
Abstract
Anticipatory planning, that is, the ability to process information in advance to prepare to act, underpins activities that unfold in time, such as spoken language, reading and handwriting. One key, though not the only, mechanism that may facilitate anticipatory planning is rhythm, which is understood as the structured grouping of events in time. Research has identified impairments in the production and processing of rhythm in individuals with dyslexia, with individual differences in rhythmic skills explaining variability in language, reading and motor abilities. In this article, we synthesise behavioural evidence suggesting that language, reading and handwriting impairments associated with dyslexia may, to some extent, be linked to a deficit in anticipatory planning. We term this the Inefficient Anticipatory Planning Hypothesis. We discuss the need for further behavioural and neural research to clarify the nature of anticipatory planning across these domains, and the extent to which different mechanisms, such as rhythm, support it. Given the interplay between language and rhythmic skills and the comorbidity of dyslexia and developmental language disorder (DLD), extending this investigation to (developmental) language disorder could refine our understanding of the distinct and shared mechanisms that underlie these conditions.
Glossary
Introduction
Dyslexia is a specific learning disorder characterised by difficulties in accurate or fluent word recognition, poor decoding and poor spelling abilities despite the provision of interventions that target those difficulties. Dyslexia may also be associated with challenges in reading comprehension and mathematical reasoning (American Psychiatric Association, 2022). Dyslexia is a complex, heterogeneous condition involving a broad range of genetic, neural and cognitive deficits (Catts et al., 2024; McGrath et al., 2020; Pennington, 2006; Pennington et al., 2012; Ring & Black, 2018). Although reading is the primary area of difficulty, research has uncovered a variety of co-occurring conditions in individuals with dyslexia. For instance, dyslexia often co-occurs with developmental language disorder (DLD; Adlof & Hogan, 2018; Catts et al., 2005; McArthur et al., 2000; Ramus et al., 2013; Snowling et al., 2020). Children with dyslexia exhibit language difficulties even before exposure to literacy, manifesting as early delays in language development (McBride-Chang et al., 2011; Scarborough, 1990; Snowling & Melby-Lervåg, 2016). Among the language deficits, poor phonological skills are recognised as a hallmark of dyslexia (Ramus et al., 2013; Snowling, 1998), although the exact nature of the phonological deficit is debated (Berent et al., 2012; Ramus & Szenkovits, 2008).
Morphology and syntax are also compromised in individuals with dyslexia (Delage & Durrleman, 2018; Guasti, 2013; Guasti et al., 2015, 2018; Moll et al., 2015; Robertson & Joanisse, 2010; Siegel, 2008; Vender et al., 2017). For instance, McArthur et al. (2000) reported that 55% (61 out of 110) of children with dyslexia showed significant deficits in syntactic and morpho-syntactic skills as indexed by their performance on the Clinical Evaluation of Language Fundamentals test (Semmel et al., 1987). Studies using measures of event-related potentials have found that the processing of sentences with morpho-syntactic violations (e.g., subject–verb agreement violations) is atypical in children and adults with dyslexia (Canette et al., 2020; Cantiani et al., 2013; Rispens et al., 2006). Evidence also points to deficits in fine and gross motor skills associated with dyslexia (Decarli et al., 2024; Nicolson & Fawcett, 1990, 2011). Nicolson and Fawcett (1990, 2011) have proposed a theoretical framework that posits that individuals with dyslexia have a deficit in skill automatisation (i.e., making a skill automatic) and that this deficit affects cognitive skills like reading and motor skills, particularly those involving balance and coordination. There is additional evidence of poor handwriting skills in children with dyslexia (Alamargot et al., 2020; Berninger et al., 2008; Gosse & Van Reybroeck, 2020; Kandel et al., 2019; Pagliarini et al., 2015).
These findings highlight the variation of deficits that co-occur with dyslexia. Reading, language and motor movements are all activities that unfold in time and in which the individual acts are interdependent. As such, they all involve coarticulation in a broad sense: what one does at time t is influenced by what one will do at time t+1. For instance, in the speech production of the word tulip, the lips start rounding for the vowel /u/ even before the consonant /t/ is pronounced. Similarly, when reading a word aloud, one’s eyes are already looking at the text ahead (Laubrock & Kliegl, 2015). In handwriting, how one writes a particular letter (e.g., movement velocity or letter shape) depends on the features of the following letter (Kandel & Perret, 2015). While doing an action at time ti, one must concurrently process information and prepare for the following action at time ti+1.To speak, read or perform motor movements, one needs to be in an anticipatory mode; that is, one must process information in advance to be ready to act at the right moment (Guasti et al., 2018). Importantly, anticipation is facilitated by the presence of temporal organisation, a grouping of events generally coherent with a regular pulsation (metrical level), that allows one to anticipate what is next.
Several theories have emphasised the role of some aspects of temporal organisation in understanding dyslexia. The Temporal Sampling Framework (Goswami, 2011; Goswami et al., 2014) posits that dyslexia arises from impaired phase-locking of neural oscillations in sensory auditory areas, which are essential for processing the temporal regularities of speech. As a result, the accurate perception of temporal information of speech is hindered, leading to phonological processing deficits, which are causally linked to reading difficulties. Beyond reading and phonology, it is proposed that the atypical neural oscillations may impact sensory processing in motor and visual areas and could explain the motor, visual and attentional processing deficits associated with dyslexia. Goswami (2011) further notes that better discrimination occurs when an auditory event is anticipated in a regular and predictable sequence, narrowing the attention window. Goswami (2011), therefore, suggests that impaired neural phase-locking may cause difficulties for individuals with dyslexia in forming internal representations of temporal organisation. In line with this view, research has demonstrated that individuals with dyslexia exhibit atypical neural oscillatory activity in the auditory and visual modalities (Lallier et al., 2017), weaker synchronisation of low-frequency oscillations to speech edges which are key for speech tracking (Lizarazu et al., 2021) and reduced neural synchrony to low-frequency speech oscillations (Molinaro et al., 2016).
More recently, Ladányi et al. (2020) introduced the Atypical Rhythm Risk Hypothesis, proposing that individuals with poor rhythmic skills are at higher risk for developmental speech and language disorders, including dyslexia. This theory draws on a large body of research showing (a) shared underlying mechanisms between the temporal processing in music and speech/language, (b) associations between temporal processing in music and speech/language in typically and atypical developing populations and (c) individual differences in temporal processing in infants which are linked to later language development outcomes. Ladányi et al. (2020), referring to the Dynamic Attending Theory (Large & Jones, 1999; Large & Palmer, 2002), hypothesise that neural oscillations synchronise with external regularities and predictable cues like beats or stress in periodic auditory stimuli. This synchronisation is suggested to structure the auditory input and focus attention on critical auditory elements, which aids in predicting when the next event will occur in a periodic sequence (Kotz et al., 2009).
Drawing on theories from different fields, Fiveash et al. (2021) proposed the Processing Rhythm in Speech and Music (PRISM) framework, which outlines three central shared elements that underpin temporal processing in music and speech: (a) precise, fine-grained auditory processing, (b) synchronisation/entrainment of neural oscillation to external metric (periodic) stimuli and (c) coupling between perception and production. Although Fiveash et al. did not include prediction and anticipatory planning as mechanisms within their framework, they indicated that prediction is related to all three mechanisms of PRISM.
The existing theories have pointed out the role of prediction at the neural level as a shared mechanism in processing temporal organisation in language and music. However, these theories have not explicitly explored anticipatory planning and its manifestation in domains beyond language and music, such as reading and motor activities. In this opinion article, we argue that anticipatory planning is a mechanism linking speech, language, reading and handwriting, which are all phenomena that unfold in time. We introduce the Inefficient Anticipatory Planning Hypothesis, proposing that a deficit in the anticipation mechanisms may partially account for the language, reading and handwriting deficits often observed in individuals with dyslexia.
We present behavioural evidence directly supporting this view; however, the available data remain limited. Our aim is to point out the role of anticipation and provide insights into the viability of our hypothesis, opening new avenues for research. Accordingly, we demonstrate that spoken language, reading and handwriting involve anticipatory planning. Where available, we provide evidence that individuals with dyslexia show inadequate anticipatory skills. Where direct evidence is lacking, particularly in language and reading, we draw on findings from neurotypical individuals and suggestive data from individuals with dyslexia that may be linked to our hypothesis. Further research is essential to refine this perspective and advance our understanding of the role of anticipatory planning in dyslexia. We acknowledge that dyslexia is a heterogeneous and multifaceted condition (McGrath et al., 2020; Pennington, 2006; Ring & Black, 2018) in which temporal processing deficits 1 are prominent. We do not claim that poor anticipatory planning is the primary or sole cause of dyslexia, but we believe it is a crucial mechanism for understanding these deficits.
This article is organised into five sections. The first, ‘What is rhythm and what is it used for,’ outlines the distinction between metre and rhythm. This distinction is commonly used in music science and linguistics and is helpful for discussing the behavioural data in the literature concerning the production and perception of events that unfold in time. The second section, ‘Temporal deficits in dyslexia,’ synthesises behavioural evidence on how and to what extent synchronisation and anticipatory planning are affected in individuals with dyslexia. The third section, ‘Anticipatory planning in phenomena that unfold in time,’ reviews the role of anticipation in language processing, reading, and handwriting and discusses evidence, where available, on how these areas are affected in individuals with dyslexia. The fourth section, ‘Anticipatory planning as a link between language, reading and handwriting,’ introduces the Inefficient Anticipatory Planning hypothesis. The final section, ‘Reflections on the Inefficient Anticipatory Planning hypothesis,’ discusses our view’s strengths, limitations, implications, and directions for future research.
What is rhythm, and what is it used for?
The literature lacks a precise, universally agreed-upon definition of rhythm (for a list of definitions, see Toussaint, 2019, pp. 1–6). This has consequences for understanding its role in human behaviour and how deficits in the production and perception of temporal events can be characterised. Since ‘rhythm’ has different meanings, we will start by defining how we use it in this article to avoid ambiguities.
In music and music cognitive science, there is a distinction between metre and rhythm (Cooper & Meyer, 1960; Essens & Povel, 1985; Lerdahl & Jackendoff, 1993; Levitin et al., 2018; London, 2012; Ozernov-Palchik et al., 2018). Figure 1 depicts the structural differences between metre and rhythm. A fundamental element for understanding metre and rhythm is the beat, the basic unit of time in music that marks time points. Beats are the basis of metre (Cooper & Meyer, 1960; Large & Palmer, 2002). The metre is a temporal frame of reference that, in the most basic form, is characterised by a segmentation of the temporal continuum into intervals of equal duration (Large, 2000). This segmentation of the temporal continuum can be marked by explicit temporal events (e.g., metronome beats, short patterns of events with strong and weak beats that define the measure in Western music notation, or even foot or finger taps) or implicit events represented in our mind. The hallmark of the different varieties of metres is periodicity, that is, the reproduction at equal intervals of the same pattern of events (see the first line in Figure 1).

The first line shows a visual representation of the basic metre – a sequence of beats of equal duration. The second line shows a rhythm that is compatible with the metre. This rhythm consists of three groups of sounds of different durations and structures. The third line introduces both rhythm and melody, where the pitch of the tones in each of the three rhythmic groups changes in distinct ways.
Rhythm consists of a perceptual grouping of a series of temporal events, as sounds and pauses of different durations (or clusters, patterns, Gestalten; see London, 2012; Overy, 2012), which are, by definition, complex and aperiodic. Thus, the hallmark of rhythm is the aperiodic groupings of events (Cummins, 2015; Turk & Shattuck-Hufnagel, 2013). Rhythm can be metrical or nonmetrical (Cummins & Port, 1998; Essens & Povel, 1985; Ozernov-Palchik et al., 2018). Metrical rhythm occurs when the rhythm is compatible with an underlying metre (See the second line in Figure 1). For instance, the interval sequence 22312114, spanning a total duration of 16 time units, can be interpreted within a metrical structure where intervals of 4 time units serve as the underlying framework (Essens & Povel, 1985). This means that the sequence can be segmented into subgroups with equal duration, such as [22][31][211][4], where each bracketed group adds up to 4 time units. Metrical rhythm typically characterises most of Western music. In this case, metre and rhythm work in synergy. However, even in music, it is not unusual to find examples of rhythms without metre, such as in Gregorian chant and recitative in operas, Baroque nonmetrical improvisation, and various forms of ethnic music. Nonmetrical rhythm occurs when an interval sequence cannot be divided into successive subgroups of equal duration, so it does not align with a metre. Essens and Povel (1985) illustrate this with the sequence 13214, which spans 11 time units (see also Ozernov-Palchik et al., 2018). Since 11 cannot be evenly divided into smaller recurring units, the sequence does not conform to a metrical structure and is an example of a nonmetrical rhythm.
Beyond some types of music, nonmetrical rhythm is typical of spoken language, which is temporally structured but does not involve an underlying grid of equal temporal units, that is, a metre (see Nolan & Jeon, 2014; Turk & Shattuck-Hufnagel, 2013). The rhythm of language can acquire a metrical periodicity as in poetry. Thus, metre and rhythm are distinct concepts, though they closely interact and complement each other. Lerdahl and Jackendoff (1993) distinguish between metre and rhythm similarly but with different terminology. They state that within rhythm, two levels must be distinguished: metre and grouping, with grouping corresponding with what we have referred to as rhythm. Throughout this paper, we use metre and rhythm for consistency, with rhythm being intended as grouping.
The distinction between metre and rhythm is helpful from a psychological point of view because it presupposes two modalities for generating and perceiving temporal behaviours. An analogy may be beneficial to introduce this difference. When one has a drawing on a sheet of squared paper, there are two ways to single out a point on the drawing: either through the cartesian coordinates in terms of the number of horizontal and vertical squares or through the indication of a specific element of the drawing, for example, the tip of the nose in a drawing of a face. This example concerns space, but an analogous situation is found in time. The squares of the sheet correspond to the metre, and the drawing corresponds to the rhythm. No musician playing in an ensemble counts the number of beats (corresponding to the cartesian coordinates in space) to establish when they have to start playing (e.g., intervene at the 132 beats), based solely on the metre. Instead, musicians rely on the musical structure of what, for example, the melody that the clarinet is playing. This example makes it clear that to plan an action (e.g., when and what to play), the best choice is not the metre (counting the beats) but the rhythm (in our example, the melody of the clarinet with its specific rhythm). In this way, musicians efficiently solve the two core problems in planning an action, that is, the serial order and timing problems (Lashley, 1951). Executing an action requires planning facilitated by rhythm (Palmer & Pfordresher, 2003). It is apparent from this description that motor planning presupposes complex cognitive skills, such as retrieval of the temporal structures (which are not just a periodic repetition of beats), anticipatory planning and, in most cases, a periodic pulsation, as a metric reference. It is unlikely that people have access to all temporal events simultaneously. Instead, as Palmer and Pfordresher (2003) pointed out, planning is incremental, so planning and producing the behaviour co-occur. In other words, as we discussed earlier in the description of coarticulation of speech sounds, planning requires one to start a subsequent action beforehand (while at the same time performing another action). During planning, one must be in an anticipatory mode and be prepared to execute the subsequent action. Thus, we use anticipatory planning or simply anticipation to refer to the ability to prepare (process information ahead) to act.
Anticipation requires focusing attention and being continuously alerted to prepare to act (Pagliarini et al., 2020). Similar processes occur when people perceive a rhythm (or process speech). As suggested by Fraisse (1982), ‘We say that we have rhythm when we can predict on the basis of what is perceived, in other words, when we are able to anticipate what will follow’ (p.150). When we hear a rhythmic structure, our perception of the stimuli that occur early in a pattern allows us to generate expectancies concerning later stimuli in real time (Martin, 1972). Rhythmic behaviour may involve coping with the metre but is more complex, and in some cases does not involve the metre, as in nonmetrical rhythms. What does this ability to cope with the metre consist of? Given that the metre underlying musical rhythm in Western music consists of a periodic succession of temporal events, people typically entrain to the beat by clapping or tapping in a periodic way along with the perceived beat (McAuley, 2010). This behaviour is called sensorimotor synchronisation (SMS), and its hallmark is periodicity, that is, both the referent and the action are periodic or regularly recurring (Repp, 2005). SMS is of paramount importance for musicians playing in an ensemble, as it allows them to synchronise their actions with the audible and visible actions of other musicians. Research suggests that SMS is controlled partly at the central level (e.g., by a timekeeper) and partly at the peripheral level through fine-motor adjustments. However, these mechanisms primarily function as perceived error correction (negative feedback loops; see Repp, 2005). Phase correction, which involves rapid adjustments to maintain synchrony with an external metre, can occur automatically and subconsciously (Repp, 2005) and does not involve the type of motor planning described earlier. Once SMS is achieved, no further planning of what or when is required, unlike in rhythmic behaviour. There is no serial order problem to solve (what), as each beat is identical to the previous one, and all beats are uniform. Similarly, there is no timing problem to solve (when), since the periodicity of the metre ensures predictability. In other words, synchronisation becomes an automatic behaviour, whereas anticipatory planning cannot become automatic (Pagliarini et al., 2020). The behaviour elicited by the metre, SMS, is typically studied using the classical synchronisation-continuation tapping paradigm, which consists of a synchronisation phase, where participants align their taps with an external periodic auditory stimulus, followed by a continuation phase, where they maintain the same periodic tapping in the absence of the auditory cue. Unlike with the metre, a continuation phase is impossible with rhythm because of its complexity and the aperiodicity of grouping events of different duration (Cummins, 2015).
In line with the literature reviewed above, we adopt this distinction between metre and rhythm, as well as between synchronisation and anticipation (or anticipatory planning). Most studies on the processing of event sequences in dyslexia, and on temporal processing in general, focus on metre and thus on synchronisation (see Kotz et al., 2018, for a review). Anticipatory planning, as we have made precise here, has been less investigated. In the next section, we review research findings on whether and how synchronisation and anticipatory planning are affected in individuals with dyslexia.
Temporal deficits in dyslexia
Converging evidence has identified temporal perception and production impairments in individuals with dyslexia and those with language impairment (for a recent review, see Ladányi et al., 2020). One of these deficits is sensorimotor synchronisation (e.g., Thomson & Goswami, 2008; Thomson et al., 2006; Wolff, 2002). One type is beat synchronisation, which is the ability to move to a steady external beat (Tierney & Kraus, 2013). Traditionally studied using a finger-tapping paradigm, beat synchronisation requires extracting the temporal pulse, tuning one’s behaviour to the stimulus, and activating an automatic, repetitive response. During tuning, behaviour is adjusted to synchronise with the auditory signal via phase error correction (Repp, 2005). Once synchronisation is established, it continues with only occasional error monitoring.
In the tapping paradigm, the synchronisation error is determined by computing the time difference between the individual’s tap and the corresponding stimulus onset and its variation over time (Chen et al., 2002). 2 Usually, while tapping in synchrony with a simple sequence of auditory tones, one displays a systematic tendency to tap their responses slightly before the signal by minus 30–50 ms (Aschersleben & Prinz, 1995; Fraisse, 1982; Repp, 2005; Repp & Su, 2013). This is known as negative mean asynchrony. In general, lower negative mean asynchrony values reflect greater synchrony, and the lower values of the associated standard deviation indicate more stability and precision (Jin et al., 2019). Negative mean asynchrony reflects an automatic tendency to act before time in a synchronisation task (Kotz et al., 2018; Repp, 2005; Repp & Su, 2013).
Beat synchronisation has been documented to be impaired in children with dyslexia (Colling et al., 2017; Flaugnacco et al., 2014; Overy et al., 2003; Thomson & Goswami, 2008; Wolff, 2002; Wolff et al., 1990). For instance, Wolff (2002) showed that 10- to 16-year-old students with dyslexia exhibited atypical mean negative asynchrony relative to neurotypical readers. When tapping with one finger to metronomic beats, the mean negative asynchrony of the dyslexic group was three to four times greater than that of neurotypical readers. Improvement was observed in both groups when tapping with two fingers, although it was not reported whether the two groups differed. When the metronome rate was altered (from 1.5 Hz to 2.0 Hz to 2.5 Hz), students with dyslexia took more time to re-tune than neurotypical readers. Wolff (2002) tentatively suggested that individuals with dyslexia may have a deficit in some components related to temporal information processing.
In a subsequent study, Thomson and Goswami (2008) employed a task that was modelled after the paradigm used by Wolff (2002) and involved tapping to metronome beats (synchronization-continuation paradigm) at three speeds: 1.5, 2 and 2.5 Hz. At each rate, children were tested in two conditions: paced (tapping along with the metronome beats) and unpaced (continuous tapping after the metronome had stopped). The absolute difference between the actual inter-tap interval (ITI; the time interval between the subject’s taps) and the expected ITI (time interval between metronome beats) was calculated. There were no significant differences between typically developing (TD) and dyslexic groups in the ITI difference scores across all rates in the unpaced condition. In the paced condition, the difference scores were significantly greater in the dyslexic group compared to the TD group at 2 and 2.5 Hz. However, no group differences were noted at 1.5 Hz. These findings indicate that children with dyslexia had difficulties entraining to fast tempos (2 and 2.5 Hz) but could still synchronise with the pattern of metronome beats at a slower tempo (1.5 Hz). Children with dyslexia did not differ significantly from TD controls in terms of the within-subject ITI variability when tapping to paced and unpaced 1.5 Hz. However, they exhibited highly variable ITIs when tapping to paced 2 and 2.5 Hz and unpaced 2.5 Hz. The inconsistent within-subject ITIs suggest that timing precision is impaired in dyslexic children.
All these findings indicate that children with dyslexia can synchronise at a slow tempo, but their difficulties are evident at faster tempos and in their timing precision. In this study, the reported difficulties could not be explained by motor control or execution difficulties as children with dyslexia were comparable to their TD peers in manual dexterity, as indexed by their scores on the pegboard task. Moreover, their scores on this task did not correlate with any of the tapping indices discussed above (Thomson & Goswami, 2008). Thus, synchronisation is not always impaired. In addition, the lack of a group difference in the unpaced tapping condition indicates that the problems experienced by dyslexic individuals are evident only when there is an external stimulus. Bégel et al. (2022) extended these findings. In a paced tapping task at 100 beats per minute (1.7 Hz), they found that children with dyslexia were poorer than controls in synchronising with the beat. This supports Thomson and Goswami’s (2008) findings, which also highlighted difficulties in synchronising with faster tempos (2 Hz) in children with dyslexia. Bégel et al. (2022) further tested children’s ability to adjust their tapping to faster or slower tempos. They found that children with dyslexia did not show differences from controls in their motor adaptation during tapping performance. However, these children had significant difficulties in perceiving tempo changes. Specifically, they were poorer than controls in their metacognitive awareness of tempo changes, mainly when the changes were large. Notably, the children with dyslexia in Bégel et al.’s (2022) study showed poorer manual dexterity (as measured by the pegboard task) than controls. However, Bégel et al. (2022) reported that their synchronisation difficulties were still observed even after controlling for manual dexterity, indicating that the synchronisation deficits were not simply due to poor fine motor skills.
Using the same paradigm, Thomson et al. (2006) documented a different performance pattern in adults with dyslexia. Dyslexic adults were comparable to controls in terms of the expected-actual ITI difference scores across all rates in both paced and unpaced conditions. The mean negative asynchrony times of both groups did not differ significantly. This suggests that they could tap and entrain to internally generated rhythms. On the other hand, adults with dyslexia showed greater ITI variability than controls when tapping at 2.5 and 2 Hz in paced conditions and at 2 Hz in unpaced conditions. This suggests that, similar to children with dyslexia, adults with dyslexia have deficits in timing precision. As with children, these deficits are evident when tapping at fast tempos but not at slower ones. Thomson et al. (2006) observed a significant difference in dominant-hand manual dexterity (measured by the pegboard task) between dyslexic and non-dyslexic adults. However, further analysis found no significant correlation between manual dexterity scores and any of the tapping variables. The authors suggested that motor execution difficulties did not account for the synchronisation difficulties observed in dyslexic adults. This evidence indicates that factors beyond motor control difficulties contribute to the synchronisation deficits observed in adults and children with dyslexia.
In sum, the evidence on beat synchronisation abilities in individuals with dyslexia confirms that a deficit in timing precision is evident at fast rates but not at slower rates. In addition, Wolff (2002) showed that individuals with dyslexia had difficulties when the tempo changed, and they had to adjust their behaviour to the new tempo. Finally, Bégel et al. (2022) reported that children with dyslexia had poor metacognitive awareness in reporting tempo changes. Some synchronisation skills are generally preserved in adults and children with dyslexia, although this is primarily the case at slow tempos (1.5 Hz) for children.
Given that individuals with dyslexia exhibit some temporal difficulties – such as imprecise timing and limited metacognitive awareness of tempo changes – it is plausible that their impairments are evident in anticipation. The subsequent discussion presents empirical support for this proposition.
Impairments in anticipation in children and adults with dyslexia were investigated for the first time by Pagliarini et al. (2020) using the warning-imperative paradigm (Walter et al., 1964). This paradigm comprises two stages, as illustrated in Figure 2. In the familiarisation phase, participants listen to a periodic sequence of beats, where no action is required, thus allowing them to get acquainted with the metre. In the testing phase, the same sequence of beats is presented again, but this time it includes a minimal rhythmic structure. This structure is obtained by randomly presenting, in the metrical sequence, ten pairs of adjacent beats auditorily different from the other beats. In each pair, the first beat is a warning beat (WB). Participants do not have to act when the WB occurs. The WB cues the arrival of the second beat, that is, the imperative beat (IB), and participants are asked to tap in time to the IB beat. Hence, the WB puts the participants in an anticipatory mode to prepare them to act to the IB.
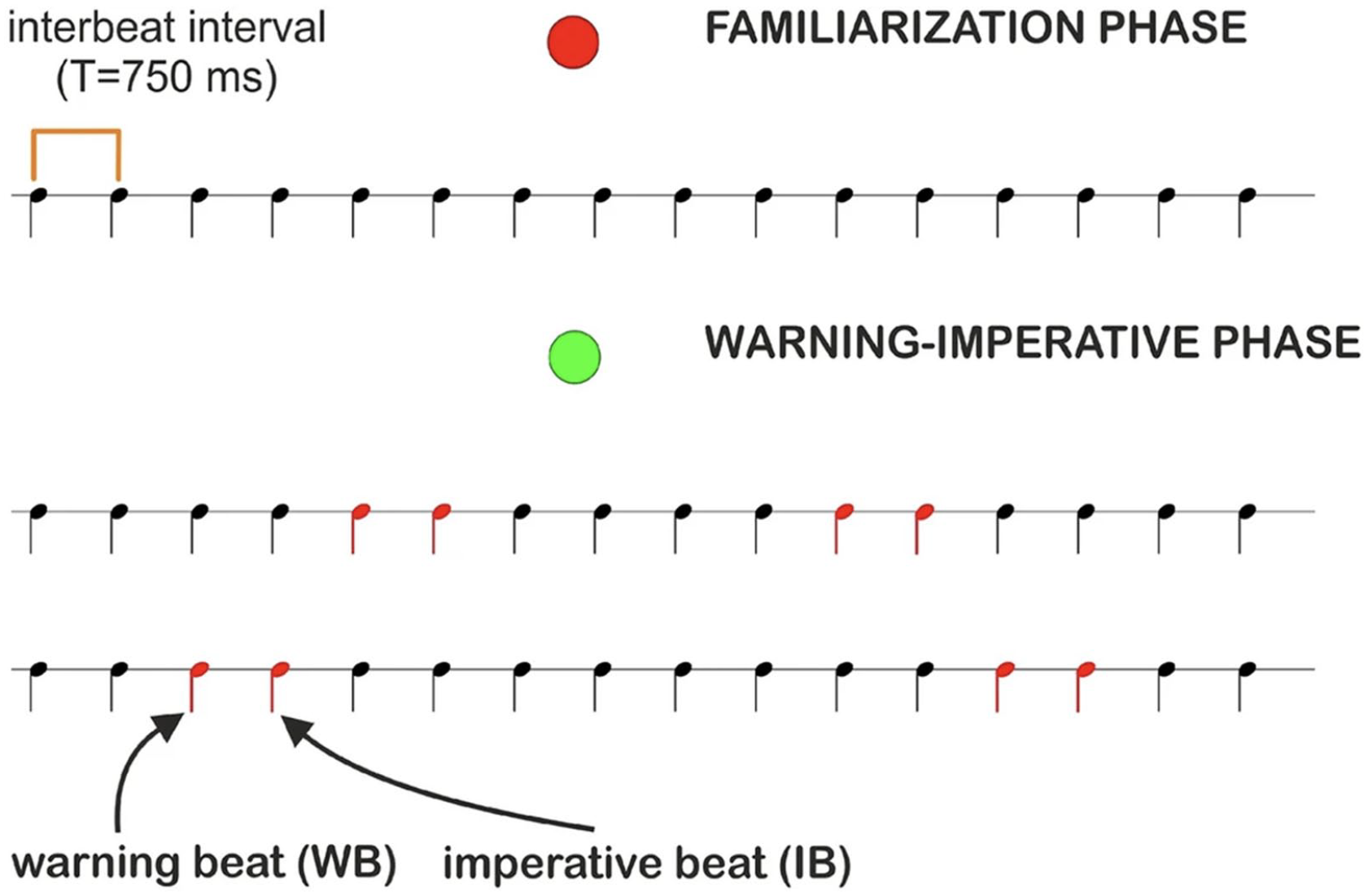
Procedure of the anticipatory timing task. For each condition, participants were presented with a familiarisation phase (preceded by a red dot), followed by a warning-imperative phase (preceded by a green dot). During the familiarisation phase, participants were asked to listen attentively to the sequence (no action was required). During the warning-imperative phase, participants were presented with the sequence they were previously familiarised with and were asked to tap the left mouse button in sync with the imperative beat (IB). In the figure, the warning beat (WB) and IB are marked in red for illustration purposes; in the experiment, both were obtained by adding the first overtone to the basic beat used to present the periodic pattern. Reprinted from “Timing anticipation in adults and children with Developmental Dyslexia: Evidence of an inefficient mechanism,” by E. Pagliarini, L. Scocchia, E. Granocchio, et al., 2020, Scientific Reports, 10, Article 17519 (https://doi.org/10.1038/s41598-020-73435-z). CC BY 4.0.
This paradigm allows for the explicit examination of anticipation based on an elementary rhythmic structure (randomly presented groupings of two beats) compatible with a metre (metrical rhythm). Since participants have been familiarised with the periodic sequence of beats, once they hear the WB, they should be able to anticipate when the IB will occur. Participants do not know when the 10 pairs of WB and IB will appear; they only know that the appearance of the WB predicts the occurrence of the IB. Therefore, they must be in an anticipatory mode, ready to act (i.e., press a button) in response to the IB. In Pagliarini et al. (2020), there were three testing conditions. In Condition 1, plain unstressed metronome beats were presented. In Condition 2, alternating strong/weak beats were presented. In Condition 3, an unpredictable sequence of beats was presented, that is, the control condition. Adults were tested across all three conditions, whereas children were tested only in Conditions 1 and 3. To measure anticipatory skills, the timing error was calculated as the difference between the observed response time to the IB and the expected IB time. A negative error indicates an early response, while a positive error indicates a delayed response. A zero difference suggests that the participant responded at the exact time of the IB. Figure 3 presents the performance of adults and children with and without dyslexia on the warning-imperative task. In Conditions 1 and 2, adult controls could anticipate the IB and press the button on time or slightly before its occurrence. In contrast, adults with dyslexia were systematically delayed, that is, they tended to press the button after the IB had occurred (see panel A in Figure 3). It is worth pointing out that the delay observed in adults with dyslexia was not merely a reaction time delay, as it was less than 150 ms, which is the average reaction time.
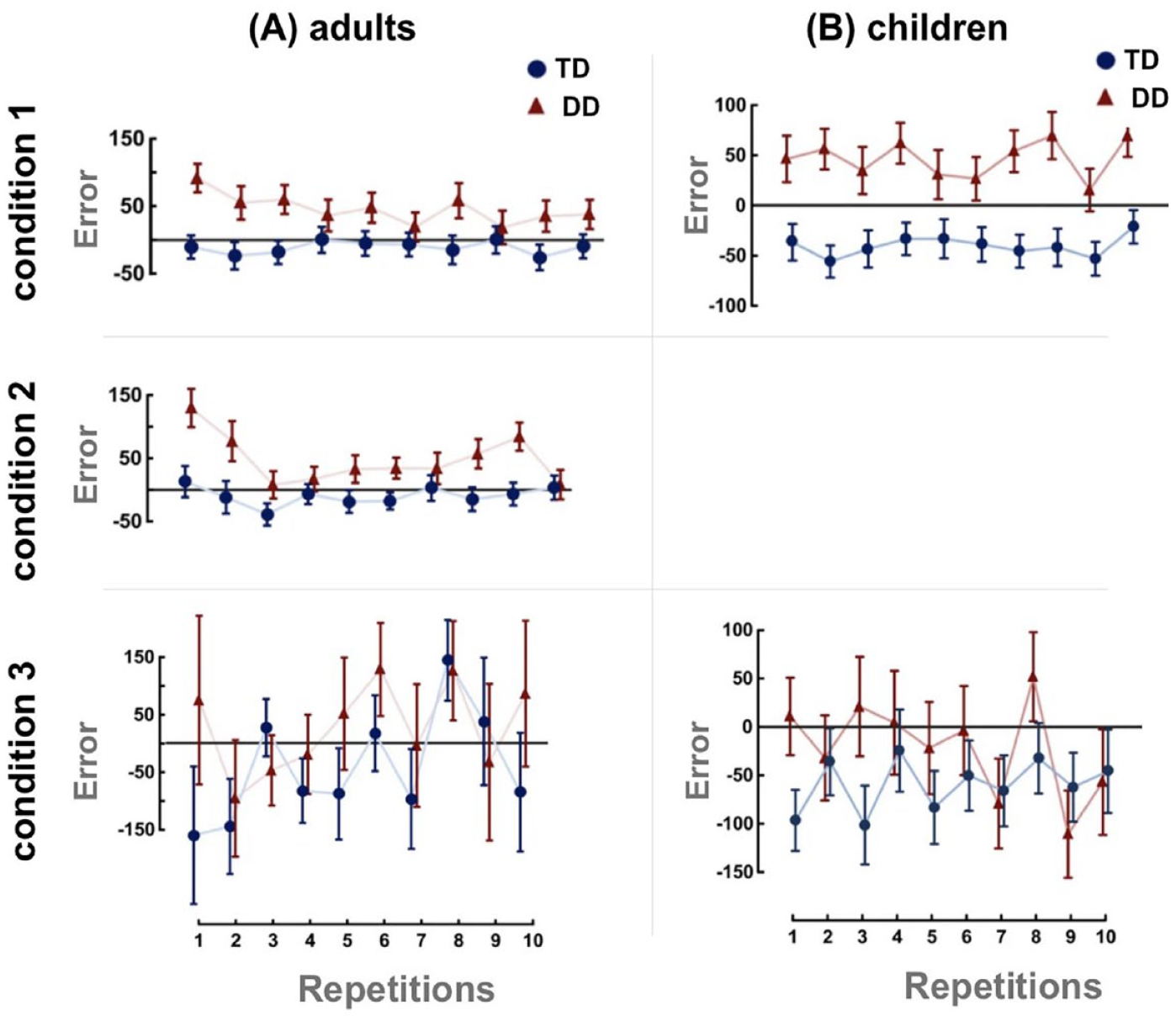
Results from typically developing (TD) and developmental dyslexia (DD) groups for the warning-imperative task. Panel A presents timing error results for adult participants in Condition 1 (unstressed), Condition 2 (stressed) and Condition 3 (unpredictable). Panel B presents results for child participants in Condition 1 (unstressed) and Condition 3 (unpredictable). The interaction Group × Repetition is reported for all conditions. The x-axis represents the mean of the ten repetitions, while the y-axis represents the timing error, which is calculated as the difference between the observed time—that is, the participant’s response – and the expected time – that is, the Imperative Beat (IB). The horizontal line at zero represents the IB time. Vertical bars indicate the standard error. Adapted from “Timing anticipation in adults and children with Developmental Dyslexia: Evidence of an inefficient mechanism,” by E. Pagliarini, L. Scocchia, E. Granocchio, et al., 2020, Scientific Reports, 10, Article 17519 (https://doi.org/10.1038/s41598-020-73435-z). CC BY 4.0.
This fact suggests that adults with dyslexia attempted to execute the action at the right moment but were inefficient. Interestingly, no between-group differences were observed in the timing error in the control condition (Condition 3), where the irregular rhythm did not allow for anticipatory planning. When no anticipation was possible, the speed of responses was similar in both groups. Therefore, Pagliarini et al. (2020) argued that slow responses could not explain the group differences in the other two conditions, that is, Conditions 1 and 2. The same pattern of results was found in children (see Panel B in Figure 3). In Condition 1, children with dyslexia were delayed, while control children tapped slightly before the IB. In Condition 3, the two groups did not differ. Finally, children and adults with dyslexia were significantly less precise in their responses across trials compared to TD peers. Collectively, the results above provide a basis for proposing the Inefficient Anticipatory Planning Hypothesis. Children and adults with dyslexia are poor at anticipating when they have to act, that is, processing information in advance (Pagliarini et al., 2020). Therefore, they struggle with the warning-imperative task, which taps precisely into the ability to anticipate.
Anticipatory planning in phenomena that unfold in time
The concept of ‘anticipatory planning’ is widely used in the motor control literature (e.g., Rosenbaum, 2009). Here, we propose that it is a specific form of prediction which can be facilitated by temporal organisation (rhythm). We also propose that anticipatory planning may, in some cases, be a component of the broader predictive process, such as when predicting the next word in a sentence based on structural constraints (e.g., a determiner predicting the presence of a noun). However, anticipatory planning can be entirely absent from the process of prediction, as in predicting future events (e.g., ‘tomorrow it will rain’). Accordingly, our evidence concerning difficulties in processing rhythmic structures (groupings of events) is not restricted to anticipatory planning but may extend to prediction more broadly. Our view is that while rhythm can facilitate anticipatory planning by providing a temporal structure, anticipation itself may not always or solely depend on rhythm. Instead, rhythm enhances predictability in contexts where temporal regularities are present; however, we acknowledge the possibility that other mechanisms may also support anticipatory planning.
Guasti et al. (2018) proposed that individuals with dyslexia are less efficient in exploiting structural regularities across oral language, handwriting, and reading, suggesting that temporal organisation plays a crucial role in these abilities. Should our hypothesis prove valid, we expect individuals with dyslexia to exhibit difficulties in other tasks or domains requiring anticipatory planning such as language, speech, reading and handwriting. So, what is the rhythmic structure underlying these phenomena? In the following sections, we will outline this structure in each case and review, where available, behavioural evidence from studies involving individuals with dyslexia who display difficulties in coping with ‘rhythmic’ structures.
Prediction and anticipatory planning in language and speech processing
Language rhythm has been extensively investigated over the last 90 years. In linguistics, it has been proposed that speech is organised based on the hierarchical ordering of phonological constituents, where phonemes combine to form syllables, syllables are grouped into feet, feet are grouped into prosodic words, which are combined into phonological phrases, forming intonational phrases that result in utterances (Hayes, 1989). This organisation is called the prosodic hierarchy. Similarly, in grammar, morphemes combine to form words, words combine to form phrases, and phrases combine to form sentences. These two hierarchies are closely related (e.g., Nespor & Vogel, 2007, pp. 49–135), suggesting that morpho-syntactic structures can be inferred from prosody (see Christophe et al., 2003; Nespor et al., 2003).
These groupings of linguistic events are the basis of the rhythmic structure of language (a view independently suggested by Arvaniti, 2009). Along with prominence and pauses, they are used during sentence production and processing to anticipate and predict what will come next (Altmann & Mirković, 2009; Anderson & Carlson, 2010; Breen, 2014; Grosjean, 1983; Huettig, 2015; Kuperberg & Jaeger, 2016; Mani & Huettig, 2012; Nakamura et al., 2012; Pickering & Gambi, 2018; Rivolta et al., 2021). This is also supported by evidence linking musical rhythmic processing to receptive and expressive grammar, as well as phonological processing (Gordon, Jacobs, et al., 2015; Gordon, Shivers, et al., 2015; Kim et al., 2024; Nitin et al., 2023), highlighting shared hierarchical and temporal structures in music and language.
It is important to note that, although language has a temporal organisation, speech rhythm is neither isochronous nor metrical, according to the terminology we adopted (Nolan & Jeon, 2014; Patel, 2007). However, as Nolan and Jeon (2014) point out, this lack of isochrony/metre does not prevent speakers from exercising anticipation. This is clear in coarticulation phenomena (discussed previously), speech errors (see Shattuck-Hufnagel, 2015, for a review), and the regulation of timing in speech production, with speakers adjusting the length of segments and syllables to prevent excessively long or short linguistic units (Jun, 2005).
Research on language production shows that speakers plan at least two or three words in advance and structure their speech into intonational-phrase-sized chunks, as defined by the prosodic hierarchy mentioned above (roughly corresponding to a sentence; see Shattuck-Hufnagel, 2015). Planning ahead is evident in several linguistic phenomena. For example, adjustments in average syllable duration are determined by phrase length (Bishop & Kim, 2018). Similarly, the placement of heavy noun phrase (NP) direct objects at the end of sentences in English (as in ‘John gave to Lucy the carpet that I bought in Morocco’) depends on their relative length and duration in relation to other phrases in the utterance (Stallings & MacDonald, 2011). These examples illustrate that speech production involves planning both what is said and when, relying on language’s temporal organisation and rhythm, including grouping, duration, pauses, and prominence.
Despite the evidence for anticipatory planning, further research is needed to clarify the precise relationship between prosodic structure and syntactic as well as semantic organisation, and how these elements interact during language production and comprehension, potentially enhancing anticipatory planning. These facts are relevant in light of the finding that individuals can predict upcoming words during sentence processing. Research shows that people can use structural, prosodic, and contextual information to probabilistically pre-activate semantic, morpho-syntactic, and phonological representations of upcoming words, unconsciously predicting upcoming linguistic input (e.g., Altmann & Mirković, 2009; Nation et al., 2003).
Evidence for prediction during language processing comes from eye-tracking studies using the visual world paradigm. These studies show that both children (Bobb et al., 2016; Fernald et al., 2010; Lukyanenko & Fisher, 2016; Mani & Huettig, 2012; Nation et al., 2003) and adults (Altmann & Kamide, 1999; Coco et al., 2016; DeLong et al., 2005; Staub et al., 2012) generate language-modulated anticipatory eye movements. For instance, Altmann and Kamide (1999) examined the eye movements of adult listeners while they viewed four referents on a screen and heard spoken sentences containing either an informative verb (e.g., ‘The boy will eat the cake’) or a neutral verb (e.g., ‘The boy will move the cake’). When the sentence included an informative verb, adults looked at the target object before it was heard, suggesting they could use lexical-semantic verb information (e.g., ‘eat’) to predict the upcoming noun (e.g., ‘cake’). This pattern has also been documented in 2-year-old children (Mani & Huettig, 2012). Additionally, Lukyanenko and Fisher (2016) showed that, while viewing plural and singular referents (e.g., two cookies and one apple), 3-year-old children could use morpho-syntactic information, that is, number markings (e.g. ‘Where are the good cookies?’ vs. ‘Where is the good apple?’) to predict upcoming referents accurately. These findings indicate that during online sentence processing, adults and children can exploit lexical-semantic and morpho-syntactic cues to generate predictions about the words they will encounter before encountering them. 3
These predictive behaviours are reflected in a motor response – eye movements – demonstrating an anticipatory planning behaviour that is not directly expressed within language but is guided by spoken input. Given the evidence on language production concerning the role of anticipatory planning facilitated by rhythm, we speculate that similar rhythmic mechanisms are engaged during comprehension. This perspective allows for a connection between the poor anticipatory planning observed in dyslexic individuals in the warning-imperative task and their difficulties in language processing. Although this remains a hypothesis requiring further empirical support, it provides a promising framework for interpreting the findings discussed and opens new avenues for research. Although we see what follows as something that can be integrated into an anticipatory planning behaviour, the term prediction will be used throughout in line with existing literature.
A few studies have examined predictive language processing in dyslexic individuals (Engelhardt et al., 2021; Huettig & Brouwer, 2015; Persici, 2019) using the visual world paradigm. Huettig and Brouwer (2015) were the first to investigate language-mediated anticipatory eye movements in adults with and without dyslexia. In the first experiment, Dutch-speaking participants heard a target word (e.g., ballon ‘balloon’) while viewing four objects: a shape competitor (e.g., zon ‘sun’), a phonological competitor (e.g., bad ‘bath’), a semantic competitor (e.g., pop ‘doll’) and a distractor (e.g., deur ‘door’). Participants were asked to indicate whether the target word was present on display or not. As information from the spoken target word unfolded, it was found that gaze fixations on the three competitors diverged significantly from fixations to the unrelated distractor for both groups. This confirmed that adults with dyslexia showed language-mediated eye gaze patterns like TD controls.
In the second experiment, participants were instructed to listen to simple sentences (e.g., kijk naar de afgebeelde piano ‘look at the piano’) while viewing four referents: a target (e.g., piano) and three distractors. The sentences varied in terms of the gender-marked definite determiner they contained: the determiner ‘de,’ which typically precedes common nouns (e.g., de piano ‘the piano’), or the determiner ‘het,’ which precedes neuter nouns (e.g., het paard ‘the horse’). In each trial, the determiner agreed on gender only with the target. Hence, participants could use the gender information carried by the determiner to predict the target object before it was heard. Results revealed that TD controls shifted their gaze to target 400 to 600 ms after the onset of the determiner, whereas the dyslexic adults shifted their gaze to target approximately 1,400 to 1,500 ms after the determiner onset. These patterns are illustrated in Figure 4. As evident there, the onset of the noun is around 2,000 ms, but eye gaze to the target noun occurs before both in adults with and without dyslexia. These findings indicate that adults with and without dyslexia were able to exploit gender information on the determiner to predict the noun of the target object well in advance of its acoustic onset. However, eye gaze to the target picture depicting the relevant noun was delayed by about 1,000 ms in participants with dyslexia compared to adult controls. Huettig and Brouwer (2015) excluded the possibility that the delayed eye gaze in adults with dyslexia is due to slower eye movements. This was supported by the results from their first experiment, which confirmed that adults with dyslexia were as fast as the control adults in modulating their eye gaze according to the linguistic input when single words were uttered.
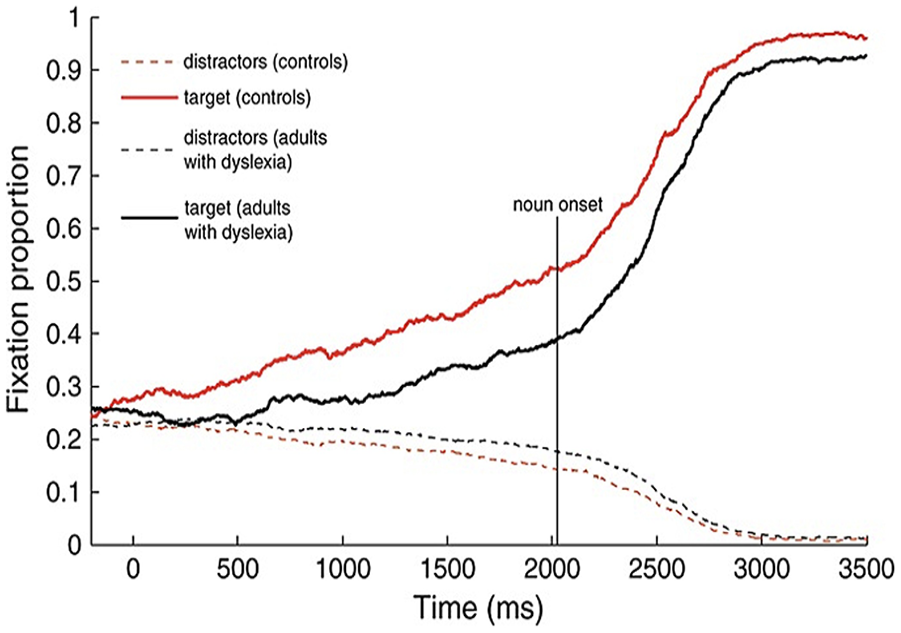
Time-course graph of the proportions of fixations to the target objects specified in the instruction and the unrelated distractors, plotted from the acoustic onset of the spoken article (‘de’ or ‘het’). The dotted lines represent the average proportions of looks to the distractors. The proportions reflect the number of trials in which participants fixated on each type of picture on the screen. Time zero represents the acoustic onset of the article. Reprinted from “Delayed Anticipatory Spoken Language Processing in Adults With Dyslexia – Evidence From Eye-Tracking,” by F. Huettig and S. Brouwer, 2015, Dyslexia, 21(2), 97–122 (https://doi.org/10.1002/dys.1497). Copyright 2015 by John Wiley & Sons. Reprinted with permission.
Several interpretations of Huettig and Brouwer’s (2015) findings are possible. It could be that delayed anticipatory eye movements arise from a difficulty processing or recognising the critical gender-marked determiners ‘de’ and ‘het,’ which tend to be phonologically underspecified in Dutch. This could make it challenging for individuals with dyslexia to use these determiners as predictive cues efficiently. Interestingly, Huettig and Brouwer (2015) found that word reading, but not nonword reading, predicted anticipatory eye gaze. Accordingly, they interpreted the delayed eye gaze in adults with dyslexia not as a primary deficit but as secondary to reduced literacy exposure. This secondary deficit results in reduced precision/sharpness of lexical representations, which consequently leads to a slower retrieval (for a discussion on how literacy enhances prediction, see Huettig & Pickering, 2019). Therefore, delayed eye gaze in dyslexic adults may also be due to difficulty retrieving the word associated with the target picture.
It is essential to note that word reading only accounted for 8% of the variance in the language-mediated anticipatory eye movement. This relatively small coefficient indicates that factors other than reduced literacy exposure may account for the delay in dyslexic adults. In addition, lexical representations were also involved in the first experiment where single words were presented, yet adults with dyslexia did not differ from controls. It is only when there is a sequence of words and one is predictive of the upcoming word that dyslexic adults have difficulty. 4 Due to these reasons, we propose an alternative interpretation to Huettig and Brouwer’s (2015) data that warrant consideration. We hypothesise that delayed anticipatory eye-gaze may be due to poor anticipatory behaviour, a primary deficit in individuals with dyslexia, as already demonstrated in the studies discussed earlier. The article and the following noun must form a syntactic and prosodic group, with the article typically being a prosodically weak element, of short duration and the noun bearing the stress, creating a ‘rhythmic unit’. Distinguishing between our hypothesis, which links delayed anticipatory eye-gaze to poor anticipatory skills, and Huettig and Brouwer’s interpretation – which attributes it to literacy exposure – requires further empirical evidence. A longitudinal study is needed to examine how these anticipatory processes develop in children before they are exposed to literacy (see also the section on handwriting). We do not deny that reduced literacy may further impact anticipatory skills in predicting words. However, we claim that anticipatory planning skills are already impaired before exposure to literacy, as supported by the data reviewed earlier. This impairment may explain the delay in guiding eye movements based on morpho-syntactic information from the determiner to predict the upcoming noun, that is, the morpho-syntactic cues are not used efficiently to rapidly process information ahead to create expectancies about the following noun and guide eye movement. Notice that the studies by Pagliarini et al. (2020), as well as those by Huettig and Brouwer (2015), did not show that individuals with dyslexia cannot display anticipatory planning and prediction, but rather that they are delayed compared to controls.
The evidence we discussed suggests that anticipation is facilitated by the rhythmic structure of language, which is linked to its morpho-syntactic organisation. However, semantic relationships can also guide prediction, which may not directly rely on rhythm. It remains unclear whether individuals with dyslexia will show similar difficulties in semantic-based prediction. Further research is needed to clarify mechanisms that facilitate anticipatory planning in language processing, and how they are affected in individuals with dyslexia.
Anticipatory planning in reading
In reading, words are grouped together, such as pairing a determiner with its noun and inserting pauses, with grouping being the main feature of rhythm. Research shows that, during reading, the duration of a pause within an utterance depends not only on the length of the preceding phrase but also on the length of the following one (Krivokapić, 2007). Additionally, silent reading has been linked to inner speech (e.g., Abramson & Goldinger, 1997; Dell & Repka, 1992), reinforcing its connection to spoken language. Reading does not have a clear rhythmic structure. However, its organisation shares some features with spoken language, particularly in how linguistic elements are processed in a sequential manner and are grouped together. This suggests that some degree of anticipatory planning is involved. Whether this process is supported by rhythm, as in spoken language, is an open question. In this section, we examine the evidence for anticipatory planning in reading, acknowledging that the underlying mechanisms require further exploration.
Readers make a series of rapid saccadic eye movements along an array of written text while reading. These saccades alternate with fixations during which new visual information is extracted from the text (Rayner, 2009). The amount of visual information obtained during a single fixation, the perceptual span (Jacobs, 1986; Rayner, 1998), is a standard measure of foveal and parafoveal processing (Sperlich et al., 2016) and is asymmetric to the direction of reading (McConkie & Rayner, 1976). For instance, in English and other languages that read from left to right (e.g., French and Dutch), the rightward extension of the perceptual span is larger than its leftward extension. In these languages, the perceptual span of skilled readers is approximately 19 letters (3–4 letters to the left of the fixation point and 14–15 letters to the right; see Rayner, 1998; Schotter et al., 2012). The parafoveal preview of the subsequent text on the right of a given fixation point allows one to process, among other things, information about word length and inter-word spaces and thus is essential for guiding eye movements on the text and plan where the eye will fixate next. As such, it may be evidence of anticipation (Schotter et al., 2012). That is, while fixating on a particular word, the eyes of the reader scout further ahead and process the following letters, thus forming some representation of them that is useful to read the upcoming words fluently, even without fixating on them. In other words, parafoveal processing alleviates the processing of future words. It may reduce it to a match between an internal representation and what is written in the text because predictions are correct in most cases.
The parafoveal preview has long been known to be impaired in individuals with dyslexia (e.g., Bouma & Legein, 1977); for example, Zoccolotti et al. (2013) suggested that individuals with dyslexia do not benefit from parafoveal processing based on Rapid Automatized Naming (RAN; Denckla & Rudel, 1976). They found that participants with dyslexia perform more poorly when naming sequences of stimuli (e.g., digits, colours in RAN) than when single stimuli are presented. To fluently name sequential material, one has to name a symbol while one’s eyes are already processing symbols ahead through parafoveal preview. In other words, in serial RAN, one must engage in anticipatory behaviour – that is, while naming symbol n, one must pre-process symbol n+1. This is not the case in single-stimuli tasks.
Further evidence for anticipatory planning in reading can be illustrated by data on the coordination of eye movement and articulation in reading aloud. The eye-voice span (EVS) measures the distance or the amount of time the eyes are ahead of the voice during oral reading (Buswell, 1921). Eye-tracking studies show that the reader’s eye fixation on a given word always precedes their articulation/production (Buswell, 1921; Inhoff et al., 2011; Laubrock & Kliegl, 2015). This phenomenon is illustrated in Figure 5 in children with typical reading. This figure shows the fixations on the text (A) and the simultaneously uttered phonemes (B). It can be observed that when the eyes fixate on the fourth word, ‘dal’, for example, the voice still pronounces the first word, ‘un’; that is, the eyes are ahead of the voice. Buswell (1921) was the first to investigate the spatial EVS and found that the eye fixations of mature readers are 15 to 17 letters (or two to three words) ahead of the voice position (e.g., the position of the articulated word). More recently, Laubrock and Kliegl (2015) described EVS in a group of university students. They found that when a reader pronounces a given word, their eyes will be 10 to 16 letters (two to three words) ahead. This suggests that reading (aloud) a given word and the upcoming words occur simultaneously. In other words, to read fluently, one must plan in advance the eye movements and be ready to process the words ahead while reading aloud a previous word.
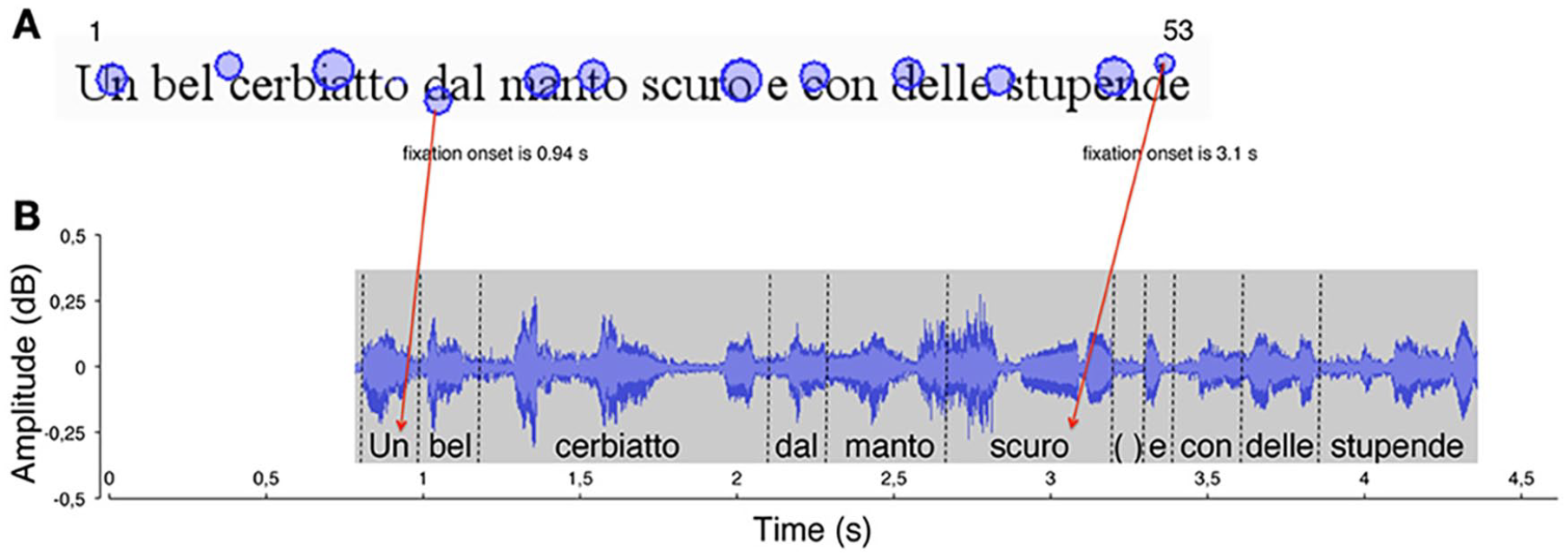
An illustration of the eye-voice span measured in letters in typically developing readers. In panel A, the circles represent the locations of eye fixations, with their sizes proportional to fixation duration. The numbers 1 and 53 indicate, for example, the first and last letter position numbers in the line. The onset time of two fixations is marked. In panel B, the waveform of reading aloud is shown as a function of time. The vertical dotted lines delimit single-word utterances labelled along the timeline. Red arrows indicate the temporal correspondence between a fixation (e.g., the grapheme ‘d’ in the word dal, letter number 18) and the phoneme uttered at the time of fixation (the phoneme/u/, letter number 1). Each fixation (e.g., ‘d’ in the word stupende, letter number 52) was linked to the phoneme uttered at the moment of fixation onset (the phoneme /o/ in the word scuro, letter number 32). The eye-voice span was calculated by subtracting the phoneme position number from the grapheme position number for all grapheme-phoneme pairs (in this example: 52–32 = 20 letters). Reprinted from “The Eye–Voice Lead During Oral Reading in Developmental Dyslexia,” by M. De Luca, M. Pontillo, S. Primativo, D. Spinelli, and P. Zoccolotti, 2013, Frontiers in Human Neuroscience, 7, 696 (https://doi.org/10.3389/fnhum.2013.00696). CC BY 3.0.
In our view, EVS could be considered a window into an individual’s predictive/anticipatory processes. It shows that readers process upcoming words while they pronounce a previous word. This information contributes to reading fluency by reducing the number of fixations during reading. The finding that the EVS predicts regressions, refixations, and fixation durations in skilled readers supports this notion (Laubrock & Kliegl, 2015). De Luca et al. (2013) evaluated the EVS in 11-year-old Italian children with and without dyslexia by examining several voices, eye movements and reading parameters. Children with dyslexia were slower and less accurate than control children in reading a meaningful text. Their slower reading pattern was characterised by more frequent and long silent pauses, sounding-out behaviours (fragmented reading pattern, with hesitations), and significantly longer word pronunciation time. Moreover, children with dyslexia exhibited more fixations, shorter forward saccades and a higher proportion of regressions than typically developing children (De Luca et al., 2013). The duration of fixations did not distinguish the two groups. In children with and without dyslexia, the eyes led the voice by a relatively constant time/space across the reading task, suggesting a consistent eye–voice lag in both groups. This indicates that both groups exhibit anticipatory behaviour. However, the amplitudes of the EVS were significantly narrower in children with dyslexia with respect to controls, which aligns with our hypothesis that the anticipatory planning process is less efficient in individuals with dyslexia. When uttering a particular phoneme in each word, the eyes of children with dyslexia were, on average, eight letters ahead of their voice. In contrast, the eyes of typically developing children were, on average, 14 letters ahead. Thus, the number of upcoming letters children with dyslexia could visually scan during oral reading was half that of control children (De Luca et al., 2013). These results extended earlier observations showing that the EVS is smaller in poor readers (Buswell, 1921; Fairbanks, 1937).
De Luca et al. (2013) suggested that the reading deficit in children with dyslexia stems from a single predominant deficit in word decoding that adversely affects all subcomponents of reading. These authors proposed that children with dyslexia are very slow in decoding written words, and thus, the voice output flow occurs close to decoding, and the distance between the voice and the eyes is very short. However, the described behaviour of children with dyslexia is also consistent with our proposal that these children have poor anticipation skills, as we have seen in previous sections, and thus are delayed in preparing to process (fixating) upcoming words while uttering a previous word. Therefore, they display more fixations and regressions than neurotypical children.
Support for our view comes from RAN, which we discussed earlier, as it does not call for many decoding skills. As Carioti et al. (2022) have shown, minority-language children are poor at reading, but their performance at RAN is indistinguishable from that of monolingual typically developing children. By contrast, poor readers obtained lower scores in reading and underperformed at RAN. Interestingly, as suggested, RAN includes some sub-tasks involved in reading: reading aloud and pre-processing a word ahead. Our interpretation of these data (a narrow EVS and a poor parafoveal preview) aligns with what we found in Pagliarini et al. (2020) with the warning-imperative task, and Huettig and Browers’ (2015) study. It is likely that the same poor anticipatory planning deficit is present in reading.
To conclude, we hypothesised that individuals with dyslexia are poor at anticipatory planning that utilises rhythm. The evidence we discussed does not prove that rhythm is involved in reading, but some data that we presented (reading involving inner speech and planning pauses based on the length of upcoming phrases) evoke the possible role of rhythm in reading. Ours is a plausible hypothesis that should be further studied in the interest of finding a common thread between reading, language and writing, a topic that we will address later.
Anticipatory planning in the rhythmic organisation of handwriting
Anticipatory planning is fundamental in behaviours that require precise timing and sequential ordering of behavioural patterns. Handwriting is one such behaviour, and as such, it is ruled by a rhythmic structure that we describe below. Handwriting is based on coarticulation in a broader sense, as the way we write a given letter is influenced by the following letter (Kandel & Perret, 2015). In cursive handwriting, we generate a continuous series of letters that vary in shape, size and stroke direction. Evidence suggests that the temporal pattern of writing a given letter is determined by the spatial and motor constraints of the adjacent letters (e.g., Kandel et al., 1993; Kandel & Perret, 2015; Orliaguet et al., 1997; Thomassen & Schomaker, 1986). For instance, Kandel and Perret (2015) asked 8-year-old children to write the letter l in three contexts (ll, le and ln in cursive script). They found that the upstroke duration for the l of le was longer than in ll and ln; the latter yielded equivalent movement times. Furthermore, the downstroke duration of l was shorter for ll than le and, in turn, shorter than ln. In general, the l downstroke durations increased as the spatial parameters to anticipate increased: ll < le (size change) and le < ln (direction change; Kandel & Perret, 2015). This is evidence that the motor system anticipates the topological changes of the subsequent letter, which translates to modifying the timing of the current production.
To write fluently, one needs to know what one has to do, that is, which is the following letter and its shape. One also has to carefully plan when the next letter should start, that is, the duration of the current letter relative to the next one. In other words, handwriting involves sequencing discrete movements in a timely manner. Accordingly, at least two rhythmic principles regulate motor timing in handwriting: isochrony and homothety. The principle of isochrony indicates that the speed of movement is proportionally tied to the linear extension (length) of its trajectory to keep the movement execution time more or less constant or of equal duration (to double the total duration of the writing movement the word size must be increased by at least 20 times; Flash et al., 2013; Stetson & McDill, 1923; Viviani & Flash, 1995; Viviani & McCollum, 1983). In other words, average movement speed increases as a function of its length. This compensatory mechanism characterises a variety of human motor acts such as drawing and handwriting (e.g., Freeman, 1914; Viviani & McCollum, 1983), typing (Viviani & Terzuolo, 1982), and hand and arm movement (Jeannerod, 1984; Yokoyama et al., 2018). Studies investigating the motor timing of handwriting have revealed that the total word writing duration remains approximately constant irrespective of changes in the word size (Freeman, 1914; Stetson & McDill, 1923; Viviani & Terzuolo, 1982). For instance, writing a bigger word is accompanied by increased handwriting speed (Viviani & Terzuolo, 1982). That is, when asked to write bigger, one would spontaneously write faster. According to the principle of homothety, the fraction of time devoted to the individual letters of a word remains constant across changes in the global writing duration of a word (Viviani & Terzuolo, 1982). To illustrate, consider the word ‘dog’. Let us assume that on an average size, one would spend 40% of the total time writing the letter ‘d’, 10% of the time writing the letter ‘o’, and 50% writing the letter ‘g’. When asked to write the word ‘dog’ bigger than the average size, the proportion of motor time spent writing each letter will remain the same, even if the total duration may slightly increase. This division of the duration in handwriting is the rhythm of word writing and is independent of changes in the total duration and size of the word. Thus, because writing is governed by rhythm (homothety), when the size of the word changes, a rescaling operation must be accomplished to respect the proportion of time dedicated to each letter.
In summary, handwriting is a rhythmic behaviour ruled at least by two rhythmic principles. The isochrony principle governs the global duration of writing a word. It states that the time to write a word at different sizes tends to remain constant by adjusting handwriting speed according to changes in size. The homothety principle governs the relative duration of the individual letters within a word. It states that the relative duration devoted to the individual letters within a word remains constant across changes in the size and total duration of the word.
If anticipation skills are poor in individuals with dyslexia, it is predicted that they will be unable to comply with the two rhythmic principles ruling anticipation in handwriting: isochrony and homothety. They will be poor at maintaining the relative and global duration constant across changes in size because to do this, they must anticipate or process the duration of the following letter relative to the one they are writing and have to decide when they have to start writing the following letter relative to the present one. Pagliarini et al. (2015) demonstrated that these two principles are compromised in children with dyslexia (but no comorbid diagnosis of dysgraphia). In their study, Pagliarini et al. (2015) evaluated the handwriting abilities of Italian-speaking, 9-year-old children with dyslexia and a group of TD children. Children were asked to write the Italian word burle (‘jokes’) using a wireless pen and a digital tablet with their dominant hand. They were instructed to write the word in both block and cursive script across different conditions: spontaneous, big and fast.
Figure 6 presents the handwriting speed of the spontaneous and big conditions for both block (panel A) and cursive (panel B) scripts. For the TD group, the letter duration curves closely align across the two conditions. This means that, in line with the homothety principle, TD children successfully maintained the relative duration of individual letters constant across conditions, regardless of whether they were writing in block or cursive script. They also adhered to the isochrony principle, as the total duration (given by the sum of the duration of single letters) remained consistent across conditions. Indeed, these children wrote faster when they were asked to write larger letters. In contrast, children with dyslexia were less able than TD children to comply with the homothety principle. Findings showed that children with dyslexia could not maintain the relative duration of individual letters constant across conditions, as illustrated in Figure 6. This lack of alignment in the curves is evident in both block and cursive script, suggesting that their difficulties in adjusting movement timing persist across different handwriting styles. Children with dyslexia were less successful than TD peers in conforming to the isochrony principle, as their total duration varied across conditions, rather than remaining stable. It is important to note that, besides the handwriting tasks described above, Pagliarini et al. (2015) assessed the children using a scribbling task – a motor task in which lexical access or orthographic coding is not required. There were no significant differences between children with and without dyslexia on the scribbling task, indicating that the poorer performance of children with dyslexia in complying with handwriting rhythmic principles does not stem from a motor deficit.
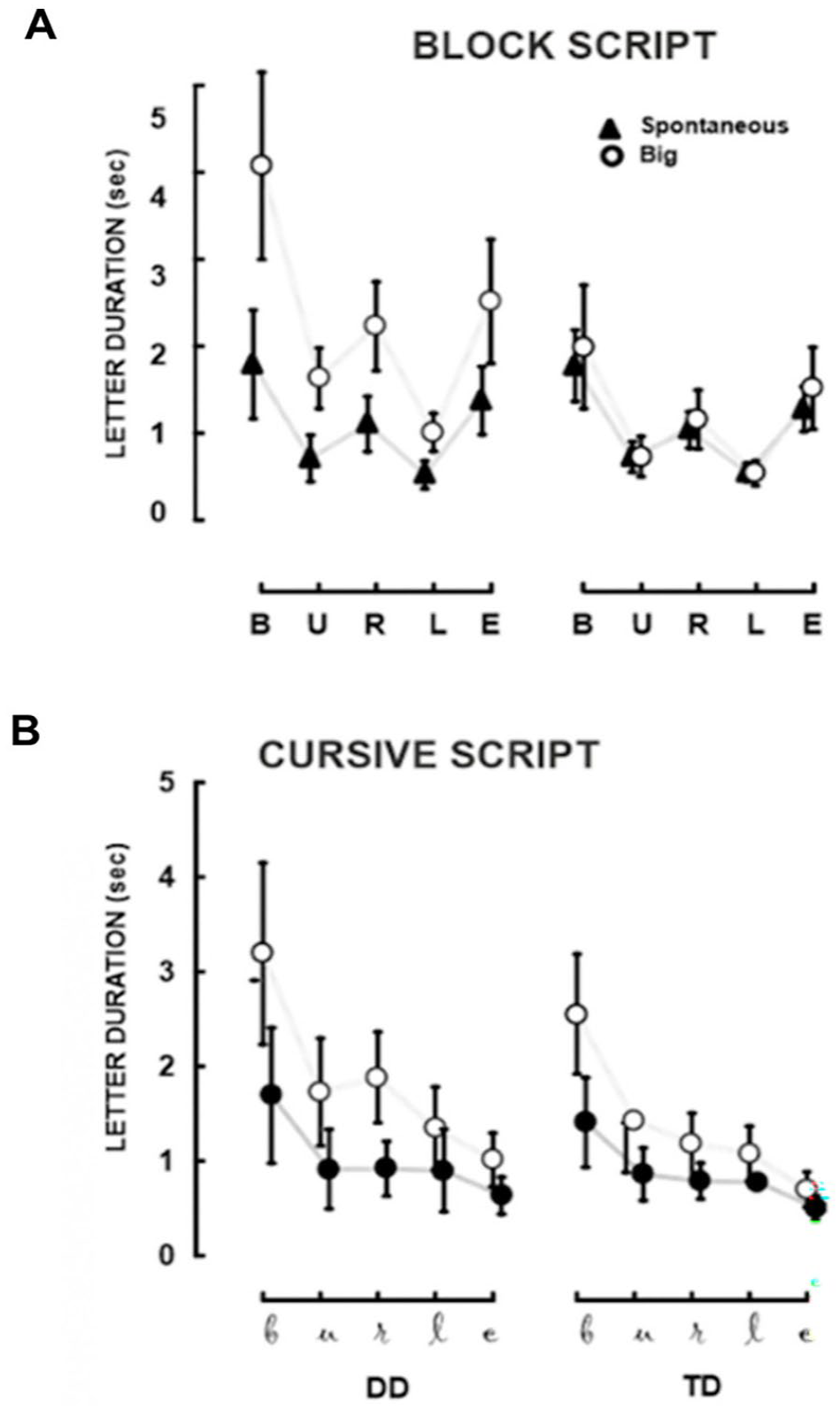
Duration of separate letters in block script (panel A) and cursive script (panel B) for children with developmental dyslexia (DD) and typically developing (TD) peers. The second-order interaction of Group (DD, TD) by Condition (Spontaneous, Big) by Letter ((b), (u), (r), (l), (e)) is shown for duration (vertical error bars represent standard error). Writing the first letter (b) requires more time than the others, indicating the burden of initiating the writing task. The curves representing the performances of TD children closely align, demonstrating adherence to the isochrony principle (keeping movement duration constant across changes in size) and the homothety principle (maintaining relative durations of movement components across changes in speed). In contrast, children with DD show greater difficulty adjusting their handwriting movement to experimental size manipulations. Additionally, for children with DD, the relative durations of letters deviate from the homothety principle. Adapted from “Dyslexic Children Fail to Comply With the Rhythmic Constraints of Handwriting,” by E. Pagliarini, M. T. Guasti, C. Toneatto, E. Granocchio, F. Riva, D. Sarti, B. Molteni, and N. Stucchi, 2015, Human Movement Science, 42, 161–182 (https://doi.org/10.1016/j.humov.2015.04.012). Copyright 2015 by Elsevier. Adapted with permission.
To ascertain that the difficulty with rhythmic production of handwriting in children with dyslexia was not due to a lack of exposure to handwriting training, Pagliarini and colleagues conducted a follow-up study. Pagliarini et al. (2017) examined the development of isochrony and homothety principles in 6- to 10-year-old TD children. Outcomes revealed that, from 6 years of age (first grade) and following only a brief exposure to formal education, TD children appropriately adhere to the rhythmic principles of handwriting. These findings suggest that TD children’s ability to comply with homothety and isochrony is not influenced by formal training and does not result from formal training. Instead, these principles are inherent in the rhythmic structures that TD children are equipped with at the earliest stage of learning to write (Pagliarini et al., 2017). Therefore, it could be argued that the poor rhythmicity in handwriting exhibited by children with dyslexia is not due to a lack of exposure to writing training. Instead, these difficulties could be attributed to a deficit in generating sequences of rhythmic events that are time-locked to each other (Pagliarini et al., 2015).
The fact that children with dyslexia exhibit a deficit in handwriting supports the Inefficient Anticipatory Planning Hypothesis. On the one hand, synchronisation is not involved, and yet we observed deficits; on the other, the lack of compliance with the principle of homothety, which requires rescaling the duration of single letters, indicates that they are not well prepared to start to or stop to act at the right moment, that is, they cannot anticipate when they have to act. As in the case of the warning-imperative task, children with dyslexia are trying hard to cope with anticipation because there is some modulation in the relative duration of single letters according to the handwriting condition. However, their compliance with homothety is very poor because, across the conditions, there is no difference in total duration (which is slightly present in TD children in the cursive script), but the relative durations of letters change at variance with the homothety principle.
So far, we have shown that individuals with dyslexia are poor at anticipating, and this impairment affects language/speech, reading, and handwriting. If the same mechanism affects all these behaviours, we expect to find correlations between the various skills or specific measures involved in these skills that tap into anticipation. The following section will discuss this aspect.
Anticipatory planning as a link between language, reading and handwriting
Thus far, we have shown that anticipatory planning is evident in spoken language, reading and handwriting. Although limited, we discussed evidence showing that anticipatory planning is impaired in individuals with dyslexia. We conjecture that inefficient anticipatory planning is a mechanism that can go awry in dyslexia. As such, anticipation deficits should be more central to studying dyslexia. If anticipation is a common thread that unites this language, reading and handwriting impairments, we expect correlations between these activities. In line with this view, Pagliarini et al. (2020) demonstrated not only that anticipatory planning was indeed impaired in children and adults with dyslexia but also showed that the timing error and the individual consistency or precision (e.g., the standard deviation of timing error of the tapping responses) correlated negatively with reading speed. The less accurate and less consistent a person is in anticipating the warning beat, the fewer syllables per second they read. Those who were less efficient in anticipation were also slower readers. Pagliarini et al. (2020) also identified a positive correlation between the children’s timing error in the warning-imperative task, individual consistency/precision, and the number of errors in reading nonwords. Children with higher timing errors and inconsistency in pressing the button at the imperative beat also made more reading errors. Thus, reading and various measures related to anticipatory planning are correlated. In addition, Pagliarini et al. (2015) found that writing speed positively correlated with the child’s speed in reading words and nonwords and receptive vocabulary and negatively correlated with the number of reading errors. Therefore, the children who wrote faster also read faster, had better vocabulary and made fewer errors in reading words and nonwords (Pagliarini et al., 2015). Conversely, children who wrote less fluently also read more slowly, made more reading errors and had poorer receptive vocabulary. Interestingly, Pagliarini et al. (2015) investigated the hypothesis that reading and writing are mediated by rhythmic competence by correlating reading speed and an index of children’s ability to comply with isochrony (in handwriting). This index was the difference in writing speed in the Big and the Spontaneous conditions. It was found that the index of the isochrony compliance positively correlated with the speed of reading words and nonwords. This suggests that the association between reading and writing could be mediated by rhythmic competence. Accordingly, Pagliarini et al. (2015) proposed that poor reading and handwriting may share an underlying mechanism: anticipatory planning facilitated by rhythm. Finally, concerning the relation between reading and language processing, we discussed research in both children and adults with dyslexia that has demonstrated difficulties in using linguistic cues to predict upcoming input. Huettig and Brouwer (2015) showed that adults with dyslexia exhibit delayed anticipatory eye movements when processing grammatical cues, and these delays correlate with their reading performance. This link between delayed anticipatory eye movements (a motor behaviour guided by spoken language in the case in question) and reading may suggest a shared mechanism. Based on various facts, we have suggested that this mechanism may be rooted in anticipatory planning. In sum, anticipatory planning seems to be a common thread linking language, reading and motor activities, such as handwriting. Accordingly, we have suggested that a deficit in anticipation may affect various behaviours that unfold in time and call for the same mechanism.
Reflections on the Inefficient Anticipatory Planning Hypothesis
Our hypothesis has several strengths. First, it offers a unifying account of the difficulties seen in dyslexia across language, reading and handwriting skills. Second, several theories (Fiveash et al., 2021; Goswami, 2011; Ladányi et al., 2020) have highlighted the temporal processing deficits associated with dyslexia, focusing on the role of neural oscillations and the role of prediction in the processing of temporal organisation in music, speech and language. Our hypothesis advances the field by specifying the nature of these rhythmic deficits. We distinguish between two cognitive mechanisms involved in processing temporal organisation: anticipatory planning – a special form of prediction facilitated by rhythm – and synchronisation (elicited by metre). This distinction has not been clearly articulated in previous research. It may have important implications for neuroimaging studies on neural oscillations and for designing experiments that differentiate between these mechanisms. As for any new hypothesis, further research is needed to explore the relationship between these two processes.
Our hypothesis contributes to the many alternative explanations proposed in the literature to account for dyslexia, including phonological deficits, automatisation failure, limitations in processing speed, temporal processing deficits, working memory deficits and attention deficits (e.g., Ramus & Szenkovits, 2008; Ramus et al., 2013; Snowling, 1998; Nicolson & Fawcett, 1990, 2011; Denckla & Rudel, 1976; Zoccolotti et al., 2013; Pennington et al., 2012; Ladányi et al., 2020; Goswami, 2011; Beneventi et al., 2010; Menghini et al., 2011; Pennington, 2006; Ring & Black, 2018; McGrath, Peterson, & Pennington, 2020). This highlights the need for further research on the interaction between anticipatory planning and other cognitive processes affected by dyslexia. We cannot claim that inefficient anticipatory planning explains all aspects of dyslexia, but we believe it is a crucial area for further investigation. For instance, it will be interesting to examine how anticipatory planning develops over time in children with dyslexia and whether early differences in anticipatory planning can predict the likelihood of dyslexia later (see Dui et al., 2020). We have provided evidence for our hypothesis based on results with dyslexic participants in the warning-imperative task and handwriting tasks, where compliance with the rhythmic organisation or lack thereof was responsible for the observed behaviour. However, in the case of language and reading, we only discussed evidence for an anticipatory behaviour, which may or may not be facilitated by rhythm. It will be important to test, more directly, the link between anticipatory planning performance on language prediction tasks, reading and handwriting and examine how these relations (if they truly exist) develop over time.
Although our hypothesis focuses on dyslexia, the role of anticipatory planning may extend to other neurodevelopmental conditions, such as developmental language disorders, where similar difficulties in reading, language and handwriting skills have been reported. Lastly, much of the evidence presented in this article has been drawn from behavioural studies, pointing to the need to explore the neural mechanisms underlying anticipatory planning in dyslexia. Future research using larger sample sizes, longitudinal designs and randomised control trials is necessary to understand better the role of anticipatory planning in developmental disorders such as dyslexia.
Conclusion
We focused on anticipatory planning, the ability to process information ahead and being prepared to act. Anticipation is fundamental in phenomena that unfold in time, particularly those that require precise timing and sequential ordering of behavioural patterns (Lashley, 1951). Examples of such phenomena include language processing, reading and motor activities such as handwriting. In this article, we looked at anticipation through the lens of dyslexia. Here, we have shown that language, reading and handwriting are all activities that involve some level of temporal organisation, which may in turn facilitate anticipatory planning. We developed the Inefficient Anticipatory Planning Hypothesis, proposing that individuals with dyslexia have an inefficient mechanism of anticipatory planning, which may partially contribute to their difficulties in tasks that tap into language processing, reading and handwriting. In line with this hypothesis, we have shown that individuals with dyslexia have difficulties in exploiting morpho-syntactic information to predict upcoming words during language processing. In the case of reading, we have discussed evidence showing that individuals with dyslexia do not benefit much from parafoveal vision, that is, they cannot process information ahead in reading words or in naming symbols (in the RAN test), which requires few decoding skills. In handwriting, we showed that children with dyslexia cannot comply with the rhythmic principles of handwriting, that is, they cannot rescale the relative durations of single letters when writing or change writing speed in relation to changes in letter/word size. We are aware that there may be alternative explanations for the data we presented in the paper, but we believe that anticipatory planning can equally contribute to our understanding of dyslexia and (developmental) language disorders.
Footnotes
Author Contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project received funding from the European Union’s Horizon 2020 program for research and innovation under the Marie Skłodowska Curie Grant Agreement Number 765556 (Principal Investigator: MTG). EP acknowledges the financial support of Grant PID2022-138413NB-I00, funded by MCIN/AEI/ 10.13039/501100011033 and by the European Union; the support of Grant 2021SGR00787, funded by the Generalitat de Catalunya; the support of the ‘Ramón y Cajal’ Grant RYC2021-033969-I, funded by MCIN/AEI/ 10.13039/501100011033 and by European Union Next Generation EU/PRTR.