
Abstract
Can farmers in sub-Saharan Africa (SSA) boost crop yields and improve food availability without using more mineral fertilizer? This question has been at the center of lively debates among the civil society, policy-makers, and in academic editorials. Proponents of the “yes” answer have put forward the “input reduction” principle of agroecology, i.e. by relying on agrobiodiversity, recycling and better efficiency, agroecological practices such as the use of legumes and manure can increase crop productivity without the need for more mineral fertilizer. We reviewed decades of scientific literature on nutrient balances in SSA, biological nitrogen fixation of tropical legumes, manure production and use in smallholder farming systems, and the environmental impact of mineral fertilizer. Our analyses show that more mineral fertilizer is needed in SSA for five reasons: (i) the starting point in SSA is that agricultural production is “agroecological” by default, that is, very low mineral fertilizer use, widespread mixed crop-livestock systems and large crop diversity including legumes, but leading to poor soil fertility as a result of widespread soil nutrient mining, (ii) the nitrogen needs of crops cannot be adequately met solely through biological nitrogen fixation by legumes and recycling of animal manure, (iii) other nutrients like phosphorus and potassium need to be replaced continuously, (iv) mineral fertilizers, if used appropriately, cause little harm to the environment, and (v) reducing the use of mineral fertilizers would hamper productivity gains and contribute indirectly to agricultural expansion and to deforestation. Yet, the agroecological principles directly related to soil fertility—recycling, efficiency, diversity—remain key in improving soil health and nutrient-use efficiency, and are critical to sustaining crop productivity in the long run. We argue for a nuanced position that acknowledges the critical need for more mineral fertilizers in SSA, in combination with the use of agroecological practices and adequate policy support.
Introduction
Agriculture is at the cross-road of sustainability issues related to food security, biodiversity loss, environmental degradation and climate change. Alternative approaches to “industrial” or “conventional” agriculture (Sumberg and Giller, 2022), such as regenerative agriculture and agroecology are gaining momentum, as noticeable from the rising number of scientific papers dealing with these topics (Giller et al., 2021b; Wezel and Jauneau, 2011). Success stories of farmers in the tropics who shift away from mineral fertilizers, pesticides, and other chemical inputs also abound on social media and communication materials of nongovernmental organizations. These are increasingly being populated with testimonies of farmers diversifying their cropping systems with cover crops, investing in livestock keeping, producing high-quality compost, and improving their cropland with the use of legumes, often with claims of great positive impact on income, food security, and soil fertility maintenance.
These promises of diversified and “fully-integrated” farming systems where the need for external inputs to sustain soil fertility and crop productivity is reduced or eliminated, along with abundant scientific literature on the farm-level impact of resource recycling and improved efficiency, have certainly inspired the “input reduction” principle of agroecology: “Reduce or eliminate dependency on purchased inputs and substitute conventional inputs that have negative impacts on the environment, replacing them by making use of agroecological alternatives” (Wezel et al., 2020).
Agroecology can be defined in numerous ways. A broad definition is “the science of the relationships of organisms in an environment purposely transformed by man for crop or livestock production” (Martin and Sauerborn, 2013). More recently, agroecology has been defined as the application of ecological concepts and principles to optimize interactions between plants, animals, humans and the environment while taking into consideration the social aspects that need to be addressed for a sustainable and fair food system (Wezel et al., 2020). A key focus has been increased reliance on knowledge and management of ecological processes, complementing and reducing the use of external inputs, like mineral fertilizers and pesticides. Agroecology revolves around a set of 10 elements (FAO, 2018) or 13 principles (HLPE, 2021), among which the recycling, input reduction, soil health, biodiversity and synergy principles are directly relevant to soil fertility maintenance. 1 These principles can be operationalized through a wide range of agroecological practices, among which are intercropping, agroforestry, rotation with legumes and crop–livestock integration. Decades of scientific work in the tropics have shown that these agroecological practices can contribute to replenishing and maintaining soil fertility, i.e., “the ability of a soil to sustain plant growth by providing essential plant nutrients and favorable chemical, physical, and biological characteristics as a habitat for plant growth” (https://www.fao.org/global-soil-partnership/areas-of-work/soil-fertility/en/).
Here, we question the “input reduction” principle of agroecology: what does it mean in resource-limited farming contexts, such as those commonly experienced by smallholder farmers throughout sub-Saharan Africa (SSA)?
The aforementioned success stories often seem based on conducive individual circumstances, for example, high remittances and above-average farm size, thus neglecting the constraints faced by the majority of smallholder farmers in SSA. Farm sizes have been shrinking in SSA, and soils are generally nutrient-depleted, due to low inherent fertility and decades of continuous cropping with low nutrient inputs, leading to low crop productivity (Sanchez, 2002). Huge productivity gains need to be achieved to match a growing food demand (van Ittersum et al., 2016). Yet, in the development community, particularly in SSA, an emerging viewpoint considers the use of mineral fertilizers as “climate stupid." 2 Advocates of this perspective argue that governments should stop subsidizing them, due to their rising costs and their negative impact on the environment, including the emission of greenhouse gases (GHGs). Instead, they advocate for the promotion of low-cost agroecological alternatives that are expected to revitalize the soil and protect the ecosystem. Within the scientific literature, there is a debate at the science-policy interface harnessing current knowledge to build views on the possible future of agricultural systems: some authors claim that there is “a growing consensus that sustainable intensification in Africa will be […] fertilizer intensive” (Jayne et al., 2019; Vanlauwe et al., 2014), while others assert, that “reducing the dependency on purchased inputs can reduce food insecurity especially for small-scale food producers” (Wezel et al., 2020) and that “the challenges farmers face can be met with appropriate management […] making the addition of external inputs largely unnecessary” (Gliessman, 2014). Côte et al. (2022) propose a reconciling view of agroecology that does not exclude “the use of […] exogenous inputs when yield gap is important,” insisting that “an in-depth agroecological transformation […] should be free of synthetic pesticides and should be parsimonious in the use of synthetic fertilizers.” Donor agencies, notably the European Union and its member countries, increasingly support development projects in SSA that engage with the agroecological transition and transition to green economies (DG DEVCO, 2020), without necessarily being clear on the role that external inputs and mineral fertilizer play.
In this article, we critically examine the input reduction claim made by agroecology in the context of smallholder farmers in SSA. Our objective is to evaluate the feasibility and desirability of this principle for SSA, specifically concerning the maintenance of soil fertility and the role of mineral fertilizer. Our analysis leads to five arguments: (i) the starting point in Africa is “agroecological” by default, meaning very low fertilizer use, that has over the years led to poor soil fertility as a result of widespread soil nutrient mining, (ii) the nitrogen (N) required by crops cannot be adequately fulfilled solely through biological N fixation from legumes or through recycling of animal manure, (iii) other nutrients like phosphorus (P) and potassium (K) need to be replaced and are also key for biological N fixation, (iv) mineral fertilizer, if used appropriately, causes little harm to the environment, and (v) reducing the use of mineral fertilizer would hamper crop productivity gains and contributes indirectly to agricultural expansion. We then discuss the role that agroecological principles can play in improving soil health and nutrient use efficiency, and in sustaining crop productivity.
Our analyses focus on rainfed annual cropping systems in SSA. This category regroups the maize crop-livestock, root and tuber crop, and cereal-root crop-livestock farming systems (Dixon et al., 2001) that predominate on the continent. Our analysis does not focus on irrigated systems (e.g., rice, vegetables) and perennial systems (e.g., coffee, cocoa, oil palm), that represent a relatively smaller share of the agricultural production area (Vanlauwe et al., 2023).
Starting point in SSA: Agroecological “by default”
Maintaining soil fertility through agroecological practices has been a common feature of mixed crop-livestock farming systems in SSA where livestock plays a great role in soil fertility maintenance. For example, in the cotton basin of southern Mali, the income earned from cotton production has been used by farmers to increase the size of their cattle herd—they can recycle amounts of manure as high as three tons per hectare per year (Falconnier et al., 2015). Farmers also rely on legumes and biological N fixation: in central Senegal, farmers have cultivated millet in rotation with groundnut for decades (Noba et al., 2014). Across the semiarid and subhumid zones of the continent, from the groundnut basin of central Senegal and the cotton basin of southern Mali, to central Ethiopia (Sida et al., 2018) and the mid-Zambezi valley in Zimbabwe (Baudron, 2011), farmers have deliberately retained native trees in their fields, creating the so-called “parklands.” Some of these trees are N-fixing trees, for example, the well-known Faidherbia albida (Delile) A. Chev. (Peltier, 1996). In the Sahel region, farmers have traditionally maintained long-term fallows that help replenish soil fertility and offer an additional source for livestock nutrition and manure production (Schlecht et al., 2004).
As a result of these long-standing agroecological practices, manure and biological N fixation have been estimated to be the largest sources of N input to cropland when considering the African continent as a whole (Billen et al., 2014). In fact, Africa is the only region of the world where mineral fertilizer is not the first source of N input to cropland (Billen et al., 2014). This conclusion also holds when looking at N inputs to cropland at the farm and village level: despite considerable variations among farms in, e.g., Kenya, Uganda, Ethiopia, and Mali, organic inputs from outside the farm/village are the primary source of N, followed by biological N fixation, atmospheric deposition and mineral fertilizer, in that order (Figure 1). The main sources of P input to cropland in Africa are manure and mineral fertilizer, which contribute approximately the same amount, around 2.5 kg P/ha/year. This is in contrast to the other regions of the world where the main P source is mineral fertilizer (>10 kg P/ha/year) (Ringeval et al., 2017).
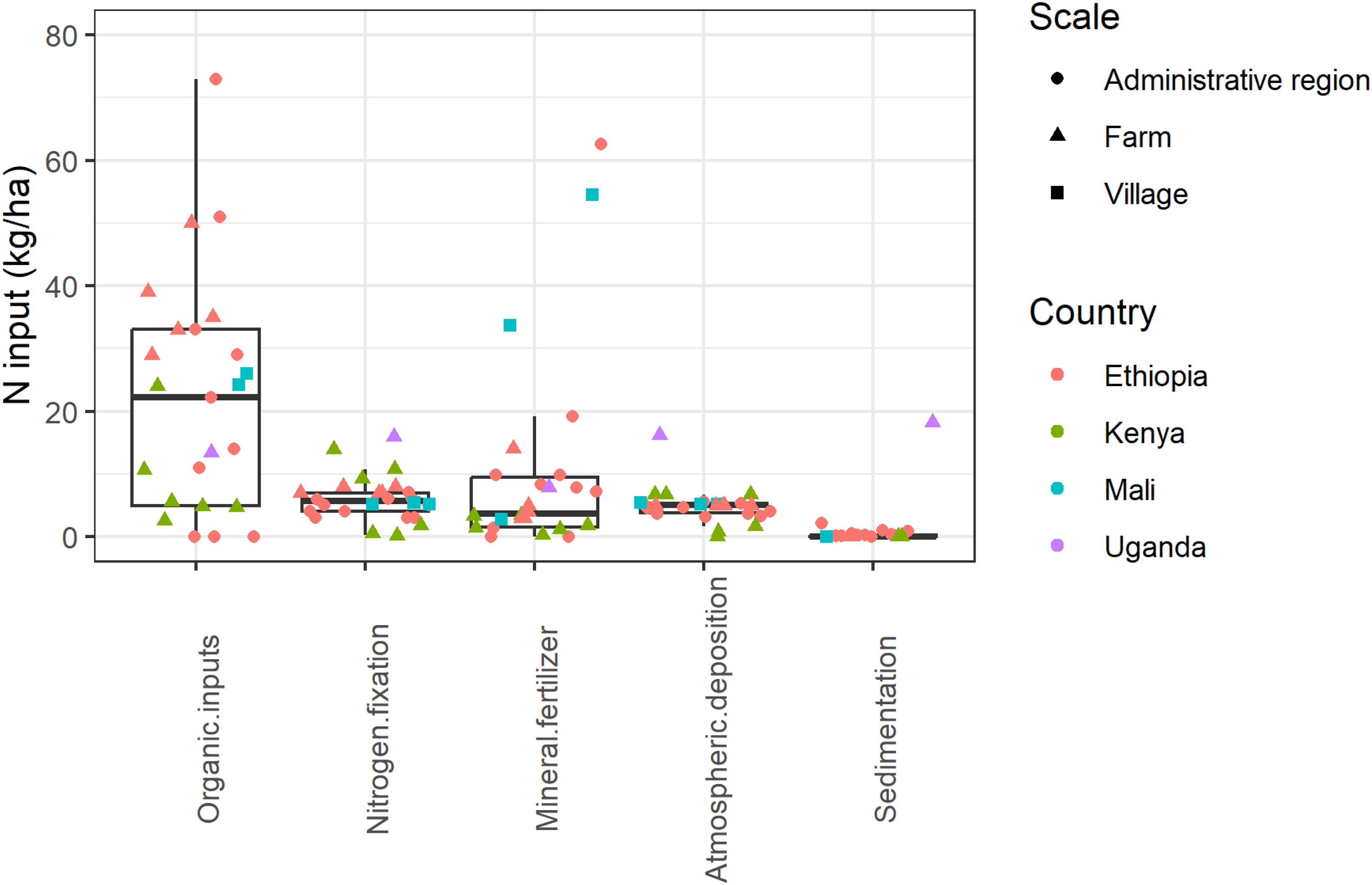
Sources of nitrogen (N) input to farming system (kg/ha/yr) in five selected studies across sub-Saharan Africa. Ethiopia: Haileslassie et al. (2005) and Haileslassie et al. (2007); Kenya: Gachimbi et al. (2005) and Onduru and Preez (2007); Mali: Ramisch (2005); Uganda: Nkonya et al. (2005).
Fertilizer use on cropland (including perennials) is currently very low in SSA, on average 13 kg N/ha/year, 3 kg P/ha/year, and 3 kg K/ha/year (FAOSTAT, 2023). This contrasts sharply with other regions of the world, e.g., Northern America with 73, 11, and 21 kg/ha/year for N, P, and K, and China with 170, 29, and 54 kg/ha/year for N, P, and K, respectively. The numbers for Africa mask, however, a great diversity across countries and agroecological regions within countries (Vanlauwe et al., 2023). In Africa, fertilizers are predominantly used in horticultural systems and large commercial farms, whereas their use is extremely limited in smallholder farms. Southern Africa has witnessed a sharper increase in mineral fertilizer use in the 1960s compared with East and West Africa, possibly because of the thriving large-scale farm sector in the region. In the 1980s, the phasing-out of governmental subsidies for inputs, as a result of structural adjustment programs, has caused mineral fertilizer use to decrease in several countries of SSA (Jayne et al., 2018).
One could therefore argue that smallholder mixed crop-livestock farming systems across SSA do not need to engage in an “agroecological” transition, given their limited dependency on external inputs in the form of mineral fertilizer, and their reliance on biological N fixation and recycling of plant biomass. Yet, a close look at the nutrient balances of cropping systems across the region suggests that long-term agro-environmental sustainability is not granted. Soil nutrient mining is widespread: Smaling et al. (1997) estimated that the cultivated land in SSA was losing annually 22 kg of N, 2.5 kg of P, and 15 kg of K per hectare. These negative balances have continued to increase over time (Vanlauwe et al., 2023). A comprehensive literature review indicated that more than 75% of nutrient balance studies across SSA concluded on negative balances for N and K, and about 60% of studies for P (Cobo et al., 2010). Long-term experiments conducted across the region also point to substantial soil nutrient mining over time; even the biennial rotation system of millet and groundnut in the F. albida parklands in central Senegal has shown evident negative N and P balances (Pieri, 1989). Similarly, the millet cropping system of western Niger with fallowing and strong livestock-mediated nutrient transfers was found to lose nutrients (Powell et al., 2004). The nutrient inputs derived from agroecological processes, as currently harnessed by smallholder farmers in SSA, combined with the current low fertilizer use, cannot compensate for nutrient exports and losses, that mainly occur in the form of crop harvest products, animal removals, leaching, and soil erosion. Demographic pressure and the related cropland expansion at the expense of rangelands and fallows add further constraints on soil fertility maintenance with current agroecological practices (Schlecht et al., 2004).
This happens in the context of low crop yields compared to the yield potential that could be achieved based on the region's climate and soil properties (Affholder et al., 2013). This discrepancy can be largely attributed to low nutrient availability in the soils (Sanchez, 2002). Yields need to substantially increase to meet the current and future food demand and alleviate rural poverty in SSA while minimizing deforestation and other forms of land-use change. Narrowing yield gaps in SSA can contribute to increasing food security at household, national, and continental levels (Gérard et al., 2020; Giller et al., 2021a; van Ittersum et al., 2016). For example, modest increases in maize yield were found to greatly contribute to improved food security in Tanzania and Uganda, though this depended strongly on local contexts (Falconnier et al., 2023). Yet, increases in cereal productivity can only be achieved if nutrient inputs increase substantially (ten Berge et al., 2019). Across several case studies, the estimated maize grain yields that are compatible with food security ranged from 3.5 to 11.9 t/ha (median 6.5 t/ha, Table 1). This translates into N uptake (grain + straw) that ranges from 54 to 228 kg N/ha, with a median of 113 kg N/ha (Table 1 and Figure 2A). For a maize-based cropping system to be sustainable, these crop exports need to be compensated by some form of nutrient inputs. If mineral fertilizers were to be reduced as per the “input reduction” claim of agroecology, other forms of input need to be drastically increased. In the next sections, we examine to what extent legumes and biological N fixation, and the recycling of plant biomass through composting and use of cattle manure, can help bring the N, P, and K required.
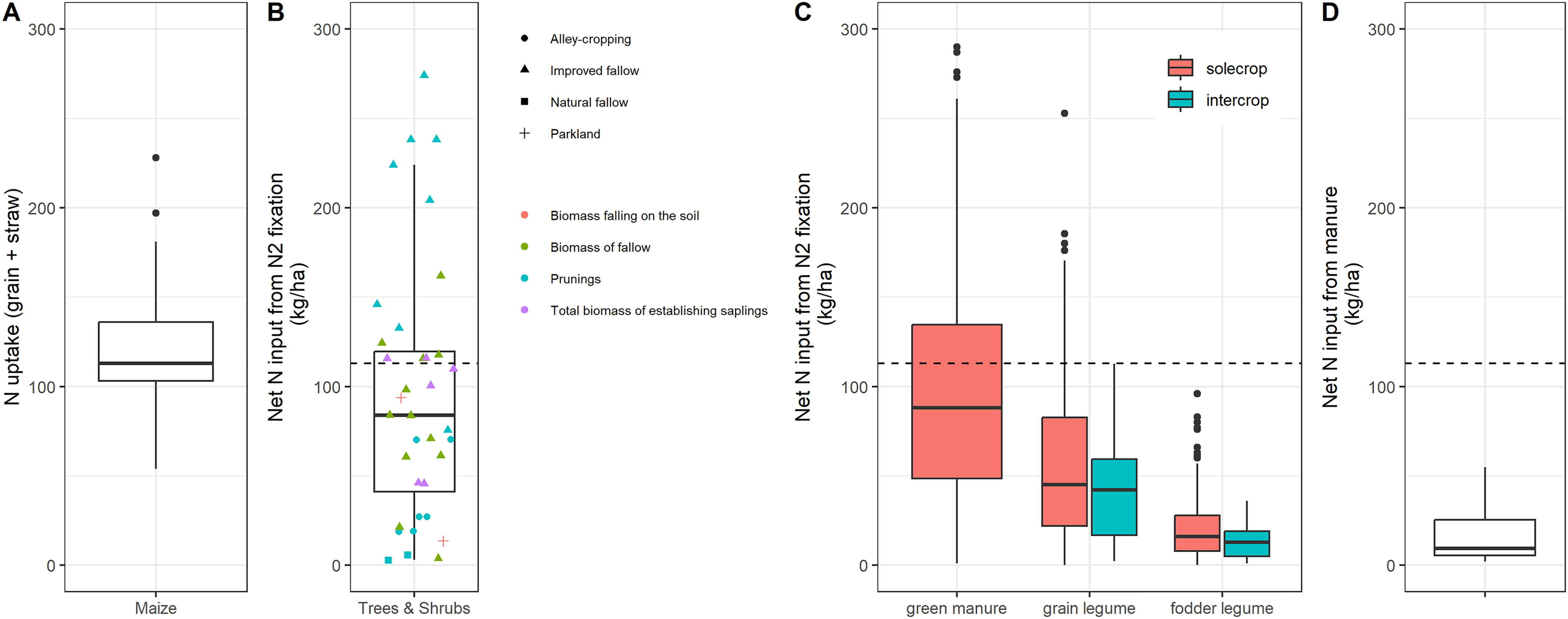
Boxplot of (A) estimated maize nitrogen (N) uptake (in grain + straw) for grain yield compatible with food security in four selected studies (see Table 1 for details of the four studies), (B) Net N input from biological N fixation from trees (13 studies), (C) Net N input from biological N fixation from green manures (21 studies), grain legumes (59 studies) and fodder legumes (60 studies), (D) Net N input from manure (four studies). For grain legumes, we assumed that all grain was exported and only stover was returned to the field, with a harvest index of 0.4 for soybean and groundnut and 0.35 for other grain legumes (Herridge et al., 2008). For fodder legumes, the studies are the same as for green manure (excluding those that are not palatable), but we assumed that only roots were returned to the soil. N fixed in roots was estimated using a root: shoot ratio of 0.3 for the majority of studies where it was not quantified. The dotted horizontal bar in B, C, and D is the median maize uptake (see A). In B, two outliers are not displayed (1063 and 1026 kg/ha, Peoples et al., 1996). See Table S1 for details on the reviewed studies.
Estimated* nitrogen (N), phosphorus (P), and potassium (K) uptake by maize (in grain + straw) (kg/ha) for grain yield (kg/ha) compatible with food security in four selected studies.
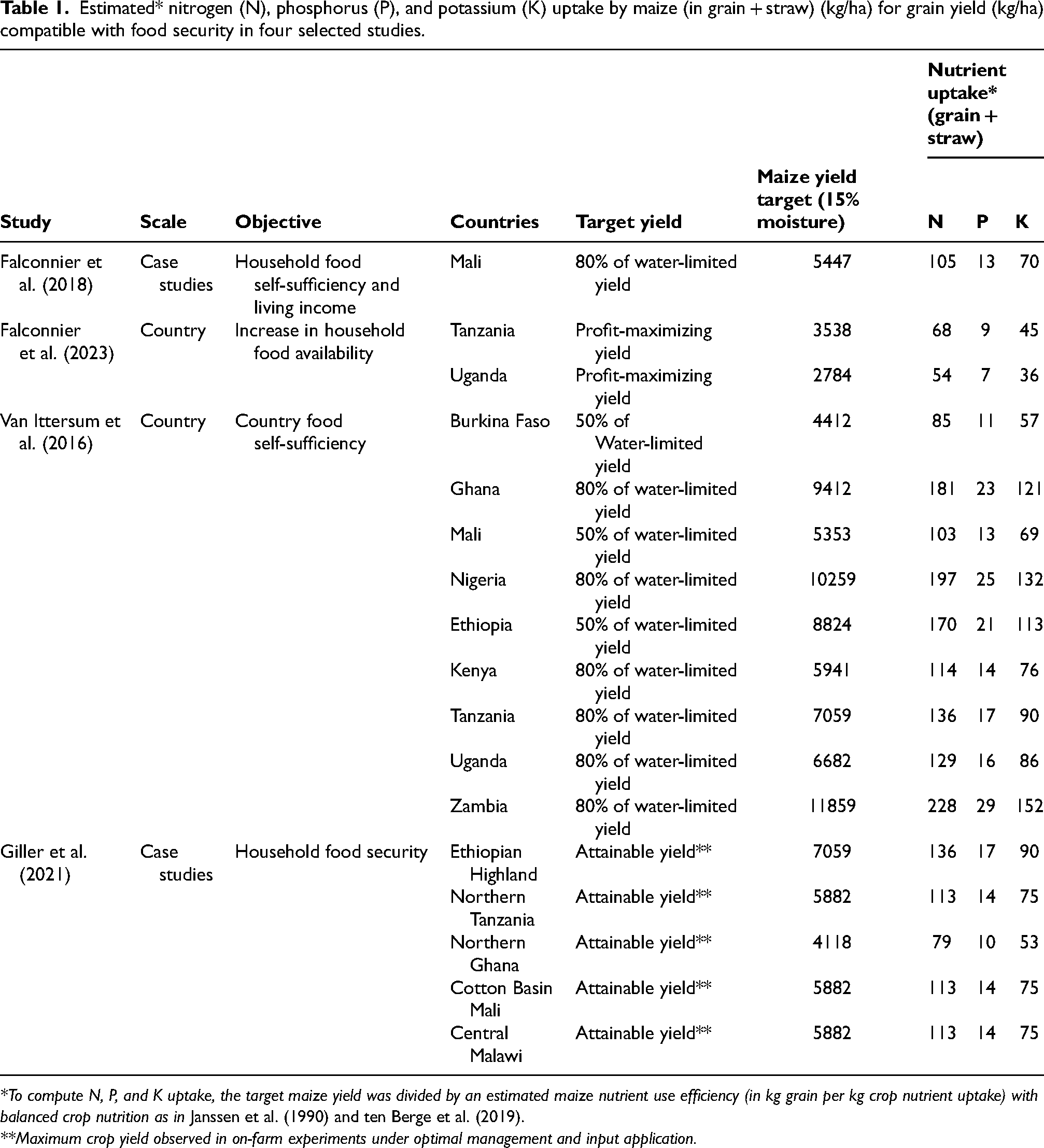
*To compute N, P, and K uptake, the target maize yield was divided by an estimated maize nutrient use efficiency (in kg grain per kg crop nutrient uptake) with balanced crop nutrition as in Janssen et al. (1990) and ten Berge et al. (2019).
**Maximum crop yield observed in on-farm experiments under optimal management and input application.
The nitrogen needs of crops cannot be sufficiently met solely through biological nitrogen fixation by legumes and recycling of animal manure
Most legumes obtain N from the atmosphere through a symbiotic relationship with rhizobia bacteria. Plant N originating from biological fixation, if not exported from the field, is a net input to the soil-crop system. Four types of legumes can be distinguished: trees and shrubs, green manures (i.e., legumes primarily cultivated for biomass to be incorporated into the soil), grain legumes (legumes primarily cultivated for food), and fodder legumes (legumes primarily cultivated to provide feed and fodder to livestock). Figures 2B–C clearly demonstrate that on average, the potential net input from biological N fixation is greatest for legume trees and shrubs, and green manures, followed by grain legumes and fodder legumes. In what follows we scrutinize the constraints to fully achieve high levels of net N inputs with the integration of legumes into cropping systems.
Nitrogen fixation by trees and shrubs
Legume trees and shrubs could contribute as much as 100 kg N/ha/year through N fixation (median of 13 studies, Figure 2B). The net N inputs vary considerably, owing to the way perennials are integrated into cropping systems, and which plant parts are left on or incorporated into the soil (Figure 2B). Trees and shrubs can be integrated into cropping systems in at least three different ways. The first is improved woody legume fallows (Sileshi et al., 2008), where trees are planted to give a large amount of high-quality biomass that can be incorporated into the soil to release nutrients for the subsequent crop. Improved fallows usually take two years to be implemented and can be considered a short-term benefit for soil fertility, but with a strong trade-off with food security because food crops cannot be grown during the fallow period. Labor for planting trees and weeding can also be a constraint to the adoption of these systems by farmers (Giller, 2001). A second option to integrate trees are parklands, i.e., the deliberate retention of scattered trees in farmers’ field so that mature trees coexist with crops. In the parklands of the Sahel region, it is common to find F. albida trees, that shed their leaves during the growing season, so that competition with the annual crop for light, water and nutrients is limited (Sida et al., 2018). However, only mature trees aged 20–40 years fix substantial amounts of N (Giller, 2001), which makes it a long-term investment for farmers. Although more observations are needed, net N inputs from parklands seem also lower than those from improved woody legume fallows (Figure 2B). Besides, there is a threshold beyond which the increase in tree density in parklands no longer benefits the crop (Leroux et al., 2020). A third option is alley cropping, where trees or shrubs are planted in rows alongside the crops, and their prunings can be left on or incorporated into the soil (Sileshi et al., 2011). Farmers’ adoption of alley cropping has been generally low (e.g., Swinkels and Franzel, 1997), unless employed on contours and used as fodder (see section on fodder legumes). Although alley cropping offers the prospect of alleviating the trade-off between food security and environmental sustainability, because food crops and trees can be mixed, trees often compete strongly with the crops for light and water, especially during dry years (Rao et al., 1991).
Nitrogen fixation by green manures, grain and fodder legumes
Green manures have similar potential net N inputs to cropland compared with trees and shrubs (median of 91 kg N/ha/year across 20 studies, Figure 2C). Although green manures rotated with cereals can bring a substantial yield benefit to the cereal (e.g., Ripoche et al., 2021), this comes at the cost of foregoing a part of the cereal yield, similar to woody fallows. The increase in cereal yield from the green manure often does not sufficiently compensate for this yield penalty (e.g., Ojiem et al., 2014; Ranaivoson et al., 2022). The small farm sizes exacerbate the challenge of losing a growing season for cereals, thereby becoming a major obstacle to the adoption of green manures. This concern has been very often voiced by farmers, along with the lack of consumable or marketable products derived from green manures (Vanlauwe et al., 2003). Additional labor is also another constraint, as incorporating the biomass from green manure into the soil can take as much as 80 persons per day per ha for a prolific Mucuna in Malawi (Vanlauwe and Giller, 2006), i.e., 40 times more than what is required to apply mineral N fertilizer (Ojiem et al., 2014).
Grain legumes offer directly consumable and marketable products, e.g., the grains of cowpea, soybean, common beans, and groundnut and are therefore preferred by farmers over woody fallows and green manures. However, the benefits in terms of soil fertility are smaller because the grains are removed from the field (Figure 2C). Franke et al. (2018) showed that on average grain legumes increase cereal yield by 0.49 t ha−1 via their precrop effects. Such yield increase is considerable, but far below the strong cereal yield increases needed to reach food security in SSA (see the first section). There is a strong trade-off between legume grain yield and the residual soil nutrient benefits (Giller et al., 1994). High-yielding soybean varieties with high N-harvest index can even be “net removers” of soil N (Vanlauwe and Giller, 2006). Overall, the grain productivity of legumes is far below that of cereals, for a labor demand that is generally greater (Ojiem et al., 2014), which is a major impediment to the adoption of grain legumes by smallholder farmers.
Plant residues, including prunings from shrubs and trees, as well as the stems and leaves of grain legumes, are often of great value as fodder for livestock (except some green manures with toxic components like in some species of Crotalaria). For example, farmers in East Africa adopted the cultivation of fodder shrubs like Leucaena trichandra on contours (Wambugu et al., 2011). However, with fodder legumes, all aboveground biomass is exported from the fields, and only roots remain, representing a small N source for the following crop (Figure 2C). On the other hand, part of the fodder biomass and its nutrients can be returned to the field in the form of manure (see the section on manure).
Nitrogen fixation in intercropping with grain legumes
Grain legume intercropping can be a promising alternative to cereal-legume rotations, as it allows for the maintenance of cereal production. Although the proportion of N uptake by the legumes through biological fixation is usually similar in intercropping compared with sole cropping (e.g., Namatsheve et al., 2020), the absolute amounts, and the potential net inputs to the cropping system, are smaller (Figure 2C). This is due to the lower plant density of the legume in intercropping compared with the sole cropping, but also because of competition for light and water with the cereal crop (e.g., Traoré et al., 2022). For example, Namatsheve et al. (2020) found that the grain productivity of intercropped cowpeas was on average around 50% of that of sole cowpeas, whereas the yields of the intercropped cereals were around 80% of those of the sole crops.
Sustaining high cereal productivity through biological N fixation?
Only the legume options that compete strongly with food production (woody fallows and green manure) can bring substantial N inputs from biological N fixation (Figure 2B) in the range of what would offset the N exports (grain + straw) of a maize crop with a yield level (∼6.5 t/ha) that is compatible with food security objectives (Figure 2A). Other options that compete less with food production and/or offer direct value for farmers (grain and fodder legumes, Figure 2C) can only bring a fraction of what would be required to offset the N exports by maize.
The estimated contributions of biological N fixation to the net N inputs to cropland, as shown in Figure 2, are possibly overestimated. Firstly, this is because only a small fraction (less than 30%) of the N in green manure and legume tree prunings is effectively recovered by the following cereal crop. Part of it can be leached, volatilized or immobilized by soil microorganisms (Giller and Cadisch, 1995). Secondly, high levels of N fixation can only be achieved if P is not limiting, and if appropriate inoculation is performed when the legume nodulates poorly. These conditions are usually observed in on-station and/or researcher-managed trials (corresponding to most of the estimates of Figure 2) but do not necessarily hold true in farmers’ fields. Thirdly, pests and diseases, usually controlled in research experiments, are an additional barrier in farmers’ fields that must be overcome for the full realization of the potential benefits of legume N fixation.
In conclusion, although legumes can contribute substantially to N input to cropping systems, it is unlikely that they alone can sustain agricultural productivity at the high level that is required for food security.
Greater amounts of manure are not achievable based on herd size and fodder resources
A simple calculation indicates that about 9 t manure (dry matter) per ha per year would be required to balance the N exports by a maize crop yielding 6.5 t/ha with a corresponding export of 125 kg N/ha in its grain and stover (see Table 1), considering that manure from smallholder farms in SSA contains on average 1.4% N (Pieri, 1989). This is largely beyond the 4 t/ha typically applied by cattle owners on a portion of their cropland in, e.g., Zimbabwe (Rusinamhodzi et al., 2013; Zingore et al., 2008). Nine Tropical Livestock Units (TLU), corresponding to approximately 13 cattle or 90 goats, are required to produce 9 t of manure (considering that one TLU produces 2.8 kg of manure per day, Berre et al., 2021). This number of TLU is far beyond what smallholder farmers typically own: for example, the average TLU across nationally representative household samples (Living Standards Measurement Study—Integrated Surveys on Agriculture, LSMS-ISA data) was 1.39 in Uganda and 1.43 in Tanzania (Figure 3). Because it is not feasible to collect all the produced manure and part of its N is lost during storage, composting and transport (Vayssières and Rufino, 2012), farmers recycle far less than what is theoretically possible. Even farmers owning 12 TLU or more cannot recycle the required 125 kg N/ha (Figure 3). Thus, recycling of N through manure falls short of what is required to sustain the necessary levels of cereal yields (Figure 2A, 2D).
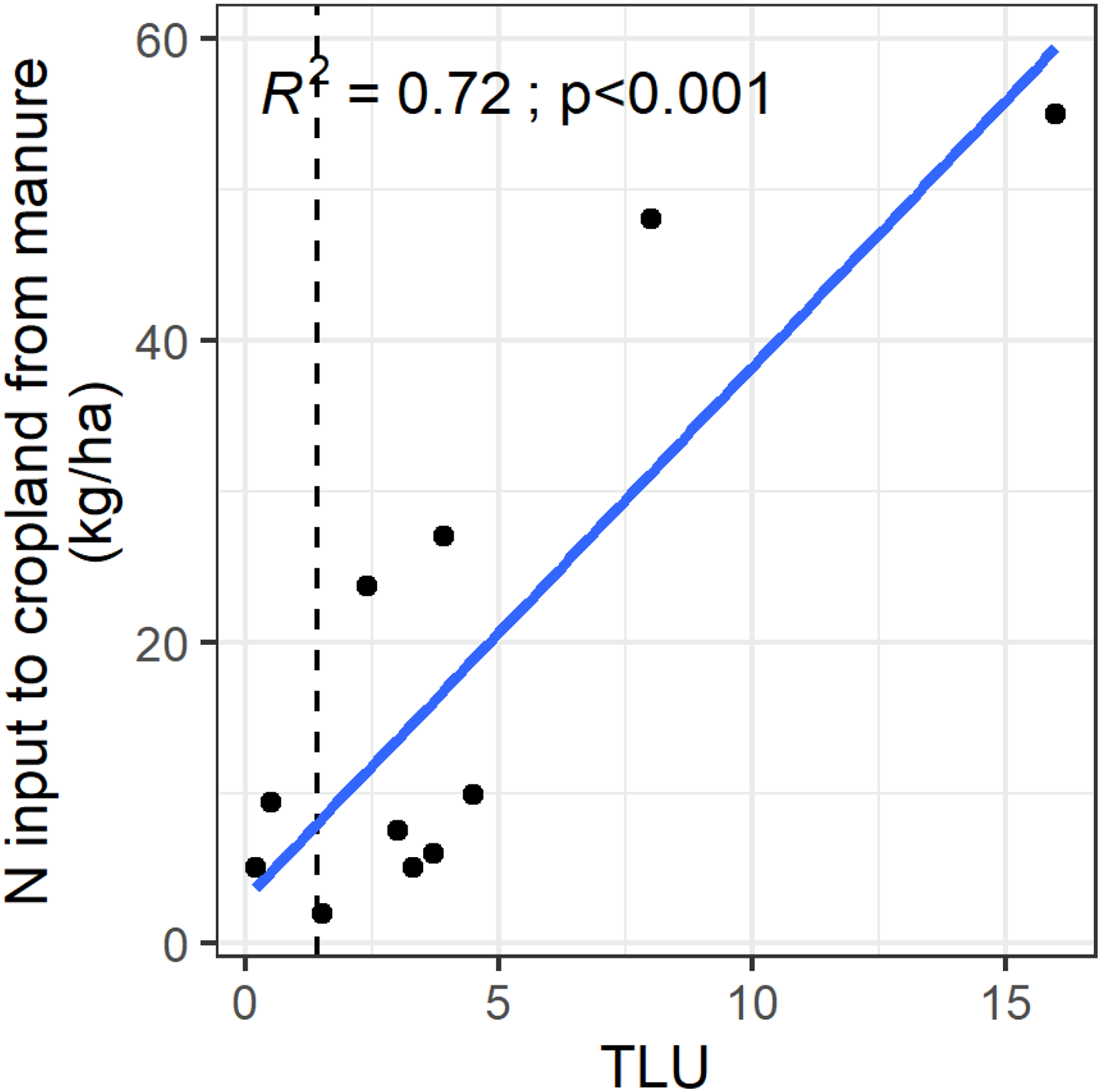
Nitrogen (N) input to cropland in the form of manure as a function of Topical Livestock Unit (TLU) ownership (four studies with 11 observations). The vertical dotted line is the mean value for TLU for a nationally representative sample of 6638 agricultural household in Uganda and Tanzania (LSMS-ISA data, Kilic et al., 2015).
During the cropping season, livestock cannot access fields and feed mainly on rangelands. One TLU—250 kg liveweight—consumes a maximum of 3% of its liveweight daily (Moran, 2005). Based on a simple calculation, it is estimated that 1.4 ha of rangeland would be required for feeding one TLU in the Sudano-Sahelian environment of West Africa, corresponding to a livestock carrying capacity of 0.7 TLU/ha. This estimation assumes that one TLU requires around 1400 kg of biomass over a six-month growing season and that rangelands produce about 1000 kg of biomass (dry matter) per hectare with 600 mm of rainfall, considering a rainfall productivity of 1.7 kg biomass (dry matter) per millimeter rain (Rufino et al., 2011). In many areas of the Sudano-Sahelian region, livestock density is far beyond this estimated carrying capacity of 0.7 TLU/ha, e.g., 4.8 TLU/ha in western Burkina Faso (Andrieu et al., 2015) and 1.3 TLU/ha in the Senegalese groundnut basin (Grillot et al., 2018). It is also important to note that high grazing pressure often leads to a decline in rangeland productivity (Hiernaux et al., 2009). The above estimations indicate that available feed resources are a strong constraint to maintaining the large herds that would be necessary to produce the manure required to sustain high cereal yields. Furthermore, manure represents a direct transfer of nutrients from rangelands to croplands. For example, Andrieu et al. (2015) estimated that N loss from the rangeland could be as high as 25 kg N/ha/year in western Burkina Faso. Enriching cropland with nutrients from manure therefore comes at the expense of soil nutrient mining in rangelands (Powell et al., 2004).
Phosphorus and potassium need to be imported and replaced continually
For P and K fertilization, agroecological practices rely on the transfers of biomass through manure and compost. It should, however, be noted that with these lateral transfers, the places of origin of the biomass (e.g., rangelands, hedgerows, fields of farmers who do not own cattle and where crop residues are left) are progressively impoverished in P and K. Therefore, we argue that P and K need to be brought by mineral fertilizers if long-term agro-environmental sustainability is to be guaranteed.
In addition, biomass transfers are also often insufficient to meet crop requirements. One reason is that low P and K contents in soil translate into low P and K concentrations in recycled plant materials. Soils of SSA have typically low plant-available P due to low P concentration in the parent material and extended weathering, and because of low soil pH and binding of phosphates by Fe3+ and Al3+ cations (Margenot et al., 2016). Similarly, plant-available K in soils of SSA is generally low because of prolonged weathering over time and insufficient external inputs (Majumdar et al., 2021).
As a result, P and K concentrations in manure and compost are rarely above respectively 0.2 and 0.6%, in SSA (Blanchard et al., 2014; Fall et al., 2000; Zingore et al., 2008). This means that manure inputs of respectively 8 and 14 t/ha would be required to sustain a maize yield of 6.5 t/ha, corresponding to P and K uptake demands of 16 kg P ha−1 and 83 kg K ha−1. It is clear that these P and K requirements for crops are difficult or even impossible to achieve by smallholder farmers, given their current herd sizes and feed resource limitations (see previous section and Schlecht et al., 2004).
Relying solely on biomass transfers in combination with the ongoing depletion of soil P and K might also have implications for the potential of legumes to efficiently fix N. In particular, insufficient availability of P can constrain biological N fixation, as adenosine triphosphate is needed in larger quantities for N-fixing legumes than for other crops (Giller, 2001). Besides, soil K deficiency affects the growth of rhizobia, nodule formation and functioning, resulting in a depressing effect on biological N fixation (Divito and Sadras, 2014).
If produced differently and used appropriately, mineral fertilizers cause little harm to the environment
Mineral fertilizers are often and rightly blamed for environmental pollution (Gruber and Galloway, 2008). Pollution arises from both their use and their production. In what follows, we analyze opportunities to address the negative environmental outcomes of mineral fertilizer, firstly related to their use, and secondly to their production.
In general, fertilizer use contributes to four negative environmental outcomes. First, direct soil emission from the application of mineral fertilizers is the most important source of N2O emission from agriculture, together with emissions from organic fertilizer (Tian et al., 2020). The estimations regarding these two emission sources carry considerable uncertainty, making it challenging to distinctly differentiate between them (IPCC, 2022). Second, NH3 volatilization from fertilizer application can have serious consequences for human health, acidification, and eutrophication of ecosystems (Liu et al., 2023; Paerl et al., 2014), and is also a source of indirect production of N2O. Third, the global increase in N and P fertilizer use has led to the eutrophication of water (Paerl et al., 2014; Peñuelas and Sardans, 2022). Fourth, N fertilizers have resulted in global soil acidification that can undermine soil carbon (C) sequestration and climate mitigation efforts (Raza et al., 2021). Yet, these negative outcomes do not come from the fertilizers per se, but from their excessive or inappropriate use. It is important to note that there are no chemical differences between NH4+ or NO3− ions coming from organic or mineral fertilizers. Plants equally absorb both these two ions regardless of their source while the origins of negative environmental impacts of nutrients vary. For example, N2O emissions are very high in Europe, North America, and East and South Asia, which together consume over 80% of the world's mineral N fertilizers (Guenet et al., 2021; Tian et al., 2020), while it is manure application that is currently the main source of agricultural N2O emissions in SSA (Davidson, 2009; Guenet et al., 2021).
Global N-use efficiency in croplands has dramatically dropped over the last decades and is now only about 40% (Lassaletta et al., 2014; Zhang et al., 2015). The potential to improve fertilizer-use efficiency and reduce pollution in regions of intensive agriculture is therefore large, with a limited negative impact on crop yield (Wuepper et al., 2020). For example, it has been reported that N-use efficiency in Greece increased from 30% in 1990 to more than 70% in 2010 (Lassaletta et al., 2014). In SSA, the scope to improve N-use efficiency is also large, as the current average agronomic efficiency of N is estimated at 14 kg maize grain per kg N applied, compared with the global average of 30 kg kg−1 (Vanlauwe et al., 2023). In general, improvement of nutrient-use efficiency can be achieved by widely adopting the 4R principles (the right nutrient source at the right rate, right time and right place) (Penuelas et al., 2023). In the context of SSA, site-specific nutrient management tailored to the high spatial heterogeneity of smallholder farming systems will be key (Chivenge et al., 2021). An analysis of 11 published studies showed that the addition of N fertilizer increased N2O emissions compared to the no input control, but there were no differences among the different rates of N fertilizer application (Vanlauwe et al., 2023). In addition, even with the high N fertilizer rates, N2O emissions remained below 1 kg N2O-N ha−1. Nitrogen fertilizer application at a rate four to five times greater than the current average in SSA (i.e., 60 kg N/ha), using one basal application and two top dressings, led to low N2O emissions in Zimbabwe, far below the IPCC default values (i.e., 0.5 to 1% of applied mineral N) (Shumba et al., 2023).
With regard to production, the synthesis of NH3 to produce mineral N fertilizers accounts for about 0.8–1.2% of the global anthropogenic CO2 emissions and 2% of global energy (Gao and Serrenhoo, 2023; Smith et al., 2020). Ammonia emissions during fertilizer production are also responsible for indirect N2O emissions. These GHG emissions could, however, be largely reduced by the production of NH3 using hydrogen from water hydrolysis with renewable energy instead of fossil fuels (Smith et al., 2020; Wang et al., 2018). This prospect is currently significant enough for the IPCC to consider NH3 as a potential low- or zero-carbon fuel similar to hydrogen (IPCC, 2022). The current expansion of fertilizer manufacturing witnessed across Africa could also largely reduce GHG emissions due to transport, besides increasing the sovereignty of the region for access to fertilizer.
Overall, it is estimated that GHG emissions from N fertilizer production and use could be reduced by up to one-fifth of current levels by 2050 with currently available technologies, increasing N-use efficiency being the most effective one, as about two-thirds of GHG emissions from fertilizers actually take place immediately after their application in croplands (Gao and Serrenhoo, 2023). The excess of fertilizers currently being used in regions where overapplication is prevalent could be redistributed to SSA (less in the Global North, more in the Global South), possibly without the need to produce more fertilizers at the global scale.
Eutrophication is a rising issue throughout Africa, particularly around large and rapidly expanding cities (Nyenje et al., 2010), but it is currently most probably driven by untreated sewage waste and erosion deposits rather than by the excessive fertilizer use in agriculture (Vanlauwe and Giller, 2006). However, if fertilizer use is to increase, its contribution to eutrophication has to be carefully monitored. Relevant “4R” fertilizer practices that are crucial to increase profitability and reduce gaseous losses will also contribute to limit N and P leaching and their contribution to eutrophication.
Reducing the use of mineral fertilizer hampers crop productivity gains and contributes indirectly to agricultural expansion
The impacts of a strategy where the use of mineral fertilizer is reduced or eliminated, as purported by the “input reduction” principle of agroecology should be carefully evaluated, and weighed against the negative environmental impacts associated with fertilizer use. Because nutrient limitations constrain crop yields (see the first section), reducing fertilizer use will further lower crop yields, and inevitably drive cropland expansion to meet the demands of a fast-growing population in SSA. Although increased fertilizer use for higher crop productivity will cause an increase in N2O emissions (e.g., Leitner et al., 2020), cropland expansion into natural ecosystems would trigger an even greater ecological footprint. Conversion of natural ecosystems into cropland is associated with accelerated vegetation and soil C loss, and loss of biodiversity (Searchinger et al., 2015). Kehoe et al. (2017) showed greater biodiversity losses with cropland expansion than with agricultural intensification, globally and in SSA in particular. Van Loon et al. (2019) projected greater GHG emissions associated with cropland expansion than with agricultural intensification to meet food security in SSA by 2050. In addition, even though SSA is generally considered as having abundant land, accounting for about 60% of the world's uncultivated arable land, this abundance is primarily concentrated in less than 10 countries. Most countries in the region are facing land scarcity. This makes cropland expansion a limited option, emphasizing the need for production intensification on existing cropland. For a net positive effect of intensification and fertilizer use on the GHG balance of Africa to take place, public policies and stronger governance of natural resources will have to be put in place to prevent rebound effects (i.e., increased deforestation due to increased profitability of farming) (Byerlee et al., 2014). In this context, it is noteworthy to highlight that a number of countries in the South, such as Vietnam, India, Bhutan, El Salvador and Chile, have successfully achieved a land-use transition in recent decades through sound policies and innovations, which has led to the simultaneous increase of their forest cover and agricultural production (Lambin and Meyfroidt, 2011).
Agroecological principles help improve soil health, nutrient-use efficiency and sustain crop productivity
Maintaining soil health
It is clear from what precedes that harnessing only biomass fluxes and biological N fixation cannot bring all the nutrients required to increase food production and meet the food demand of an increasing population. However, these processes and the related practices are key to maintain or improve soil health in general, and soil organic C, in particular (Vanlauwe et al., 2023). Soil health is one of the 13 HLPE principles of agroecology, and is reflected in three of the ten FAO agroecology elements, namely diversity, synergy and resilience (Wezel et al., 2020). Degraded soils, i.e., soils with poor health and with chemical and physical properties of low quality are estimated to occur in 25% of croplands globally, across all environmental conditions (Le et al., 2016). In what follows we show that two agroecology principles (recycling and biodiversity) and associated practices (the use of organic amendments and agroforestry) are key to maintain and improve soil health.
Degraded soils tend to be poorly responsive to mineral fertilizers (Nezomba et al., 2015). The application or retention of organic resources is critical to maintain or restore soil health (Laub et al., 2023). Crop residues are the most abundant and available organic resource, and their retention in croplands can play a significant role. However, due to other competing demands for animal feed, fuel and construction, amounts of crop residues returned to cropland are usually low (Valbuena et al., 2012). In this context, manure is often the most critical resource to maintain soil health, but required application rates may be prohibitive to smallholders (see section on manure). On the other hand, there is a large potential to increase the biomass and nutrients being recycled through manure by adopting better herd management (e.g., cattle housing) and improved manure storage (e.g., heap coverage) (Rufino et al., 2007). Improved recycling is thus required if soil organic matter is to be maintained in soils. Manure application can also mitigate soil acidification resulting from the continuous application of N fertilizers on soils with low cation exchange capacity and low pH buffering capacity (Vanlauwe et al., 2001).
Agroforestry—defined in its broad sense as “land-use systems and practices in which woody perennials are deliberately integrated with crops and/or animals on the same land management unit” (Leakey, 1996)—can also maintain and improve soil health. Agroforestry is based on the biodiversity principle. Compared to annuals, some perennials have deeper root systems and fix more C, capture nutrients from deeper soil layers and produce biomass and litter for a longer period of time; their inclusion in cropping systems can thus contribute to a greater input of organic C to the soil (Corbeels et al., 2019), but also lead to nutrient cycles (e.g., N, P) that are more tightly coupled to the C cycle (by stoichiometric relations in plant biomass, organic matter and microbial biomass), and thus are less leaky (Lemaire et al., 2014).
Improving nutrient-use efficiency
Efficiency is a key principle of agroecology (captured in both 13 HLPE principles and the 10 FAO elements), but also a concept that is central to the ecological and economic sustainability of any agricultural production system, whether agroecological or not (Kumar et al., 2020).
Long-term improvement of soil health as described above can lead to more nutrients being supplied by the decomposition of soil organic matter, complementing or substituting some of the nutrients that need to be supplied by mineral fertilizers (Myers et al., 1994). Although nutrients other than N “need to be imported and replaced continually” (see section on P and K), agroecological practices help maximize their use efficiency. In the case of P in particular, the inclusion of legumes (principle of biodiversity) can lead to higher nutrient-use efficiency, as legumes tend to have greater rates of P acquisition than cereals (e.g., Tang et al., 2021). Often, the efficiency of mineral fertilizer improves with the use of agroecological practices. For example, the incorporation of Crotaliaria-Mucuna green manures into the soil improved N-use efficiency of the subsequent upland rice in Madagascar (Ranaivoson et al., 2022); the use of cattle manure was responsible for increased maize response to mineral fertilizer in Kenyan smallholder farms (Njoroge et al., 2019); intercropping maize with soybean gave greater fertilizer N-use efficiency across 106 observations in Africa (Xu et al., 2020). More generally, experimental evidence suggests that increased plant diversity—in particular functional diversity tends to lead to higher nutrient-use efficiency (Furey and Tilman, 2021). Biomass recycling (through manure, crop residues; principle of recycling) also leads to P being retained in the labile organic soil pools, which may lead to higher P-use efficiency (Drinkwater et al., 2017). Similarly, nutrient-use efficiency was shown to be promoted by nutrient recycling through livestock grazing (e.g., Hiernaux and Turner, 1996). Finally, better nutrient-use efficiency also decreases the risk of negative cost-benefit ratios when farmers invest in fertilizers (Bielders and Gérard, 2015).
Maintaining other ecosystem services for sustained crop productivity
Bommarco et al. (2013) highlighted the link between supporting and regulating ecosystem services, crop productivity and yield gaps, both in low-input and intensively cultivated production systems. Soil fertility maintenance and nutrient cycling are key supporting ecosystem services for agriculture and have been reviewed in the previous sections of this article. Biological pest and weed control, and crop pollination are two regulating ecosystem services that are equally critical for food production.
Crop losses due to pests are projected to increase with climate change (Deutsch et al., 2018) while the use of insecticides in agriculture is increasingly regulated and its cost-benefit ratio is declining globally (Zhang, 2018). Weeds also contribute substantially to yield gaps globally for a range of crops (maize, wheat, cotton, soybean) (Oerke, 2006). As a result, biological pest and weed control is becoming more critical, in both low-input and intensively cultivated production systems. Similarly, the global pollinator decline is threatening all regions of the world. Yield losses due to pollinator decline have been recorded globally for crops with a moderate dependence on pollination. On the other hand, yields of crops with a high dependence on pollination may have been maintained by their stronger reliance on domestic honeybees, whose stocks are, however, growing slower than the demand for pollination (Aizen and Harder, 2009; Garibaldi et al., 2009).
Biological pest and weed control, and pollination may be stimulated by increasing on-farm diversity, a key agroecology principle. For example, the populations of wild pollinators, and predatory and parasitoid arthropods may increase following the retention of natural and semi-natural patches in cropland, that provide nectar, alternative preys, and overwintering and nesting habitats (Bianchi et al., 2006). These patches may also host and provide cropland with insectivorous birds and bats (Sow et al., 2020). Natural areas may also provide agricultural areas with herbivores feeding on weeds, and with pathogens attacking crop pests (Blitzer et al., 2012). The optimal amount of noncrop habitat and the optimal spatial arrangement are species-specific, and depend on the dispersal ability and sensitivity to disturbance (including edge effects) of the species of interest (Ekroos et al., 2016). With regard to weeds, diversified crop rotations including legumes can help decrease weed pressure and its impact on cereal yield (e.g., Ripoche et al., 2021). Appropriate biological pest and weed control is a critical step to ensure appropriate crop growth and improved nutrient-use efficiency (ten Berge et al., 2019).
Conclusions
The analysis of existing scientific literature on nutrient balances in SSA, biological N fixation of tropical legumes, and manure production and use in smallholder farming systems shows that agroecological practices have a clear role to play when it comes to maintaining and improving soil fertility in the context of smallholder farmers in SSA. However, further reducing the already low use of mineral fertilizer—as per the “input reduction” principle of agroecology—may aggravate the current challenge of nutrient depletion in croplands and rangelands throughout SSA. Our analysis shows that biophysical production factors, but also shortages of land and farm labor, constrain the opportunity to close crop nutrient gaps and increase productivity by relying solely on N fixation and transfers of biomass using manure. More research at the farm and landscape level where issues of nutrient availability and sustainability operate, is needed to explore the benefits and constraints of agroecology. It is, however, clear that agroecological practices can improve nutrient-use efficiency and should complement the use of mineral fertilizer, provided that financial and physical access to fertilizer is guaranteed through adequate market development. Moreover, effective policy support to agroecological practices that deliver great environmental benefits for the society, but often limited short-term benefits to farmers, requires urgent attention.
Supplemental Material
sj-docx-1-oag-10.1177_00307270231199795 - Supplemental material for The input reduction principle of agroecology is wrong when it comes to mineral fertilizer use in sub-Saharan Africa
Supplemental material, sj-docx-1-oag-10.1177_00307270231199795 for The input reduction principle of agroecology is wrong when it comes to mineral fertilizer use in sub-Saharan Africa by Gatien N Falconnier, Rémi Cardinael, Marc Corbeels, Frédéric Baudron, Pauline Chivenge, Antoine Couëdel, Aude Ripoche, François Affholder, Krishna Naudin, Emilie Benaillon, Leonard Rusinamhodzi, Louise Leroux, Bernard Vanlauwe and Ken E Giller in Outlook on Agriculture
Footnotes
Author's note
François Affholder, Universidade Eduardo Mondlane, Faculdade de Agronomia e Engenharia Florestal, Maputo.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the European Union’s Horizon 2020 research and innovation program, delEU-DESIRA program [grant numbers 101083653, project CANALLS (Driving agroecological transitions in the humid tropics of Central and Eastern Africa through transdisciplinary agroecology Living Labs) and 424-933, project RAIZ (Promoting agroecological intensification for resilience building in Zimbabwe)].
Supplemental material
Supplemental material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.