
Abstract
Objective
This study aimed to isolate and characterize endophytic fungi associated with different tissues of Alocasia macrorrhizos and to evaluate their antibacterial and antioxidant potentials, supported by chemical profiling and in silico validation of bioactive metabolites.
Methods
Endophytic fungi were isolated from the leaves, corm, and roots of A. macrorrhizos and identified through morphological and molecular analyses. Ethyl acetate extracts of all isolates were screened for antibacterial activity using the disc diffusion method and for antioxidant potential using 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging and ferric reducing antioxidant power (FRAP) assays. Bioactive extracts were analyzed by gas chromatography–mass spectrometry (GC–MS) to identify secondary metabolites. Molecular docking studies were performed against the bacterial target protein lactotransferrin to predict binding affinities and interaction patterns of selected compounds.
Results
Six endophytic fungi were isolated and identified as Curvularia sp. (AML-F1), Rhizopus sp. (AML-F2), Aspergillus spp. (AMC-F3 and AMC-F6), Paecilomyces sp. (AMR-F4), and Tulasnella sp. (AMC-F5). Molecular analysis confirmed Paecilomyces lecythidis (AMR-F4) and Aspergillus fumigatus (AMC-F6). Among all isolates, AMR-F4 and AMC-F6 showed the strongest antibacterial activity, with inhibition zones of 21.0 ± 1.52 mm and 22.0 ± 0.60 mm against Bacillus subtilis, and 17.7 ± 1.50 mm and 17.7 ± 0.31 mm against Staphylococcus aureus at 500 µg/disc, comparable to kanamycin. Both extracts also demonstrated notable antioxidant activity, with DPPH IC50 values of 74.13 ± 1.49 µg/mL and 100.56 ± 0.16 µg/mL, and strong ferric ion reduction in the FRAP assay. GC–MS analysis identified 47 metabolites from AMR-F4 and 39 from AMC-F6. Molecular docking revealed higher binding affinities of selected compounds than kanamycin.
Conclusions
Alocasia macrorrhizos hosts bioactive endophytic fungi, particularly P. lecythidis and A. fumigatus, highlighting their potential as natural sources of antimicrobial and antioxidant agents.

Keywords
1. Introduction
Nature has long served as the primary source of healing, providing medicinal compounds derived from leaves, roots, and seeds long before the development of modern medicine. 1 Apart from plants, endophytic microbes have been recognized as valuable sources of bioactive compounds since the discovery of Taxol and have emerged as important contributors to modern drug discovery. These microorganisms produce a wide range of metabolites with anticancer, antimicrobial, antioxidant, antiviral, antidiabetic, anti-Alzheimer’s, and immunosuppressive activity, thereby enriching pharmaceutical pipelines. 2 However, only a limited number of plant species have been investigated for their endophytic richness and bioactivity. 3 Endophytic fungi are essential components of the plant microecosystem and affect host physiological processes via multiple mechanisms, including the production of phytohormones such as indoleacetic acid and the release of metabolites that support stress tolerance. These fungi generate secondary metabolites such as terpenoids, steroids, xanthones, and phenols, which exhibit strong antibacterial, antifungal, and herbicidal activities. 4 Compared to soil-derived fungi, endophytes display higher bioactivity and industrial relevance, contributing to enzyme, organic acid, riboflavin, and alcohol production, thus underscoring their therapeutic and biotechnological potential.2,5–7 Secondary metabolites from endophytic fungi represent a major source of novel therapeutic agents. For example, the anticancer drug Taxol was derived from the Pacific yew (Taxus brevifolia) in the 1990s. Similarly, the anticancer drug vincristine was developed from the endophytic fungi Mycelia sterilia. 8 These discoveries highlight the critical importance of endophytic fungi as reservoirs of novel compounds that drive drug discovery, therapeutic breakthroughs, and pharmaceutical innovation. 9 Therefore, there is an urgent need to explore additional plant hosts as sources of endophytic fungi to further expand this vital area of research.
Alocasia macrorrhizos is a medicinally important plant in the family Araceae, traditionally used to treat inflammation, infections, pain, and metabolic disorders. Phytochemical investigations have shown that this species is rich in phenolics, flavonoids, alkaloids, and terpenoids—compounds frequently associated with antimicrobial and antioxidant activities. Plants that produce such chemically diverse and bioactive metabolites provide favorable ecological niches for endophytic fungi, as prolonged host–endophyte associations can drive co-evolution and stimulate the biosynthesis of structurally diverse secondary metabolites with complementary biological functions. Consequently, medicinal plants that display robust chemical defense systems are increasingly regarded as promising reservoirs of endophytes with pharmaceutical potential.10–12
Despite its well-documented ethnomedicinal use and bioactive phytochemical profile, A. macrorrhizos has not been systematically investigated as a host for endophytic fungi. The scarcity of reports on endophytes from this plant suggests that its associated fungal communities and their metabolic potential remain largely unexplored. Therefore, the present study aims to isolate and characterize endophytic fungi from different tissues of A. macrorrhizos and to evaluate their antibacterial and antioxidant activities. By integrating biological assays, chemical profiling, and molecular docking analyses, this work seeks to uncover bioactive fungal metabolites and to establish A. macrorrhizos as a promising yet underexplored source of endophytic fungi with potential relevance to natural product–based drug discovery.
2. Materials and methods
2.1. Collection and identification of the selected plant
A healthy, mature Alocasia macrorrhizos plant was collected in June 2024 from Dapunia Union, Pabna Sadar Upazila, Pabna District, Rajshahi Division, Bangladesh (approximately 24.04° N, 89.16° E). The specimen was obtained from a shady site after rainfall. Botanical authentication was performed by an expert at the Bangladesh National Herbarium, Dhaka, and a voucher specimen (No. DACB 135481) was deposited for future reference. Collected samples were transported to the laboratory in sterile bags, processed immediately upon arrival, and separated into corm, root, and leaves for further study.
2.2. Isolation and cultivation of endophytic fungi
Endophytic fungi were isolated following surface sterilization of plant tissues to eliminate external contaminants. Fresh plant tissues were rinsed three times with sterile distilled water, each rinse lasting 10 min. Next, they were dried using sterile paper towels and cut into 1-cm segments. Roots and corms were subjected to sequential sterilization with 75% (v/v) ethanol for 1 min, 5.3% sodium hypochlorite for 5 min, and a final rinse in 75% ethanol for 30 s. Leaves were sterilized with 75% ethanol for 2 min. Following sterilization, all tissues were air-dried on sterile filter paper inside a laminar airflow cabinet. To prevent bacterial growth, each explant was placed onto potato dextrose agar (PDA) supplemented with 0.05 g/L of chloramphenicol. The final rinse water was plated onto PDA as a negative control to verify the effectiveness of sterilization. Plates were sealed and incubated at 26–28 °C for 7 days and observed daily for fungal growth. Emerging fungal colonies were subcultured onto fresh PDA plates using flame-sterilized inoculating loops and incubated for an additional 7 days to obtain pure endophytic fungal isolates. 12
2.3. Identification of the endophytic fungi
2.3.1. Morphological identification
The isolated endophytic fungal strains were identified based on their macroscopic and microscopic characteristics.12,13 Morphological identification was based on color, texture, morphology, and growth patterns from both the front and reverse sides of the Petri dishes. Microscopic characteristics were determined via examination of features such as spore shape, size, color, hyphal structure, septation, and the presence of fruiting bodies. Lactophenol Cotton Blue staining was used for microscopic analysis, in which phenol acts as a disinfectant, lactic acid preserves fungal structures, and cotton blue selectively stains chitin in the fungal cell wall. Stained slides were prepared and examined under 40× magnification using a light microscope to visualize fungal structures. 11
2.3.2. Molecular identification and phylogenetic analysis
Genomic DNA from the selected endophytic fungi was extracted using the BioFACT Genomic DNA Prep Kit (BIOFACT Co. Ltd., Korea) according to the manufacturer’s instructions. High-quality DNA obtained via the salting-out method was used to amplify the internal transcribed spacer (ITS) region by PCR using the HotStar Taq Master Mix Kit (Qiagen, USA). ITS5 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATGATGC-3′) primers (Invitrogen, USA) were employed under the following thermal cycling conditions: initial denaturation at 95 °C for 2 min, 35 cycles at 95 °C for 1 min, 56 °C for 40 s, and 72 °C for 1 min, and a final extension at 72 °C for 10 min. DNA concentration and purity were measured using a NanoDrop spectrophotometer, and amplification was confirmed via 2% agarose gel electrophoresis. Purified PCR products were subjected to Sanger sequencing, and the resulting ITS sequences were analyzed using BLAST against the NCBI GenBank database for species identification. Phylogenetic trees were constructed using the Neighbor-Joining method in MEGA 11, and all ITS sequences were deposited in GenBank.14,15
2.4. Extraction of secondary metabolites
Isolated endophytic fungi were cultivated using liquid fermentation for the extraction of secondary metabolites. Fungal cultures were inoculated into 2,000-mL Erlenmeyer flasks containing 1,000 mL of potato dextrose broth and incubated at 28 °C for 21–28 days on a rotary shaker maintained at 150 rpm. 16 Following incubation, the fermented broth was filtered through three layers of muslin cloth to remove the fungal mycelia. The resulting filtrate was extracted three times with ethyl acetate using a separating funnel. The organic solvent phase was then concentrated under reduced pressure using a rotary evaporator. The concentrated extract was further dried in a water bath to obtain the crude secondary metabolites. The dried crude extract (approximately 400 mg) was dissolved in dimethyl sulfoxide (DMSO) to prepare a 20 mg/mL stock solution. The solution was filtered through a 0.22-μm syringe filter and stored at 4 °C for further use in biological assays. For all bioassays, the stock solution was diluted in the appropriate assay medium such that the final DMSO concentration did not exceed 1% (v/v), a level shown not to interfere with biological activity. 17
2.5. Evaluation of antimicrobial activity via disc diffusion
The antimicrobial activity of the isolated endophytic fungi was determined following the methods described by De Siqueira, with some modifications. 18 Agar media, Petri dishes, and glassware were sterilized by autoclaving at 121 °C at 15 lb/sq inch for 15 min. Inoculating loops and forceps were exposed to UV light for 30 min in a laminar airflow cabinet prior to use. Fungal extracts (5 mg) were dissolved in ethanol and vortexed to prepare 25 µg/µL and 50 µg/µL solutions. Sterile filter paper discs (5 mm diameter) were impregnated with 10 µL of the corresponding test solutions to achieve final concentrations of 250 µg/disc and 500 µg/disc. Kanamycin (30 µg/disc) was used as a positive control, and discs treated with ethanol served as a negative control. All plates were incubated in an inverted position at 37 °C for 16–18 h, after which antibacterial activity was evaluated.
2.6. Evaluation of antioxidant activity of crude metabolites via DPPH free radical scavenging
Antioxidant activity of the fungal crude extracts was evaluated using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging assay following the method described by Yadav and colleagues with minor modifications.
19
Extracts were dissolved in methanol and serially diluted to concentrations of 3.125–100 µg/mL, while ascorbic acid served as the standard. A freshly prepared solution of 0.004% DPPH in methanol was stored at 4 °C in the dark until use. For the assay, each sample (2 mL) was mixed with 3 mL of DPPH solution and incubated at room temperature in the dark for 30 min. Absorbance was measured at 517 nm using a UV-Vis spectrophotometer, with methanol serving as the blank. The percentage of DPPH radical scavenging activity was calculated using the following formula:

Here, IC50 is the concentration of a sample required to scavenge 50% of DPPH free radicals.
2.7. Determination of antioxidant activity by ferric reducing antioxidant power assay
The FRAP assay was carried out according to the method described by Benzie and co-workers by monitoring the reduction of Fe3+- tripyridyl triazine (TPTZ) to the blue-colored Fe2+-TPTZ. 20 In the reducing power assay, fungal extracts and ascorbic acid (standard) at varying concentrations (1.5625-100 µg/mL) were mixed with phosphate buffer and potassium ferricyanide and incubated at 50 °C for 20 min. After adding TCA, FeCl3, and water, the absorbance was read at 700 nm. Increased absorbance indicated greater reducing activity, which was calculated as the percentage increase in reducing power.
2.8. Identification of bioactive components via GC–MS analysis
Two fungal extracts, AMR-F4 and AMC-F6, were subjected to gas chromatography–mass spectrometry (GC–MS) analysis following the evaluation of their biological properties. 21 Analyses were conducted at the CSIRL Laboratory, Jashore University of Science and Technology Bangladesh, using a PerkinElmer Clarus® 690 gas chromatograph equipped with an Elite-35 column (30 m × 0.25 mm × 0.25 µm) coupled to a Clarus® SQ 8C mass spectrometer. Samples (1 µL) were injected in splitless mode, with helium (99.99%) as the carrier gas at a constant flow rate. Electron ionization was carried out at 70 eV. The oven temperature was programmed from 60 °C to 240 °C at 5 °C/min, with the final temperature held at 240 °C for 4 min to ensure complete elution of analytes. The injector temperature was maintained at 280 °C. Components were identified based on molecular weight, structure, and fragmentation patterns using standard spectral databases. This analysis provided information about individual metabolites, supporting their potential biological or therapeutic relevance and guiding subsequent extraction strategies.
2.9. Molecular docking
In silico molecular docking was performed to predict interactions between bioactive compounds identified from GC–MS analysis and selected protein receptors. The target protein, 1B0L (Lactotransferrin), is a human iron-binding glycoprotein with well-established antimicrobial properties. The recombinant structure, expressed in Aspergillus awamori and resolved through X-ray crystallography at 2.2 Å resolution, comprises 691 amino acids with a theoretical molecular weight of 76.29 kDa. This structure is highly conserved and functionally comparable to native human lactoferrin, containing two Fe3+ and two CO32- ions essential for its biological activity. A chemical library was constructed using compound information retrieved from the PubChem database, including molecular weight, formula, and 3D structure. ADMET profiling was performed using pkCSM and SwissADME to evaluate pharmacokinetic properties and drug-likeness based on Lipinski’s rule of five.21,22 Ligand structures were optimized in BIOVIA Discovery Studio by adjusting geometry, adding hydrogen atoms, assigning partial charges, and minimizing energy using the CHARMm force field. For flexible compounds, multiple conformers were generated using the conformer generation protocol. Protein structures were obtained from the RCSB Protein Data Bank, refined by removing bound ligands and water molecules, and optimized by adding hydrogen atoms and assigning charges using BIOVIA Discovery Studio. 23 A grid box was generated around the receptor’s active site to define the docking region, and ligands were positioned within this grid to explore all feasible binding orientations. Molecular docking was performed using PyRx (AutoDock Vina) with parameters optimized for the receptor binding pocket. Binding affinities were evaluated based on docking scores and interaction energies, including hydrogen bonding, van der Waals forces, and electrostatic interactions. The top-ranked poses were further refined through energy minimization to improve binding accuracy and stability. Protein–ligand interactions were visualized and analyzed using PyMOL and BIOVIA Discovery Studio Visualizer to identify key binding modes, hydrogen bonds, and hydrophobic contacts. Docking accuracy was validated against available experimental data, where applicable. 24
2.10. Statistical analyses
All experiments were performed in triplicate, and results are presented as the mean ± standard deviation (SD) of three independent measurements to reflect within-group variability. Data organization and preliminary descriptive statistics were conducted using Microsoft Excel (version 2016). All inferential statistical analyses were performed using IBM SPSS Statistics (version 26). Prior to statistical testing, data distributions were assessed for normality using the Shapiro–Wilk test. Homogeneity of variance between groups was evaluated using Levene’s test. When assumptions of normality and equal variance were satisfied, parametric tests were applied. Comparisons among multiple groups were conducted using one-way analysis of variance (ANOVA), followed by appropriate post hoc tests with adjustment for multiple comparisons where applicable. All statistical tests were two-tailed, and a p-value < 0.05 was considered statistically significant. Graphical representations were generated using the same software to ensure consistency between statistical analysis and data visualization. No artificial intelligence tools were used in conducting the experimental work or in the statistical analysis.
3. Results
3.1. Isolated endophytic fungi from Alocasia macrorrhizos
Six endophytic fungi were isolated from different parts of Alocasia macrorrhizos, including the corm, leaves, and roots. Among these, two isolates (AML-F1 and AML-F2) were obtained from the leaves, one (AMR-F4) from the roots, and three (AMC-F3, AMC-F5, and AMC-F6) from the corm. The preliminary identification of these fungal isolates was based on their colony color and growth characteristics on PDA medium.
3.2. Identification of isolated endophytic fungi
3.2.1. Morphological identification
Morphological features of the isolated fungal strains.


Macroscopic view (a) front side view, (b) back side view and (c) microscopic view (40X) of AML-F1.
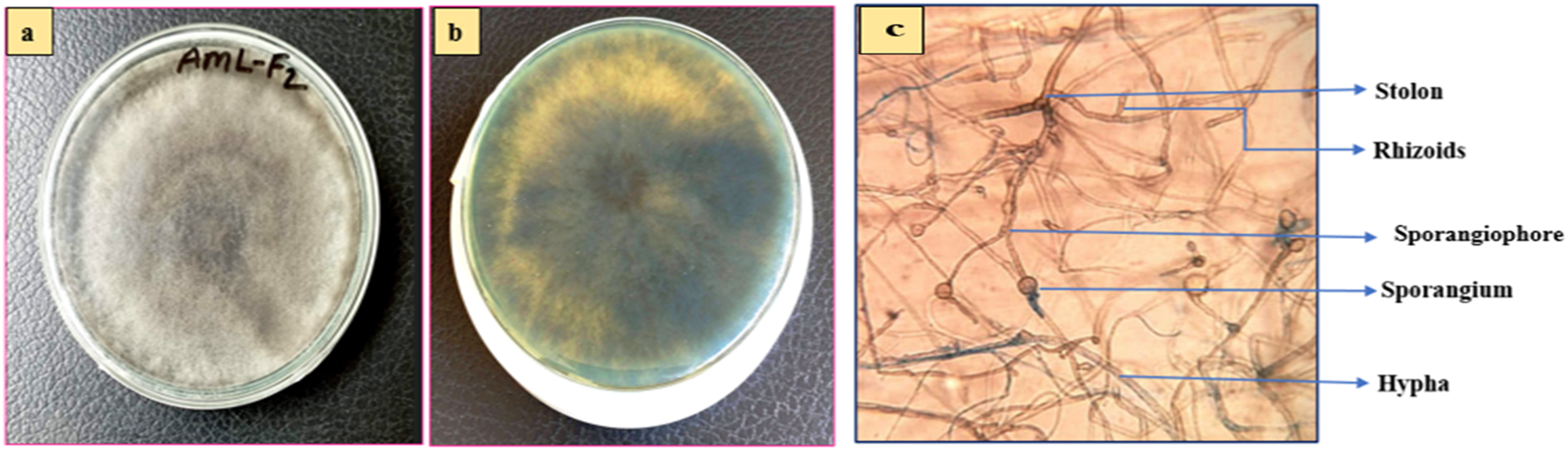
Macroscopic view (a) front side view, (b) back side view and (c) microscopic view (40X) of AML-F2.

Macroscopic view (a) front side view, (b) back side view and (c) microscopic view (40X) of AMC-F3.

Macroscopic view (a) front side view, (b) back side view and (c) microscopic view (40X) of AMR-F4.

Macroscopic view (a) front side view, (b) back side view and (c) microscopic view (40X) of AMC-F5.
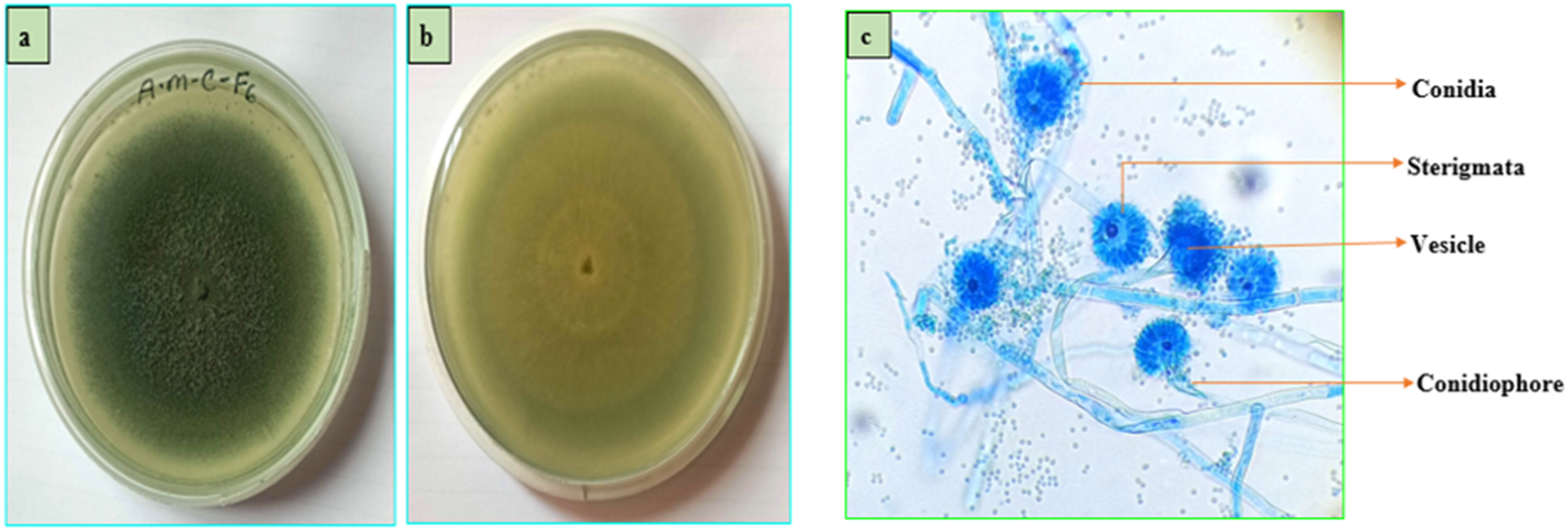
Macroscopic view (a) front side view, (b) back side view and (c) microscopic view (40X) of AMC-F6.
3.2.2. Molecular identification
The ITS sequence of isolate AMR-F4 exhibited 99.05% similarity with Paecilomyces lecythidis (GenBank Accession No. PX459735), confirming its identity as Paecilomyces lecythidis. Similarly, the ITS sequence of isolate AMC-F6 showed 99.03% similarity to Aspergillus fumigatus (GenBank Accession No. PX459737), confirming its identity as Aspergillus fumigatus. Phylogenetic tree analysis further validated these findings, as both isolates clustered closely with their respective reference species, supported by high bootstrap values (Figures 7 and 8). These clustering patterns strongly suggest that AMR-F4 and AMC-F6 are genetically affiliated with the Paecilomyces and Aspergillus lineages, respectively. Phylogenetic relationship between AMR-F4 and the other related species. Using the maximum likelihood approach (1000 bootstrap replication), the phylogenetic tree was built. Phylogenetic relationship between AMC-F6 and the other related species. Using the maximum likelihood approach (1000 bootstrap replication), the phylogenetic tree was built.

3.3. Evaluation of antimicrobial activity
In-vitro antibacterial activity of crude metabolites of AMR-F4.

*Results are means of three replicates (n = 3) ± SEM.
In-vitro antibacterial activity of crude metabolites of AMC-F6.

*Results are means of three replicates (n = 3) ± SEM.
The antibacterial activity of all active extracts was concentration-dependent, with significantly larger inhibition zones observed at 500 µg/disc than at 250 µg/disc. One-way ANOVA revealed significant differences in inhibition zones among the tested bacterial strains (P < 0.0001). Dunnett’s post-hoc test further confirmed that several fungal extracts, particularly AMR-F4 and AMC-F6, produced inhibition zones that were significantly different from those of the standard antibiotic kanamycin (P < 0.001). These findings demonstrate statistically significant variation in antibacterial efficacy among the fungal metabolites evaluated.
3.4. Evaluation of antioxidant activity
3.4.1. DPPH free radical scavenging activity
All fungal extracts exhibited DPPH radical scavenging activity in a concentration-dependent manner, although the extent of activity varied among the isolates (Figure 9). Among the tested samples, AMC-F3 demonstrated the highest antioxidant potential, with an IC50 value of 18.45 ± 0.19 μg/mL, which was the closest to that of ascorbic acid (4.44 ± 0.04 μg/mL). The AML-F1 extract also showed moderate scavenging activity (IC50 = 32.34 ± 0.91 μg/mL), followed by AMC-F5 (IC50 = 57.50 ± 0.15 μg/mL) and AMR-F4 (IC50 = 74.13 ± 1.49 μg/mL). By contrast, AML-F2 and AMC-F6 displayed relatively weaker activity, with IC50 values of 97.21 ± 1.20 μg/mL and 100.56 ± 0.16 μg/mL, respectively. At higher concentrations (100 μg/mL), AMC-F3 achieved a mean inhibition of 78.06 ± 0.26%, while that of AML-F1 and AMC-F5 was 62.04 ± 0.60% and 59.19 ± 0.14%, respectively. Ascorbic acid, the positive control, showed the highest inhibition (94.86 ± 0.20%), confirming the validity of the assay. These findings indicate that several fungal metabolites possess notable free radical-scavenging ability, likely due to the presence of phenolic or flavonoid compounds capable of donating hydrogen atoms or electrons to neutralize DPPH radicals (Supplementary Tables S5–S14, Figure 9, and Supplementary Figure S7). DPPH free radical scavenging activity of fungal crude extracts and ascorbic acid. (a) Dot plot showing percentage inhibition of DPPH radicals at different concentrations (3.125–100 μg/mL) for each fungal extract and the positive control (ascorbic acid). Each dot represents an individual experimental data point obtained from independent technical replicates (n = 3 per concentration). Horizontal lines indicate the mean value for each group, and error bars represent the standard error of the mean (SEM). (b) Box-and-whisker plot summarizing IC50 values of the fungal extracts and ascorbic acid. Boxes represent the interquartile range (IQR), the central line indicates the median, whiskers denote the minimum and maximum values, and individual points represent independent measurements. IC50 values were calculated from concentration–response curves derived from three independent experiments (n = 3).
3.4.2. Ferric reducing power assay
The reducing power of the fungal extracts increased with increasing concentration, reflecting their ability to donate electrons and reduce Fe3+ to Fe2+. Among the tested samples, AMR-F4 exhibited a stronger ferric reducing capacity (93.24 ± 0.40% at 100 μg/mL) than AMC-F6 (91.47 ± 0.40% at 100 μg/mL), although both values were lower than that of ascorbic acid (96.65 ± 0.20%). The progressive, concentration-dependent increase in reducing power suggests that these fungal extracts contain redox-active secondary metabolites capable of acting as electron donors, thus contributing to their overall antioxidant potential.
3.5. Chemical profiling of fungal extracts AMR-F4 and AMC-F6 through GC–MS
Major bioactive compounds identified in the AMR-F4 extract through GC-MS.

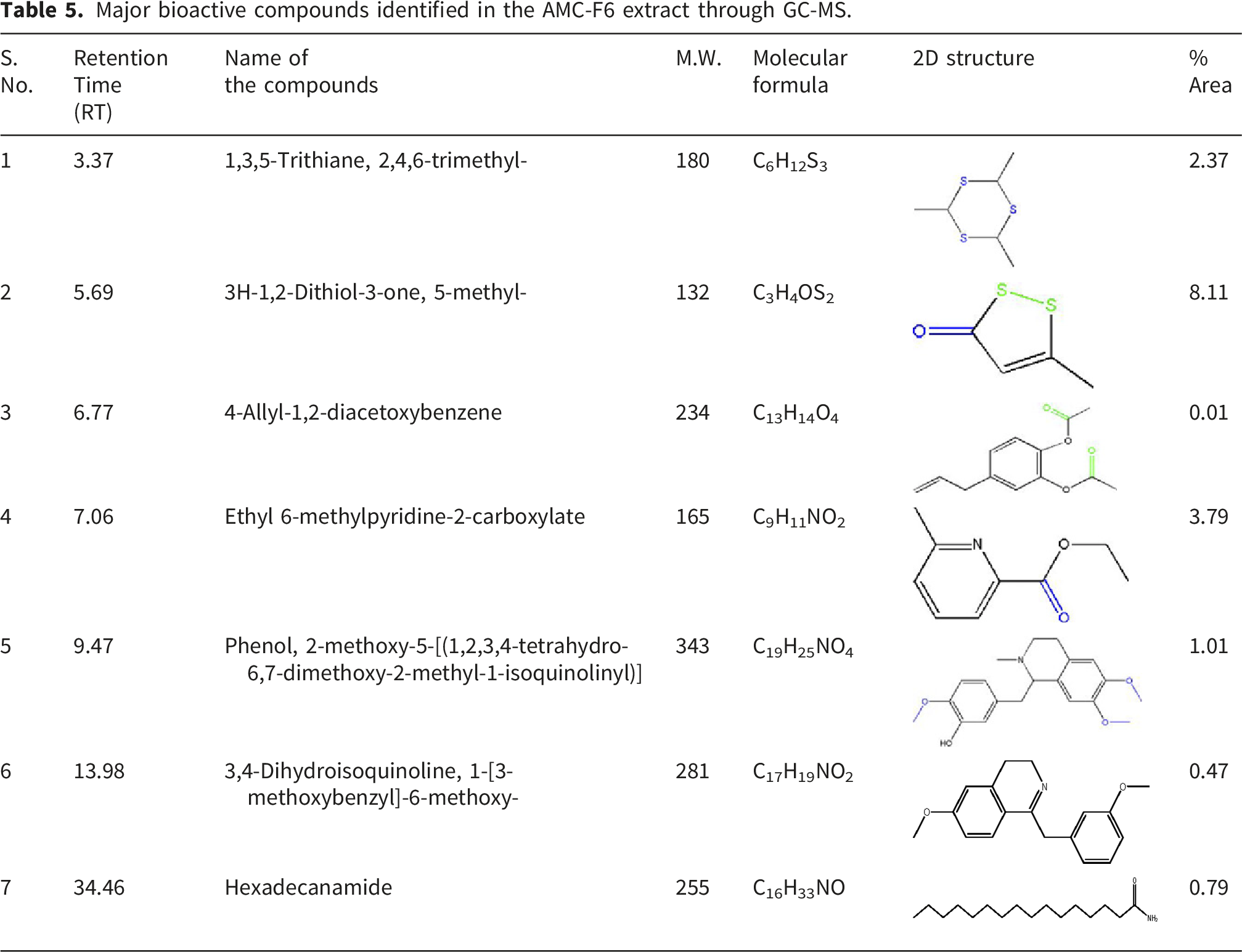
Major bioactive compounds identified in the AMC-F6 extract through GC-MS.
3.6. Molecular docking study to evaluate antibacterial activities of GC–MS identified major compounds
Binding affinity of major ligands with targeted 1B0L protein.


Protein-ligand complex with 1B0L protein and (a) kanamycin, (b) furo(3,4-e)-1,3-benzodioxol-8(6h)-one, 6-(1,2,3,4-tetrahydro-6,7-dimethoxy), (c) 2-furancarbonitrile, (d) phenol, 2-methoxy-5-[(1,2,3,4-tetrahydro-6,7-dimethoxy-2-methyl-1-isoquinolinyl)], (e) benzeneacetic acid, 2-methoxyphenyl ester.
4. Discussion
Endophytic fungi are widely recognized as prolific producers of structurally diverse secondary metabolites with significant biological activities, particularly antibacterial and antioxidant properties.35,36 In the present study, endophytic fungi were isolated for the first time from different tissues of Alocasia macrorrhizos, expanding our knowledge of endophyte diversity associated with this medicinal plant and supporting previous observations that tropical plants harbor metabolically rich fungal communities. 37 The recovered isolates belonged to taxonomically diverse genera commonly reported from plant endospheres, suggesting that A. macrorrhizos provides a favorable ecological niche for endophytes with biosynthetic potential. Notably, the variation in antibacterial and antioxidant activities among the isolates reflects the strain-specific nature of secondary metabolite production, which is strongly influenced by host association, ecological adaptation, and genetic diversity.36,37 Although one isolate affiliated with Tulasnella calospora displayed sequence divergence suggestive of intraspecific variation, its definitive classification as a novel variant cannot be established without comprehensive multi-locus phylogenetic analysis, as recommended by contemporary fungal taxonomic frameworks. 38
Antibacterial screening revealed differential activities among the fungal extracts, with AMR-F4 and AMC-F6 exhibiting the most pronounced inhibitory effects against both Gram-positive and Gram-negative bacteria. This observation aligns with previous findings that Gram-positive bacteria are generally more susceptible to fungal metabolites due to their comparatively less complex cell wall structures.39,40 The broad-spectrum activity of AMR-F4 (Paecilomyces lecythidis) and AMC-F6 (Aspergillus fumigatus) is consistent with previous studies showing that Paecilomyces and Aspergillus species demonstrated antibacterial efficacy through the production of compounds such as paecilotoxins, helvolic acid, and fumigaclavines. 41 The concentration-dependent increase in inhibition zones supports prior findings, indicating that antibacterial activity correlates positively with extract concentration. 42 The comparable inhibition levels between AMR-F4 and the standard antibiotic kanamycin further suggest that these fungal metabolites possess substantial antimicrobial potency.
In antioxidant assays, all fungal extracts exhibited measurable DPPH radical scavenging activity, with AMC-F3 displaying the strongest effect, followed closely by AMR-F4 and AMC-F6. The antioxidant capacity of these isolates is comparable to values reported for endophytic Penicillium and Aspergillus species.39,40 FRAP assays further validated the electron-donating ability of AMR-F4 and AMC-F6, which demonstrated a reducing power exceeding 90% at 100 μg/mL, consistent with the results reported by Almanaa et al., who noted similar activity in A. terreus and P. citrinum. 43 Such activity is likely attributable to phenolic, flavonoid, and terpenoid compounds capable of donating electrons or hydrogen atoms to neutralize free radicals. 44
GC–MS analysis of the ethyl acetate extracts from Paecilomyces sp. (AMR-F4) and Aspergillus sp. (AMC-F6) revealed a chemically diverse profile of volatile and semi-volatile secondary metabolites, highlighting the biosynthetic potential of these endophytic fungi. A total of 47 compounds were detected in AMR-F4 and 39 compounds in AMC-F6, comprising amides, esters, sulfur- and nitrogen-containing heterocycles, and phenolic derivatives—chemical classes frequently implicated in antimicrobial and antioxidant activities.45,46 Several of the identified metabolites have been previously reported in bioactive fungal extracts, supporting their relevance to the observed biological effects. Among the predominant constituents, sulfur-containing heterocycles such as 3H-1,2-dithiol-3-one and thiomorpholine derivatives are of particular interest. Dithiol-based compounds participate in thiol–disulfide exchange reactions, thereby interfering with bacterial redox homeostasis and enzyme function, especially within redox-sensitive metabolic pathways. Similarly, thiomorpholine analogs disrupt bacterial membrane integrity and inhibit key enzymatic targets through sulfur-mediated interactions, thereby contributing to growth inhibition rather than nonspecific toxicity.47,48 These mechanisms provide a clearer biochemical rationale for the antibacterial activity associated with the sulfur-rich metabolites detected in both isolates.
Furan derivatives and phenolic compounds identified in the extracts are recognized for their ability to quench reactive oxygen species through electron-donating and hydrogen-transfer mechanisms, contributing to antioxidant capacity. In addition, substituted furans interact with bacterial membranes and intracellular proteins, leading to impaired cellular respiration and membrane destabilization.47,48 The presence of long-chain amides and fatty acid derivatives, including undecanamide and tetradecanamide, further supports the antibacterial potential of the extracts, as such compounds can alter membrane fluidity and permeability in Gram-positive and Gram-negative bacteria. Moreover, these metabolites have been previously isolated from Paecilomyces and Aspergillus species with confirmed antimicrobial activity. 49
The identification of 2-nitropropanoic acid methyl ester as a major metabolite in AMC-F6 is noteworthy, as nitro-containing esters are known to exert bacteriostatic effects through redox cycling and electrophilic interactions with cellular nucleophiles, thereby inhibiting essential metabolic enzymes. Importantly, the co-occurrence of multiple bioactive compound classes within each extract suggests that the observed antibacterial and antioxidant effects likely arise from additive or synergistic interactions, rather than from the action of a single dominant metabolite. Such multi-component bioactivity is well documented in fungal secondary metabolite systems and enhances biological efficacy while reducing resistance development.50–52 Overall, the GC–MS profiles demonstrate that A. macrorrhizos-associated endophytic fungi produce structurally and functionally diverse metabolites with defined biochemical roles. These findings provide a mechanistic basis for the observed biological activities and reinforce the potential of these endophytes as promising sources of pharmacologically relevant natural products for further isolation, characterization, and drug development.
To complement the biochemical assays, molecular docking studies were conducted to elucidate the potential interactions between the major GC–MS-identified metabolites and the bacterial target protein 1B0L (Lactotransferrin). The docking simulations, validated using Lipinski’s Rule of Five and ADMET parameters,53,54 showed that several compounds from the AMR-F4 and AMC-F6 extracts exhibited favorable binding affinities relative to the standard antibiotic Kanamycin (–7.1 kcal/mol). Notably, Furo(3,4-e)-1,3-benzodioxol-8(6H)-one, 6-(1,2,3,4-tetrahydro-6,7-dimethoxy-) displayed the strongest binding energy, suggesting stable accommodation within the protein’s binding pocket. Compounds such as 2-furancarbonitrile, 1H-indole, 1-butyl-3-methyl-, and 3,4-dihydroisoquinoline, 1-[3-methoxybenzyl]-6-methoxy- also demonstrated favorable binding energies and extensive residue interactions, which may contribute to the antibacterial effects detected in vitro. By contrast, ligands with weaker binding affinities exhibited fewer stabilizing interactions, indicating a reduced contribution to target engagement. Although docking results do not confirm biological activity, these findings support the experimental antibacterial observations and highlight specific fungal-derived compounds for further mechanistic validation using enzymatic assays and in vivo models. The consistent engagement of catalytically active residues implies that these metabolites may interfere with microbial iron acquisition and disrupt cell wall integrity, thereby providing a plausible molecular basis for their antibacterial action. 55
Collectively, the chemical, biological, and molecular findings confirm that endophytic fungi isolated from A. macrorrhizos are rich sources of bioactive metabolites with significant antibacterial and antioxidant activities. The strong agreement between GC–MS composition, in vitro assays, and molecular docking outcomes underscores the pharmaceutical potential of these isolates. The results are consistent with those reported by Shaaban et al. and Naveen et al., highlighting the untapped capacity of filamentous fungi to yield novel antimicrobial and antioxidant compounds.39,41 However, the present study was limited to crude extracts; the isolation and structural elucidation of individual active components are required to define precise mechanisms and therapeutic relevance. Future studies should therefore employ bioassay-guided fractionation, spectroscopic characterization (LC–MS, NMR), and in vivo evaluation to confirm efficacy and safety.
5. Conclusions
The present study establishes Alocasia macrorrhizos as a previously unexplored reservoir of biologically active endophytic fungi and demonstrates that its associated endophytes possess antibacterial and antioxidant potential. Rather than emphasizing individual analytical approaches, our findings collectively indicate that specific endophytic strains associated with this medicinal plant can produce chemically diverse metabolites that translate into measurable biological effects. This observation supports the broader concept that plant–endophyte associations in tropical medicinal species represent an underutilized resource for natural product discovery. Importantly, the observed bioactivities highlight the functional relevance of endophytic fungal metabolites beyond their taxonomic identification or chemical complexity. The results reinforce the idea that endophytes contribute to the pharmacological value of their host plants and may serve as sustainable and scalable sources of bioactive compounds. Future work should focus on targeted isolation, structural elucidation, and mechanistic evaluation of individual metabolites, as well as on validating their efficacy and safety in biological models. Such efforts will be essential to translate endophytic fungal metabolites into viable candidates for combating antimicrobial resistance and oxidative stress–related diseases.
Supplemental material
Supplemental material - Bioprospecting endophytic fungi, Paecilomyces lecythidis and Aspergillus fumigatus, from Alocasia macrorrhizos as potential sources of bioactive agents
Supplemental material for Bioprospecting endophytic fungi, Paecilomyces lecythidis and Aspergillus fumigatus, from Alocasia macrorrhizos as potential sources of bioactive agents by Amina Khatun, Rajib Das, Abdul Ali Bhuiyan, Nowsad Zahan Sathi, Sazzad Mahmud, Md. Selim Uddin, and Shariful Haque in Science Progress.
Footnotes
Acknowledgments
The authors would like to express their gratitude to the Department of Pharmacy, Pabna University of Science and Technology, Pabna for their invaluable assistance, resources, and infrastructural facilities necessary to perform this research. The use of AI tools solely for language refinement and confirming that no scientific data, analyses, or results were generated or modified using AI. The authors affirm full responsibility for the accuracy, originality, and integrity of the submitted work, in accordance with Sage’s AI policy.
Author contributions
Conceptualization and methodology, M.A.K., and S.H.; experimentation, M.A.K., S.M., and R.D. data analysis, M.A.K., M.S.U., R.D. and M.N.Z.S.; investigation, M. A.K., A.A.B., S.M., and S.H.; writing—original draft preparation, M.A.K., R.D., M.N.Z.S. & A.A.B; writing—review and editing, M.A.K, M.S.U., R.D., A.A.B., and S.H.; supervision, S.H. All authors have read and agreed to the published version of the manuscript. Authors: Mst. Amina Khatun (M.A.K), Rajib Das (R.D.), Abdul, Ali Bhuiyan (A.A.B.), Mst Nowsad Zahan Sathi (M.N.Z.S.), Sazzad Mahmud (S.M.), Md. Selim Uddin (M.S.U.), Shariful Haque (S.H.).
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data will be available on request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.