
Abstract
Exposure to ethanol in utero can result in Fetal Alcohol Spectrum Disorders, which may cause long-lasting cognitive and behavioral abnormalities. Preclinical studies indicate that choline ameliorates the behavioral effects of developmental alcohol exposure in rodents, and clinical studies on the effectiveness of choline, administered early in pregnancy, showed that the adverse effects of heavy prenatal alcohol exposure on postnatal growth, and cognition in human infants were mitigated. However, little is known on the mechanisms behind the effects of choline. We have previously reported that astrocyte pre-treatment with 75 mM ethanol, in vitro, reduces neurite outgrowth in hippocampal neurons co-cultured with the pre-treated astrocytes. Our in vitro system allows us to study the effects of chemicals on astrocyte functions, able to modulate neuronal development. The main objective was to test the hypothesis that choline can ameliorate the astrocyte-mediated effects of ethanol on neurite growth. In this study, we exposed primary rat cortical astrocytes to ethanol, choline, ethanol plus choline, or control conditions for 24 h. Culture media was then removed, replaced with fresh media containing no ethanol or choline treatments and primary rat hippocampal neurons were plated on top of the astrocyte monolayer and cultured for 16 h. Neurons were then stained for β-III Tubulin and neurite outgrowth was measured. Astrocyte exposure to ethanol (25, 50, and 75 mM) decreases neurite outgrowth in co-cultured hippocampal pyramidal neurons, while astrocyte treatment with choline had no effect. Astrocyte treatment with ethanol and choline in combination, however, prevented the effect of ethanol, leading to levels of neurite outgrowth similar the control condition. Choline prevents the inhibitory effect of ethanol-treated astrocytes on neurite outgrowth while not altering normal neuronal development. These results suggest a new, astrocyte-mediated mechanism by which choline ameliorates the effects of developmental alcohol exposure.
Introduction
In utero alcohol exposure can induce Fetal Alcohol Spectrum Disorders (FASD), which have been increasingly recognized as a major concern in the US as well as world-wide. 1 In the United States, FASD prevalence can be as high as 5% of the population, 2 and is even higher in some communities across the word.1,3 The most severe and long-lasting consequences of in utero alcohol exposure include cognitive and behavioral dysfunctions that have been associated with neuroanatomical abnormalities.2,4
Several preclinical studies suggest that choline supplementation may ameliorate the cognitive and behavioral effects of developmental alcohol exposure. For instance, choline supplementation improves ethanol-induced hyperactivity, decreases the number of reversal learning errors in a serial spatial discrimination task, 5 and rescues ethanol-induced impairment of spatial memory in the Morris Water Maze. 6 However, choline does not rescue ethanol effects on motor coordination 7 in a human gestation third-trimester equivalent FASD rat model. Choline was also shown to ameliorate balance deficits caused by neonatal alcohol exposure in mice. 8 Furthermore, low dietary choline exacerbates the effects of developmental alcohol exposure in motor development tasks. 9 A comprehensive review of the ameliorating effects of choline in rodent models of FASD has been published. 10 Based on this strong preclinical evidence, clinical trials assessing the effectiveness of choline in the treatment of children with FASD have been carried out and more clinical trials are recommended before a conclusion on the effectiveness of choline treatments on FASD children can be reached. 10 Choline, administered early in pregnancy, appeared to mitigate adverse effects of heavy prenatal alcohol exposure on postnatal growth, and cognition in human infants. 11
Choline is an essential nutrient found in many foods, including liver, eggs, and wheat germ. 12 Like ethanol, it can pass through both the placenta and the blood-brain barrier, and therefore has the ability to affect the development of the fetal brain. 13 There is an increased demand for choline during pregnancy, and it is essential for healthy fetal development. 14 Choline supplementation decreases ethanol induced hypermethylation in the prefrontal cortex, 15 and stabilizes ethanol induced micro-RNA variability in the hippocampus. 16 Gestational choline supplementation normalizes the effects of ethanol on DNA methylation, histone acetylation, the expression of the pro-opiomelanocortin gene in hypothalamic neurons, 17 and it also prevents the effects of ethanol on neurite outgrowth, cell adhesion molecule L1 signaling, and L1 distribution in lipid rafts in cerebellar neurons in culture. 18 Finally, choline supplementation normalizes the upregulation of M2/4 muscarinic receptors induced by ethanol in the dorsal hippocampus. 19
Astrocytes play essential roles in all aspect of brain development, as they contribute to the development of the blood-brain barrier and are highly involved in synapse formation, functional maturation, and pruning.20–22 Our previous research has indicated that astrocytes also play an important role in neurite outgrowth and mediate the effects of ethanol on neuronal development.23–25 Specifically, we have reported that when neurons are cultured in vitro with astrocytes treated with ethanol, they display a delayed maturation measured as decreased neurite outgrowth. 25
In this study we investigated the hypothesis that choline treatments may prevent the inhibition of pyramidal neuron neurite outgrowth induced by ethanol-treated astrocytes. In order to investigate the effect of the treatments on hippocampal neuron development, we treated primary cultures of astrocytes with ethanol and/or choline for 24 h, using a method previously described by us. 25 We report that, when neurons were cultured with ethanol-treated astrocytes, they displayed reduced neurite outgrowth in all ethanol concentrations tested. Neurons cultured with astrocytes treated with choline or with choline and ethanol together were not significantly different from neurons incubated with control astrocytes. These results suggest choline prevents the effects of ethanol on neuronal development via an astrocyte specific mechanism.
Methods
Animals
Timed-pregnant Gestational Day (GD) 15 Sprague-Dawley rats were purchased from Charles River (Wilmington, MA). Upon arrival, animals were maintained at the Portland VAMC Veterinary Medical Unit, under a 12 h light/dark cycle at 22°C ± 1°C. The animals were allowed a week to acclimate following travel before tissue collection. Animals had ad libitum access to water and food (chow diet). All animal procedures were approved by the Portland VA Health Care System Institutional Animal Care and Use Committee, and followed US National Institutes of Health animal welfare guidelines.
Cortical astrocyte primary cultures
Cortical astrocyte cultures were prepared from GD21 fetuses, as previously described.25,26 Fetuses were sexed, and female and male astrocytes were plated in separate flasks. Astrocytes were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) containing 10% Fetal Bovine Serum (FBS), 100 units/mL penicillin, and 100 µg/mL streptomycin (all from Gibco, Grand Island, NY), in a humidified incubator at 37°C under a 5% CO2/95% air atmosphere for 10–16 days. Two flasks (one with female-only astrocytes and one with male-only astrocytes) were then trypsinized; cell suspensions derived from female and male astrocyte flasks were combined and plated into 24-well plates containing circular glass coverslips (1 × 105 cells/well). Cultures prepared in this way are balanced, mixed-sex cultures. Astrocytes were cultured for three additional days and then switched to serum-free DMEM supplemented with 0.1% Bovine Serum Albumin (BSA, Sigma Aldrich, St. Louis, MO) and Penicillin/Streptomycin for 24 h, followed by ethanol, choline, ethanol plus choline, or control treatments, prepared in DMEM/0.1% BSA medium, for 24 h. The medium containing treatments was then removed from astrocyte cultures and replaced with treatment-free DMEM/0.1% BSA.
Cortical astrocyte-hippocampal neuronal co-cultures
Hippocampal neurons were prepared from GD21 fetuses as previously described.23–25 The same number of female and male pups were used in these cultures. Neurons were plated at a concentration of 1 × 104 neurons/coverslip on top of the astrocyte monolayer for 16 h, starting 2 h after the removal of treatments from the astrocyte cultures.
Ethanol and choline treatments
Astrocytes plated on glass coverslips placed in 24-well plates were incubated with 1 mL DMEM/0.1% BSA medium containing 25, 50, or 75 mM ethanol or control (ethanol-free) medium for 24 h. Ethanol treatments took place in sealed chambers with a dish of water containing the same alcohol concentration present in the cultures; each chamber contained astrocytes treated with a single ethanol concentration. 25 A gas mixture of 5% CO2/95% air was run through these chambers, after which the chambers were sealed and incubated at 37°C for 24 h. Choline treatments were carried out by incubating astrocyte cultures with 1 mL DMEM/0.1% BSA medium containing 0 (control), 10, 50, or 100 mM choline chloride (Sigma Aldrich, St. Louis, MO) for 24 h. To test the effects of both ethanol and choline on neurite outgrowth, astrocytes were incubated with 75 mM ethanol, 100 mM choline, 75 mM ethanol + 100 mM choline, or control (treatment-free) DMEM/0.1 BSA medium for 24 h.
Ethanol concentration determination
Ethanol concentrations in the medium were determined at the beginning and the end of each treatment by head-space gas chromatography after mixing 20 µL of medium with 500 µL of a solution of 4 mM propanol in water, as previously described. 27 There was no significant evaporation of ethanol during the exposure period.
Osmolality
Changes in osmolality have been shown to affect neurite outgrowth. 28 For this reason, we determined the osmolality of control medium, medium containing 100 mM choline at the beginning and at the end of the 24 h incubations, and in medium incubated on astrocytes for 2 h, corresponding to the medium that neurons experience when co-incubated with astrocytes. Osmolality was measured using a Vapro Pressure Osmometer 5520 (Wescor Inc., Logan, UT). For each time-point samples from three independent replicates were tested; each sample was run three times and results are expressed as mmol/kg.
Cell viability
Astrocytes exposed to choline or ethanol were tested for cell viability using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetraxzolium bromide (MTT) assay (Sigma Aldrich, St. Louis, MO), as previously described.29,30 Cells were incubated with 500 µL DMEM/0.1 BSA containing 0.5 mg/mL MTT for 2 h. The MTT-containing medium was then removed from the astrocyte monolayer, 500 µL of DMSO was added, and the resulting solution was read on a spectrophotometer at 570 nm; data are expressed as percent of control.
Immunocytochemistry
Following astrocyte-neuron co-culture, cells were fixed with 4% paraformaldehyde in phosphate buffered saline, and immunostained with a β-III Tubulin antibody (1:150 dilution; catalog #: MAB1637, Millipore, Burlington, MA) followed by the Alexa Fluor 488 secondary antibody (1:300 dilution; catalog #: A11001, Thermo-Fisher Scientific, Waltham, MA). Coverslips were then mounted on microscope slides (25. Zhang et al., 2014).
Microscopy
Neurons were imaged on a Leica DM500b microscope with a DFC36 FX camera, using the program Neurolucida (Version 11, MBF Bioscience, Williston, VT, USA). Neurons were selected for morphometric analysis by a researcher blind to the experimental treatments and traced from pictures taken with a 40× objective. Only cells that (1) had three or more neurites all longer than the cell body, (2) did not overlap with other neurons, (3) were fully on top of the underlying astrocytes, (4) were pyramidal neurons, and (5) were intact were selected for analysis. Neurites were traced using the software Neurolucida, and analyzed by Neurolucida Explorer. For each cell, three measurements were taken: the length of the longest neurite, the average length of the minor neurites, and the number of neurites.
Statistics
Data were analyzed using a multi-level analysis in the statistical program R 31 with the package lme4. 32 Outliers were identified using a value equal to 1.5 the Inter-Quartile Range. We conducted nested multilevel analyses to account for multiple cells measured in the same coverslip and multiple coverslips within the same experiment 33 (See Table 1 for a summary of the sample sizes).
Neuron morphometric analysis experimental sample size.
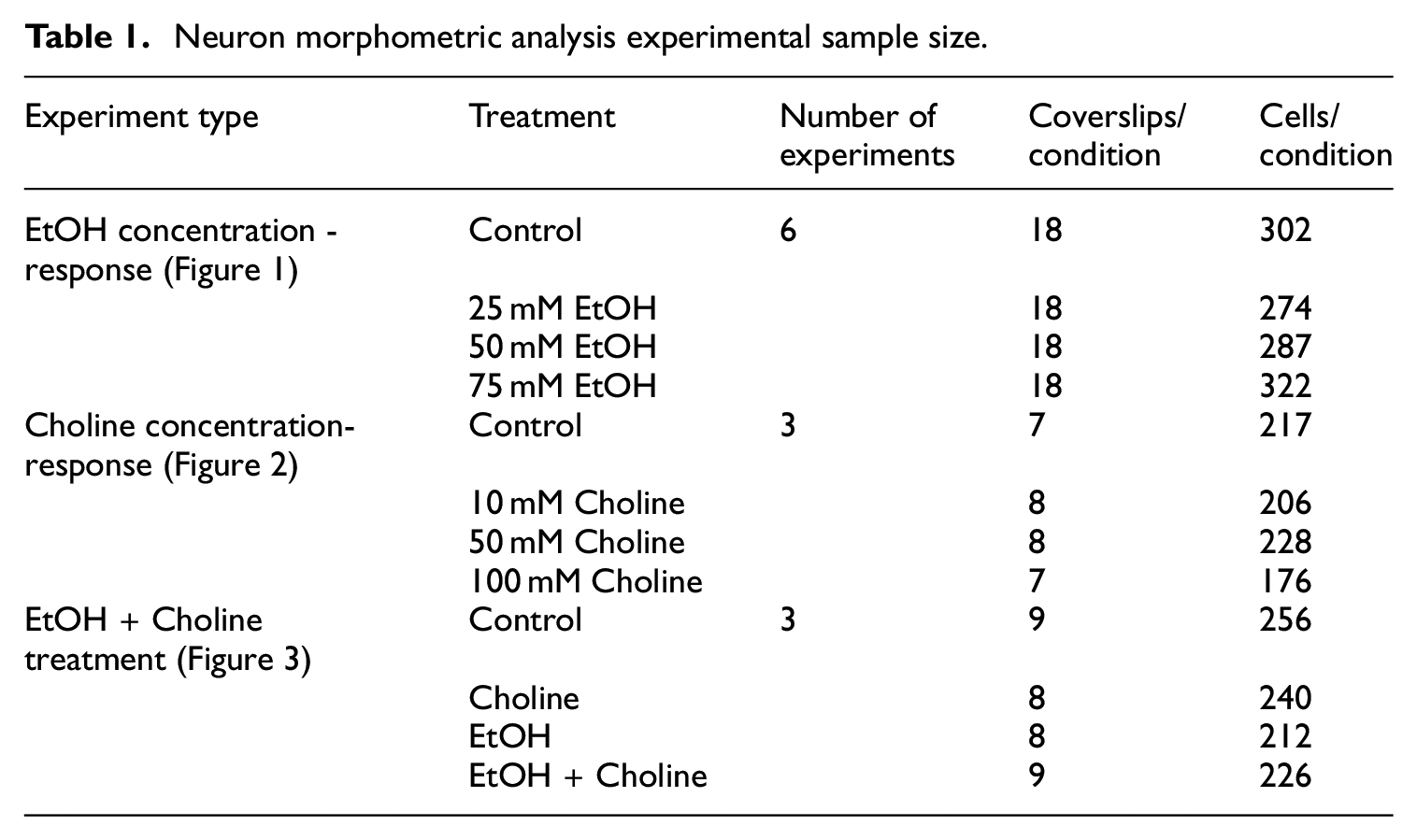
Benjamini-Hochberg tests were run to correct for multiple comparisons and control for the false discovery rate (FDR). Tukey’s post-hoc test was run when appropriate. A post-hoc p-value of <0.05 and a false discovery rate q-value of <0.05 were considered significant. Data are reported as mean ± S.E.M.
Results
Ethanol exposure decreased neurite outgrowth
Astrocyte cultures were exposed to 25, 50, or 75 mM ethanol or control conditions for 24 h. The medium containing ethanol was then removed and replaced by fresh, medium with no ethanol. After 2 h in treatment-free medium, hippocampal pyramidal neurons were cultured on top of the astrocytes for 16 h, and processed for staining with neuronal specific β-III tubulin antibody, and measured using fluorescence microscopy. A multilevel analysis with corrections for multiple comparisons showed an effect of ethanol exposure on the longest neurite length (q < 0.0001, Table 2).
EtOH concentration-response multilevel analyses and FDRs.

Tukey’s post-hoc analysis showed that ethanol exposure decreased the longest neurite length at all tested ethanol concentrations (Control vs 25 mM p = 0.003, Control vs 50 mM p < 0.001, Control vs 75 mM p = 0.001; Figure 1(a)). Furthermore, there was an effect of ethanol on average minor neurite length in the nominal p-value, but this effect was not significant following correction for multiple comparisons (q = 0.063, Figure 1(b), Table 2). Ethanol-treated astrocytes did not affect the number of neurites (Figure 1(c), Table 2).

Effect of ethanol-treated astrocytes on neurite outgrowth of pyramidal neurons. Astrocytes plated on glass coverslips were treated for 24 h in the presence or absence of ethanol (25, 50, or 75 mM); treatments were then washed out and replaced with fresh medium. Two hours later, freshly isolated hippocampal neurons were plated on top of the astrocytes for an additional 16 h, at the end of which neurons were fixed, labeled with a β-III tubulin antibody followed by a fluorescent secondary antibody, and morphometrically analyzed using the software Neurolucida: (a) length of the longest neurite, (b) average minor neurite length, and (c) number of neurites/neuron. All data were analyzed using a multilevel analysis with Benjamini-Hochberg to correct for multiple comparisons followed by Tukey’s post-hoc test.
Choline exposure did not change neurite outgrowth
Astrocytes were exposed to 0 (control), 10, 50, or 100 mM choline, and neurons were cultured on the treated astrocytes as described above. Choline did not affect the length of the longest or the minor neurites, nor the number of neurites, regardless of choline concentration (Figure 2(a)–(c), Table 3).

Effect of choline-treated astrocytes on neurite outgrowth of pyramidal neurons. Astrocytes plated on glass coverslips were treated for 24 h in the presence or absence of choline (10, 50, or 100 mM); treatments were then washed out and replaced with fresh medium. Two hours later, freshly isolated hippocampal neurons were plated on top of the astrocytes for an additional 16 h, at the end of which neurons were fixed, labeled with a β-III tubulin antibody followed by a fluorescent secondary antibody, and morphometrically analyzed using the software Neurolucida: (a) length of the longest neurite, (b) average minor neurite length, and (c) number of neurites/neuron. All data were analyzed using a multilevel analysis with Benjamini-Hochberg to correct for multiple comparisons.
Choline concentration-response multilevel analyses and FDRs.

Choline prevents the effect of ethanol on neurite outgrowth
Because we saw no effect of any choline concentration on neurite outgrowth (Figure 2), we chose to use the highest concentration of choline (100 mM) in the ethanol and choline co-treatment experiments. Similarly, because we saw a similar response to all concentrations of ethanol tested (Figure 1) on neurite outgrowth and because our previous work showing the effects of ethanol treated astrocytes reducing neurite outgrowth used 75 mM ethanol, 25 we used the 75 mM ethanol to determine if the presence of choline during ethanol treatment would reverse or reduce the effects of ethanol on neurite outgrowth. Astrocytes were exposed to 100 mM choline, 75 mM ethanol, 100 mM choline + 75 mM ethanol, or control conditions for 24 h, after which the treatments were removed. Two hours after removal of treatments, hippocampal neurons were cultured on top of the astrocytes, and neurite outgrowth was measured following 16 h of growth. We found a significant interaction between ethanol and choline (q < 0.001) and a main effect of ethanol on the longest neurite length (q = 0.01), but no effect of choline (Table 4).
EtOH and choline treatment multilevel analyses and FDRs.

Ethanol and choline together resulted in longer neurite lengths than the ethanol treatment (p < 0.0001) and the choline treatment (p < 0.05) by the Tukey’s post-hoc test (Figure 3(a)). Ethanol exposure also trended toward a reduction in neurite length when compared to controls (p = 0.056). There was no effect of choline or ethanol on the average length of the minor neurites (Figure 3(b)).

Effect of choline-plus-ethanol-treated astrocytes on neurite outgrowth of pyramidal neurons. Astrocytes plated on glass coverslips were treated for 24 h in the presence or absence of choline (100 mM), ethanol (75 mM), or ethanol and choline together; treatments were then washed out and replaced with fresh medium. Two hours later, freshly isolated hippocampal neurons were plated on top of the astrocytes for an additional 16 h, at the end of which neurons were fixed, labeled with a β-III tubulin antibody followed by a fluorescent secondary antibody, and morphometrically analyzed using the software Neurolucida: (a) length of the longest neurite, (b) average minor neurite length, and (c) number of neurites/neuron. All data were analyzed using a multilevel analysis with Benjamini-Hochberg to correct for multiple comparisons followed by Tukey’s post-hoc test.
We also found a main effect of ethanol on the number of neurites (q = 0.01), but no main effect of choline or interaction (Figure 3(c); Table 4).
Tukey’s post-hoc analysis showed that neurons cultured with astrocytes treated with ethanol and choline in combination had significantly more neurites than the ethanol only treatment (p = 0.028). Representative images of neurons in all four conditions can be seen in Figure 4.

Pyramidal neurons immunocytochemically stained with β-III tubulin antibody. Representative hippocampal neurons co-cultured with control (a), 75 mM ethanol-treated (b), 100 mM choline-treated (c) and 75 mM ethanol + 100 mM choline-treated (d) astrocytes.
Changes in neurite outgrowth following choline exposure were not due to changes in osmolality
Astrocytes were exposed to 100 mM choline for a period of 24 h. Samples of media were taken at the beginning (t0) and end (t24) of the incubation. At the end of the 24 h incubation with choline, the medium was replaced with treatment-free DMEM/0.1 BSA and sampled following 2 h of incubation. All samples, as well as control medium were tested for osmolality (Figure 5).
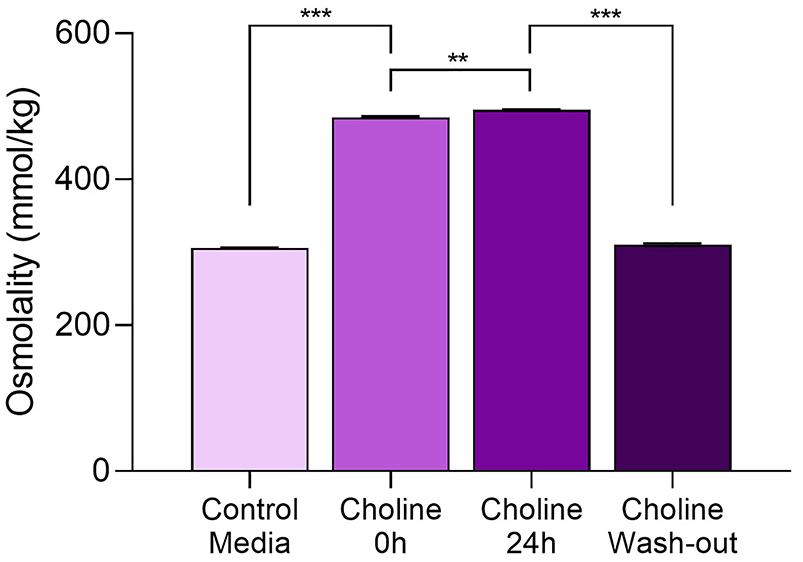
Effect of choline on osmolality. Osmolality was evaluated in treatment-free DMEM/0.1% BSA medium (control; left column), in the medium containing 100 mM choline chloride before (t0, second column from the left) and after (t24, third column from the left) 24 h incubation on astrocytes. In some of the wells containing choline-treated astrocytes, the medium containing choline was removed and fresh DMEM/0.1% BSA was added to the cultures for 2 h before being evaluated for osmolality (column on the right). Data were analyzed using two-way ANOVA, followed by Tukey’s post-hoc test.
The purpose of this experiment was to determine if there was a significant difference in osmolality in the cellular environment at the time when neurons were added, as changes in osmolality could be responsible for changes in neurite outgrowth. 28 As expected, 100 mM choline chloride significantly increased medium osmolality compared to control medium both before (p < 0.0001) and after (p < 0.0001) the 24 h incubation by Tukey’s post-hoc (ANOVA: F(2,6) = 5730.882, p < 0.0001). Interestingly, there was a small but significant increase in the osmolality of the choline-containing medium incubated with astrocytes for 24 h compared to the medium before incubation (see the two middle columns in Figure 5; p < 0.01). We hypothesize that this increase is due to the factors that are released by astrocytes during the 24 h incubation. The osmolality of the medium incubated with astrocytes for 2 h following the 24 h incubation with choline was not different from the osmolality of control medium, indicating that the effect of choline-treated astrocytes on neurite outgrowth is not due to changes in osmolality (Figure 5, right column).
Changes in neurite outgrowth are not due to astrocyte cell death
To test whether ethanol and choline treatments affect astrocyte viability, astrocytes were incubated for 24 h in the presence or absence of 25, 50, 75 mM ethanol (Figure 6(a)) or 10, 50, 100 mM choline chloride (Figure 6(b)). Following this, the treatment-containing medium was removed and replaced with fresh, treatment-free medium for 16 h to mimic the timing of astrocyte-neuron co-cultures. Cell viability, assessed by the MTT assay, was not reduced by any of the ethanol (Figure 6(a)) or choline (Figure 6(b)) treatments. There was, however, a small but significant increase in cell viability induced by the highest concentration of ethanol (75 mM) (ANOVA: F(3,32) = 2.429, p = 0.01; control vs ethanol 75 mM p = 0.026 by the Dunnett’s post-hoc test. Figure 6(a)). Furthermore, we observed no alterations in astrocyte morphology or number in any of the ethanol, choline, or ethanol + choline treatments described.

Effect of ethanol and choline on astrocyte viability. Astrocytes plated in 24-well plates were treated for 24 h in the presence or absence of ethanol (25, 50, or 75 mM) (a) or choline (10, 50, or 100 mM) (b); treatments were then washed out and replaced with fresh medium for 2 h. Cell viability was then measured by the MTT test. Data were analyzed using a one-way ANOVA, followed by a Dunnett’s post-hoc test, and are expressed as percent of control.
Discussion
Cognitive and behavioral consequences of developmental alcohol exposure may, at least in part, be due to alterations in neuronal plasticity and connectivity as evidenced by rodent preclinical studies and human imaging studies.34–36 Our laboratory has been investigating the hypothesis that some of the effects of ethanol on neuronal plasticity may be mediated by astrocytes.4,37 In culture, neurons develop faster and reach full morphological and functional maturation only when they are plated in the presence of astrocytes,38–40 underscoring the role of astrocytes in neuronal development.
We optimized in vitro models to assess how astrocyte treatments with ethanol affect neuronal development.24,25 We previously reported that neurons co-cultured on top of astrocytes pre-treated with 75 mM ethanol displayed reduced neurite outgrowth in comparison to neurons co-cultured with control astrocytes. 25 In the present study we expand on this observation to report that neurons cultured with ethanol-treated astrocytes had significantly reduced longest neurite length (which is the one that will likely develop into the axon24,38) starting at the lowest ethanol concentration used, 25 mM (Figure 1, Table 2). The ethanol concentrations we used in this study, 25, 50, and 75 mM (corresponding to 0.115, 0.23, and 0.35 g/dL, respectively) are clinically relevant, as they can be found in the blood of individuals after high ethanol intake 41 and are within the range of concentrations recommended for in vitro studies. 42 Furthermore, we verified that these ethanol concentrations did not reduce astrocyte viability (Figure 6(a)).
Ethanol pre-treatment of astrocytes in an astrocyte-neuron co-culture model has been shown to reduce the length of longest neurite, but has shown inconsistent results on the average minor neurite length with one report showing no changes, 24 and one report showing a reduction. 25 Here we report effects of ethanol, with or without choline, only altering the longest neurite length (Figures 1 and 3), which may reflect specific effects on the developing axon or may be a result of differences being easier to observe in neurites that are longer or growing at a faster rate. Our in vitro observation of altered astrocyte-specific ethanol effects on neurite outgrowth, determined by pre-treating astrocytes and removing any treatments prior to the plating of neurons on top of the astrocyte monolayer, makes interpretation of the results in the larger context of brain development complex. For example, we have observed increased pyramidal neuron complexity in a rat neonatal intragastric gavage FASD model, 43 and others have observed gestational ethanol consumption of ethanol in a mouse model caused increased length of cortical neuron projections at postnatal day 0 that was reduced by the co-administration of choline with ethanol in the diet. 44 Besides the obvious differences between in vitro and in vivo models, the results presented here are distinct in the measurement of astrocyte-specific effects of ethanol with or without choline supplementation. In vivo models have the drawback of both neurons and astrocytes (as well as microglia, oligodendrocytes, and other cell types) being present and responding to the treatments in a complex manner. Our findings of astrocyte-specific effects sheds light on how astrocytes respond to ethanol and/or choline and the effects of these responses on neuronal development. Candidates for mediating the altered neuronal morphology elicited by ethanol pre-treatment of astrocyte are abundant as astrocytes produce many factors that facilitate proper neuronal development 45 but a potential mechanism is via tissue-type plasminogen activator (tPA; Plat) which we have previously shown is up-regulated by ethanol in astrocyte cultures and in the whole rat hippocampus. 46
Choline supplementation has been shown to ameliorate behavioral abnormalities in rodents developmentally exposed to ethanol and is currently being investigated for its effectiveness in children with FASD.5–8,10 An important goal of this study was to test whether choline supplementation in astrocytes ameliorates the effects of alcohol on neurite outgrowth. We report that, while morphometric parameters of neurons cultured with choline-treated astrocytes were not statistically different from the ones of neurons co-cultured with control astrocytes (Figure 2, Table 3), data from neurons co-cultured with astrocytes exposed to both ethanol and choline together showed a protective effect of choline on the reduced length of the longest neurite caused by ethanol (Figure 3, Table 4). It should be noted that the culture medium used does contain choline chloride at a concentration of 28.6 µM which is sufficient for normal astrocyte growth and astrocyte-neuron co-cultures and may explain why choline treatment alone has no observed effects on neurite outgrowth. However, the choline in the culture medium is no longer sufficient for normal astrocyte functions including supporting neurite outgrowth in co-cultured neurons when the astrocytes are pre-treated with ethanol.
Because in this study we used high concentrations of choline, which are, however, in the range of concentrations previously used in in vitro experiments,47,48 we verified that they do not affect astrocyte viability (Figure 6(a)). We also verified that choline treatments in astrocytes do not change the osmolality of the medium during neuron co-culture (Figure 4, compare the first column to the left and the first column to the right); this is an important consideration given the fact that changes in osmolality of the medium in which neurons are grown can affect neurite outgrowth. 28 Together these data indicate that choline counters the changes induced by ethanol in astrocytes, which are therefore able again to foster neuronal development similarly to control astrocytes.
Choline is an essential nutrient and a precursor of several molecules involved in important cellular functions. Among the most important choline metabolites are betaine, which can be converted to methionine and, subsequently, to S-adenosylmethionine, the universal methyl donor used by DNA methyl transferases in DNA methylation reactions, and the neurotransmitter acetylcholine.49,50 However, because the conversion of choline to betaine and then to methionine is catalyzed by enzymes present only in the liver and kidneys and the conversion of choline to acetylcholine by choline acyltransferase occurs only in cholinergic neurons, 49 it is reasonable to assume that these two metabolites do not play a role in the observed effect mediated by astrocytes. Another major metabolite of choline is phosphatidylcholine (PC), the major membrane phospholipid which accounts for about 95% of choline in most of the tissues; the enzyme necessary for the biosynthesis of PC is present in all nucleated cells. 49 PC also plays an important role in signal transduction as the hydrolysis of PC by the enzyme phospholipase D (PLD) generates phosphatidic acid (PA), a second messenger that can induce neurite outgrowth. 51 Interestingly, it is well established that ethanol alters PA signaling. Indeed, ethanol competes with water as a substrate for PLD whereby PA production is inhibited and phosphatidylethanol is formed instead.52,53 This mechanism may be involved in the protective effect of choline on the inhibition of neurite outgrowth mediate by ethanol-treated astrocytes. The reason why choline supplementation does not, on its own, have an effect on neuronal development may be that cellular homeostatic processes regulate the incorporation of choline into PC or other choline metabolites only when needed by the cell, as, for instance, when in the presence of ethanol.
In conclusion, we show that choline can prevent the effects of ethanol-treated astrocytes on neurite outgrowth. This study is the first to suggest that the beneficial effects of choline may be attributed to an effect not on neurons, but on astrocytes. Astrocyte involvement in brain development is now well established; 21 our research has been aimed at investigating the effects of ethanol on astrocyte functions that can then result in altered neuronal development. It is therefore not surprising that astrocytes can also be the target for possible interventions for the treatment of FASD, such as choline. Future studies will be aimed at characterizing the mechanisms by which choline ameliorates the effects of ethanol-exposed astrocytes on neurite outgrowth.
Footnotes
Acknowledgements
We thank Mr. Adam Munhall for assisting with osmolality measurements, Dr. Deborah Finn and Mr. Jeremiah Jensen for measuring medium alcohol concentrations, and Ms. Shelley Bloom for her assistance with astrocyte cultures. The opinions expressed in this paper are of the authors only, and the contents do not represent the view of any other person or entity, including the United States Department of Veterans Affairs, or the United States Government.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the NIH/NIAAA F31AA024950-01A1 to CG, and by NIH/NIAAA R01AA022948 and VA Merit Review Award I01BX001819 to MG, and by facilities and resources at the Portland VA Health Care System.