
Abstract
Long-term, low–dose rate (LDR) ionising radiation has been found to increase the risk of leukaemia and other haematological effects. Understanding the mechanisms underlying these effects can be useful for guiding radiation protection policies. This study explores the utility of an organ-on-a-chip (OOC) system as a platform for prolonged LDR studies on haematopoietic stem cells (HSCs). Control samples exhibited a higher population of cells in the primitive state than irradiated samples, revealing a nuanced impact of prolonged LDR exposure on HSC behaviour. RNA profiling further identified differential expression in 166 genes between these samples, offering insights into molecular mechanisms underlying the radiation response. This study underscores the usefulness of OOC systems in advancing our understanding of longer-term responses to LDR irradiation.
Keywords
INTRODUCTION
Haematopoietic stem cells (HSCs) are known to balance self-renewal and differentiation within the dynamic microenvironment of the niche, but there is currently little understanding of the effects of prolonged low–dose rate (LDR) on this intricate biological process (Pajonk and Vlashi, 2013).
Cohort studies investigating individuals working in the nuclear industry, exposed to prolonged periods of LDR irradiation, have demonstrated an elevated risk of leukaemia (Boice Jr et al., 2022). Additionally, a separate cohort study observed alterations in platelet counts among medical workers following cumulative long-term exposure to LDR irradiation (Liu et al., 2021). A better understanding of the mechanisms underlying these effects will be useful to guide radiation protection policies, but there are limitations posed by traditional experimental approaches to study this, such as short-term human peripheral blood culture. In response to the challenge of the short ex-vivo survival of peripheral blood cells, this study instead strategically employs haematopoietic stem and pluripotent cells (HSPC) cultured with mesenchymal stromal cells (MSC) on a porous ceramic scaffold, whilst having actively circulating media from a co-compartment reservoir, creating a physiologically relevant niche. This organ-on-a-chip (OOC) approach is recognised for extending the survival of HSPCs and facilitating the preservation of a self-renewing cell population and generation of mature haematopoietic lineages, while allowing for flexibility in experimental setups that animal models may not have (Frenette et al., 2013; Sieber et al., 2018; Hashem-Boroojerdi et al., 2022). In this way, the effect of prolonged LDR irradiation on long-term self-renewal, differentiation capacity, and gene expression profiles of HSC can be studied.
MATERIALS AND METHODS
Organ-on-a-chip system
Human MSCs (Stemcell Technologies; #70071) were seeded onto hydroxyapatite-coated zirconium oxide–based Sponceram 3D ceramic scaffolds (Zellwerk GmbH) to mimic the bone marrow (BM) microenvironment. Subsequently, human BM–derived CD34+ cells (Stemcell Technologies; #70002) were seeded onto each ceramic scaffold and transferred into microfluidic chips (TissUse GmbH; C2-96) connected to a pump control unit. Media within chips were circulated at a rate of 27 beats per minute. This facilitates extended culture durations by promoting the circulation of HSPCs within a microenvironment resembling the BM.
Irradiation
Microfluidic chips containing BM organoids were irradiated with a 335 ± 50 MBq, 0.662 MeV photopeak, and 1 mGy h−1 dose rate cesium-137 irradiator with culture conditions of 37°C and 5% CO2 over a period of 57 days for a total dose of 3.012 Gy.
Sample collection and analysis
At the conclusion of the experimental phase, RNA extracted from cells isolated from the scaffold was analysed for differential gene expression (DGE) using the NanoString nCounter SPRINT platform and the stem cell characterisation panel (NanoString; 115000499). Data analysis was performed using NanoString nSolver and Partek Genomics Suite. Harvested cells also underwent flow cytometry analysis with DURAClone SC Haematopoietic Stem Cell kit (Beckman Coulter Life Sciences; C49589).
RESULTS
Cell differentiation in control and irradiated HSPCs
The analysis of haematopoietic cell populations in control and irradiated samples revealed variations across distinct cell subsets (Fig. 1). For mature HSPCs (Fig. 1B), irradiated samples showed a greater ratio for percentage singlets relative to freshly thawed HSPCs, compared to control samples. The same trend was observed for granulocyte-monocyte progenitors (GMPs) (Fig. 1F). These results suggest an inclination of HSPCs cultured under prolonged LDR irradiation towards the myeloid lineage. Similarly, for the more immature, self-renewal multipotent progenitors (MPP), irradiated samples displayed a smaller ratio compared to control samples, suggesting a proclivity towards more mature haematopoietic lineages (Fig. 1C). In general, a decrease was observed in the populations of primitive state HSPC, MPP, myeloid lineage progenitors (MLP), and common lymphoid progenitors (CLP) (Fig. 1A, C–E), in irradiated compared to control samples. This observation reinforces the indication of a tendency towards myeloid lineages over lymphoid lineages.
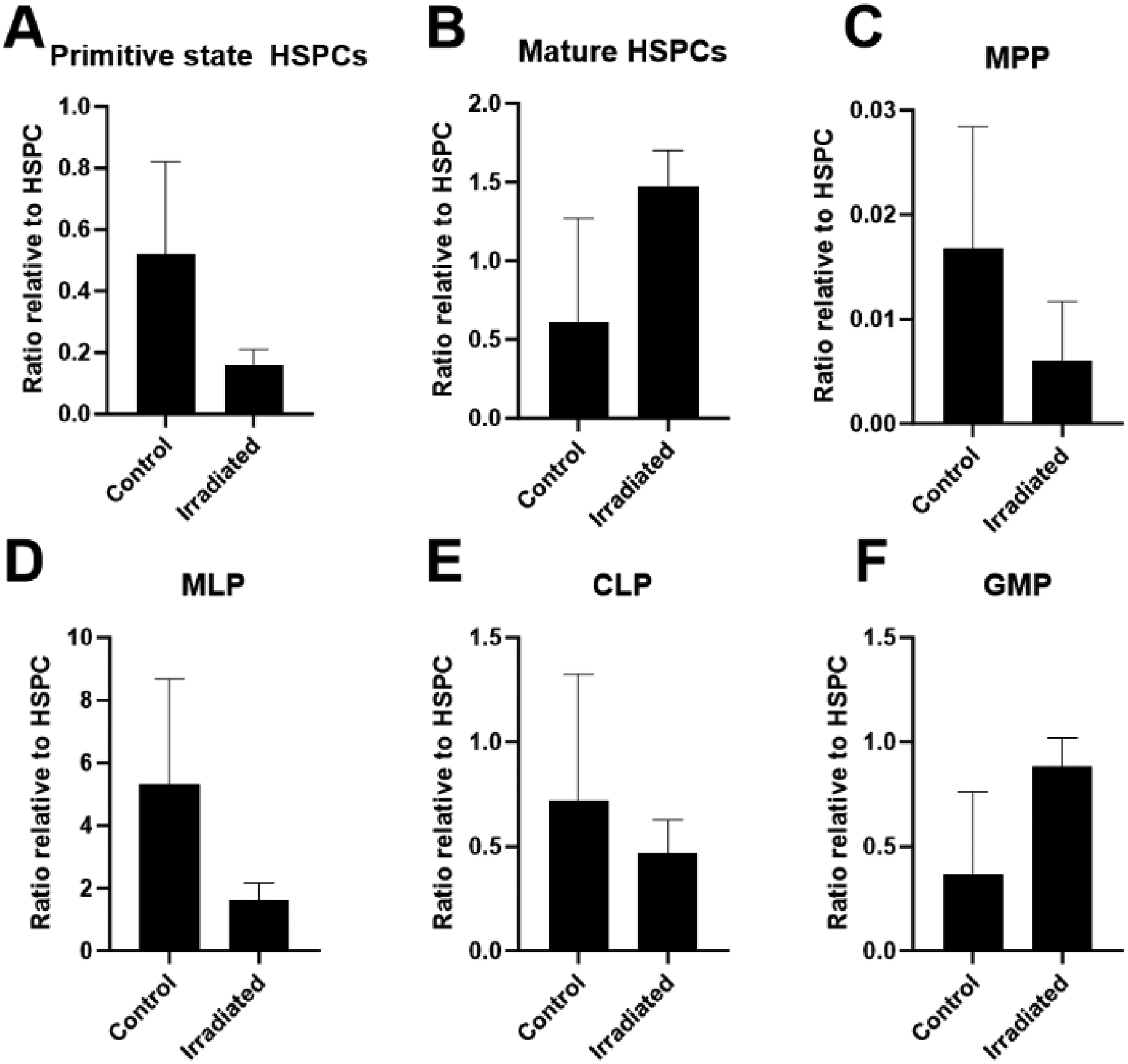
Ratios of percentage of singlets postculture relative to freshly thawed HSPCs on Day 1 of culture (n = 2). Error bars represent geometric standard deviations. (A) Primitive state HSC CD34+CD38−, (B) mature HSPC CD34+CD38+, (C) multipotent progenitors (MPP) CD34+CD38−CD45RA−CD49f−CD90−, (D) myeloid lineage progenitors (MLP) CD34+CD38−CD45RA+CD90−, (E) common lymphoid progenitors (CLP) CD34+CD38−/loCD45RA+CD90−, (F) granulocyte–monocyte progenitors (GMP) CD34+CD38+CD45RA+.
A three-dimensional principal component analysis (PCA) plot (Fig. 2) encompassing 770 genes provided a snapshot of the gene expression landscape. The segregation between control and irradiated samples suggests a discernible impact of prolonged LDR on the gene expression profile. One-way ANOVA was used subsequently to identify DGE between irradiated and control samples (Fig. 3).
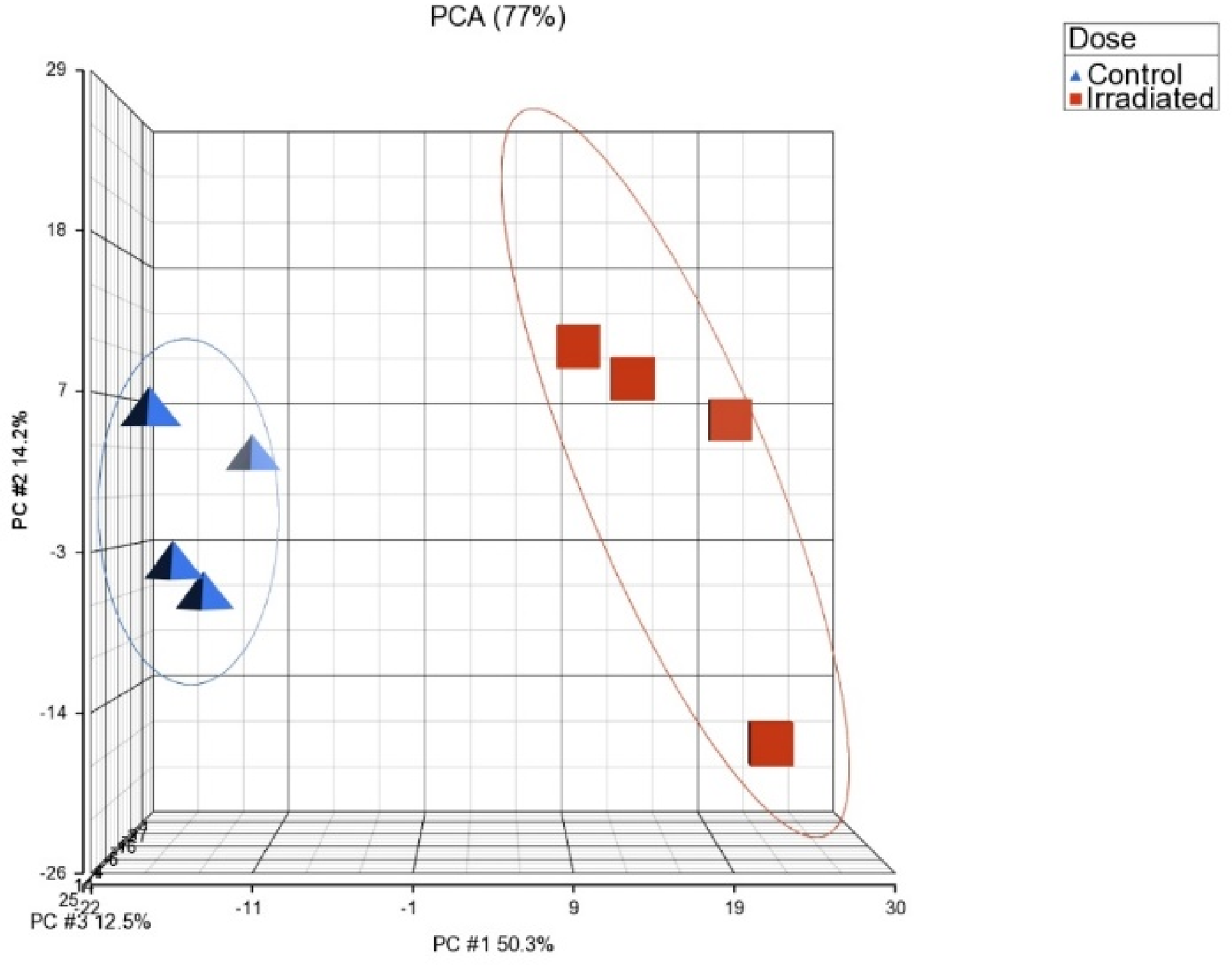
Three-dimensional principal component analysis (PCA) plot depicting the clustering of samples based on experimental conditions (n = 4). The first three principal components collectively account for 77% of the dataset's variance, with PC1 playing a prominent role (50.3%).
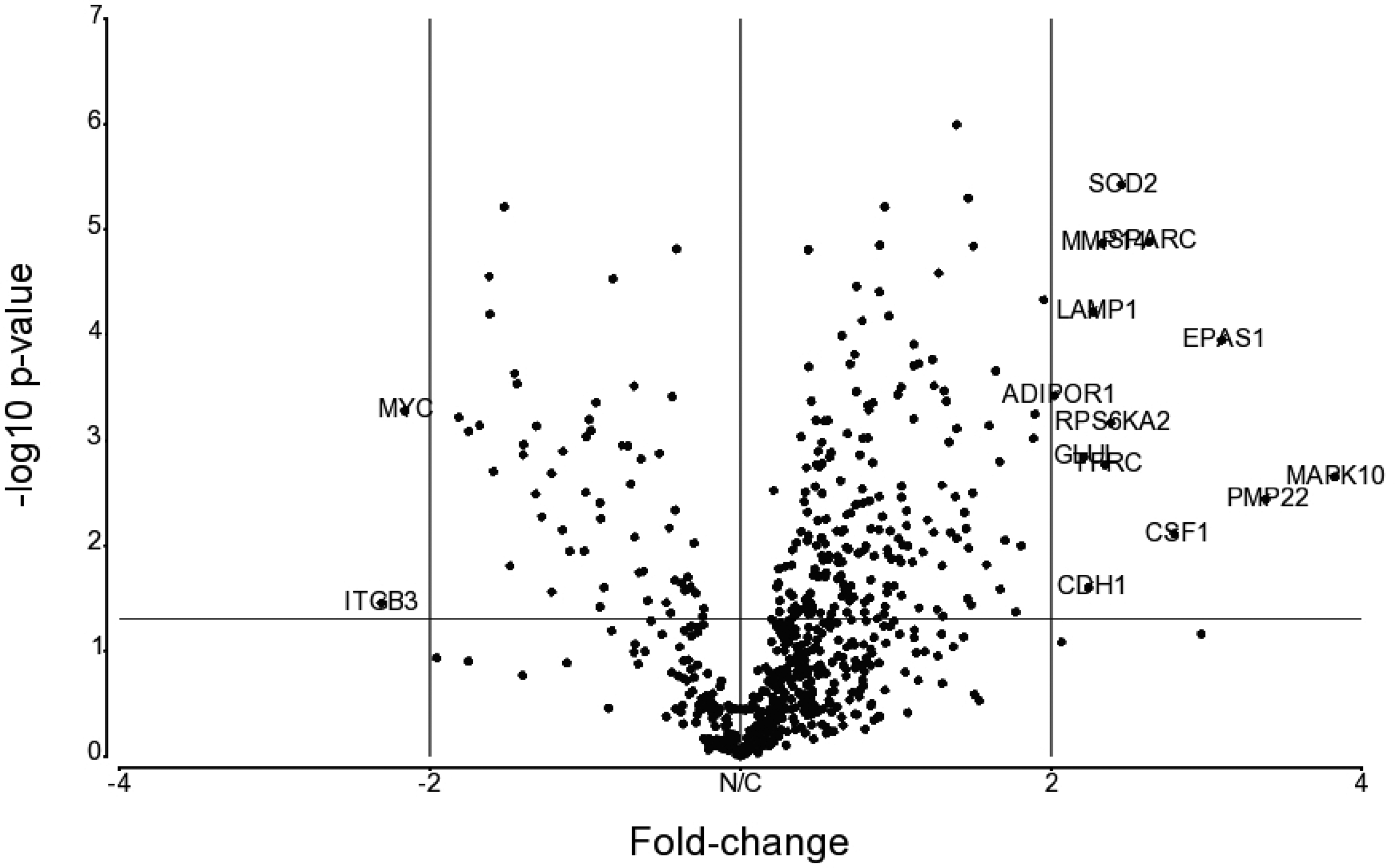
Volcano plot of DGE between irradiated versus control samples. Genes are plotted based on their fold-change against unadjusted p-value after one-way ANOVA (n = 4). Labelled on the plot are genes exhibiting fold-changes of magnitude 2 or greater.
A subsequent refinement through FDR correction narrowed down 166 genes, each demonstrating statistical significance (FDR < 0.05). From this group, 13 genes with fold-changes of magnitude 2 or greater are depicted in Table 1. This includes the well-established radiation-responsive gene MYC, while another biomarker of acute radiation syndrome WNT3 was also seen with FDR = 0.01 and fold-change = 1.32. It would be of interest to analyse the similarities and differences in such DGE between prolonged LDR and acute exposure (Woloschak and Chang-Liu, 1995; Cruz-Garcia et al., 2021; Abend et al., 2023).
Top 13 differentially expressed genes with fold-changes of magnitude 2 or greater.

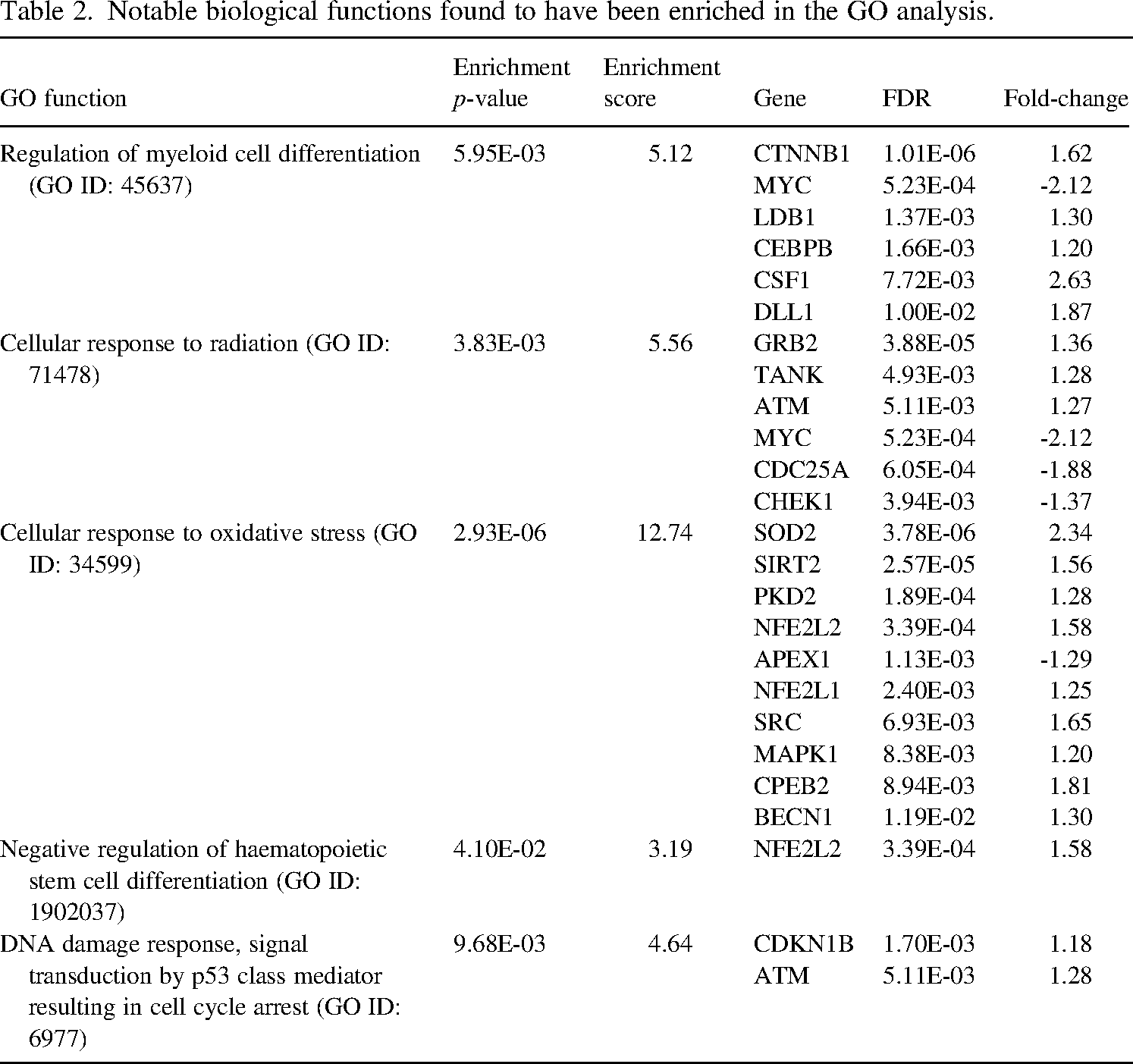
Functional enrichment analysis was subsequently performed, and Gene Ontology (GO) analysis showed enrichment in notable biological functions known to be related to radiation exposure (Table 2). An observable trend of gene upregulation of irradiated versus control samples is observed across the enriched biological functions. This underscores a pervasive molecular response within the experimental context, highlighting activation of key pathways associated with prolonged LDR.
Notable biological functions found to have been enriched in the GO analysis.
In prior unreported experiments, attempts to conduct prolonged LDR studies using peripheral blood cultures faced significant challenges, primarily attributed to the poor viability of blood cell populations and extensive haemolysis. In contrast, the cultivation of BM organoids in OOC systems for 57 days exposed to LDR irradiation enabled exploration of discernible molecular changes. Notably, HSPCs exhibited a propensity towards myeloid lineages, evident in the increased CD38 expression, consistent with other LDR studies (Fujishiro et al., 2017). Further exploration, with an increased sample size, into potential factors influencing this tendency would be warranted.
DGE in irradiated BM organoids also raises the prospect of identifying potential biomarkers and molecular pathways involved in prolonged LDR exposure. GO analysis revealed genes associated with oxidative stress and DNA damage responses, which are established effects of ionising radiation (UNSCEAR, 2000; Sudprasert et al., 2006). Providing additional support for the inclination of irradiated HSPCs towards myeloid lineages, the elevated expression of CSF1 observed in this study aligns with findings from a complementary mouse model investigation (Li et al., 2004).
In summary, the utilisation of BM organoids in OOC cultures emerges as a promising alternative for studying the effects of long-term LDR exposure, to offer valuable insights into the underlying mechanisms governing cell fates and molecular pathways.
Footnotes
DECLARATION OF CONFLICTING INTERESTS
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this research was provided by the National Research Foundation, Singapore.