
Abstract
In this study, the authors aimed to investigate the role of oxidative stress on the hepatic damage caused by methotrexate (MTX) and the possible protective effects of β-carotene against this damage. The rats were divided into four groups as control, MTX (20 mg/kg ip), β-carotene (10 mg/kg/day ip) + MTX, and β-carotene. Histopathologic alterations were evaluated for defining the liver damage. The tissue, malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione peroxidase (GP-x) contents and serum aspartate aminotransferase (AST) and alanine aminotranferase (ALT) activities were also examined. Histopathologic damage for each group score findings have been determined as control: 0.66 ± 0.33; MTX: 7.0 ± 0.68; β-carotene + MTX: 3.3 ± 0.42; and β-carotene: 0.5 ± 0.3. In the MTX-treated group, MDA, AST, and ALT values were increased, while SOD and GP-x values were decreased compared with the control group. In the β-carotene + MTX-treated group, AST and ALT values significantly decreased, while all other parameters were similar to the control group. This study shows that β-carotene has a protective effect on MTX-induced oxidative hepatic damage. Consequently, it seems that an antioxidant agents like β-carotene may be useful in decreasing the side effects of chemotherapy.
Introduction
Methotrexate (MTX), which is a folic acid antagonist, is used commonly as a cytotoxic agent in the treatment of leukemia and other malignancies as well as in the inflammation diseases such as psoriasis and rheumatoid arthritis in low doses (Bayram et al. 2005; Şener et al. 2006a). With the widespread use of MTX, although hepatotoxicity is the most important potential major side effect (West 1997). The toxicity of MTX in the liver seems to relate to the generation of reactive oxygen species (ROS) (Şener et al. 2006a; Uraz et al. 2008). However, the number of laboratory animal studies in which microscopic and biochemical changes have been evaluated together are scarce.
There is an abundance of evidence that the regular consumption of fruit and vegetables is associated with a reduced risk of chronic and degenerative diseases. For this reason, many researchers have been focused on this subject (Vardi et al. 2008; Keen et al. 2005; Ramos et al. 2005). The protective effects of fruits and vegetables are thought to derive from vitamins and carotenoid, flavonoid, and phenolic composites that they contain. Although more than 600 different compounds have been characterized, until now β-carotene is the most potent one of the antioxidants (Stahl and Sies 2003). β-carotene exhibits antioxidant activity by suppressing singlet oxygen, scavenging peroxide radicals, and directly reacting with peroxy radicals, thus stabilizing membrane lipids from free radical attack (Bast, Haenen, and Van den Berg 1998).
This experimental study was designed to clarify the role of oxidative stress in MTX-induced hepatic damage and the possible protective effect of β-carotene against it using histological and biochemical parameters.
Material and Methods
Experimental Conditions
For this study, 24 Wistar albino rats weighing 210–310 g were obtained from Inonu University Laboratory Animals Research Center. The rats were kept at a temperature of 21 ± 2°C and relative humidity of 60 ± 5% in a 12-hour light/dark cycle. The experiments were carried out according to the standards of animal research of the National Health Research Institute and with the approval of the Inonu University Ethical Committee. Rats were divided into 4 groups randomly, each group including 6 animals.
Group I (control) rats received the equivalent volumes of saline intraperitoneally (ip).
Group II (β-carotene) rats received β-carotene (10 mg/kg/day ip) for 24 days. The dose of β-carotene used in this study was selected on the basis of a previous related study (Matos et al. 2006). β-carotene was obtained from Sigma (St. Louis, MO, USA).
Group III (MTX) rats were given MTX (David Bull Laboratories, Mulgrave-Victoria, Australia) as a single ip dose (in saline, 20 mg/kg) on day 21 of the experiment.
Group IV (β-carotene + MTX) rats were given β-carotene by ip injection in vehicle (saline) 10 mg/kg/day for 24 days and were further administered MTX at a dose level of 20 mg/kg on day 21 of the experiment.
At the end of the 24th day, all the rats were sacrificed with cervical dislocation; their blood samples and liver tissues were excised. Tissue samples were fixed in 10% neutral buffered formalin during 24 hours and embedded in paraffin. For biochemical analysis, the centrifuged serum from blood (4 cc) taken from the inferior vena cava was stored at –30°C for aspartate amino transferase (AST) and alanine amino transferase (ALT) measurements, and the liver tissues that folio covered were stored at –30°C until assay for MDA, SOD, and GP-x measurements. The sections taken from the paraffin blocks were examined by Leica DFC-280 microscope following staining with hematoxylin-eosin (H-E) and Masson trichrome staining method.
Histological Analysis
Score of liver damage severity was semi-quantitatively assessed as follows: disruption in radial arrangement around central vein, sinusoidal dilatation, and nuclear changes in hepatocytes. The microscopic score of each tissue was calculated as the sum of the scores given to each criteria. Scores were given as absent (0), slight (1), moderate (2), and severe (3) for each criteria. Maximum score is noted as 9.
Biochemical Assessment
After the decomposed liver tissue sections were stored in deep freeze, 200 mg of liver tissue was homogenized over the ice particules in the glass and then centrifugation approximately 10 min for assessment of enzyme activities and lipid peroxidation.
SOD Assay
SOD activity was assayed using the nitroblue tetrazolium (NBT) method of Sun, Oberley, and Li (1988). The samples were subjected to ethanol-chloroform (62.5/37.5%) extraction prior to the enzyme activity assay. NBT was reduced to blue formazan by O2 –, which has a strong absorbance at 560 nm. One unit (U) of SOD is defined as the amount of protein that inhibits the rate of NBT reduction by 50%. The calculated SOD activity was expressed as U/mg protein in the tissue.
CAT Assay
CAT activity was measured at 37°C by following the rate of disappearance of hydrogen peroxide (H2O2) at 240 nm (ϵ240 = 40 M–1 cm–1) (Luck 1963). One unit of CAT activity is defined as the amount of enzyme catalyzing the degradation of 1 mmol of H2O2 per min at 37°C and specific activity corresponding to transformation of substrate (in µmol) (H2O2) per min per mg protein.
GP-x Assay
GP-x activity was determined in a coupled assay with glutathione reductase by measuring the rate of NADPH oxidation at 340 nm using H2O2 as the substrate (Lawrance and Burk 1976). Specific activity is given as the amount of NADPH (µmol) disappeared per min per mg protein.
Lipid Peroxidation Assay (MDA)
The analysis of lipid peroxidation was carried out as described by Buege and Aust (1978) with a minor modification. The absorbance of the supernatant was recorded at 532 nm. MDA results were expressed as nmol mg–1 protein in the homogenate. The protein content of samples was determined using the colorimetric method of Lowry et al. (1951) using BSA as the standard.
AST and ALT
Determination of the plasma concentrations of the liver enzymes ALT and AST were measured in serum samples obtained from all groups of rats. Activities were expressed as IUL–1. The measurements were done in accordance with the methods of the diagnostic kits (Biolabo Reagents, Maizy, France).
Statistical Analysis
A computer program (SPSS 11.0) was used for statistical analysis. The results were compared with Kruskal-Wallis variance analysis. Where differences among the groups were detected, group means were compared using the Mann-Whitney U (Bonferroni) test. Values of p < .05 were considered significant. All results were expressed as means ± standard error (SEM).
Results
Light Microscopic Evaluations
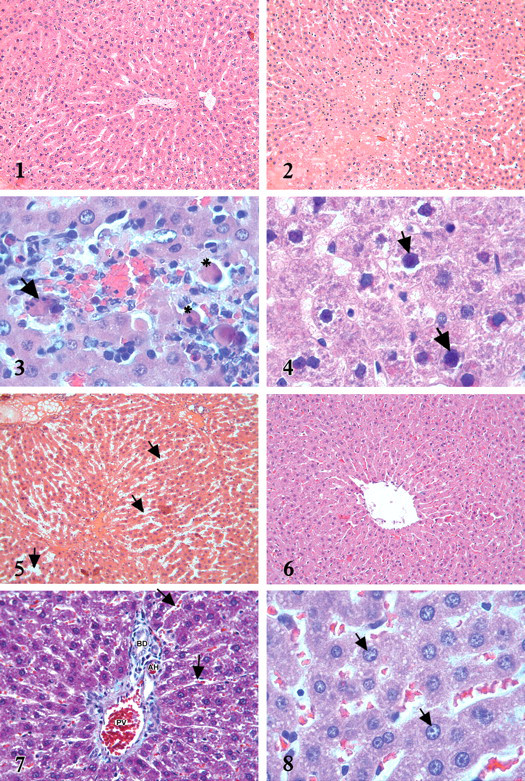
In the livers of rats from the control and β-carotene-treated rats, hepatocytes showed a normal histological appearance (Figure 1 ). In the MTX-treated rats, the normal radial arrangement of hepatocytes from central vein were abnormal (Figure 2). Foci of apoptopic cells were detected in some slides inside the lobule (Figure 3). Another conspicuous finding in this group was the difference in the appearance of the hepatocyte nucleus. Shrinkage and inflection were observed in the nuclear contour, and condensation in the structure of chromatin was also observed (Figure 4). In the sinusoid lumen of some sections, dilatation was observed (Figure 5 ). In rats given β-carotene + MTX, affected to a significantly less degree than those in the group given MTX alone (Figure 6). The histological appearance of the sinusoidal lumen in the β-carotene + MTX group was also similar to that in the control group (Figure 7). Most of the nuclei of hepatocytes were euchromatic and normal shape (Figure 8). Apoptotic cells were not observed in this group. However, β-carotene treatment did not completely alleviate the lesions, and milder degenerative alterations were still present. The microscopic damage score for each group was determined in the histological section, and results are given in Table 1.
Figure 1. Normal histological appearance of liver in control group. H-E X20.
Figure 2. Disruption of radial arrangement of hepatocytes in MTX group. H-E X20.
Figure 3. Apoptotic cell (star) and apoptotic body (arrow) distinguished by dense eosinophilic cytoplasm and pyknotic nucleus in MTX group. H-E X100.
Figure 4. Nucleus with irregular contour and dense chromatin structure (arrows) in MTX group. H-E X100.
Figure 5. Dilatation in sinusoids (arrows) observed in MTX group. H-E X20.
Figure 6. Radial hepatocytes from central vein to periphery in β-carotene + MTX group. H-EX20.
Figure 7. The appearance of sinusoids in β-carotene + MTX group (arrows). PV: Portal vein, AH: Hepatic artery, BD: Bile duct. Masson trichrome staining X40.
Figure 8. Hepatocytes whose nucleus (arrows) are euchromatic and its contours regular in β-carotene + MTX group. H-E X100.
Comparison of the effect of β-carotene on microscopic damage caused by MTX in liver.

a Significantly increased when compared with control group, p = .003.
b Significantly decreased when compared with MTX group, p = .007.
c Significantly increased when compared with control group, p = .004.
Biochemical Results
Biochemical results are given in Tables 2 and 3 .
Comparison of the effect of β-carotene on lipid peroxidation caused by MTX and the changes in antioxidant enzyme parameters.
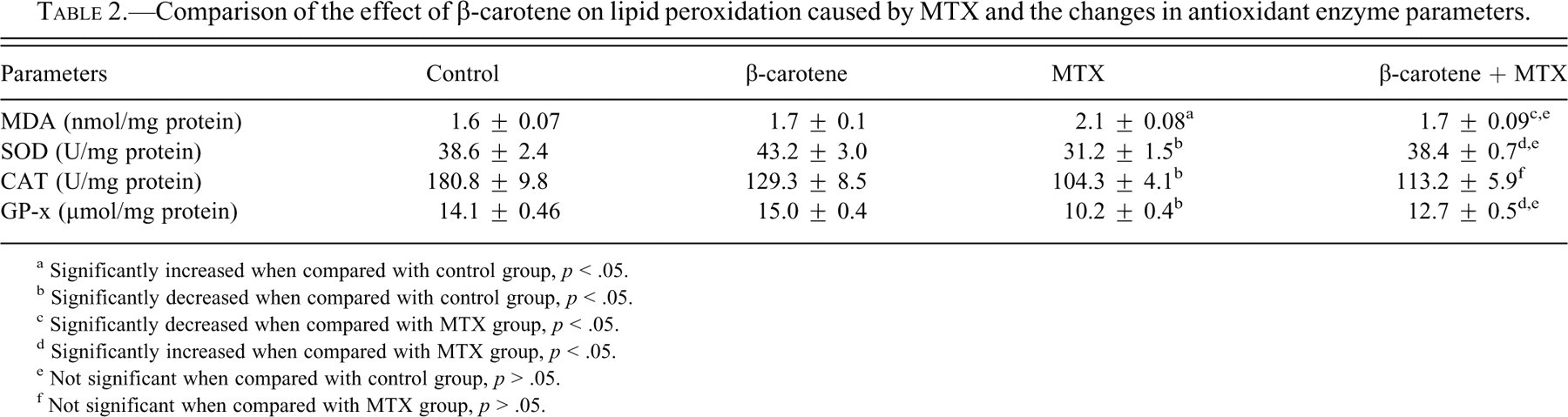
a Significantly increased when compared with control group, p < .05.
b Significantly decreased when compared with control group, p < .05.
c Significantly decreased when compared with MTX group, p < .05.
d Significantly increased when compared with MTX group, p < .05.
e Not significant when compared with control group, p > .05.
f Not significant when compared with MTX group, p > .05.
Comparison of the effect of β-carotene on the changes in AST and ALT enzyme parameters caused by MTX.
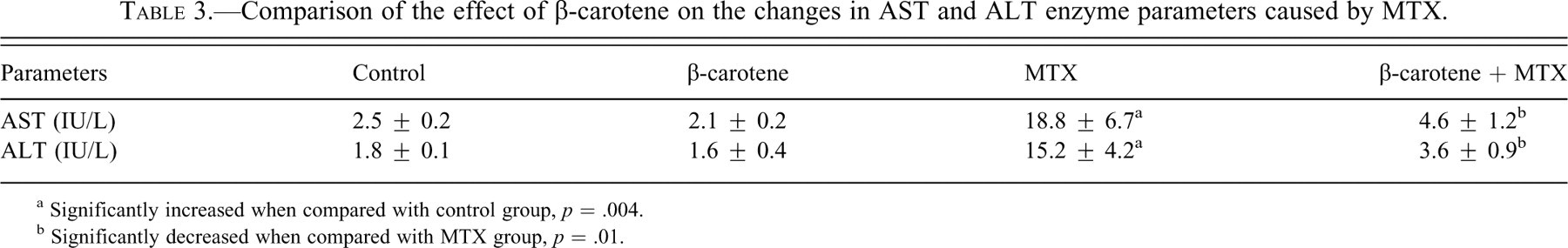
a Significantly increased when compared with control group, p = .004.
b Significantly decreased when compared with MTX group, p = .01.
In the MTX group, the level of MDA, the end product of lipid peroxidation, significantly increased (p = .008), while SOD (p = .03), CAT (p = .004), and GP-x (p = .004) activities significantly decreased compared with the livers of rats in the control group. When rats were given β-carotene + MTX, their biochemical parameters were statistically different from those of the MTX-alone group. β-carotene administration caused significant decreases in the hepatic MDA levels (p = .019) and increases in SOD (p = .004) and GP-x (p = .016) enzyme activities when compared with the levels in the livers from rats given MTX alone. But β-carotene administration before MTX had no statistically significant effect on CAT (p = .26) levels. AST and ALT activities showed increases in rats given MTX alone in comparison with the levels in control rats (p = .004). These values were decreased in the β-carotene + MTX group but were still higher than those seen in control animals. This decrease was found significant when compared with MTX group (p = .016).
Discussion
The results of the studies indicate that MTX causes oxidative tissue damage by increasing lipid peroxidation in the liver tissue and decreasing the level of antioxidant enzymes. Moreover, increased AST and ALT values, biochemical indicators of liver damage, and histopathological findings supported this conclusion. It has also been shown that β-carotene given for 21 days before the MTX application provided significant protection from the hepatotoxicity of MTX.
Drugs used for cancer chemotherapy are well known to produce acute toxic side effects in multiple organ systems. The most common target organs are tissues that contain self-renewing cell populations such as bone marrow, gastrointestinal tract, mucosal membranes, and hair follicles (Kim, Kim, and Chung 1999). It has been reported that liver damage may occur as well in particular high doses or following chronic administration of MTX (Şener et al. 2006b; Uraz et al. 2008).
The conversion of MTX to its major extracellular metabolite, 7-hydroxymethotrexate, takes place in the liver, where it is oxidized by a soluble enzymatic system. Inside cells, MTX is stored in a polyglutamated form. Long-term drug administration can cause accumulation of MTX polyglutamates and decreased folate levels. The presence of higher levels of polyglutamates causes a longer intracellular presence of the drug, and this has been suggested as a mechanism for MTX hepatotoxicity (Johovic et al. 2003). Moreover it is known that oxidative stress plays a role in the tissue damage caused by MTX (Şener et al. 2006b; Johovic et al. 2003; Cetinkaya et al. 2006). It is demonstrated that the cytosolic nicotinamide adenosine diphosphate (NADP)–dependent dehydrogenases and NADP malic enzyme are inhibited by MTX, suggesting that the drug could decrease the availability of NADPH (nicotinamide adenosine diphosphate hydrogen) in cells (Johovic et al. 2003). Under normal conditions, NADPH is used by glutathione reductase to maintain the reduced state of cellular (GSH) glutathione, an important cytosolic antioxidant. Thus, the significant reduction in glutathione (GSH) levels promoted by MTX could lead to a reduction of effectiveness in the antioxidant enzyme defense system, sensitizing the cells to ROS (Babiak et al. 1998). In this study, it has been shown that MTX administration increases the level of MDA in the liver and decreases the enzyme activities of SOD and GP-x capacity. In previous studies (Vardi et al. 2008, 2009), we have observed similar effects of MTX on intestinal and testicular tissue. This finding is in agreement with several reports demonstrating that MTX induces oxidative stress in tissues as demonstrated by increasing MDA levels (Uraz et al. 2008; Johovic et al. 2003; Cetinkaya et al. 2006; Cetin et al. 2008) and decreasing SOD activities (Cetinkaya et al. 2006; Cetin et al. 2008). The oxidative damage increasing with MTX in liver tissue was prevented by dosing β-carotene for 21 days prior to MTX administration. It plays an important role in protecting cell membrane against oxidative damage because of its property of scavenging lipid and peroxyle radicals (El-Demerdash et al. 2004; Sergio and Russel 1999). Berryman et al. (2004) reported that in the liver of diabetic rats, MDA levels significantly increased when compared a control group, whereas the addition of β-carotene returned levels back to normal. In this same study, the addition of β-carotene also increased the activity of GP-x. Similar to our findings, β-carotene is reported to increase antioxidant enzymes in tissues, or prevent their decrease in a series of different experiment models (Vardi et al. 2008, 2009; El-Demerdash et al. 2004; Kheir-Eldin et al. 2001). Under the current experimental conditions, pretreatment with β-carotene may have protected the hepatocyte membrane integrity by preventing the peroxidation of fatty acids by free radicals and inhibiting an important mechanism that causes liver damage. In our study, after MTX application, AST and ALT levels, known indicators of hepatic damage, had increased 7 fold compared with the control group. It is known that in the use of drugs causing liver toxicity transaminase levels increase in the plasma depending upon the degree of hepatic damage. Kuvandik et al. (2008) reported that the levels of AST and ALT in plasma serum have increased significantly in acetaminophen hepatotoxicity; and Uraz et al. (2008) reported that MTX also increased ALT levels. In our study; the level of AST and ALT in the group supplied with β-carotene was statistically lower than in those animals given MTX alone. Sugiura et al. (2006), reported that carotenoids may prevent increases of serum aminotransferase increase in the early periods of liver damage in hyperglycemic patients. In our microscopic investigations in the livers of rats given MTX, hepatic nuclei were larger and karyolemma contours were irregular; and in some areas, apoptopic hepatocytes could be seen. MTX acts as a dihydrofolic acid analogue that binds to the dihydrofolic acid reductase enzyme by inhibiting the synthesis of tetrahydrofolate, which is required for DNA synthesis (Babiak et al. 1998). De novo inhibition of purine and pyrimidine synthesis leads to DNA defects, which results in apoptosis (Tsurusawa, Saeki, and Fujimoto 1997). Increased oxidative stress may cause shape and structural changes of the nucleus by causing DNA fragmentation and denaturation, which play a critical role in the initiation of apoptosis (Prahalathan, Selvakumar, and Varalakshmi 2005). It has been shown that production of ROS may mediate a signal for apoptotic cell death (Vardi et al. 2009). The current study has shown that β-carotene + MTX group nucleus changes were limited, and apoptosis was not observed. β-carotene may have protected cells from DNA changes and apoptosis with the ability of connecting singlet oxygen radicals and scavenging peroxyle radicals.
As a result, β-carotene probably protects from MTX-induced liver damage by scavenging of free radicals. Therefore, β-carotene might be an effective therapy in protecting the liver in patients given MTX treatment. Although further experimental and clinical studies are required to confirm these findings.