
Abstract
Protected areas require long-term monitoring to understand the influence and extent of ecosystem stress to inform management and conservation decisions. As long-term data are not always available, paleolimnological methods offer a way of extending our knowledge of past environmental conditions necessary to use as context for remediation. Here, we examine four sediment cores and additional surface sediments from 14 ponds located on Sable Island National Park Reserve Canada (SINPR), where long-term ecological changes and vulnerability to disturbance are not well defined. We develop a paleolimnological approach to assessing environmental vulnerability through the use of biological indicators (Diptera: Chironomidae), where shifts in the environment are inferred by shifts in chironomid assemblages over time. Analysis of surface sediments show four distinct assemblage types reflecting four different habitat conditions; primarily represented by the presence of Glyptotendipes, Chironomus, Microtendipes, and Dicrotendipes. Differences in habitat conditions through time based on these results are then compared to biostratigraphic analysis of sediment cores from four of the ponds. We found that two ponds had large shifts in chironomids assemblages that were associated with changes in habitat over time, while two others that were not as exposed to the influence of erosion and influx of sand dunes did not. Our findings established a baseline of historical change in SINPR, broadening the scope of long-term monitoring, which is essential for defining goals for management and conservation of the ecological integrity of Sable Island.
Introduction
A common challenge for effective ecosystem management is sufficient long-term historical data to be able to understand natural variability over time. Vulnerability can be defined as a function of risk, resilience, exposure, sensitivity, and adaptive capacity of a system,1,2 but is difficult to quantify. 3 Paleolimnology is a science that uses proxies preserved in lacustrine sediments to make inferences about historic and prehistoric environments. 4 This expands the scope of monitoring and quantifies susceptibility to change in freshwater ecosystems, inherently providing baseline information necessary for informed management and conservation. 5 Establishing pre-disturbance or reference conditions of an ecosystem can provide evidence-based targets that may help inform restoration.6,7 Closed freshwater systems are valuable sentinels for change due to their unique ability to collect and chronologically preserve surrounding ecological information.8,9
The impacts of stress can often have cascading effects on aquatic biota, especially in responsive groups, such as the Chironomidae (chironomids). Chironomid assemblages are sensitive to environmental factors such as temperature, productivity, and landscape-related change.10–12 Thus, analysis of chironomids are useful to infer the environmental conditions that influence their abundance during their aquatic lifecycle. 4 Reconstructing past conditions can distinguish differences between an ecosystem’s natural variability (unforced) and major climate-driven changes (forced) 13 ; this leads to an understanding of the parameters impacting ecological integrity, which describes the extent that an ecosystem’s diversity and function is comparable to similar local natural ecosystems as well as its ability to support its own ecological processes. 7 Furthermore, targets for ecosystem restoration can be defined based on insight provided by the reconstruction of long-term changes and natural ecosystem trajectory, 14 having major implications for making informed management decisions.5,15
Informed management is essential for protected areas, which are often established with the intent of long-term conservation of ecosystem services. Sable Island National Park Reserve (SINPR) is an emergent sandbar in the Atlantic Ocean that is mandated as a National Park Reserve of Canada to maintain the ecological integrity of the island’s ecosystem. 16 The freshwater ponds of SINPR are vital for supporting the unique biodiversity of the island; however, these systems are also experiencing a number of environmental stressors, including saltwater intrusion events from storm-surge, windblown sand intrusion, increased evaporative stress, and nutrient inputs from wildlife.16–18 Additionally, the continuity of the fresh groundwater lens that replenishes the ponds is unknown and faces threats of flooding or sea level rise that could move seawater into the source. 19
A bioassessment using aquatic macroinvertebrates recorded 27 taxa in the ponds of SINPR, with the largest diversity reflected by chironomids. 20 As chironomids are well known to offer insights for the environmental conditions in which they develop, 10 a paleolimnological assessment of changes in subfossil diversity and abundance over time allows for both the establishment of a pre-disturbance condition, as well as inferences of responses to changes the environment.4,10 Here, we examine pond systems from SINPR to understand the implications of centuries of environmental stress on the island’s freshwater ecosystems. Due to the proximity of the marine environment, we hypothesize that modern chironomid assemblages of the freshwater ponds of SINPR will primarily reflect differences in lake morphology and water quality as a function of differences in lake catchment, and that the evolution of lake catchments over time will correspond to shifts observed in the subfossil assemblage. Through assessment of subfossil biological indicators from the surface sediments of 14 freshwater systems on SINPR as well as sediment cores from 4 of these 14 ponds, we examine historic ecological trends of the SINPR ponds in order to: (1) identify chironomids indicators that may differentiate the freshwater habitats; (2) identify trends in chironomids assemblages through time, and 3) infer causes of ecosystem stress to better inform management and conservation. We aim to demonstrate how paleolimnological assessment of long-term environmental change in freshwater systems can be used to address issues of conservation and vulnerability and responses to stress. Establishing baseline conditions and the pre-disturbance context of these systems will be integral to helping Parks Canada determine the extent of risk facing the freshwater ecosystems on SINPR, and help define necessary goals for management and preservation of the ecological integrity of the island.
Study site
Sable Island is a 30 km2 emergent sandbar, 49 km long, 1.5 km wide at its maximum, and 161 km southeast from the nearest point of mainland Nova Scotia16,21 (Figure 1). Sable Island is the only emergent land near the edge of the continental shelf, 16 with a highly dynamic form influenced by winds, waves, ocean currents, and storms. 22 Several freshwater ponds across the island are vital to supporting its biodiversity. Many ponds contain macrophyte communities that include pondweed or watermilfoils, and herbaceous communities such as cranberry heath and other low growing species on pond edges. 23 We examined 14 isolated ponds on SINPR that are part of an aquatic monitoring program conducted by Parks Canada. 20 These 14 sites represent most/all (as of 2020) of the freshwater systems (e.g. we excluded ponds that were predominantly salt-water) on Sable Island that have a sufficient volume of water to allow for sampling.

Map of Sable Island National Park Reserve and the freshwater ponds selected for sampling. Cores were extracted from PD03, PD37, PD38, and PD67, indicated by the red boxes, and surface samples were collected from all ponds indicated.
The vegetation of SINPR is influenced by a moderate climate, cooler growing season temperatures, infrequent frosts, 23 high winds, and sea spray. 24 Terrestrial vegetation is characterized by sparse grasslands, heath, and marram grass, 23 with a variety of low growing shrubs such as black crowberry (Empetrum nigrum). 24 A population of genetically distinct feral horses resides on SINPR, descending from settlement and agriculture attempts in the mid-eighteenth century. 25 With a population that had fluctuated from 150 to over 250, the horses were given legal protection from human interaction in 1960, and have since reached over 500 members.26,27 In their free roam of the island, the horses are known to affect the vegetation through trampling and grazing, with the greatest influences on marram grassland communities and vegetation near freshwater ponds. 28 The population is not currently managed.
Methods
Field sampling
During August 2019, an Uwitec gravity coring device (8.4 cm tube diameter) was used to collect sediment cores from the middle of four ponds (PD03, PD37, PD38, and PD67) using an inflatable boat (Figure 1). Cores were divided into 0.5 cm intervals in the field; the length of the cores collected were 25, 12, 17, and 15-cm, respectively. The sediment core of PD03 and PD37 each had large amounts of sand visible in the cores; PD03 had a sand interval from 5.5–10 cm, and PD37 was predominantly sand from 7 cm to the base of the core (12 cm). While PD67 had macrophytes at the very top of the core and small amounts of sand visible at 1 cm, the core consisted of brown gyttja for much of the entire record. PD38 consisted of gyttja for the entirety of the core until a basal sand interval was found at 17 cm. Each extruded interval was placed in labeled Whirl-Pak® bags, transported in a cooler, and stored at 4 °C for processing. Additional samples from the surface sediments of 10 additional ponds were also collected using the same approach; however, only the top 1-cm of sediments were processed (Supplementary Table S1).
Water quality parameters were also collected monthly (May – August) from 2015–2020 by Parks Canada for the 14 ponds on SINPR (Figure 1) as part of an independent water quality monitoring program; further details about the water monitoring program can be found in Jacks et al. (2021). 20 Temperature, dissolved oxygen, conductivity, pH, salinity, and turbidity were measured with the use of an YSI multi-parameter probe at the same time as water samples were collected. Pond water samples were collected from the middle of each pond by hand and tested for major ions, nutrients, and metals at the Environment and Climate Change Canada (ECCC) laboratory in Moncton, New Brunswick. We related differences in chironomids assemblages to water quality in the supplemental information (Table 1).
Location and basic parameters of ponds cored for analysis on Sable Island National Park Reserve. Environmental parameters measured on August 20, 2020.

Laboratory analysis
Sediments from both surface samples as well as selected intervals of the four sediment cores were processed for subfossil chironomids following standard methods. 10 Not all intervals collected were processed. Each sample was treated with potassium hydroxide (KOH) at 75 °C for 30 min, with intermittent stirring, to deflocculate the sediment. The residual was then poured through nested sieves of 212 and 106 µm, and rinsed with 95% ethanol. While PD03, PD37, and PD67 were analyzed at 1-cm intervals, only a partial analysis of PD38 was completed as few discernable differences in the chironomid assemblages were apparent; samples were completed from the sediment intervals of 0.5, 5, 10, 15, and 17 cm.
Subfossil chironomids were sifted and counted for each sample with the use of a dissecting microscope and fine forceps. A minimum of 50 head capsules from each interval were collected as per recommended methods, 29 with the exception of the intervals between 6.5–9.5 cm for Pond PD03 due to extremely low abundances. For the other intervals, if less than 50 head capsules were found then more sediment was subsampled until total identifiable remains were greater than 50. The collected specimens were then permanently mounted on glass slides using Entellen® then identified using a Stereo microscope. Head capsules were identified to the lowest possible taxonomic resolution with the use of Oliver and Roussel (1983) 30 and Brooks et al. (2007). 31
Dating analysis
Chronologies of each core were established using alpha 210Pb dating techniques at the University of Ottawa by Chronos Scientific Ltd (formally myCore). The Constant Rate of Supply (CRS) model 32 was used to establish the age-depth relationship with linear interpolation used beyond the range of the CRS models. To determine percent organics and percent carbonate content throughout each core, weight loss on ignition (LOI) analysis (following Heiri et al. 2001 33 ) was carried out for each 0.5 cm interval by measuring loss following sequential burns of sediment at 550 (4 h) and 950 °C (2h).
Statistical and numerical methods
Data analysis were performed using R statistical software v4.2.0. Relative abundances were calculated as the percentage of identifiable chironomids in each sample. Rare taxa were removed from analysis; only taxa representing > 2% relative abundance in at least 2 ponds were retained. Taxa represented by their relative abundance were plotted stratigraphically over depth and time. A constrained hierarchal cluster analysis of chironomid assemblages using the function ‘chclust’ of the rioja package was used to describe the biostratigraphic results; where the number of zones for each core was determined a broken stick model. 34 The concentration and accumulation of subfossil chironomids were calculated as head capsules per gram of dry weight (HC g−1 DW), and the number of head capsules per dry density of each interval over time (HC cm−2 yr −1), respectively.
A hierarchical cluster analysis of the relative abundance of surface samples was used to differentiate between sites, where the Ward’s D2 method grouped ponds based on minimum variance and squared dissimilarities into clusters as determined by a Mantel-optimal number. 35 Significant chironomid indicators that represented each cluster were determined through Indicator Species Analysis (IndVal 36 ) using the R package labdsv. A Principal Component Analysis (PCA) was then used to outline the trends in the chironomid communities of each core over time plotted passively across the chironomid assemblages identified from the surface samples using the function ‘timetrack’ in the R package analogue. 37
Results
The modern environment
More than 1200 subfossil head capsules were extracted from the 14 surface samples, with an average of 91 head capsules (range = 54.5–143.5). The most dominant taxa were Microtendipes (17.85% relative abundance), Paratanytarsus (16.8%), Tanytarsus (13.86%), Cricotopus (11.28%), Dicrotendipes (10.34%), Glyptotendipes (8.73%), and Chironomus (7.83%). A hierarchical cluster analysis of the 14 ponds showed four distinct clusters based on the Mantel-optimal number, and four distinct habitat types based on chironomid indicator taxa (Figure 2). The taxa that were statistically significant (p < 0.05) for each cluster were Glyptotendipes (p < 0.01) for Cluster 1 (PD03, PD66, and PD67); Chironomus (p < 0.05) for Cluster 2 (PD06, PD07, PD63, PD64, and PD70); Microtendipes (p < 0.05) for Cluster 3 (PD37, PD47, PD51, and PD53); Dicrotendipes (p < 0.01) for Cluster 4 (PD38 and PD52). Paratanytarsus had a non-significant p-value of p = 0.08, but was considered to have influence in determining the assemblage of Cluster 2.
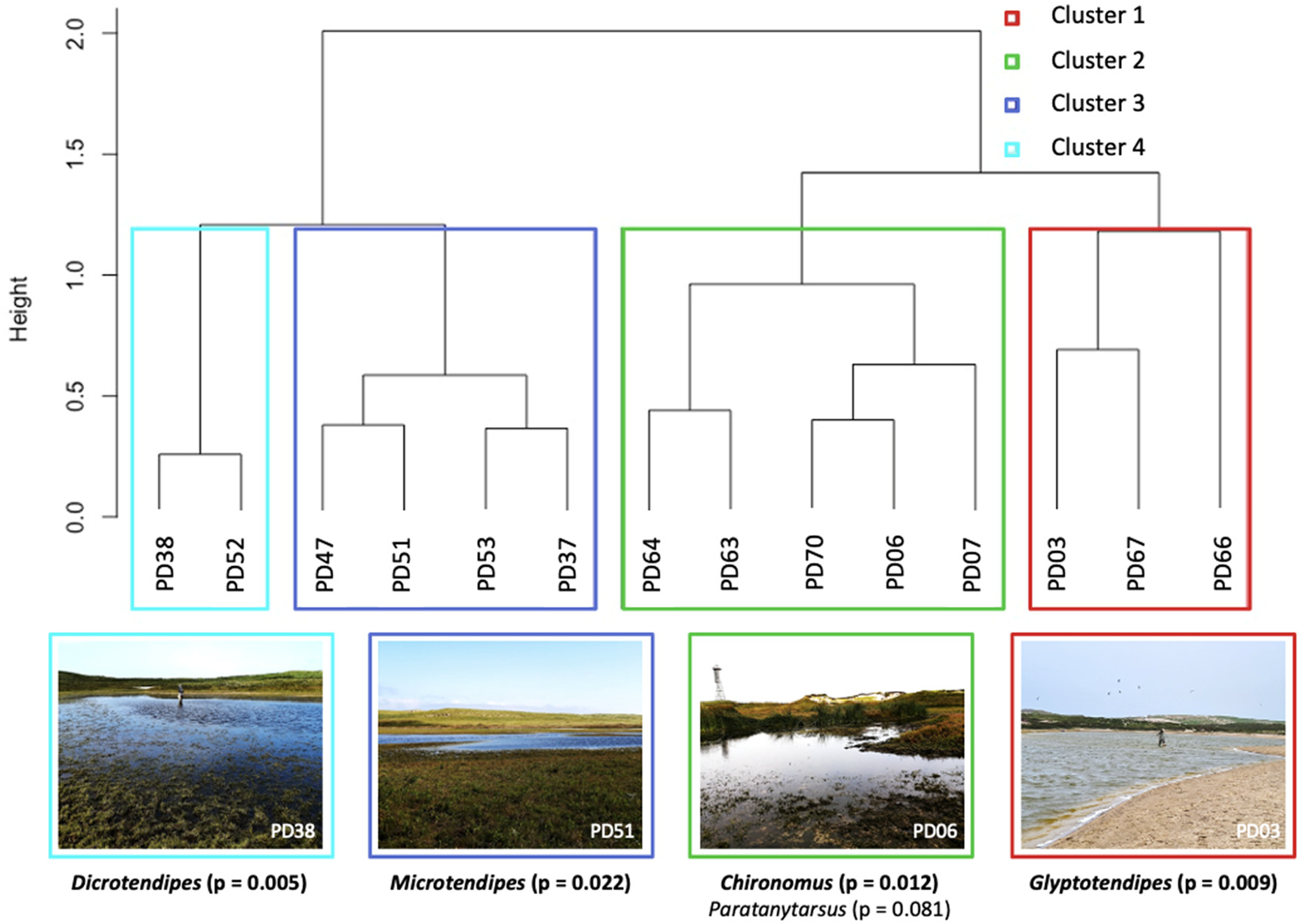
A hierarchical cluster analysis of all SINPR ponds using Ward’s minimum variance method and the Mantel-optimal number of clusters. The significant indicator taxa and a photo of a pond from each cluster is included beneath. The p-value for Paratanytarsus is > 0.05, but is still noted for its influence in determining Cluster 2.
Water quality measurements from 2015–2020 were compared for all the ponds based on the parameters included in Jacks et al. (2021). 20 The values for chloride, dissolved organic carbon (DOC), nitrogen (TN), pH, phosphorus (TP), and turbidity were compared between ponds (Supplementary Figure S5). As Jacks et al. (2021) 20 described small differences in water quality between seasons and years, we then averaged the parameters (from 2015–2020) for the ponds in each identified cluster from our hierarchical cluster analysis; notable overall differences are observed in the average concentrations of TP and TN between the clustered ponds (Table 2). Cluster 1–4 have TN concentrations of 2.4, 1.9, 2.3, and 1.1 mg/L TN, respectively. Cluster 1–4 have TP concentrations of 0.46, 0.28, 0.41, and 0.11 mg/L, respectively. We averaged 5 years worth of water samples as a reflection of the fact that the surface sediments (the upper 1 cm of sediments) sampled would likely represent multiple years of subfossil chironomids.
Indicator taxa and the average water chemistry per cluster (values were averaged for all of the lakes in the cluster from 2015-2020 water samples) based on Ward’s minimum variance method of all sampled SINPR ponds.

*Indicates water quality data is missing.
Sediment core chronology
The chronology for PD67 (Figure 3(a)) was established based on 22 210Pb measurements. The level of unsupported 210Pb activity decreased from 1.202 to 0.0075 Bq/g between 1.0–12.0 cm; the lowermost three samples were below background. A CRS model was used to estimate the age of each interval from 1–11 cm, and linear interpolation thereafter to a basal date of 1767 at 15 cm. The chronology for PD03 (Figure 3(b)) was established based on 8 210Pb measurements. The level of unsupported 210Pb activity decreased from 0.650 Bq/g at 1.0 cm to 0.563 Bq/g at 5.0 cm. An influx of sand content between 5.0 and 10.0 cm caused an unconformity in the unsupported 210Pb activity below 5.0 cm. As such, we used a linear interpolation based on the CRS model for each interval from 10.0–25.0 cm, subtracting the sand-interval from the chronology. Each interval from 5.0–10.0 cm was assigned a date of 1960, followed by the linear interpolation of dates to the basal section of the core. Due to the difficulty in estimating age for PD03 beyond 5 cm, dates prior to 1950 should be interpreted with caution. Chronologies for PD37 and PD38 were established based on 8 and 9 210Pb measurements, respectively (See supplemental materials).

Age-depth model for (a) PD67 and (b) PD03 based on 210Pb and linear interpolation.
Biostratigraphic analysis
A total of 1326 subfossil chironomid head capsules (HC) were enumerated from the lacustrine sediment core of PD67. There were 17 different taxa found in the 18 sampled intervals (HC per interval: average = 73.75; and range = 52–129.5). PD67 had an average concentration of 43.5 HC per gram of dry weight (HC g−1 DW), with a range of 12.3 to 59.3 HC g−1 DW throughout the core, and a peak of 150.2 HC g−1 DW in 2012 CE. We inferred changes in the ecosystem based on known environmental tolerances of chironomid indicators during three significant time periods as indicated by a constrained cluster analysis; assemblage shifts for PD67 occurred at circa 1950 and ca 1990 (Figure 4(a)).

Biostratigraphy of chironomid relative abundance (%) in cores (a) PD67 and (b) PD03 plotted over depth and time. The light grey represents a 10x exaggeration of low abundances for increased visibility. Sand intervals in PD03 are indicated by horizontal grey bar.
The base of the core, designated Zone 1, 15 to 8 cm (ca 1770 to 1950 CE), was a period defined by a dominance of Microtendipes (40% average relative abundance) and Glyptotendipes (32%). Cricotopus (5%) and Paratanytarsus (7%) were also found in smaller abundances during this zone (Figure 4(a)). Microtendipes peaked at 64% in ca 1800 CE (14.0 cm), then decreased to ∼43% ca 1950. Microtendipes pedellus, which is the likely taxa found on Sable Island based on Jacks et al. (2021) 20 examination of larvae, is a collector-gatherer 38 known to prefer coarse sediments low in organic content. 31 Glyptotendipes, which have been associated to mesotrophic ponds and macrophytes that are mined by many species of the genus,39,40 increased from 23 to 52% between ca 1770 and 1885 CE (15.0–11.0 cm). Paratanytarsus, which is associated with aquatic vegetation,31,39,41 increased from 3 to 7% between ca 1770 and 1950 (15–8 cm). Cricotopus remained < 7% between ca 1770 CE and 1950 (15.0–8.0 cm).
During Zone 2, from 8.0 to 4.0 cm (1950–1990 CE), the diversity of the assemblage increased from ∼8 to 13. Glyptotendipes decreased from a high of 40% of the assemblage to ∼19% from ca 1950 to 1990 CE and Cricotopus and Paratanytarsus increased from 5 to 18% and 7 to 19%, respectively. The genus Cricotopus is comprised of many taxa, however, they are not easily distinguishable from each other as subfossils. Based on our morphological identifications, we feel Cricotopus sylvestris, Cricotopus cylindraceus, and Cricotopus elegans are likely those found within the sediment record. The most recent period (Zone 3), from 4–0.5 cm (1990 to 2019), diversity decreased from a high of 13 to 10, with notable increases in the Tribe Tanytarsini. Several taxa of Tanytarsini were identified by Jacks et al. (2021) 20 were identified, but due to difficulty in identification of worn subfossil specimens, we were often unable to identify them to a higher taxonomic resolution. However, undifferentiated taxa of the Tanytarsini increased from < 2 to 5–10% of the total assemblage in Zone 3. Cricotopus increased from 18% at 1990 to > 40% at the top of the core. Paratanytarsus also increased to ∼30% of the assemblage during this interval. In contrast, abundances of Glyptotendipes further declined from 19% at 1990 to < 5% of the total assemblage at the top of the core.
For PD03, a total of 1369 HC were enumerated with 26 different taxa found in the 25 sampled intervals (HC per interval: average = 59.52; and range = 7–106). PD03 had an average abundance of 201.2 HC g−1 DW, with a range of < 5 to 502.5 HC g−1 DW. Abundances of < 20 HC g−1 DW occur between 6 and 10 cm during the sand layer where < 50 head capsules were collected due to extreme low abundances. Prior to the sand intervals from 25 to 10 cm (ca 1860–1960 CE), maximum HC accumulation was 157.5 HC g−1 DW. After the sand intervals, the concentration of HC ranged from 148.4 to 502.5 HC g−1 DW. The sand layer between 7–9.5 cm was the significant zone outlined by our constrained cluster analysis with a broken stick model (Figure 4(b)). As abundances were extremely low during this period, we discuss PD03 with respect to its two primary intervals, from 25 to 10 cm (Zone 1) and 5 to 0.5 cm (Zone 2). While we have attempted to interpolate dates prior to the sand interval, we can only discuss pre-sand from post-sand assemblages with any certainty, and interpolated dates prior to 1950 should be interpreted with caution.
Prior to 1960, the assemblage of Zone 1 of PD03 was dominated by Chironomus Cricotopus, Glyptotendiepes, Microtendipes, and smaller abundances of Orthocladius and Corynoneura (Figure 4(b)). Abundances of Cricotopus increased from 18 to 41% between the base of the core and the sand interval (25–10 cm). Glyptotendipes reached a maximum abundance of 28% in 1900 CE (20.0 cm) but decreased following ca. 1930 to ∼10% of the record. Likewise, Microtendipes pedellus also declined from its maximum abundance of 20% in 1920 CE (15.0 cm) to 5% ca. 1930 for the remainder of Zone 1. Chironomus anthracinus-type decreased from 4% from the base of the record to 1920 CE to < 1%, while abundances of Chironomus plumosus-type, an indicator of eutrophication, 42 decreased from 10% at 15 cm (1920 CE) to ∼10% for the remainder of Zone 1. Dicrotendipes, which is associated with littoral areas and mesotrophic conditions,40,43 were abundant only in the basal interval (31% in ca 1860 CE at 25.0 cm), then remained < 8%.
Between 1950 and 2019 CE (5–0.5 cm), during Zone 2, Glyptotendipes ranged from 15 to 20% and Microtendipes pedellus remained < 6% compared to Zone 1 where both taxa were found at higher abundances. Chironomus anthracinus-type was only found as < 2% of the assemblage in Zone 2, while Chironomus plumosus-type, which is often associated with productive conditions, 31 was 5–10% of the assemblage. Endochironomus increased from 3–7% and Polypedilum from 5–10% of the assemblage during Zone 2. The predatory chironomid Procladius was also found in small abundances during Zone 2 and increased slightly to 5% of the total assemblage from 2010 to 2019.
A total of 515 HC were processed for PD37; however, we were only able to obtain > 50 specimens from 0.5–7 cm of the core. Low abundances (1–10 HC) were observed until 12 cm, after which no chironomids were present in the core (Supplemental Figure S3). We enumerated 368 HC from PD38 in the five intervals processed (Supplemental Figure S4). Chironomids were present in sufficient abundance throughout the 17 cm core. Both PD37 and PD38 had a similar chironomids assemblage, dominated by high abundances of Microtendipes, Dicrotendipes, and Cricotopus. While the assemblages were similar in both lakes, PD37 did not contain a sufficient amount of chironomids for interpretation from 7.0 cm to the base of the core. The only difference observed in PD37 from our constrained cluster analysis with a broken stick model was the point at which low abundances of chironomids were found pre-1910 (7.0 cm). While PD38 had a sufficient amount of chironomids available for analysis, we did not find a discernable difference between intervals processed to warrant a full analysis, and no significant zones were noted. Assemblage data for both PD37 and PD38 can be found in our supplemental information.
A PCA demonstrated the trajectory of each core interval passively across the assemblages of the surface samples from the 14 sampled ponds (Figure 5). Variation of the downcore assemblages across the PCA shows periods of time where conditions may have been similar to the assemblages of present-day ponds. The surface interval of both PD67 and PD03 show similarities, reflected by higher abundances of Cricotopus. High abundances of Microtendipes downcore (prior to 1820 CE) distinguished PD67 from other ponds, and as the abundance decreased it became more similar to the assemblages of present-day ponds with higher proportions of Glyptotendipes (from 1830–1980), and subsequently by Cricotopus (from 1980-present), reflecting a present-day vegetated catchment influenced by increasing amounts of sand. The trajectory of the downcore PD03 record shifted towards PD38 and PD52, which had similar high of Dicrotendipes in the present-day assemblage. An increase in Cricotopus abundance since 1890 CE makes PD03 dissimilar to most other ponds. In contrast, both PD37 and PD38 had little change between their past and current chironomid assemblages. As such, we have used PD37 and PD38 as a reference for a habitat that is primarily represented by heath, as is the current present-day environment. However, surface samples did show some differences between the two systems with noted abundances of Chironomus plumosus-type in recent sediments of both ponds, and higher abundances of Dicrotendipes for PD37 in contrast to higher abundances of Microtendipes for PD38 (Figure 2).

PCA biplot of chironomid communities from the surface samples from each pond sampled and the passive trajectories of each sediment core. Note differences between surface sediment samples and the core intervals are expected; surface sediments (grey dots) were collected August of 2019 and represent the contemporary chironomids assemblages.
Discussion
Little is known about the baseline conditions of the freshwater systems of SINPR. While PD67 is distinguishable on a hand drawn map produced in 1766, 44 most of the other ponds are not shown and some of these areas are instead covered by a large saltwater lagoon. SINPR has long had a history of shifting sands from coastal erosion processes, whereby sediment erodes from the western tip and accumulates at the eastern tip, resulting in gradual migration of the island.44,45 Additionally, shoreline retreat, overwash processes, and windblown sand have resulted in burial of vegetation and loss of some freshwater ponds. 18 Harsh coastal weather and intense storms are also known to cause infiltration of ponds with sand, as well as alter pond chemistry. 46 Most freshwater chironomids are intolerant to saltwater; however, we found no evidence of saltwater intrusion events from the water quality monitoring performed by Parks Canada, albeit records only exist from the establishment of the park in 2013. Here lies the opportunity for paleolimnological methods to fill this knowledge gap. In order understand the extent of change for optimal park management, we focus on how ponds have evolved over their paleohistory. Although there are limited comparable freshwater ecosystems located on sandbar islands in the Atlantic Ocean, we are still able to describe changes in ecological condition through shifts in chironomid assemblages in a paleolimnological approach as the baseline context for ecosystem management.
Ecosystem changes
The chironomid assemblages surveyed from recent conditions of the 14 freshwater ponds of Sable Island provides perspective of the habitats and associated chironomids assemblages that exist on the island today. We combine past knowledge of water quality and benthic invertebrate assemblages 20 with additional characterization of primary differences between ponds through the assessment of the surface sediment samples for subfossil chironomid comparisons. Presently, both PD67 and PD03 have sandy catchments and littoral zones with nearby grassland, as opposed to many of the other ponds that have surrounding heath communities and are more densely vegetated, such as PD37 and PD38. In our cluster analysis of the ponds, Cluster 1 (PD03, PD66, and PD67), defined by the presence of Glyptotendipes, can be associated to a grassland catchment and higher nutrient conditions. Clusters 2–4 are associated with productivity (mainly inferred from the presence of Chironomus plumosus-type), a coarse sandy habitat surrounded by heath vegetation (Microtendipes), and mesotrophic conditions (Dicrotendipes), respectively (Figure 2). Although PD67 and PD03 have similar communities today, they had divergent communities in the past. Shifts in the abundances of key indicator taxa throughout the lacustrine sediment cores allow for indication of habitat change over time, as shifts in the chironomid assemblages indicate a change in ecosystem conditions.
The chironomid assemblages of PD67 experienced a shift from higher abundances of Microtendipes and Glyptotendipes in the past to Cricotopus and Paratanytarsus in recent sediments (Figure 4(a)). The decrease observed in Glyptotendipes and Microtendipes, particularly ∼1990, suggests the community assemblage responded to changes in sediment habitat over time. The increased abundances of Cricotopus and Paratanytarsus in the core since 1969 CE could reflect the prominence of submerged macrophytes and a sandy catchment with marine influence, consistent with the onset of present-day conditions. Whereas conditions of the past likely reflected those of PD64 in the past (Figure 2), which is a smaller, shallower, system surrounded by health vegetation. While PD03 has a similar present-day chironomid assemblage as PD67, it was different prior to 1964 CE, when a large inflow of sand occurred (between 6.0 and 10.0 cm depth in the sediment record). Prior 1930 CE, the assemblage of PD03 appeared more like PD70 (Cluster 2; Figure 2), with higher abundances of Microtendipes, Dicrotendipes, Glyptotendipes, and Chironomus, which suggests a shallow system with coarse sediment, macrophytes, and a densely vegetated catchment that included grasslands and heath. Following the sand interval ∼ 1960, abundances of Cricotopus increased 10–15%, likely reflecting receding grass cover and increased sandy margins as seen today (see Supplementary Figure S6). Increased %Organic content of sediments after the sand intervals alludes to increased productivity following 1964 CE (Supplementary Figure S1). The present-day chironomid assemblage of PD03, with notable abundances of Glyptotendipes, Endochirnomus, and Polypedilum, are common indicators of nutrient-rich waters.31,39,42 Water quality measurements from 2015–2020 also show that TN and TP concentrations are higher in PD03 than the 12-pond average (Supplementary Figure S2). However, Cricotopus and Glyptotendipes are also present prior to the sand intervals, as well as a high abundance of Chironomus, suggesting that PD03 has always been relatively productive. High abundances of C. plumosus-type suggest lower oxygen conditions have long existed in PD03, although productivity has increased post-sand deposit.
Potential causes of ecosystem stress
With intensifying global environmental change, climate variability is expected to accelerate and increase the frequency and severity of storms. 47 These threats could increase likelihood of storm-induced habitat shifts, or even a change from a freshwater to brackish ecosystem as seen in some ponds in the past.18,21,48 In addition to sea level rise, these projections of climate variability threaten isolated oceanic islands whose stability is of concern with these increased risks. 49 SINPR is located on the storm track for low-pressure systems, and is known as the windiest place in Nova Scotia 50 ; its offshore location and exposure to harsh weather causes increased vulnerability to environmental changes and further habitat shifts.
The sand intervals observed in the PD03 record below 5.0 cm (1964 CE) seem to have been caused by a severe weather event. Based on historical aerial imagery of the island dating back to 1959, it appears PD03 was a more open system in the past (JBR Eamer, personal communication, June 25, 2021). A group of the island’s western ponds, slightly east of PD03, experienced storm overwash entering the system from the south beach caused by dune breaching prior to 1971; active terrain management eventually restored the dunes and closed off the system to prevent inflow in 1991, but another breach eventually caused those ponds to become brackish. 50 While it is not clear if PD03 also experienced the same impacts, the sand influx seen in the core occurred prior to 1963 and was possibly a result of the same storm surge that affected the ponds to the east. Aerial imagery of PD03 from 1963 appears to show depositional input of sand, with an open path to the south beach where the sand would have likely come from (Supplementary Figure S3). While there is no clear indication of saltwater intrusion in PD03, the pond could have experienced recovery from saltwater in addition to the sand input. The extreme low abundance of chironomids in the sand intervals are likely a direct result of a significant source of material entering the lake basin at a single point in time. The intrusion of sand in the catchment of PD03 is predicted to have caused the shift to the habitat seen today.
The unmanaged, naturalized horse population could also be increasing nutrient enrichment to the freshwater systems of Sable Island. Although pond shorelines make up less than 1% of the island’s area, they experience disproportionately frequent grazing and trampling, which alters the surrounding vegetation and catchment. 28 While these impacts cannot be definitively attributed to the horses, there is a high possibility that the increasing abundance causes further reduced vegetation cover and increased erosion, 51 as well as higher rates of defecation and runoff into the ponds, resulting in nutrient enrichment.7,52 In a study on the effects of agricultural land-use on chironomid communities, Campbell et al. (2009) 39 found that cattle grazing had a particularly strong impact on water quality and chironomid compositions by disturbing pond margins and increasing turbidity and nutrient concentrations. We found similar contexts for both PD67 and PD03, where habitat-driven changes are associated with elevated nutrients, mostly occurring after 1960 CE; this is demonstrated in the stratigraphic constrained cluster analyses for both ponds where chironomid assemblages are statistically different after 1960 CE (Figure 4), coinciding with the protection and increase of the horse population. Nutrient enrichment in the ponds can also be attributed to input from nearby seal and horse carcasses, as well as habitual roosting by seabirds such as gulls and terns. The water quality of PD03 also reflects the prevalence of eutrophic conditions defined by high nutrients and turbidity. Recession of surrounding vegetation in both ponds can also be caused by over-grazing and trampling of surrounding areas.
Vulnerability and implications for management
The concept of vulnerability is defined by a complex set of factors, some of which include risk, exposure and ecosystem sensitivity.1,2 Based on the chironomid-inferred habitat shifts over time, we can see that both PD67 and PD03 have experienced long-term changes that are important for the context of management and conservation. Shifts in the chironomid assemblages of both systems indicate changes in habitat conditions that represent exposure and sensitivity to stress, indicating inherent ecosystem vulnerability. Furthermore, the changes that have occurred in PD67 and PD03 have caused convergence towards analogous habitats in recent decades as a result of morphological changes, making these ecosystems more ecologically similar now than in the past. In contrast, PD37 and PD38 were similar in their chironomids assemblages throughout the sediment record. The only differences observed in either system was an increase in Chironomus plumosus-type in recent sediments, suggesting that these ponds are also experiencing elevated nutrient enrichment and potential periods of eutrophication. 42
Paleolimnology can fill long-term monitoring gaps and expand the available timeframe of the system, establishing the bases for monitoring, whereby the current conditions can be compared with historic patterns. 53 The monitoring of current conditions is necessary to quantify short-term, present-day changes, and the long-term outlook offered by paleolimnology puts these changes into perspective. The paleo record from PD03 and PD67 show a cascading effect, whereby terrestrial vegetation has decreased, erosional processes have increased, and water quality has diminished. Benthic community compositions and water quality should continue to be monitored for analyzing the present conditions and trajectory of future changes. Investigation into the efficacy of physical ecosystem enclosures is currently in the beginning stages by Parks Canada and the Sable Island Institute, which may help to minimize and understand the direct and indirect influence of horses on different ecosystems, including the freshwater ponds. The short-term nature of pond enclosures combined with the long-term paleolimnological perspective will provide a complementary quantification of changes and better-defined previous conditions, which will continue to improve understanding of ecological stressors on the ponds and enable conservation of ecological integrity. Furthermore, previous attempts of terrain management have included dune restoration to protect ponds from storm surge, although this technique was not successful at all locations. 49 While it is evident in the core and water quality data that PD03 remains a freshwater system after the sand and possible saltwater intrusion, the habitat conditions that were present prior to the dune breaching have not been restored, as indicated by the altered chironomid assemblages present.
The paleolimnological perspective is essential for defining these changes, as the current conditions of PD67 and PD03 do not reflect the past. PD67 likely has higher nutrient enrichment now than in the past, and PD03 was not always as sandy as it is now. Quantifying past conditions and change over time informs management that these ecosystems were different, which is an important implication to consider when developing management objectives. For example, we found that the chironomids assemblages for both PD37 and PD38 are similar today as they were in the past; primarily reflecting productive conditions in a vegetated catchment comprised of heath. It is important to note that management of natural ecosystems can be based on unpredictable circumstances through time. Paleolimnology can establish reference conditions and evidence-based goals for restoration 6 ; yet, the ecosystem may never be able to return to the previous state because the parameters to support those conditions may no longer exist. The existence of horses on the island goes back to the eighteenth century, further than our paleo record was able to collect. While impacts from horses are likely caused by the expansion and persistence of the population, the pre-disturbance conditions remain unknown. If the horse population itself was to be managed, the changes done to the landscape may still be difficult to reverse, such as return of the formerly well-established heath around PD67 and PD03. Even if the horse impact was minimized through complete fencing of the ponds, the pre-disturbance reference conditions for ecosystem recovery do not currently exist.
The dynamic nature of Sable Island makes it impossible to rule-out other factors contributing to ecosystem change, and the continuous shifting morphology makes it more unlikely that returning to previous conditions is a possibility. Ongoing sea-level rise threatens the island’s subaerial sediments with possibility of eventual submergence. 18 With the island’s longevity in question, management plans must consider accelerated morphological change. Therefore, reference conditions established for ponds should be considered “benchmarks”, or a standard for comparison, rather than fixed targets to aim for when working towards ecosystem recovery where complete restoration is difficult to obtain. 54 When multiple stressors are heavily impacting an ecosystem, it is difficult to isolate and manage overlapping factors. 55 The combined stressors impacting the changing Sable Island pond ecosystems results in increased complexity for management, which needs to be considered in decision-making processes. Although SINPR’s pond systems have been significantly impacted, informed management of stressors could reduce continued ecological change and result in stabilization over time.
Conclusion
Based on the chironomid-inferred ecosystem changes observed in the sediment records of four ponds of SINPR, we can infer that shifts in habitat because of nutrient enrichment and geomorphologic change are some of the most prominent stressors on the ponds. Both PD67 and PD03 exhibit a change in their chironomid community assemblages that convey shifts in habitat; the decrease in Microtendipes and increase in Cricotopus in both ponds indicates loss of previously well-established heath. PD67 also exhibits impacts of erosion and increased submerged vegetation, while PD03 exhibits a shift after prominent inflow of sand. We found only a moderate influence of nutrient enrichment on either PD37 or PD38. The list of stressors is not definitive, yet these are key impacts to consider for management based on long-term ecosystem change seen in the paleo record.
As a national park, management decisions are called to prioritize maintaining and restoring the ecological integrity of SINPR. Historical data provided by the paleo record provides context for management and conservation decisions as well as the means to create reference benchmarks. These findings show that while paleolimnological analysis is predominantly utilized to analyze changes in lakes, it is also effective for determining long-term changes in small dynamic ponds and has potential for application in other similar ecosystems. For continued understanding of the vulnerability of SINPR, future work should include further analysis of chironomids in the adult stage to increase taxonomic resolution, as well as further assessment of ecological indicators such as diatoms to broaden the scope of defined ecosystem conditions. Continued monitoring of the ponds is essential for tracking ecosystem changes.
Supplemental Material
sj-docx-1-sci-10.1177_00368504221126865 - Supplemental material for A paleolimnological context of ecological vulnerability for the freshwater ecosystems of Sable Island National Park Reserve, Canada
Supplemental material, sj-docx-1-sci-10.1177_00368504221126865 for A paleolimnological context of ecological vulnerability for the freshwater ecosystems of Sable Island National Park Reserve, Canada by Victoria Theresa Watson, Dan Kehler and Andrew Scott Medeiros in Science Progress
Footnotes
Acknowledgements
Nova Scotia and Sable Island National Park Reserve are located in Mi’kma’ki, the unceded and ancestral territory of the Mi’kmaq People; we are grateful for the opportunity to research on this land. Logistical and field sampling support was provided by Parks Canada. We thank Frederica Jacks and Michael Bakaic for field sampling assistance, and Connor Nishikawa, Kathleen Hipwell, and Jennifer Eamer for laboratory assistance. We thank Jordan Eamer for discussions on Sable Island’s changing morphology. Logan Horrocks assisted with creation of the map used in ![]() . Samples were collected under a Parks Canada research permit: SINP-2019-32317.
. Samples were collected under a Parks Canada research permit: SINP-2019-32317.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Marine Environmental Observation Prediction and Response Network, the City of Halifax, and Dalhousie University (Fathom Fund, projectDAL)
Data availability
Raw abundance values of taxonomic data associated with this manuscript are archived; https://doi.org/10.5281/zenodo.7034792
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.