
Abstract
Drug-induced valvulopathy is a serious liability for certain compound classes in development and for some marketed drugs intended for human use. Reports of valvulopathy led to the withdrawal of fenfluramines (anorexigens) and pergolide (antiparkinson drug) from the United States market in 1997 and 2007, respectively. The mechanism responsible for the pathogenesis of valvulopathy by these drugs is likely a result of an “off-target” effect via activation of 5-hydroxytryptamine (5-HT) 2B receptor (5-HT2BR) expressed on heart valve leaflets. Microscopically, the affected valve leaflets showed plaques of proliferative myofibroblasts in an abundant extracellular matrix, composed primarily of glycosaminoglycans. However, the valvular effects caused by fenfluramines and pergolide were not initially predicted from routine preclinical toxicity studies, and to date there are no specific validated animal models or preclinical/toxicologic screens to accurately predict drug-induced valvulopathy. This review covers the structure and function of heart valves and highlights major advances toward understanding the 5-HT2BR-mediated pathogenesis of the lesion and subsequently, development of appropriate animal models using novel techniques/experiments, use of functional screens against 5-HT2BR, and more consistent sampling and pathologic evaluation of valves in preclinical studies that will aid in avoidance of future drug-induced valvulopathy in humans.
Keywords
Heart Valves: Structure and Functional Histology
Normal semilunar (i.e., aortic and pulmonary) and atrioventricular (i.e., mitral and tricuspid) heart valves play an important role in maintaining unidirectional blood flow through the cardiac chambers of the heart, and this function requires not only structural integrity, but also coordinated interactions among several critical valvular components (i.e., for aortic and pulmonary valves, cusps/leaflets, commissures, and their respective supporting structures [roots]; and for mitral and tricuspid valves, leaflets, commissures, annulus, chordae tendineae, papillary muscles, and atrial and ventricular myocardium). In humans, semilunar valve (SV) leaflets are lined with endothelium and have three histologically well-defined tissue layers: (1) the ventricularis, facing the inflow surface, composed primarily of collagen and elastic fibers, provides for elasticity when the cusp changes shape during the cardiac cycle; (2) the subjacent spongiosa, composed of loose collagen and abundant proteoglycans, provides a cushion for physical forces; and (3) the fibrosa, facing the outflow surface, consisting predominantly of dense collagen fibers, provides strength (Figure 1 ). Additionally, a poorly demarcated fourth layer called the aortalis (facing the aortic surface), composed of scant collagen and elastin, has been described in selected publications (Schoen and Edwards 2001; Schoen 2005a; Schoen 2005b; Schoen 2006). The atrioventricular valve (AV) leaflets are structurally similar to those of the SV and have analogous components: atrialis (also known as auricularis), spongiosa, fibrosa, and ventricularis (from inflow to outflow, respectively). Both the AV and the SV are innervated by adrenergic and cholinergic neural networks (Marron et al. 1996).
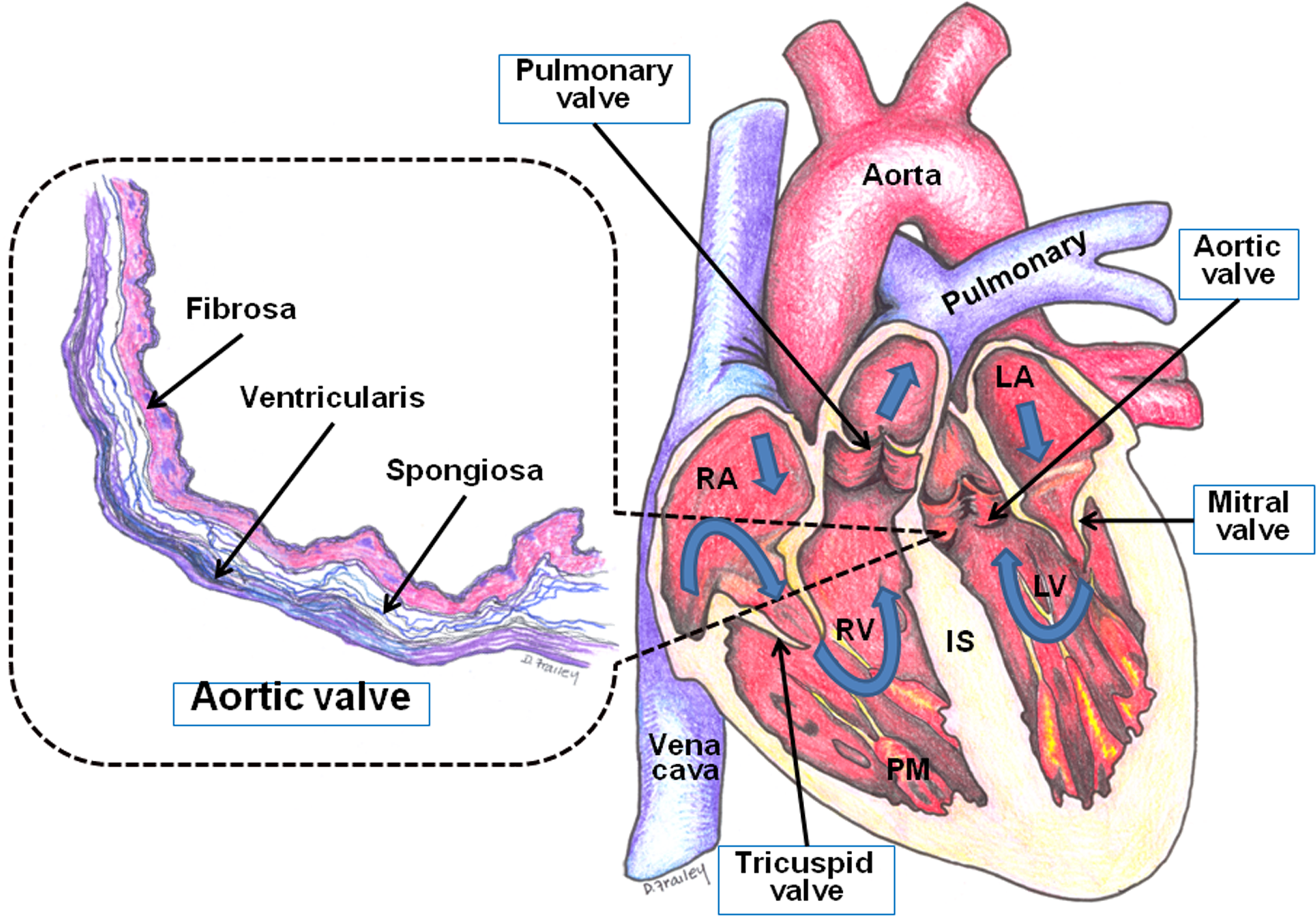
Systematic representation of heart chambers, valves, and valvular histology. Blue arrows indicate unidirectional blood flow through the various chambers of the heart. Histology of aortic valves, mimicking Elastic Van Giesen stain (collagen, red; elastic fibers, black). Three well-defined layers: (1) fibrosa, facing the outflow surface, consisting predominantly of dense collagen fibers; (2) spongiosa, composed of loose collagen and abundant proteoglycans (clear spaces); and (3) ventricularis, facing the inflow surface, composed of primarily collagen and elastic fibers. LA, left atrium; LV, left ventricle; RA, right atrium; RV, right ventricle; IS, interventricular septum; and PM, papillary muscles.
Valvular Cells
Two types of cells are present in fully formed heart valves: valvular endothelial cells (VECs), covering all surfaces of the valve leaflet, and valvular interstitial cells (VICs), located deep below the surface. It has been shown that VECs express different transcriptional profiles and respond differently to injury at various regions of the valve (e.g., aortic side vs. ventricular side) (Simmons et al. 2005). For example, VECs in normal aortic valves of adult pigs have been shown to differentially express 584 genes on the aortic side versus the ventricular side (Simmons et al. 2005). Several of these observed differences could help explain the vulnerability of the aortic side of the valve cusp to calcification in disease (Simmons et al. 2005). Valvular endothelial cells also show phenotypic differences in response to in vitro shear when compared with vascular endothelial cells (Butcher et al. 2006). However, reasons for and implications of these differences are not yet known.
Valvular interstitial cells, the most prevalent cells in all three valvular layers, maintain the integrity and stability of normal valves and regulate repair processes during disease and following valve injury (Messier et al. 1994; Mulholland and Gotlieb 1996; Mulholland and Gotlieb 1997; Rabkin et al. 2001). Valvular interstitial cells have morphologic and functional features characteristic of fibroblasts, smooth muscle cells, and myofibroblasts (Schoen 2006), and they express muscle structural genes, including the cardiac isoforms of troponin-T, -I, and -C; β-myosin heavy chain; and cardiac myosin light chain 2 (Roy et al. 2000). Approximately 57% of VICs in human valve leaflets express α-smooth muscle actin (α-SMA, a marker for distinguishing smooth muscle cells and myofibroblasts from fibroblasts). In vitro, however, nearly 80% of VICs stain for α-SMA (Taylor et al. 2000). Valvular interstitial cells have specific features determined at the time of embryonic endothelial-to-mesenchymal transformation, and therefore, the term “valve fibroblasts” should be abandoned and replaced by the term “VICs” (Liu et al. 2007). The valves were previously thought to be composed of a phenotypically homogeneous population of VICs, but now there are currently five identifiable phenotypes of VICs: (1) embryonic progenitor endothelial/mesenchymal cells; (2) quiescent VICs (qVICs); (3) activated VICs (aVICs); (4) progenitor VICs (pVICs); and (5) osteoblastic VICs (obVICs) (Liu et al. 2007). These phenotypes may exhibit plasticity and convert from one form to another. The embryonic progenitor endothelial/mesenchymal cells represent the cells that are derived embryonically through endothelial-to-mesenchymal transformation, which initiates the process of valve formation in the embryo (Norris et al. 2008). Quiescent VICs are the fibroblast-type cells in the adult valve and maintain normal valve physiology. During valvular injury, qVICs become aVICs, express α-SMA, and become involved in the repair and remodeling of valves (Aikawa et al. 2006; Cushing et al. 2005; Liu and Gotlieb 2008; Walker et al. 2004). These cells are a type of myofibroblast cell similar to those found at sites of wound repair in a variety of tissues (Desmouliere et al. 2005; Hinz et al. 2007). The pVICs are poorly defined and consist of a heterogeneous population of progenitor cells that may be important in repair (Visconti et al. 2006). In conditions promoting calcification, such as the presence of osteogenic and chondrogenic factors, qVICs can undergo osteoblastic differentiation into obVICs (Chen et al. 2009).
Valvular Extracellular Matrix
The primary components of valvular extracellular matrix (ECM) are proteoglycans (PGs), collagens, and elastin. The overwhelming determinant of valve durability is valvular ECM, produced and maintained by VICs. The quantity and quality of valvular ECM, therefore, depend on the viability and function of VICs. Proteoglycans are integral ECM components of valves and play an important role in mediating ECM organization (Kinsella et al. 2004). Proteoglycans consist of glycosaminoglycan (GAG) chains (linear molecules of repeating disaccharides) attached to a core protein. Depending on biological and biomechanical needs, the sulfation pattern of GAG chains (i.e., whether 4-sulfated or 6-sulfated) varies. 4-Sulfated GAGs (i.e., small leucine-rich dermatan sulfate PGs), such as decorin and biglycan, are abundant in tensile loading regions such as chordae tendineae; these PGs mediate collagen fibrillogenesis (Reed and Iozzo 2002) and sequester transforming growth factor-β (Kresse and Schonherr 2001). Glycosaminoglycan hyaluronan and 6-sulfated GAGs (versican) are found in regions experiencing compression, such as the leaflet free edges; versican often co-localizes with elastic fibers and numerous cell-associated molecules to regulate cell adhesion, proliferation, and migration (Grande-Allen et al. 2004; Wight et al. 1991; Wight 2002). Additionally, valves contain collagen types I, III, and IV; elastin; fibronectin (ECM protein); and tenascin-C (ECM glycoprotein) (Fayet et al. 2007).
Valvular interstitial cell–regulated repair involves synthesis, degradation, and reorganization of ECM, which depends on matrix-degrading enzymes such as matrix metalloproteinases (MMPs). Matrix metalloproteinases are calcium- and zinc-dependent proteases that are produced and secreted by VICs as inactive zymogens. Matrix metalloproteinases are activated by plasmin, interleukin-1β, and tumor necrosis factor-α, and they are inhibited by four types of specific tissue inhibitors of metalloproteinases (TIMPs) through the formation of irreversible complexes (Li et al. 2000; Visse and Nagase 2003). TIMP-1, -2, and -4 are present in soluble form, whereas TIMP-3 binds to the ECM through heparan sulphate proteoglycans (Kassiri and Khokha 2005). Matrix metalloproteinases and TIMPs play an important role in the physiological maintenance of ECM, as well as in the pathogenesis of a variety of diseases (Li et al. 2000; Visse and Nagase 2003). Extracellular matrix changes occur in many heart valve pathologies, including myxomatous mitral valve disease (Grande-Allen et al. 2004; Gupta et al. 2008; Mahimkar et al. 2009) and drug-induced valvulopathy in humans and animals (Droogmans, Cosyns et al. 2007; Droogmans, Franken et al. 2007; Elangbam et al. 2006; Elangbam et al. 2008; Gustafsson et al. 2005; McDonald et al. 2002).
Assessment of Heart Valves in Toxicologic Pathology
In veterinary medicine, valvular histology has been inconsistently described in recent editions of histology/pathology textbooks; that is, it is described as three or four well-defined tissue layers as in humans, with no mention of specific layers, but characterized as a central core of collagen fibers, reinforced by thin layers of elastic fibers (Aupperle, Marz et al. 2009; Aupperle, Thielebein et al. 2009; Aupperle et al. 2010; Maxie and Robinson 2007; Plendl 2006; Van Vleet et al. 2002). Among the preclinical species for drug safety screening, gross evaluation of valves and associated structures is routinely performed in nonrodents (dogs, primates), including sampling and characterization of changes and location of affected areas for further histopathological evaluation. In contrast, rodent hearts are routinely fixed intact in 10% neutral buffered formalin, and heart valves are not grossly examined. Inconsistencies in rodent heart-valve sampling are often unavoidable because of the inherent small heart size, thin valve leaflets, and the various trimming/sectioning methods used. For most standard rodent studies, the heart is sectioned through its longitudinal axis to include one or ideally both the right and left ventricles, interventricular septum, portions of both atria, and major blood vessels at the base of the heart. As a result, some sections may contain one or more valves, whereas others may not contain any. Review of nine long-term rat studies (four from Charles River, Montreal and five from the National Toxicology Program [NTP]) revealed a substantial variability in the presence of individual valves in heart sections, and some heart sections had no valves (Elangbam, unpublished data). Furthermore, even in the presence of prominent valvulopathy, no alteration in the valves was described and recorded in any of these studies (Elangbam et al. 2006). There is also a paucity of published reports on age-dependent valvular changes and drug-induced valvular disease in rodents, and most importantly, chemically exacerbated spontaneous valvulopathy has not been fully explored. For example, although valvular changes were not described and recorded in the original NTP study report with dl-amphetamine in Fischer 344 (F344) rats (NTP 1991), subsequent re-evaluation of hearts from this study found a treatment-related exacerbation of spontaneous valvulopathy (Elangbam et al. 2006). Therefore, unless specified for a particular rodent study, it is not customary or mandatory practice to examine and record heart valve findings, particularly in two-year studies. As a result, the valvular findings in rodents may have been underdiagnosed and underreported, mainly because of inevitable inconsistency of histological sampling and inconsistency in reporting.
Drug-induced Valvulopathy in Humans
Valvular abnormality from any cause can lead to hemodynamic overload on ventricles (left or right ventricle, or both ventricles), and this overload eventually leads to myocardial dysfunction, congestive heart failure, and sudden death in humans. Even a subtle, nondestructive valvular thickening may result in leaflet restriction and regurgitation (Carabello and Crawford 1997). Valvular disease, or valvulopathy, is a main cause of morbidity and mortality in humans worldwide, requiring valve replacement surgery, the second most common heart operation performed in the United States (Keane et al. 1993; Thom et al. 2006). Four drug classes that have been linked to valvulopathy in humans are described below.
Anorexigens (fenfluramine, dexfenfluramine)—Appetite Suppressants
Anti-obesity drugs, such as fenfluramine and dexfenfluramine (stereoisomer of fenfluramine), have been linked to valvulopathy in humans (Connolly et al. 1997; Dahl et al. 2008; Greffe et al. 2007; Hershel 2000; Jick et al. 1998). Fenfluramine, dexfenfluramine, and their major metabolites norfenfluramines stimulate 5-hydroxytryptamine (5-HT, serotonin) release from neuronal and platelet stores (Fishman 1999; Yanovski and Yanovski 2002). Connolly et al. (1997) first reported twenty-four cases of valvulopathy in women who had received treatment with the fenfluramine and phentermine combination (“Fen-Phen”) for a mean duration of eleven months. Five of these patients subsequently underwent cardiac surgery and valve replacement. Microscopic examination of the valve leaflets showed “stuck-on” plaques of proliferative myofibroblasts in an abundant extracellular matrix. A subsequent larger study with valve samples from sixty-four patients described an “atypical plaque” containing myxoid stroma, proliferative myofibroblasts, and often small vessels and lymphocytic infiltrations (Steffee et al. 1999; Volmar and Hutchins 2001). These valvular changes resembled lesions associated with carcinoid heart disease (valvular disease in patients with carcinoid tumors of the gastrointestinal tract that have metastasized to the liver) or ergotamine-induced valvulopathy. Prevalence varies considerably between studies, ranging from approximately 6% to just over 30% (Bowen et al. 1997). One case-control study reported a prevalence of 23% in patients treated with fenfluramine. A meta-analysis of the published studies of patients treated with fenfluramine estimated that one in eight patients receiving these agents for more than ninety days had valvulopathy (Sachdev et al. 2002). It has also been shown that the duration of treatment played a role in the development of clinical valvulopathy, namely, that low prevalence rates are associated with shorter durations of therapy, and higher prevalence rates are associated with longer exposures (Jollis et al. 2000). In September 1997, both fenfluramine and dexfenfluramine were withdrawn from the United States (U.S.) market, including a multibillion-dollar settlement by the manufacturer of these drugs (SoRelle 1999). Recently, several cases of fenfluramine-induced valvulopathy requiring surgery have been identified, seven years after discontinuing the medication, suggesting that anorexigen-induced valvulopathy may continue to manifest well into the future (Dahl et al. 2008; Greffee et al. 2007).
Dopamine Agonists (pergolide, cabergoline)—Parkinson’s Disease and Hyperprolactinemic Disorders
Over the past three decades, there has been widespread use of ergot-derived dopamine agonists (pergolide and cabergoline) to treat Parkinson’s disease, restless leg syndrome, and hyperprolactinemic disorders. An association between valvulopathy and pergolide was first reported in 2002 (Pritchett et al. 2002), and this association is now widely accepted. Typically, the morphological and histological features of pergolide-related valvulopathy resemble fenfluramine- and ergotamine-induced valvulopathy and carcinoid heart disease (Baseman et al. 2004). Recently, two large European studies independently verified the association of valvulopathy with pergolide and cabergoline. In the first study, Zanettini et al. (2007) reported moderate or severe valvular regurgitation with a significantly greater frequency in patients given pergolide (23.4%) or cabergoline (28.65%) than in those given non-ergot dopamine agonists such as pramipexole and ropinirole (0%) or controls (5.6%). In the second study, Schade et al. (2007) concluded that the incidence of heart valve regurgitation was increased in pergolide (7.1-fold) and cabergoline (4.9-fold) groups, but not with use of other dopamine agonists such as ropinirole, pramipexole, bromocriptine, or lisuride. This study also noted that the severity of valvular disease was associated with higher cumulative doses of either cabergoline or pergolide. Similarly, a recent systematic literature review on valvulopathy in Parkinson’s disease patients showed that in eleven of thirteen studies, the use of cabergoline and pergolide was associated with significantly increased frequency of heart valve disease compared with non-ergot group and controls (Steiger et al. 2009). Pergolide was withdrawn from the U.S. market in August 2007 because of increased risk for valvulopathy. Although the European Medicines Agency has added new warnings and contraindications to the product information, cabergoline and pergolide remain in use in the rest of the world (Antonini and Poewe 2007; EMEA 2008).
Ergot Alkaloids (methysergide, ergotamine)—Migraine Prophylaxis
Migraine is a complex, common, and disabling disorder of the brain that occurs in 34.5% of adult women and 20.1% of adult men in the general population (Bigal et al. 2006). Ergot alkaloid drugs include methysergide and ergotamine, which are very effective drugs for migraine headache prophylaxis. In 1967, Graham reported the development of valvular insufficiency and retroperitoneal fibrosis in twenty-seven patients taking methysergide. Although initially reported as left-sided valve effect, autopsy studies and case reports showed tricuspid valve involvement (Bana et al. 1974; Hendrikx et al. 1996; Misch 1974). Furthermore, several patients developed dyspnea and eventually required heart valve replacement. Ergotamine, an ergot alkaloid structurally similar to methysergide, has been shown to cause left-sided valvulopathy primarily affecting the mitral valve. Ergotamine- or methysergide-associated valvulopathy is morphologically similar to that seen in carcinoid heart disease (Hauck et al. 1990; Redfield et al. 1992; Van Camp et al. 2004). Although rarely used now because of their adverse effects, methysergide and ergotamine remain licensed for migraine prophylaxis.
Recreational Drug: 3,4-Methylenedioxymethamphetamine (Ecstasy)
3,4-Methylenedioxymethamphetamine (MDMA), an amphetamine-based drug, is a psychoactive stimulant used for recreational purposes. Use of MDMA has increased over the past two decades and has become one of the most popular party drugs in Western Europe and the United States (EMCDDA 2005; Martins et al. 2005). Setola et al. (2003) first reported that MDMA and its metabolite 3,4-dimethoxyamphetamine (MDA) are potent 5-HT2BR agonists, and these drugs elicited prolonged mitogenic responses in human VICs in vitro. As predicted by Setola et al. (2003), Droogmans, Cosyns et al. (2007) reported the development of significant valvular regurgitation in eight (28%) people (mean age of 24.3 years) who took MDMA (average of 3.6 tablets of MDMA per week for six years) compared with none in the control group. Analysis of valve morphology identified restrictive valvulopathy similar to that seen in patients taking pergolide or cabergoline. The investigators also noted a correlation between dose and severity of valvulopathy. Most MDMA use is intermittent and recreational, but in heavy and frequent MDMA users, an increased risk of valvulopathy seems probable. 3,4-Methylenedioxymethamphetamine is currently being evaluated as a treatment for post-traumatic stress disorder.
Molecular Mechanism of Drug-induced Valvulopathy
Pharmacologically, anorexigens (fenfluramine, dexfenfluramine, and their active metabolite norfenfluramine), dopamine agonists (pergolide and cabergoline), amphetamines (MDMA and its metabolite MDA), and ergot alkaloids (ergotamine, dihydroergotamine, and methysergide and its active metabolite methylergonovine) are potent 5-HT2B receptor (5-HT2BR) agonists (Table 1 ). Therefore, the mechanism responsible for the pathogenesis of valvulopathy by these drugs is likely a result of an “off-target” effect via activation of 5-HT2BR expressed on heart valve leaflets (Fitzgerald et al. 2000; Roth 2007; Rothman and Baumann 2009; Rothman et al. 2000; Setola et al. 2003). The critical role of 5-HT2BR activation in the pathogenesis of valvulopathy is further highlighted by the observation that chemically similar drugs, such as lisuride and terguride, which are selective agonists for 5-HT2C and 5-HT2A receptors and antagonists for 5-HT2BR, were not associated with valvulopathy in humans (Jahnichen et al. 2005; Roth 2007). Bromocriptine and quinagolide have a weaker affinity for the 5-HT2BR and, therefore, are thought to be less valvulopathic. However, evidence from a relatively small study suggests that high doses of bromocriptine are associated with an increased risk of valvulopathy in patients with Parkinson’s disease (Antonini and Poewe 2007; Serratrice et al. 2002). Similarly, no increase in the risk of valvulopathy was observed in patients treated with the dopamine agonist pramipexole, which has low affinity to the human 5-HT2BR (Mulholland and Gotlieb 1997; Millan et al. 2002; Schade et al. 2007; Zanettini et al. 2007).
Drugs with 5-HT2BR functional profiles and their association with valvulopathy in humans.

There are seven distinct families of 5-HT receptors (5-HT1-7) and several subtypes (Nichols and Nichols 2008). However, only 5-HT2BR has been implicated in valvulopathy because of its high concentration in human heart valves and pulmonary arteries (Fitzgerald et al. 2000; Rothman et al. 2000; Roth 2007). Recent studies have implicated preferential activation of 5-HT2BR as a key initiating step in the pathogenesis of anorexigen-induced valvulopathy in humans. 5-Hydroxytryptamine has a direct mitogenic effect on VICs, and this mitogenic effect is mediated by 5-HT receptors (Fanburg and Lee 1997; Lopez-Ilasaca 1998; Rajamannan et al. 2001). 5-Hydroxytryptamine 2B receptors are G protein coupled and known to activate mitogenic pathways through the phosphorylation of Src kinase and extracellular-regulated kinases (ERK) (Roth 2007). Activation of 5-HT2B receptors also leads to activation of phospholipase C-β and subsequent activation of protein kinase C. 5-Hydroxytryptamine and Src-P may mediate and enhance the activity of the transforming growth factor β1 receptor (also known as activin receptor–like kinase, ALK-5) (Ignotz and Massague 1986; Jian et al. 2002; Roth 2007; Waltenberger et al. 1993). Once activated, ALK-5 subsequently phosphorylates the major downstream signaling molecules SMAD2 and 3 proteins. Phosphorylated SMAD2 and 3 form a complex with SMAD4, and this complex translocates into the nucleus and regulates the transcription of specific genes involved in cell growth, differentiation, development, and immune response (Gordon and Blobe 2008; Heldin et al. 1997; Liu and Gotlieb 2008; Massague et al. 2005) (Figure 2). It has also been suggested that the final common pathway by which activation of 5-HT2B receptors leads to mitogenesis probably involves phosphorylation of retinoblastoma protein (Roth 2007).
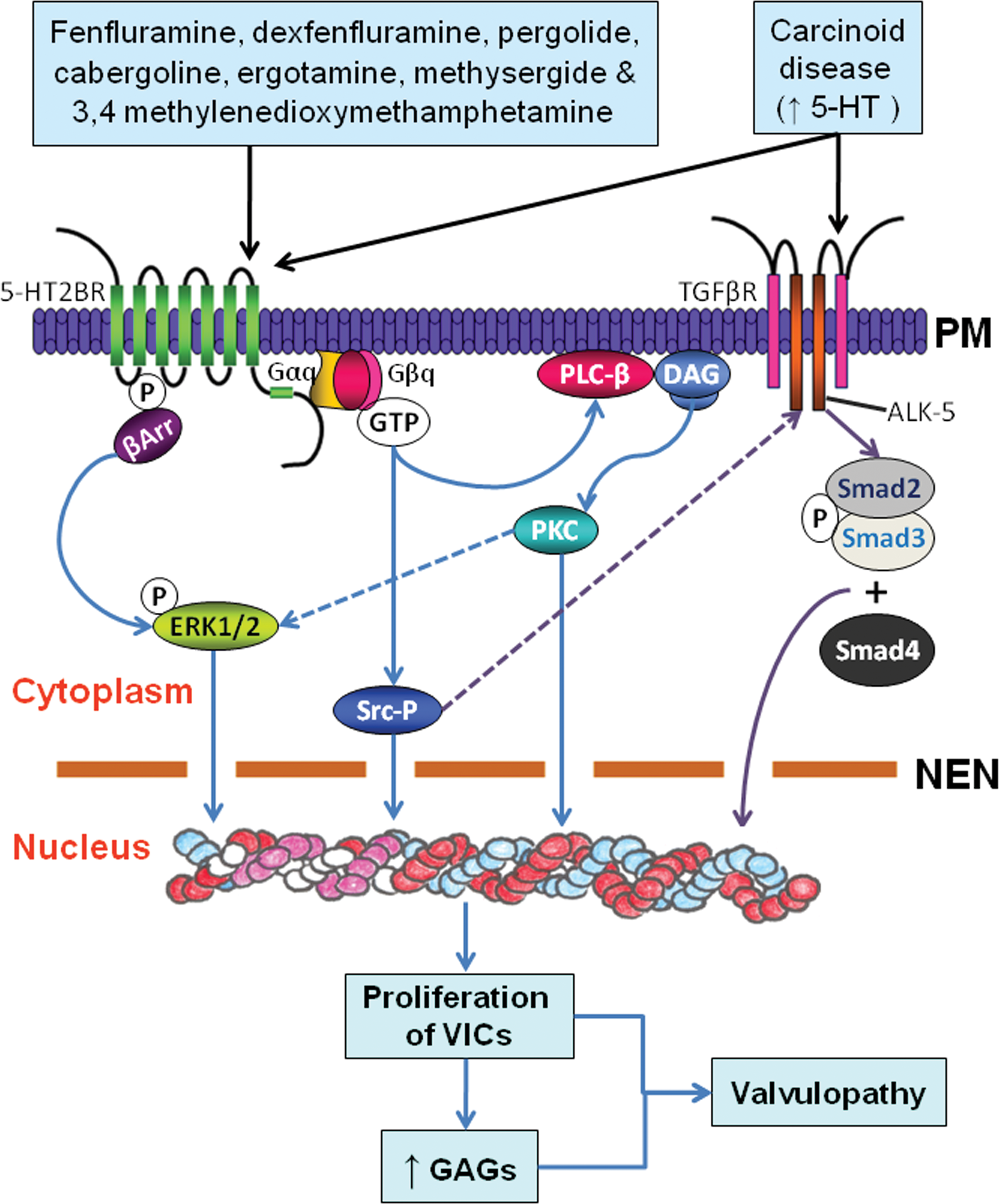
Schematic representation of 5HT signaling pathways involved in the pathogenesis of drug-induced valvulopathy by 5-HT2BR agonists (fenfluramine, dexfenfluramine, pergolide, cabergoline, 3,4-methylenedioxymethamphetamine, ergotamine, and methylsergide). Activation of 5-HT2BR by these drugs causes dissociation of guanine nucleotide regulatory proteins (G-proteins), which leads to activation of protein kinase C (PKC) via activated phospholipase C-β (PLC-β) resulting from intracellular calcium mobilization and diacylglycerol (DAG) release. G-proteins may also cause phosphorylation and activation of Src with the formation of phosphorylated Src (Src-P) and activation of extracellular regulated kinases 1 and 2 (ERK1 and ERK2), facilitated by β-arrestin (βArr) binding. Extracellular regulated kinases 1 and 2 are also activated by PKC. Additionally, 5-HT and possibly Src-protein enhance the activity of TGFβ receptors. Activation of TGFβ receptor 1 (also known as activin-like kinase-5, or ALK-5) leads to phosphorylation of SMAD 2 and 3 proteins and formation of a complex with SMAD4; this complex translocates to the nucleus and regulates the transcription of specific target genes. All of these pathways lead to proliferation of aVICs, matrix secretion/accumulation, and subsequent valvulopathy. PM, plasma membrane; NEN, nuclear envelope.
In addition to transcriptionally controlled proliferative effects on VICs, there is also evidence of oxidative stress (i.e., generation of reactive superoxide, O2•–) in heart valves by serotonin and dopamine in vitro (Pena-Silva et al. 2009). Increased O2•– levels in homogenates of heart valves and blood vessels after incubation with serotonin and dopamine were prevented by inhibitors of flavin-oxidases (diphenyliodonium) or monoamine oxidase (MAO) (ranylcypromine and/or clorgyline), but not by inhibitors of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, suggesting a novel mechanism for MAO’s role in oxidative stress in human heart valves by serotonin and dopamine (Pena-Silva et al. 2009). Another study by Connolly et al. (2009) showed a significant imbalance of ECM production versus proliferation in mitral VIC cultures with fenfluramine plus 5-HT exposure, suggesting that this imbalance may in part explain the pathophysiology of fenfluramine-induced valvulopathy. Because of the clear association between 5-HT2BR agonism and drug-induced valvulopathy, it has been strongly recommended to screen future drugs with 5-HT activity and their metabolites at the 5-HT2BR comprehensively before launching clinical trials (Huang et al. 2009; Roth 2007; Rothman et al. 2000; Setola and Roth 2005).
Renewed Interest in Currently Prescribed Drugs with 5-HT2BR Agonism
Huang et al. (2009) identified a number of United States Food and Drug Administration (FDA)–approved drugs that are currently on the market as 5-HT2BR agonists and their potential for development of valvulopathy. The study included the screening of approximately 2200 FDA-approved or investigational drugs for 5-HT2BR agonism using calcium-based, high-throughput screening and five additional readouts of 5-HT2BR activation such as nuclear factor of activated T cells-mediated transcription of a β-lactamase reporter gene, ERK2 phosphorylation, β-arrestin recruitment to agonist-occupied 5-HT2B receptors, accumulation of inositol phosphates, and cell proliferation. Of the 2200 drugs screened, twenty-seven were 5-HT2BR agonists, including seven drugs known to be associated with valvulopathy and six currently prescribed drugs (guanfacine, quinidine, fenoldopam, oxymetazoline, xylometazoline, and ropinirole). Furthermore, the study revealed that the patterns of 5-HT2BR functional selectivity might help to differentiate drugs that are likely to cause valvulopathy in humans from those that are not. For example, ropinirole, a 5-HT2BR agonist approved for treating Parkinson’s disease and restless leg syndrome, is not associated with valvulopathy (Schade et al. 2007; Zanettini et al. 2007) and is distinct from the known valvulopathic 5-HT2BR agonists in 5-HT2BR functional assays (Huang et al. 2009). Additionally, ropinirole has low affinity to the human 5-HT2BR. The authors also recommended five currently prescribed drugs, such as guanfacine, oxymetazoline, quinidine, xylometazoline, and fenoldopam to be studied further in terms of their pharmacodynamics and potential for valvulopathy. Xylometazoline and oxymetazoline are nasal decongestants for short-term use, whereas fenoldopam is an antihypertensive agent used postoperatively and in-hospital during a hypertensive crisis for one-time use. In contrast, guanfacine (an antihypertensive agent) and quinidine (an antiarrhythmic agent) are indicated for prolonged use and therefore carry a greater risk for development of valvulopathy than drugs for short-term use. Additionally, recent FDA approval of guanfacine for the treatment of attention-deficit–hyperactivity disorder might further expose an increasing number of patients, including children, to a potentially valvulopathic agent (Huang et al. 2009).
Drug-induced Valvulopathy in Animal Models: Need for Understanding of Valvular Pathophysiology and Its Causes
Given the serious consequences of drug-induced valvular effects, there is a growing demand for sensitive preclinical screening strategies, including detailed and systematic examination of heart valves and comprehensive diagnosis and recording of valvular lesions in rats and mice, two commonly used rodent species. To date, there are no validated animal models or preclinical/toxicologic screens to accurately predict drug-induced valvulopathy in humans (Greaves 2007), and therefore, establishing animal models to understand mechanisms and pathogenesis of valvulopathy is warranted (Donnelly 2008). The first evidence of valvular changes caused by anorexigens was reported in rats by Bratter et al. in 1999, two years after the withdrawal of fenfluramine and dexfenfluramine from the U.S. market in 1997. In that study, there was obvious thickening of mitral valves in prenatal rats co-administered with dexfenfluramine and phentermine for fourteen days. However, microscopic evaluations of these thickened valves were not performed. It is therefore unknown whether mitral valve thickening in rats (Bratter et al. 1999) has microscopic features similar to those of human valvulopathy caused by dexfenfluramine and phentermine combinations. Pharmacologically, fenfluramines are potent 5-HT releasers as well as re-uptake inhibitors, whereas phentermine inhibits monoamine oxidase (an enzyme responsible for degradation of 5-HT), and as a result, the major emphasis has been placed on 5-HT and 5-HT receptors and their role in the pathogenesis of drug-induced valvulopathy. In 2005, Gustafsson et al. showed that daily subcutaneous injection of serotonin (20 mg/kg) in Sprague Dawley (SD) rats for three months resulted in valvular changes that are morphologically and echocardiographically similar to those seen in human carcinoid heart disease. In a separate study, Droogmans, Franken et al. (2007) showed that rats given 5-HT (20 mg/kg/day, subcutaneously) and pergolide (0.5 mg/kg intraperitoneally) for five months induced valvular regurgitation and microscopic changes similar to those seen in anorexigen-induced valvulopathy in humans. A subsequent study by Droogmans et al. (2009) showed that cyproheptadine (5-HT2BR antagonist) co-treatment prevented the development of pergolide-induced valvulopathy in rats with a reduced number of 5-HT2BR–positive VICs, suggesting that 5-HT2BR plays an important role in the pathogenesis of valvulopathy.
We previously reported the immunostaining and quantitative transcript levels of 5-HT receptors (5-HT2BR and 5-HT1BR) in the normal mitral, aortic, tricuspid, and pulmonary valves of SD rats and cynomolgus monkeys (Elangbam et al. 2005). Our results showed that the 5-HT receptor expression in four heart valves was comparable among cynomolgus monkeys, rats, and humans, and therefore, the potential exists to gain mechanistic insight to anorexigen-induced valvulopathy from animal studies. A possible association between spontaneous mitral valvulopathy (SMV) and increased number of 5-HT2BR–positive cells in valve leaflets has been described in SD rats, suggesting that 5-HT2BR may play a role in its pathogenesis. Additionally, 5HT2BR may play a role in the exacerbation of SMV (i.e., significant increase in incidence and severity of SMV) in Fischer 344 rats with dl-amphetamine treatment (Elangbam et al. 2006). Amphetamine analogs such as MDMA and MDA are potent 5-HT2BR agonists, which have been shown to cause valvular cell proliferation in vitro (Setola et al. 2003) and valvulopathy in people who took MDMA for six years (Droogmans, Cosyns et al. 2007). Interference with 5-HT transmembrane processing by knocking out the 5-HT transporter (5-HTT) gene has been shown to cause cardiac fibrosis and valvulopathy in mice, establishing a link between 5-HTT deficiency and the development of valvulopathy in vivo. Interference with 5HTT processing may also result in increased and persistent 5-HT receptor interactions, and increased valvular mitogenic activity and extracellular matrix production in the 5-HTT–knockout mice (Mekontso-Dessap et al. 2006).
Valvular thickening and subendocardial fibromyxoid proliferation noted in SMV in rats were similar to those seen in the heart valves of patients with carcinoid heart disease and in people medicated with ergot alkaloids, anorectics, or MDMA (Droogmans, Franken et al. 2007; Elangbam et al. 2006). McDonald et al. (2002) reported distinctive morphological and compositional changes to aid in the differentiation of anorexigen-induced valvulopathy from floppy (also known as myxomatous valvular degeneration), rheumatic, and carcinoid heart diseases. The size and number of onlay (refers to neo-tissue or thickening) lesions, the amount of GAGs, the location of inflammation and neovascularization are important in this distinction. Normal valves are composed of roughly equal amounts of GAGs and collagens in humans, and they rarely harbor leukocytes and blood vessels, even as they age. McDonald et al. (2002) demonstrated that anorexigen-exposed valves contain more GAGs than normal or floppy valves. The valvular lesions associated with carcinoid heart disease have GAG-rich fibromyxoid tissue and contain a large number of leukocytes and vessels per square millimeter of tissue area. Floppy valves have no significant neovascularization but have a number of onlays. In contrast, rheumatic valves have few large, fibrous onlays. Although the neovascularization is more prominent as compared with floppy valves, anorexigen-exposed valves are still less vascular but more GAG rich than valves in carcinoid heart disease. These features are indicative of a distinctive pathologic process in anorexigen-exposed valves (McDonald et al. 2002). Spontaneous mitral valvulopathy in SD rats has a morphology and matrix composition that are strikingly similar to anorexigen-induced valvulopathy in humans. Compared with normal valve leaflets, SMV exhibited a greater valve thickness, a higher amount of GAGs, and a lower amount of collagen (Figure 3 ) (Elangbam et al. 2006).
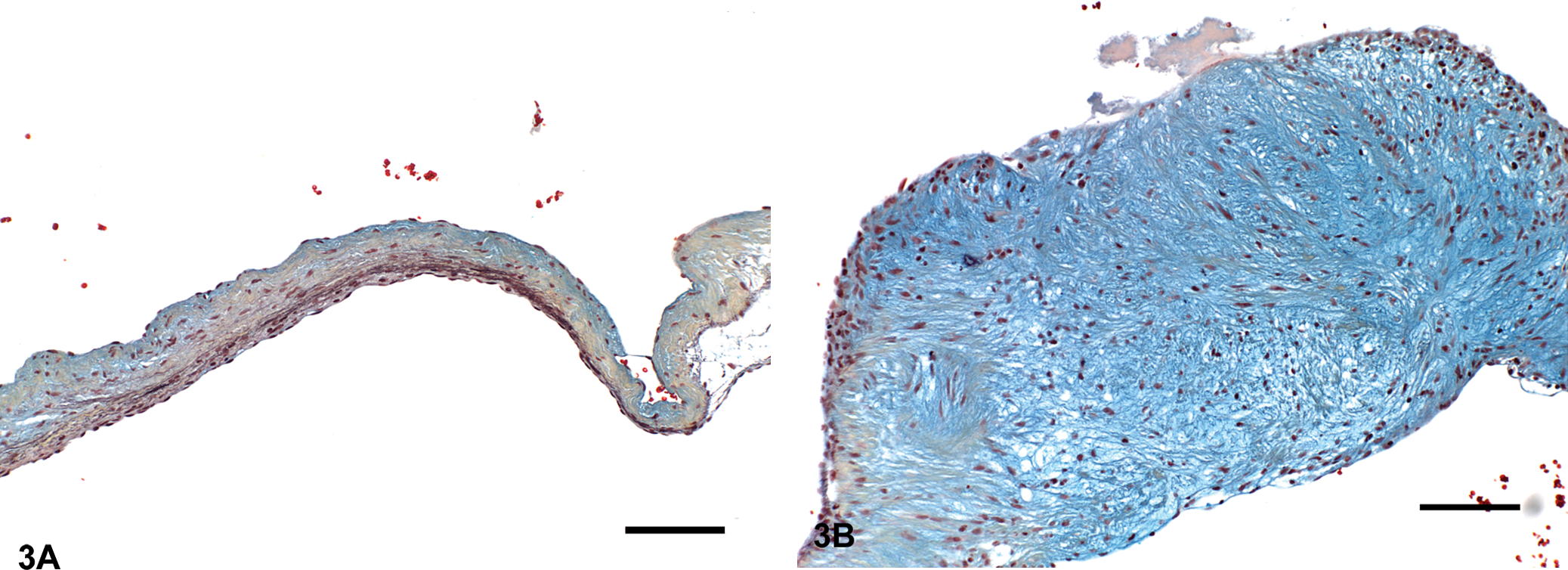
Mitral valve leaflets, Movat’s pentachrome stain. (A) Normal mitral valve leaflet from a Sprague Dawley rat. Movat’s pentachrome highlighted the glycosaminoglycans (GAGs) and collagen as cyan blue and dark yellow, respectively. Bar = 100 μm. (B) Mitral valve leaflet with spontaneous mitral valvulopathy from a Sprague Dawley rat. Note marked thickening of the valve leaflet resulting from GAGs-rich fibromyxoid tissue, and apparent segmental loss or disorganization of collagen. Bar = 100 μm.
Recently, we investigated the role of 5-HT in the development of valvulopathy in SD rats using morphological and compositional valvular analyses, as well as transcriptomic modulation of 5-HT2BR and 5-HTT genes in the aortic and mitral valves. Subcutaneous injections of 5-HT for seven days resulted in valvular thickening and compositional alterations (higher GAGs and lower collagen contents) in aortic and mitral valves of SD rats. These compositional alterations were associated with up-regulation of 5-HT2BR and down-regulation of 5-HTT genes (Elangbam et al. 2008), and therefore, 5-HT2BR activation and 5-HTT inhibition may play a significant role in the pathogenesis of 5-HT–induced valvulopathy in SD rats. In this study, we were able to recognize a distinctive 5-HT–related valvular matrix alteration after seven days in the aortic and mitral valve leaflets, thus providing a possible short-term investigative tool to study in vivo drug-induced valvulopathy by serotonergic compounds, as a way to determine or support in vitro selectivity data. Our findings further highlight the necessity and use of animal models to screen potential valvular effects of serotonergic compounds in humans.
In conclusion, valvulopathy is a serious liability for certain compound classes in development and for some marketed drugs intended for human use. Reports of valvulopathy in 1997 led to the withdrawal of anorexigens such as fenfluramine and dexfenfluramine from the U.S. market, as well as a multibillion-dollar settlement by the manufacturer of these drugs. Similarly, pergolide (for Parkinson’s disease) was withdrawn from the U.S. market in August 2007 because of increased risk of valvulopathy. The valvular effects of both fenfluramines and pergolide were not initially predicted from routine preclinical toxicity studies, although subsequent studies have shown standard laboratory models are capable of developing valvular lesions. It is probable that inconsistent sampling and interpretation may have contributed to this detection failure. To date, there are no specific validated animal models or preclinical/toxicologic screens to accurately predict drug-induced valvulopathy. Functional screens against 5-HT2BR and more consistent sampling and pathologic evaluation of valves in preclinical studies appear to hold the greatest promise for detecting drugs with the potential to cause valvulopathy. A recent report suggests that some drugs with 5-HT2BR activity with the potential for developing drug-induced valvulopathy are still in the U.S. market for long-term use, which may require careful risk–benefit analysis.
Footnotes
Acknowledgments
The author thanks Denise M. Frailey for illustrations and Drs. Roger H. Brown, Richard T. Miller, Alan H. Stokes, and Christine L. Merrill for their critical review of the manuscript.