoke, polluted air, and charcoal-broiled and smoked food, has been implicated in various toxicological effects, including atherogenesis and carcinogenesis (Curfs et al. 2004; Miller and Ramos 2001; Van Schooten et al. 1998; Waldman et al. 1991). For example, exposure to PAHs, including DMBA, can lead to toxicological changes in the liver, including oxidative stress and production of carcinogenic metabolites (DiGiovani and Juchau 1980). Notably, DMBA is among the most potent chemical carcinogens known (Samy et al. 2006; Singletary and Liao 1989). In this context, in vitro experiments have indicated that DMBA in rat liver microsomes causes dose- and time-dependent production of the oxidant H2O2, which is closely associated with tumor promotion (Frenkel et al. 1981; Frenkel et al. 1993). For these reasons, in many studies DMBA is used to induce oxidative stress in mice, and the animals' livers are used to assay the level of oxidative state (Choi 2008).
It is well known that mitochondria are particularly vulnerable to oxidative damage. Lipid peroxidation and protein oxidation can affect many metabolic enzymes in the mitochondrial matrix, including the electron transport system (Netto et al. 2002; Oliveira et al. 2006; Szeto 2006). In the present study, we investigated the potential involvement of the 18 kDa mitochondrial translocator protein (TSPO) in harmful effects of DMBA in the liver. The TSPO is present in various organs, including the liver (Gavish et al. 1999). The TSPO’s primary intracellular location is the outer mitochondrial membrane (Anholt et al. 1986; Krueger and Papadopoulos 1990; O’Beirne et al. 1990; Veenman et al. 2007). Studies have suggested that the TSPO is involved in the regulation of the mitochondrial electron transport system (Hirsch et al. 1989; Moreno-Sánchez et al. 1991).
In addition, a large number of studies have indicated that mitochondrial TSPO is involved in various other functions. It is well documented that the TSPO is involved in the regulation of cholesterol transport into mitochondria in relation to bile production and steroidogenesis, in oxidative stress and apoptosis, and in inflammatory and immune responses (Gavish et al. 1992; Knudsen et al. 1993; Krueger and Papadopoulos 1990; Kunduzova et al. 2004; Papadopolous et al. 2006; Veenman and Gavish 2006; Veenman et al. 2007; Veenman et al. 2008; Veenman et al. 2010; Veenman et al. forthcoming). Previous studies have suggested that TSPO density may be correlated with the antioxidant status of liver cells (Fischer et al. 2001) and that it may play a role in organisms' response to pollution (Giannaccini et al. 2004). Interestingly, we have found that the TSPO appears to be an active participant in the generation of reactive oxygen species (ROS) at mitochondrial levels and in the modulation of the mitochondrial membrane potential, thereby participating in the induction of the mitochondrial apoptosis cascade (Kugler et al. 2008; Veenman et al. 2008; Zeno et al. 2009). In turn, ROS levels are also known to affect TSPO function (Delavoie et al. 2003). Recently, we found that the TSPO is down-regulated in the aorta wall of rats in correlation with pathological changes in the aorta wall resulting from DMBA exposure (Dimitrova-Shumkovska et al. 2010a). Thus, we assumed that, in addition to causing oxidative stress and pathological changes in the liver, DMBA exposure of rats would also lead to changes in TSPO binding characteristics in this organ.
Several studies have focused on the effects of DMBA on biochemical and antioxidant parameters in the liver (Choi 2008; Girolami et al. 2008; Muqbil and Banu 2006). As the liver is considered to suffer from oxidative stress owing to toxicological stress following DMBA exposure (Choi and Kim 2009), we applied various doses and times of DMBA exposure and assayed potential dose-dependent effects of DMBA on hepatic histopathology and oxidative stress. In the present study, we applied DMBA to otherwise healthy, outbred animals to be able to better study the effects of DMBA per se, that is, without concomitant phenotypic and/or genotypic peculiarities. Apart from evaluation of liver histopathology and oxidative stress parameters, we assayed changes in TSPO binding characteristics in the liver following DMBA exposure of rats.
Materials and Methods
Materials
Wistar rats were obtained from the Military Medical Academy (VMA, Belgrade, Republic of Serbia) and maintained and bred in our own animal facilities at the Department of Physiology and Biochemistry of the Faculty of Natural Sciences and Mathematics, Skopje, Republic of Macedonia. Rat feed was obtained from Filpaso (Filpaso, 52.11, Skopje, Republic of Macedonia). [3H]PK 11195 was obtained from New England Nuclear (Boston, MA, USA) and PK 11195 from Sigma-Aldrich (Rehovot, Israel). 7,12-Dimethylbenz[a]anthracene (DMBA) was purchased from Sigma-Aldrich. Enzyme-colorimetric test kits were obtained from Human Diagnostics (Wiesbaden, Germany). The SOD determination kit RA20408 was obtained from Fluka-Biochemika (Steinheim, Germany). The glutathione assay kit (CS0260) and glutathione reductase assay kit (GRSA114K4000) were from Sigma-Aldrich (Steinheim, Germany). Trichloracetic acid (TCA), 2-thiobarbituric acid (TBA), trifluoroacetic acid (TFA), 1,1,3,3-tetraethoxypropane (TEP), 5,5′-dithio2-nitrobenzoic acid (DTNB), T-chloramine, and 1M phosphate buffer solution (PBS) were from Sigma-Aldrich. 2,4-Dinitrophenylhydrazine (DNPH), Folin-Ciocalteu reagent, guanidine chloride, ascorbic acid, and ammonium iron (II) sulfate -(NH4)2 Fe (SO4)2 were obtained from Merck, Darmstadt, Germany. Standard chemicals and consumables were obtained from various commercial sources.
Research Subjects
Male Wistar rats, 20 weeks old at the start of the experiment, were used. The rats were kept on a regular dark/light cycle, and they received water and regular chow ad libitum. All animals procedures were reviewed and approved by the local ethics committee, in accordance with National Institutes of Health (USA) Guidelines for the Care and Use of Experimental Animals (NIH publication No. 85-23, revised 1996).
Treatment and Tissue Handling
Administration of DMBA
For each experiment, DMBA was dissolved in sesame oil, as described previously (Carter and Carter, 1988; Whitsett et al. 2006; Dimitrova-Shumkovska et al. 2010a). The animals received different doses of DMBA by intragastral intubation, as described previously (Löcher et al. 2001; Motoyama et al. 2008; Dimitrova-Shumkovska et al. 2010a) and were sacrificed at various periods after DMBA application, as listed below. The control group (C) was exposed to vehicle, that is, 1 mL of sesame oil administered once by intragastral intubation, and sacrificed 18 weeks after DMBA administration (n = 16). The DS 10 group (n = 6) received a single dose of DMBA (10 mg in 1 mL of sesame oil). Animals of the DS-10 group were sacrificed 10 weeks after DMBA administration. The DS 20 group (n = 18) was exposed to a single dose of DMBA (20 mg /1 mL sesame oil), and sacrificed 18 weeks after DMBA administration. The DR 20 group (n = 24) was exposed to DMBA four times at weekly intervals (i.e., 4 × 5 mg of DMBA in 0.5 mL of sesame oil), and sacrificed 18 weeks after the first dose of DMBA.
One of the goals of this experiment was to correlate the stoichiometry of dose, frequency of administration, and time period after DMBA initiation. The main set of goals was to determine potential correlations in the liver between TSPO binding characteristics, oxidative stress, and histopathological changes after exposure to DMBA.
Tissue Collection
The rats were heavily anesthetized with a ketamine/xylazine mixture (90 mg/kg ip and 10 mg/kg ip, respectively), applied after an overnight fast of twelve hours. Blood was taken from the heart by cardiac puncture. Liver tissue samples were harvested between 10:00 AM and noon. The livers were flash-frozen, weighed, and stored at –80°C until further analysis.
As described previously (Dimitrova-Shumkovska et al. 2010a,b), for assays of the oxidative stress markers thiobarbituric acid–reactive substances (TBARs), glutathione (GSH), protein carbonyls (PC), superoxide dismutase activity (SOD), and glutathione reductase (GSSG-Red), tissue homogenates were prepared in 1.12% KCl at + 4°C. For advanced oxidation protein products (AOPPs), tissue homogenates were prepared in 50 mM PBS at + 4°C. For these oxidative stress assays, we used an ultrasonic homogenizer (Cole-Parmer Instrument Co., Chicago, IL, USA), as described previously (Dimitrova-Shumkovska et al. 2010a,b). For TSPO binding assays, tissue homogenates were prepared in 50 mM PBS on ice using a Kinematika Polytron, as described previously (Danovich et al. 2008; Dimitrova-Shumkovska et al. 2010a,b).
Histopathology
Livers of the rats in the four experimental groups were subjected to histopathological analysis. The liver was blocked in 10 × 5 × 5 mm pieces. After removal and dissection, the liver blocks were fixed in 10% buffered formalin and processed for embedding in paraffin. Sections (3–5 µM) from blocked liver of animals from each experimental group were cut with a microtome and examined with a light microscope mounted with a video camera (Nikon-Eclipse Düsseldorf, Germany). All slides were routinely stained with hematoxylin and eosin (HE) and with the Periodic Acid Schiff method (PAS) for glycogen deposits (Merck, Germany).
Protein Quantification
General protein content of liver homogenates for carbonylization assays was measured according to Levine et al. (1990). For [3H]PK 11195 binding assays and specific assays for oxidative stress, general protein content in the tissues in question was measured according to Lowry et al. (1951).
Lipid Peroxidation Assays
Lipid peroxidation products (LPO) from liver were measured as TBARs in 2.5% (w/v in 1.12% KCl) of liver homogenates according to the method of Okhawa et al. (1978), modified by Draper and Hadley (1990). For oxidant-stimulated lipid peroxidation, liver homogenates were incubated for thirty minutes with 4 × 10−5 M Fe2+ − 2.6 mM ascorbic acid. The results of the TBAR assays were presented as nanomoles of TBARs per gram liver tissue, using 1,1,3,3-tetraethoxipropane (TEP) as external standard, measured at 535 nm with a spectrophotometer (Cintra, UV/Vis Spectrometer, GBC, Melbourne, Australia).
Protein Carbonyl Assays
Levels of protein carbonyls (PC) were determined in liver homogenates (2.5 % w/v in 1.12% KCl), according to Levine et al. (1990). The carbonyl content was calculated from the spectrophotometric absorbance measurement at 340 nm (Cintra) with the use of a molar absorption coefficient of 22,000 M-1cm−1.
Advanced Oxidation Protein Product Assays
Levels of advanced oxidation protein products (AOPPs) in liver homogenates were determined spectrophotometrically at 340 nm (Cintra, UV/Vis Spectrometer) and calibrated with chloramine-T solutions, which in the presence of potassium iodide absorb at 340 nm (Witko-Sarsat et al. 1996). Briefly, 1 mL of 2.5% tissue homogenate diluted 1/20 in 50 mM PBS (containing butylated hydroxytoluene, 100 µM and EDTA, 1 mM) was placed in quartz cuvettes and 50 µL of 1.16 M potassium iodide was added, followed by 200 µL of acetic acid after two minutes. In quartz cuvettes, used as blanks, 50 µL of 1.16 M potassium iodide was added to 1 mL of PBS followed by 200 µL of acetic acid. Advanced oxidation protein product concentrations were expressed as nanomoles per mg protein of analyzed tissue.
Tissue Antioxidant Analysis
Flash-frozen liver tissue was ground to a fine powder and treated with 5% trichloroacetic acid (TCA) followed by centrifugation at 10,000 × g, at 4°C, for ten minutes. The supernatant was assayed spectrophotometrically for activity of SOD, GSH, and GSSG-Red. Briefly, liver superoxide dismutase activity (SOD) (EC 1.15.1.1) was assayed according to Winterbourn et al. (1975), with an SOD determination kit (RA20408, Fluka, Biochemika, Steinheim, Germany), according to which one unit of SOD activity is defined as the amount of enzyme activity that causes 50% inhibition of the reduction of tetrazolium salt, WST-1 (2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4 -disulfophenyl)-2H-tetrazolium, monosodium salt). Superoxide dismutase is capable of inhibiting the formation of the water-soluble formazan dye typically formed by reduction of WST-1 with superoxide anions. Superoxide dismutase activity in liver was expressed as units of SOD per milligram protein according to the instructions of the manufacturer (Fluka, Biochemika).
Liver glutathione (GSH) content was assayed by measuring acid-soluble glutathione levels using 5,5′-dithiobis (2-nitrobenzoic acid) (DTNB) to give a TNB that absorbs at 412 nm with a glutathione assay kit (CS0260), according to methods described previously (Akerboom et al. 1981). Liver glutathione reductase (GSSG-Red; EC 1.6.4.2) activity was assayed with a glutathione reductase assay kit (GRSA 114K4000) according to previously described methods (Dolphin et al. 1989; Garcia-Alfonso et al. 1993).
TSPO Binding Characteristics
Maximal binding capacity (Bmax) and equilibrium dissociation constant (Kd) of the binding of the TSPO-specific ligand [3H]PK 11195 were determined in whole cell membrane homogenates from the liver, as previously described (Awad and Gavish 1987). Radioactivity was counted with a 1600CA Tri-Carb liquid scintillation analyzer (Packard, Meriden, CT, USA). Scatchard analysis of [3H]PK 11195 binding was done to determine the Bmax and Kd values. In addition, TSPO binding characteristics were determined in other organs such as kidney, brain, testes, and heart, according to the same methods as described for the liver.
Statistical Analysis
Data are presented as means ± SD (n > 5). Significance was determined using analysis of variance and post hoc analysis as appropriate, whereas for dependent groups, the paired-sample t test was used. Significance of association was determined by the two-tailed t test. STATISTICA 5.0 (Stat Soft Inc., Tulsa, USA) was used, and p < .05 was considered indicative of significant differences. Pearson’s correlation coefficient was used as a measure of linear association between two variables.
Results
The rats of this study were observed daily, and the vast majority appeared healthy throughout the course of the experiment, with the exception of the death of one rat in the DS 20 group (5.6%) and two rats in the DR 20 group (8.3%). No rats died in the DS 10 group. No morphologically visible abnormal outgrowths appeared in any of the rats. At sacrifice, gross examination of internal organs generally revealed no abnormalities. Thus, no obvious tumorigenic effects were observed in the DMBA-exposed male rats with the gross anatomical investigation of the present study. However, in some rats treated with 20 mg of DMBA, reductions in testis size of approximately 30% were observed (in two DS 20 rats, and in three DR 20 rats).
At gross anatomic levels, DMBA treatment could have resulted in pale and swollen livers in individual rats of the different groups. Histopathology of liver tissue (Figure 1, Table 1) included glossy appearance of hepatocytes and heterochromatic nuclei (Figure 1B, Table 1). In the animals of the DR 20 group, areas of glossy appearance appeared throughout the affected livers of the group (in 15 of 23 animals) (Figures 1B–D), whereas in the DS 10 and DS 20 groups, the glossy appearance was subcapsular, that is, restricted to peripheral regions (in 4 and 11 animals of these groups of 6 and 12 animals, respectively; Figure 1C, Table 1). In affected livers, erythrocytes appeared coagulated, as condensed conglomerates attached to the endothelial surfaces of sinusoids, which were visibly thickened (in 4 animals from the DS 10 group; in 11 animals from the DS 20 group; and in 15 animals from the DR 20 group, which corresponds to 67%, 61%, and 65% of the analyzed samples, respectively) (Figure 1D, Table 1). Mentioned changes did not differ substantially between DMBA treated groups, but some could be more pronounced in animals of the DR 20 group. For example, in some liver slides of the latter, the sinusoid was no longer visible, accompanied by disorganized hepatic parenchyma with mild cell swelling (Figure 1E), observed in 6 animals (26%) of the total analyzed (Table 1). In the DR 20 group, vacuolar degeneration was also visible (Figure 1E). The DMBA treatments also provoked glycogen overload, which was prominent in the midzonal region of the hepatic lobes. This overload was visible in affected animals of all three DMBA-treated groups. In the other animals of the three groups, no signs of pathology in the liver were observed (1, 5, and 2 animals for the DS 10, DS 20, and DR 20 groups, respectively).
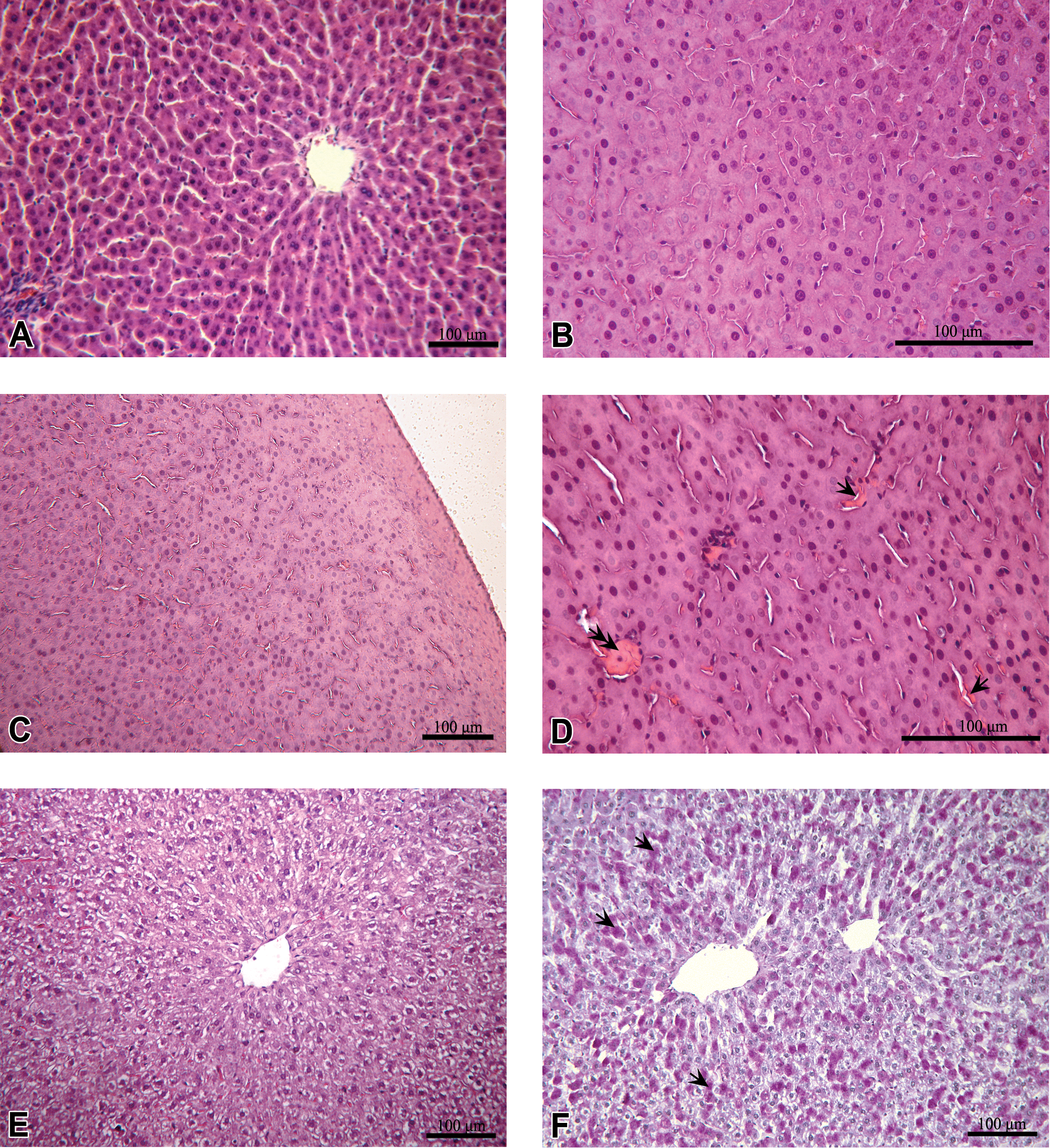
Histopathological effects of 7,12-dimethylbenz[a]anthracene (DMBA) treatments on the liver. (A) Normal liver tissue. (B) Glossy appearance of hepatocytes with heterochromatic nucleus examples (outlined in left part of the figure), visible in all groups with DMBA administered (DS 20 given as example). (C) Outlined, peripheral, subcapsular glossy appearance that is characterized for groups DS 10 and DS 20 (DS 10 given as example). (D) Coagulated erythrocytes as condensed conglomerates (arrow head) at visible thickened endothelial surface of sinusoids (arrows) (evident only in DR 20). (E) Disorganization of hepatic parenchyma with vacuolar degeneration in DR 20. (F) An increase in glycogen granules (visible as darker dots, examples indicated with arrows) distributed in a centrolobular, midzonal region (group DR 20). Hematoxylin and eosin staining for micrographs A–E, and periodic acid–Schiff staining for micrograph F. Scale bars are 100 μm, as indicated. Experimental groups of DMBA treatment (DS 10, DS 20, DR 20) are as described in the Methods section.
Histological abnormalities in liver in all groups after DMBA exposure (n [%]).

N indicates number of animals from each group analyzed with light microscopy; n indicates the number of rats with pathology described in the table. The percentages indicate how big the fraction of rats with pathological signs is compared to the total number of rats from the groups investigated in this respect.
Abbreviation: DMBA, 7,12-dimethylbenz[a]anthracene.
Accompanying the histopathological pathology, DMBA exposure resulted in considerable and significant reductions in liver GSH levels in all three DMBA-treated groups (Figure 2A). In particular, average GSH content was decreased by 79% for the DS 10 group, 77% for the DS 20 group, and 73% for the DR 20 group compared to control liver (Figure 2A). Significant enhancement in average activity of GSH reductase was observed in the DS 20 and DR 20 groups, compared to vehicle control (Figure 2A). Regarding GSH-reductase activity, the DS 10 group showed no difference with the vehicle control. Tissue activity of superoxide dismutase (SOD) remained unchanged following the administration of DMBA to all three experimental groups (DS 10, DS 20, DR 20; Figure 2A).
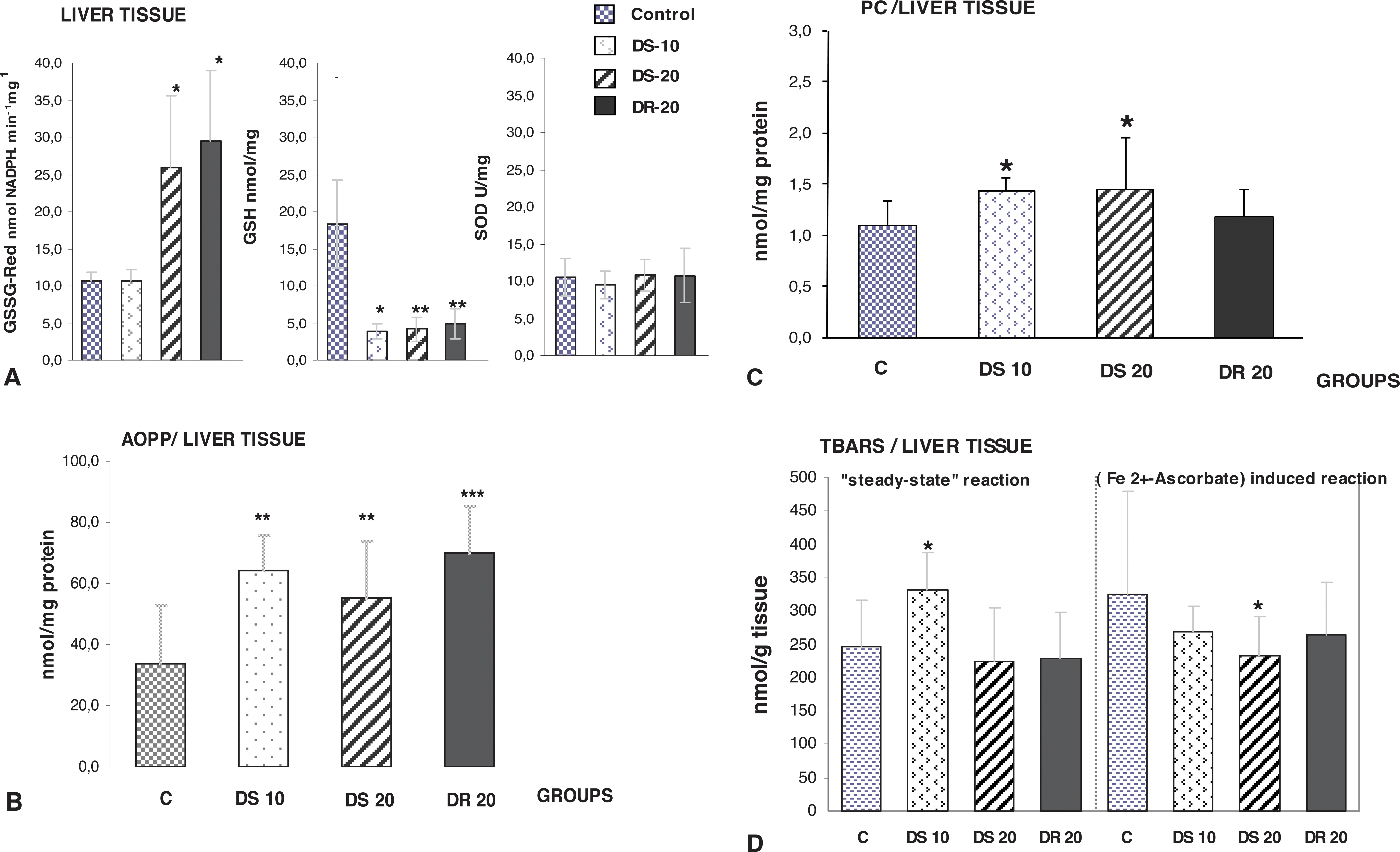
Oxidative stress assays in liver of 7,12-dimethylbenz[a]anthracene (DMBA)-exposed rats: (A) Significant increases in activity of glutathione reductase GSSG-Red, decreases in glutathione content GSH, and no change in superoxide dismutase SOD levels in DMBA-exposed rats compared to vehicle control rats. (B) Increases in advanced oxidation protein product levels in liver of DMBA-exposed rats compared to vehicle control rats. (C) Increases in protein carbonyl PC levels in liver of DMBA-exposed rats compared to vehicle control rats. (D) Increases in thiobarbituric acid reactive substance (TBAR) formation in liver homogenates of DMBA-exposed rats compared to vehicle control rats. On the left side, TBAR production nmol/g tissue in tissue sample steady-state reaction. On the right side, TBAR production nmol/g tissue after incubation with 4 × 10−5 M Fe2+ - 2.6 mM ascorbic acid induced lipid peroxidation. C, vehicle control. Experimental groups of DMBA treatment (DS 10, DS 20, DR 20) are as described in the Methods section. *p < .05 versus vehicle control; **p < .01 versus vehicle control.
Reminiscent of the effects on the GSH levels, all groups exposed to DMBA treatment displayed significantly increased average AOPP content in liver, compared to vehicle control (Figure 2B). In detail, this parameter of protein oxidation was enhanced in the DR 20 group by 106%, in the DS 20 group by 62%, and in the DS 10 group by 89%. Measurement of carbonyl residues indicative of oxidative protein damage showed that protein carbonyl content in the liver was moderately, but significantly, higher in the DS 10 (31%) and DS 20 (33%) groups, compared to vehicle control (Figure 2C). The animals of the DR 20 group did not show a significant change compared to the vehicle control.
Regarding TBAR production in liver (Figure 2D), we measured steady-state concentration, which corresponds to physiologic values of lipid peroxide formation (concentration in vivo). Only the DS 10 group showed a significant increase in steady-state levels of TBARs (35%) (Figure 2D, left panel). To further assess this finding, we assayed whether lipid oxidation could actually be activated in vitro. Inducible lipid peroxidation in vitro, as assayed with Fe2+ - acorbic acid was significantly lower only in the DS 20 group, by 31%, and unchanged in the other groups (Figure 2D, right panel).
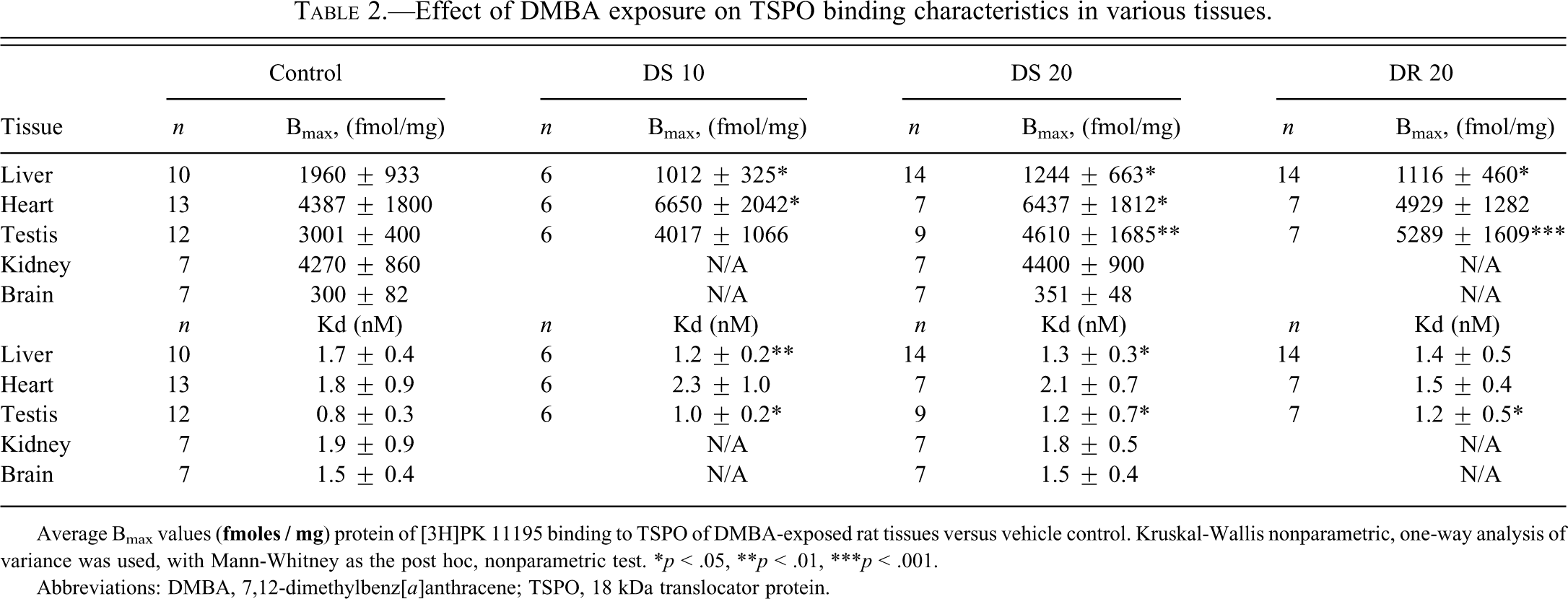
Binding assays with the TSPO-specific ligand [3H]PK 11195 were performed to determine potential effects of DMBA administration on TSPO binding characteristics in rat liver (Figure 3). In the liver of vehicle control rats, the Bmax and Kd values for TSPO were in the range of previously described results (Gavish et al. 1999). Representative Scatchard plots in liver presenting both Bmax and Kd are shown in Figure 3 for the control and experimental groups. The Bmax of [3H]PK 11195 was significantly lower in rat liver in all groups that were administered DMBA compared to vehicle (by 48%, 37%, and 43% for the DS 10, DS 20, and DR 20 groups, respectively, compared to vehicle control, Table 2).
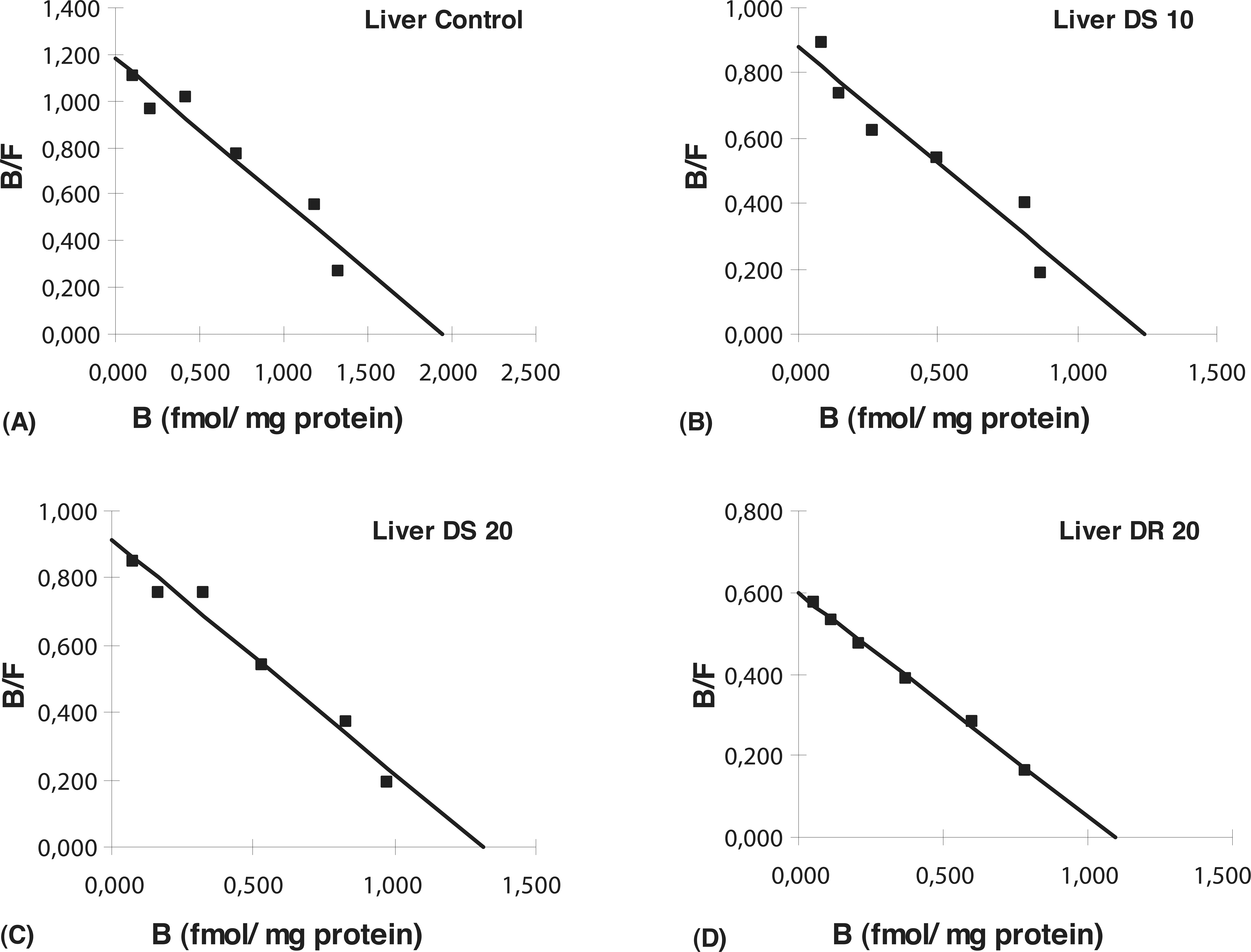
Representative Scatchard plots of saturation curves of [3H]PK 11195 binding to membrane homogenates of liver, respectively of vehicle control rats (A) and of rats exposed to DMBA (B–D). B, bound; B/F, bound over free; C, vehicle control. Experimental groups of DMBA treatment (DS 10, DS 20, DR 20) are as described in the Methods section. Explanation of further abbreviations as in Figure 1.
Effect of DMBA exposure on TSPO binding characteristics in various tissues.
Average Bmax values (
*p < .05
**p < .01
***p < .001.
Abbreviations: DMBA, 7,12-dimethylbenz[a]anthracene; TSPO, 18 kDa translocator protein.
Regarding tissues other than liver, compared to vehicle control, [3H]PK 11195 binding assays of the testis of our experimental groups showed a significant enhancement in the Bmax of TSPO resulting from DMBA administration, in two DMBA-treated groups, namely, DS 20 and DR 20 (54% and 76%, respectively), whereas for the DS 10 group no statistically significant increase was observed (Table 2). In homogenates from the heart, the Bmax for [3H]PK 11195 binding was also significantly higher in the groups with onetime application of DMBA (52% and 47%, respectively, for the DS 10 and DS 20 groups, compared to vehicle control), whereas no effects of DMBA treatment regarding TSPO binding characteristics were observed for the DR 20 group (Table 2).
Regarding tissues other than liver, heart, and testis, [3H]PK 11195 binding levels in kidney and brain appeared not to be affected by the DMBA. In particular, the binding analysis done on the DS 20 group showed no differences in binding capacity in kidney and brain homogenates, compared to vehicle control (Table 2). Kidney: Control group, Bmax = 4270 ± 860 fmol/mg and Kd = 1.9 ± 0.9 nM; DS 20 group Bmax = 4400 ± 900 fmol/mg and Kd = 1.8 ± 0.5 nM Brain: Control group Bmax = 300 ± 82 fmol/mg and Kd = 1.5 ± 0.4 nM and DS 20 group Bmax = 351 ± 48 fmol/mg and Kd = 1.5 ± 0.4 nM
Discussion
One part of this study was to assay potential consequences of dose, frequency of administration, and term from initial DMBA application on liver pathology, oxidative stress, and effects on TSPO binding characteristics in the liver. The repeated dose paradigm (fragmenting one large dose of DMBA to smaller, nonlethal fractions) is considered the optimal application model to induce malignant tumors (Gao et al. 2007; Huggins et al. 1966; Löscher et al. 1993; Mevissen et al. 1996). Depending on the particular report, term from administration of PAH to sacrifice ranges from 24 hours to 270 days. We used three regimens of DMBA application to outbred Wistar rats (DS 10, DS 20, and DR 20) and terms of 70 days (10 weeks) and 126 days (18 weeks) to determine the effect of DMBA per se, that is, under otherwise normal conditions. The different effects of the three regimens are discussed below.
This study served to determine potential correlations between DMBA exposure and oxidative stress in the liver, liver pathology, and TSPO binding characteristics in the liver. We paid special attention to the liver, since metabolic activation and detoxification of DMBA in vivo occur primarily in this organ (Moore et al. 1986; Muto et al. 2003). On the other hand, it is known that DMBA toxicity requires its conversion to 3,4-dyhydrodiol intermediates in the liver (Gao et al. 2007; Miyata et al. 2001). As determined with liver histopathology in the present study, the three DMBA treatments resulted in moderate toxic effects, comparable to cyclosporine-A–induced hepatotoxicity in rat liver (Rezzani et al. 2005). As another parameter of liver histopathology, it has been reported that drug toxicity, including PAH, may be accompanied by glycogen overload owing to disturbed carbohydrate metabolism (Muto et al. 2003; Pereira et al. 2006; Singh et al. 1997). Therefore, we also examined hepatocellular glycogen. It appeared that the rats that were subjected to DMBA exposure suffered from irreversible impairment in glycogen metabolism.
Studies by others applying PAH to outbred rodents described dose- and duration-dependent liver pathology, as we also found in this study (Table 1), including foci of altered hepatocytes, such as hepatocyte hypertrophy, multinucleated hepatocytes, inflammation, pigmentation, necrosis, and highly increased incidence of hepatocellular adenoma (Baker et al. 2001; Mishra et al. 2009; NTP technical report 2006; Safer and Al-Nughamish 1999; Tsuda and Farber 1980).
It has been suggested that possible mechanisms of hepatotoxicity include enhanced production of oxygen-derived free radicals resulting in depletion in intracellular reduced glutathione (Bironaite and Ollinger 1997; Stadlbauer et al. 2005). The present study shows a strongly reduced GSH levels in all three DMBA treated groups. This finding was accompanied by decreased GSH/GSSG-Red ratios in the DS 20 and DR 20 groups, indicating reduced capacity of the liver to scavenge free radicals produced in response to DMBA (Arulkumaran et al. 2007; Wu et al. 2004). It has been suggested that such a reduction in antioxidant capacity may also lead to a decrease in DMBA clearance (Muqbil and Banu 2006; Tsai-Turton et al. 2007). Our results demonstrated no differences in SOD levels following any of the DMBA treatments, which could simply mean that the potential DMBA effects are balanced out by compensatory responses (Valko et al. 2006).
Regarding the potential damaging effects of the oxidative stress in the liver, enhancement of AOPP levels was observed in all three DMBA treatment groups, and enhancement of protein carbonyls were observed in the DS 10 and DS 20 groups. Studies by others have suggested lipid oxidation in the liver resulting from DMBA exposure (Choi 2008; Mugbil and Banu 2006). We measured the profiles of the “steady-state” lipid peroxidation products (LPO) as TBARs. The DS 10 group displayed significantly increased TBAR values in liver, whereas the DS 20 and DR 20 groups did not show significant differences in this respect. To further assess this finding, we also assayed whether lipid oxidation could actually be activated in vitro. For this purpose, we assayed ferro-ascorbate pro-oxidant activity. Under these conditions, the DS 20 group showed decreased levels of TBARs, whereas no effects for the DS10 and DR20 groups were observed. Decreased content of highly unsaturated fatty acids, or modification of structure, composition, and stability of the membranes, as well as decreased concentration of NADPH, or changes in antioxidant status and protective enzymes, could be contributory factors to the lower rates of induced LPO (Cheesman et al. 1986; Kolanjiappan et al. 2003). This may also be the caase for our findings. In any case, in contrast to the effects on protein oxidation, lipid peroxidation does not appear to be a major effect of DMBA treatment in our paradigm.
Taken together, our data deriving from the GSH/GSSG-Red and AOPP suggest significant oxidative stress in the liver induced by acute exposure of rats to DMBA. This finding is consistent with previous reports that DMBA induces oxidative damage in liver in vivo characterized by induced depletion in liver glutathione content, and increased peroxidative damage (Arulkumaran et al. 2007; Choi and Kim 2009; Girolami et al. 2008; Letchoumy et al. 2006; Mishra 2009).
Unexpectedly, the oxidative stress assays regarding the liver, other than GSH and AOPP applied in this experiment (e.g., SOD, protein carbonylation, lipid peroxidation), provided less conclusive results than the GSH and AOPP data, suggesting that DMBA may have specific oxidative stress effects in the liver with the regimens we applied, as opposed to more general toxic effects. No statistical differences were observed in liver LPO measurements in the DS 20 and DR 20 groups, and also not in protein carbonyls in the DR 20 group, which suggests that the liver is able to deal with the induced stress by DMBA. In another study, we applied a high-fat, high-cholesterol (HFHC) diet to rats to evaluate oxidative stress in the liver (Dimitrova-Shumkovska et al. 2010b). In that study, we found not only enhanced protein oxidation but also enhanced lipid peroxidation in the rats' livers. In fact, all parameters indicated enhanced oxidative stress in the rats' livers. As the HFHC diet was chronic, whereas the DMBA exposures of the present study were acute, either the insult of DMBA to the liver was less severe than the insult caused by the HFHC diet, or the liver recovered from the DMBA treatment with regard to lipid peroxidation (LPO) in the time period after the DMBA application. Interestingly, in this respect, although the DS 20 and DR 20 groups were sacrificed 18 weeks after DMBA exposure and showed no effects with the TBAR steady-state assay, the DS 10 group was sacrificed only 10 weeks after DMBA treatment and (still) showed a significant increase with the TBAR steady-state assay of the liver. This finding may suggest that 18 weeks allows for at least a partial recovery from oxidative stress, whereas 10 weeks does not allow for such a recovery. As described above, the obtained results regarding LPO could not fully reflect a picture about levels of liver injury (Fang and Lin 2008). It would be interesting to study other, shorter periods after DMBA exposure, to see whether in such paradigms higher levels of oxidative stress in the liver are detected.
In addition to above mentioned effects on liver toxicity, which are in accord with other studies, our results showed significant decreases in TSPO binding capacity in the livers of all groups given DMBA (Figure 3). In particular, these novel findings of changes in TSPO binding capacity in the liver after DMBA exposure correlated well with (1) increased protein oxidation (AOPP), (2) decreased GSH levels, and (3) histopathological signs. It has been suggested previously that TSPO binding characteristics, such as affinity, may correlate with oxidative stress in the liver (Courtiere et al. 1995). As the TSPO binding capacities in the liver are typically reduced in the present study, this may be a compensatory response directed toward reducing the damaging effects of DMBA. Such a mechanism has been proposed with in vitro studies with the toxin CoCl2 (Zeno et al. 2009). This conclusion is based on studies in which reductions in TSPO levels via TSPO knockdown by genetic manipulation in vitro resulted in reduced cell death levels, and reduced ROS generation (Levin et al. 2005; Shoukrun et al. 2008; Veenman et al. in press; Zeno et al. 2009). Presently, we cannot definitely conclude whether the reduced TSPO binding capacity is a physiological response to the toxin DMBA and/or oxidative stress induced by DMBA, or that reduction in TSPO binding capacity in the liver is part of damaging effects of DMBA.
Since TSPO appears to control cholesterol transport and modulate steroidogenesis in endocrine tissues (Papadopoulous et al. 1997; Veenman et al. 2007), we also assayed expression of TSPO in testis homogenate of DMBA-treated rats. A significant enhancement in TSPO expression was observed in the testes of DS 20 and DR 20 rats. In another study regarding glandular tissue, with DMBA applied to female Sprague Dawley rats (80 mg/kg, i.e., comparable to our DS 20 and DR 20 groups), a significant increase in TSPO capacity in the nucleus of developed breast tumors compared to control tissue was observed (Mukhopadhyay et al. 2006). Our data and studies of others may allow us to hypothesize that stress induced by DMBA exposure may result in enhanced TSPO binding in glandular and steroidogenic tissues, potentially leading to tumorigenic developments. In parallel, the TSPO response in heart tissue was also significantly higher in the groups with onetime application of DMBA (52% and 47%, respectively, for the DS 10 and DS 20 groups, compared to vehicle control), whereas no effects of DMBA treatment regarding TSPO binding characteristics were observed for the DR 20 group. It is well known that TSPO responds to ischemia (Veenman and Gavish 2006). Thus, we hypothesize that lesions in the aorta observed after DMBA treatment in rats (Dimitrova-Shumkovska et al. 2010a) may contribute to the increase in TSPO binding capacity observed in the heart in the present study.
In brain and kidney, binding assays of kidney and brain in the DS 20 group did not reveal differences with the control group. We did not do assays of the other groups because we thought there would be no difference. In any case, only in the liver and the testes do the changes appear to be consistent.
As in the different organs, TSPO binding capacity may either increase, decrease, or not change, suggesting that TSPO responses to DMBA exposure in the liver and some other organs are organ specific rather than part of a uniform general body response. Thus, the observed TSPO responses in the various organs may be of a physiological, functional nature rather than simply part of the damage caused by DMBA. More studies are needed to verify this hypothesis for the individual organs.
In conclusion, our exposure of rats to DMBA caused oxidative stress in liver, as well as pathological changes in this organ. The pathological changes appeared to be related to dose or duration of DMBA exposure, and they were most consistently elevated upon repeated doses of DMBA totaling 20 mg. The binding capacity of TSPO was decreased in liver after DMBA treatment in correlation with indicators of oxidative stress in this organ. In other organs, such as heart and testis, TSPO binding was enhanced, or not affected (e.g., brain and kidney), suggesting that the effect of DMBA on TSPO binding in liver is tissue specific. We suggest that the reductions in TSPO binding capacity may present protective mechanisms. For example, reductions in ROS generation and cell death may follow reductions in TSPO binding capacity, as demonstrated previously with in vitro studies (Zeno et al. 2009).
Footnotes
Acknowledgments
L.V. and M.G. received support from internal Technion Research Funds: Elias Medical Research Fund; Lazarov Research Fund; K. Rozenblatt Research Fund; L. Aronberg Research Fund in Neurology; Trauma Research Fund; Fund for Interdisciplinary Research and Collaboration; Daniel Horovitz Fund–Secondary Brain Damage and Neurodegeneration; Atkins Fund–Gerontology; Alexander Goldberg Memorial Research Fund; Nattie Fisher Alzheimer Research Fund; Jessie Kaplan Research Fund, Alzheimer’s. The Center for Absorption in Science, Ministry of Immigrant Absorption, State of Israel, is acknowledged for their support to S.L. and L.V. Presently, L.V. is supported by a joint grant from the Center for Absorption in Science of the Ministry of Immigrant Absorption and the Committee for Planning and Budgeting of the Council of Higher Education under the framework of the KAMEA program, Israel. We gratefully thank Radmila Crceva Nikolovska, Svetlana Stojanovska and Marietta Sthakleva for their support with the technical work. This work was further made possible by the continuous and generous support, advice, and encouragement of Mr. Gradimir Shumkovski.
The authors declare no conflict of interest.