
Abstract
Diversification measures are widely recognized for their potential to enhance the resilience of agri-food production systems under global change. Nitrogen-fixing legumes commonly feature in diversification schemes, where they exert positive impacts on soil structure, nutrient retention, carbon sequestration or fertilizer reduction. Though legumes are premier tools for conservation agriculture, their impacts on (invertebrate, microbial) biological control are far less clear. As legumes secrete copious amounts of energy-rich (floral, extra-floral) nectar and provide alternative host or prey items for resident natural enemies, their deployment is expected to benefit biological control. Yet, its mechanistic basis is poorly understood and scientific underpinnings weak. In this Perspective piece, we collate data from the published literature and open-access databases on the extent to which different natural enemy guilds interact with 25 legume species that are commonly used as intercrops in the (sub)tropics. Our work unveils how natural enemies regularly forage on legumes, but the portrayal of those interaction linkages –or “biostructure” data- is profoundly incomplete and their ensuing implications for biological control are under-investigated. For instance, merely 4 (out of 167) parasitoids of the globally invasive fall armyworm Spodoptera frugiperda have been recorded in association with legumes. We hope that this Perspective piece will stimulate further thought, drive a new wave of biodiversity x ecosystem function research, and ultimately result in the design and implementation (or preservation) of more resilient, resource-frugal and pest-suppressive farming systems.
Keywords
Legume intercrops: core pillars of ecological intensification
Across the globe, the food system has become a foremost contributor to global change, driving land-use change and biodiversity loss, depleting non-renewable natural resources and releasing more than one third of anthropogenic greenhouse gas emissions (Crippa et al., 2021; Rockström et al., 2020). In the absence of large-scale “transformative” change and dedicated mitigation efforts, feeding a swelling world population without jeopardizing Earth system resilience may soon prove unfeasible (Springmann et al., 2018). To redress global change, ambitious goals need to be set and urgent action is required to implement best-bet solutions (Díaz et al., 2019). Several options are on the table to alleviate the environmental pressures that emanate from agri-food production. Crop diversification constitutes a particularly lucrative solution. Increasing (plant) biodiversity in cropping systems e.g., through intercropping can help to conserve or even raise a wide arc of regulating and supporting ecosystem services – which translate in 33–38% higher gross outputs and incomes (Isbell et al., 2017; Li et al., 2020; Martin-Guay et al., 2018; Rosa-Schleich et al., 2019; Tamburini et al., 2020). More diverse cropping systems benefit from enhanced plant-animal interactions, which ultimately bolster services such as insect-mediated pollination or biological pest control (Letourneau et al., 2011; Lichtenberg et al., 2017; Wan et al., 2020). The agronomic, environmental and economic spin-offs of diversified cropping systems are particularly appealing in the Global South, where (formal, farmer-led) intercropping research is flourishing (Hufnagel et al., 2020). This cornucopia of benefits can be fully reaped when farmers pursue an ecological intensification pathway (Bommarco et al., 2013; Kleijn et al., 2019) i.e., by harnessing ecological processes in order to maintain yields while curtailing petroleum-derived inputs (Isbell et al., 2017).
Legumes (Fabaceae) are intercrops of choice for many farmers, and legume integration is increasingly seen as a crucial lever to achieve more sustainable food systems (Foyer et al., 2016; McDermott and Wyatt, 2017; Rubiales et al., 2021; Snapp et al., 2019). This is nothing new, as legume-cereal intercrops constituted the foundation of native American societies e.g., the Iroquois or Haudenosaunee and the Maya Indians in the Yucatan Peninsula (Emerson, 1953). Through their distinctive capacity of fixing atmospheric nitrogen in symbiotic root nodules and favoring mycorrhizal fungi, legumes reduce reliance on synthetic fertilizer and lower energy use in agri-food production. Specifically, legume intercrops could cut nitrogen fertilizer use by a quarter in global cereal systems and potentially achieve net land savings (Jensen et al., 2020). Legumes also provide non-nutritional benefits to their non-legume partners, given their renowned role in averting runoff and soil erosion (Rodríguez et al., 2022). Legume integration bears consistent yield benefits among prime food staples or forage crops, which are especially pronounced in low-input environments e.g., in African smallholder pigeon pea or maize systems (Daryanto et al., 2020; Zhao et al., 2022). Equally, grass–legume bicultures in Andean highlands, yield 65% more biomass than monocultures (Meza et al., 2022). Legume-mediated impacts are also felt outside the agronomic realm. As an example, legume-cereal intercrops offer rich foraging habitats to wild and domesticated bees, aiding pollinator conservation (Kirsch et al., 2023; Suso et al., 2016). A deliberate assessment and communication of those multi-service benefits of legume intercrops, can help to take this practice forward (Brooker et al., 2015). Yet, recognition of the multi-service or multi-function role of legumes, is severely impeded by a dearth of multidisciplinary “systems” research and efforts that deliberately look outside of the agronomy “box” (Couëdel et al., 2019; Hufnagel et al., 2020; Martin-Guay et al., 2018). Equally, an informed choice of the “right” type of legume multifunctional intercrop is often impossible due to a deficient understanding of the underlying ecological mechanisms e.g., plant-plant facilitation (Brooker et al., 2021; Cappelli et al., 2022). Also, there is an overwhelming number of cases solely relying on a few species – not including native or locally adapted plants – where positive as well as negative feedbacks differ from those described for the cultivated species in most studies.
Scientific attention is inordinately skewed towards a narrow slice of ecosystem services e.g., food provisioning, erosion prevention or nitrogen fixation. Meanwhile, the impacts of (legume) intercropping on crop health constitute a major “blind spot” (Beillouin et al., 2019; Seppelt et al., 2020). Recently meta-analyses have sought to fill this knowledge gap for disease and weed control (Gu et al., 2021; Zhang et al., 2019). There is however ample scope for natural pest or disease control through legume integration (Gurr et al., 2017; Kocira et al., 2020), even though legumes' actual contribution to crop health may be contingent on the composition, structure, and disturbance regime of the surrounding agro-landscape. Intercrops can either exert direct effects through plant-mediated or bottom-up effects or indirect (natural enemy-mediated or top-down) effects (Civitello et al., 2015; Root, 1973). Plant-mediated effects may then lower the likelihood that specialist herbivores or pathogens find competent hosts or persist in diverse plant communities vs. monocultures. Enemy-mediated effects can provide indirect top-down regulation of herbivores. As legumes often provide shelter, nectar, alternative prey/host items or pollen to resident natural enemies, their integration in habitat management strategies for biological control carries particular promise (Gurr et al., 2017; Landis et al., 2000).
Indeed, legume integration could slow pest population build-up and avert pest-inflicted losses. Legumes such as faba bean flower prolifically and constitute coveted sources of floral nectar or pollen for pollinators and natural enemies alike (Cole et al., 2022; Köpke and Nemecek, 2010). Moreover, at least 153 legume genera bear extrafloral nectaries (Marazzi et al., 2019), which provide freely accessible energy-rich nectar to a suite of natural enemies (Heil, 2015; Lérault et al., 2022; Lundgren, 2009). Lastly, legumes host a range of (non-pest) herbivores that constitute alternative hosts or prey for generalist parasitoids or predators. The above is mirrored in consistently higher levels of biological control in the few legume intercrop systems that have been adequately studied (Iverson et al., 2014). These findings are in line with those of other vegetational diversification tactics (Andow, 1991; Daryanto et al., 2020; Letourneau et al., 2011), with the added advantage that focal crop yields are routinely raised. For example, associating Brazilian coffee plants with extra-floral nectar-bearing legumes raises yields, enhances predatory mite abundance and reduces pest damage (Ferla et al., 2023; Rezende et al., 2021). Yet, all too often, sufficient field-level data are lacking to properly quantify the impact of legume intercrops on biological pest control (Shackelford et al., 2019). Also, broader usage of (legume) companion plants for biological control may have been hampered by a near-exclusive focus on their production of biocidal compounds (Couëdel et al., 2019). Only tangential attention has been given to arthropods – often limited to a simple cataloging of natural enemy visitors without necessarily examining the resulting ecosystem functions or services (Murrell, 2017). Equally, relevant agronomic indicators (e.g., intercrop management, yield or revenue), are often ignored in biological control studies (Chaplin-Kramer et al., 2019). Hence, notwithstanding the major advances in habitat management over the past decades (Gurr et al., 2017; Landis et al., 2000), legume integration for biological control may still be in its infancy.
In this Perspective piece, we conduct a retrospective assessment of the extent to which legume-associated natural enemies (NE) and their ensuing biological control, have been studied in intercrop systems in (sub-)tropical settings. Our work centers on a panel of 25 legume species that are commonly used as intercrops in tropical or subtropical environments of the Global South. Specifically, we use open-source data to log the identity of arthropod predators and parasitoids that are associated with each legume at a global level. Next, we use systematic literature searches to assess the level of scientific attention that has been directed to each legume-associated NE with regard to (1) arthropod biological control in maize or sorghum i.e., two globally-important cereal grains, and (2) biological control in the respective legume-cereal intercrops. This focus is justified as maize is the most commonly studied focal cropin intercropping research (Beillouin et al., 2019). In addition, we conduct an in-depth assessment centered on a globally important pest of cereal grains i.e., the fall armyworm Spodoptera frugiperda (Kenis et al., 2022). For all known S. frugiperda parasitoids, we record their interaction linkages with each legume species and the related extent of scientific attention. As such, our work unveils whether the necessary baseline information on NE biodiversity and biotic (plant x animal) interactions is available to underpin arthropod biological control research and practice in legume-cereal intercrops in the (sub-)tropics.
Cataloguing legume x natural enemy interaction linkages
Based upon Baligar and Fageria (2007) and Li et al. (2013), we composed a panel of 25 legumes that are commonly used as intercrops in tropical or subtropical settings. Next, for each legume species, we queried the Global Biotic Interactions or GloBI search engine (Poelen et al., 2014) using the terms “interacts with,” “has parasite,” “flowers visited by” or “pollinated by.” Queries were conducted between April 15 and May 31, 2023. GloBI automatically extracts (plant, animal, microbial) species interaction data from existing data infrastructures, and resolves and integrates these in a centralized, searchable biodiversity database (https://www.globalbioticinteractions.org/). Using the above search terms, we logged all arthropod parasitoid or predator species that were generated by the GloBI search engine. Species belonging to families with inconsistent zoophytophagous feeding habits e.g., Miridae were not included. Thus, for each legume species, we compiled a list of (trophically, non-trophically) associated natural enemies or biological control agents (BCAs). Next, we compiled a list of all known dipteran and hymenopteran parasitoid species of S. frugiperda in its native and invasive range (Kenis et al., 2022; Molina-Ochoa et al., 2003). For all parasitoid species, we revised the nomenclature and collated all known synonyms using the desktop version of Taxapad 2012 (Yu et al., 2012). Given the absence of searchable taxonomic databases for predator species, no synonyms were considered for this particular natural enemy guild. For each of the S. frugiperda parasitoids, we logged the number of known plant x animal interaction records with either of the 25 legume species. As such, we methodically assessed whether baseline ecological insights in terms of food web linkages or interaction records are available to support S. frugiperda biological control – using a particular NE – in sorghum or maize intercropped with legumes.
Next, systematic literature searches were carried out using Web of Science (WoS). The WoS Core Collection database, which covers a 1900–2023 timespan, was queried using a University of Queensland staff subscription during May 15–30, 2023. Three different literature queries were performed. A first query aimed at gauging the number of studies considering key regulating ecosystem services (ES; i.e., soil stability, plant nutrition or nitrogen fixation, weed control, and biological control) for each legume species, when used in crop diversification schemes. A matrix of literature searches was thus conducted based on legume species x ES. The following search string, specifically geared towards blackgram effects on soil stability, was used and adapted for each given ecosystem service: TS = ((“Vigna mungo” OR blackgram OR “black gram”) AND (“inter-crop*” OR “cover-crop*” OR intercrop* OR covercrop* OR “crop diversification”) AND (“soil erosion” OR “soil conservation” OR “soil loss” OR “runoff” OR “soil stability”)). For the remaining ecosystem services, the following keywords were used: nutrient cycling, plant nutrition or nitrogen; weed control, weed management, weed suppression or soil cover; biological control, biocontrol, biological pest control or natural enem*. One-way analysis of variance (ANOVA), followed by a Tukey post-hoc analysis, was used to compare the number of studies between the four ES. A second query aimed at assessing the extent of scientific attention to a given natural enemy with regard to biological control in either maize or sorghum systems. For the particular case of Aphelinus albipodus (an aphid parasitoid associated with cowpea, faba bean and pigeon pea), the following string was used: TS = (((maize OR corn OR “Zea mays”) AND (“biological control” OR biocontrol OR “biological pest control” OR “natural enem*”) AND (“Aphelinus albipodus”))). This string was duly modified to cover all other NEs and to also gauge the related scientific output for sorghum. Other commonly intercropped cereals such as wheat or oats were not considered. A third query was specifically geared towards assessing the number of studies of a given natural enemy in cereal x legume systems, covering all species within the legume panel, their associated NEs and maize or sorghum focal crops. Hence, the above search string for A. albipodus was expanded by adding in the common and scientific names of a particular legume e.g., (“Vigna mungo” OR blackgram OR “black gram”). The resulting publication titles and abstracts were then manually screened to assess their actual coverage. The number of scientific publications that were obtained through each of the WoS queries, eventually filtered through manual screening, served as a proxy of global scientific attention.
Building on weak foundations
Our panel of 25 legume species comprised several popular inter- or cover-crops such as pea, cowpea, groundnut, Sunn hemp, faba bean, common bean or pigeon pea (Table 1). Within this panel, four key regulating ecosystem services received differing levels of scientific attention (ANOVA, F3,96 = 12.108, p < 0.01). Plant nutrition proved the most commonly investigated ES across legumes, featuring in 173.5 ± 244.9 (mean ± SD) scientific publications per species (Figure 1). This compares to a respective 31.4 ± 53.8, 23.5 ± 45.0 and 11.2 ± 17.4 publications for weed control, soil stability and biological control. Four legumes i.e., soybean, pea, cowpea and groundnut receive the bulk of studies from an agronomic angle, while pigeon pea and velvet bean were also commonly investigated for weed control. Biological control received merely 6.4% of the total studies in the plant nutrition domain, with 32% of legumes featuring in 10 or more all-time publications on this topic.
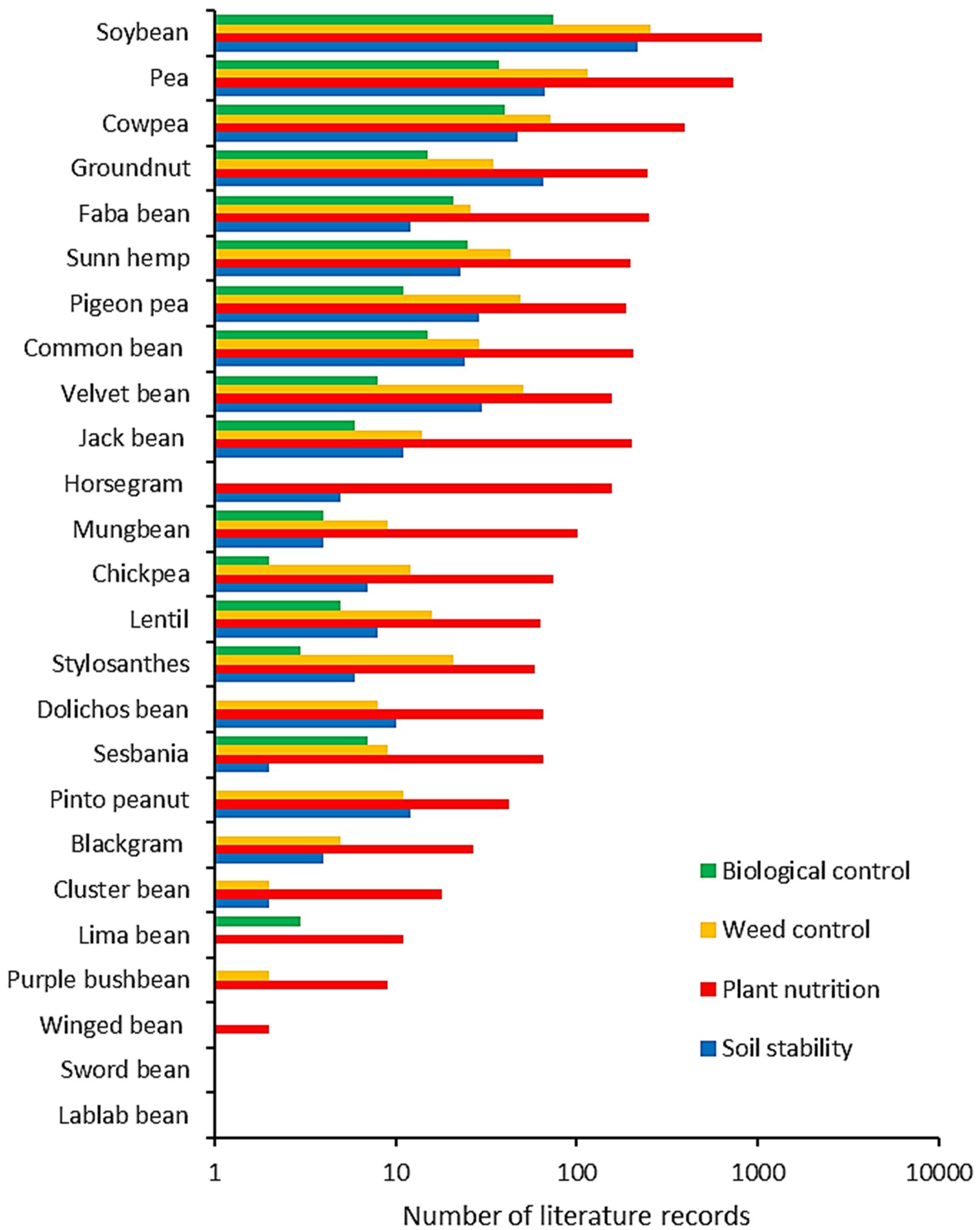
Comparative extent of scientific attention to regulating ecosystem services (ES) in legume-based inter- or cover-cropping systems. For 25 legume species that are commonly included in diversification schemes, we plot the number of all-time peer-reviewed publications regarding specific ES – as logged through systematic Web of Science queries. Patterns are shown for the following ES: biological control, weed suppression, plant nutrition or nitrogen fixation, and soil stability or erosion prevention. Data are plotted on a logarithmic scale. Lablab bean refers to Lablab niger i.e., the outdated name of Lablab purpureus (dolichos or hyacinth bean).
Number of arthropod biological control agents with known interaction links to 25 commonly cultivated legumes, as assessed using the GloBI interface. The 12 legume species with most species interaction records are highlighted in gray. Per legume, the number of parasitoid or predator species and families is listed.
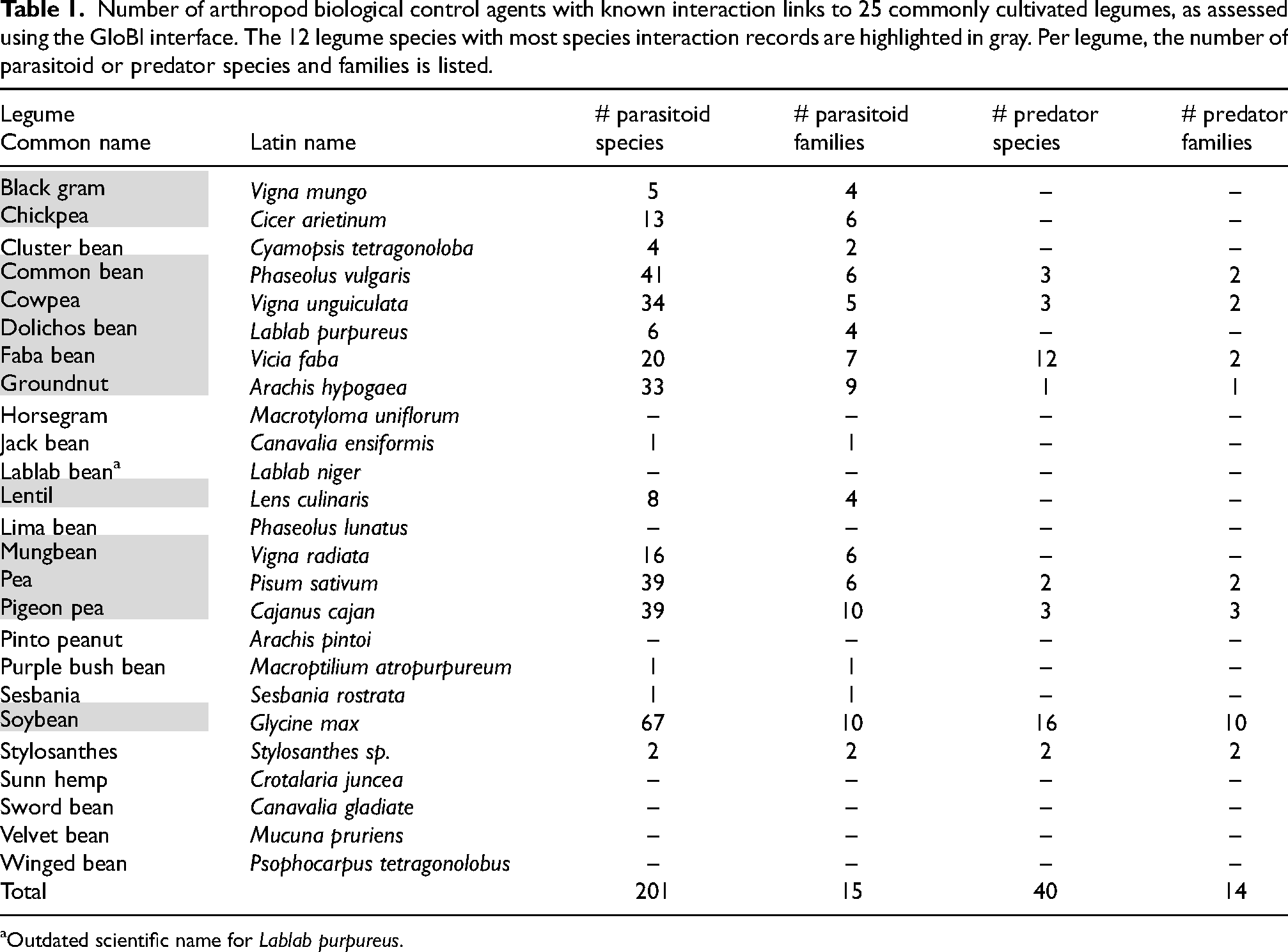
Outdated scientific name for Lablab purpureus.
Across all 25 legume species, interactions were logged for a total of 241 natural enemy species i.e., 201 parasitoids and 40 predators (Table 1). The most common parasitoid families are Eulophidae (n = 56 species), Pteromalidae (36) and Aphelinidae (31), while no records were found for parasitoids such as Aphidiinae. Parasitoids with interaction links to a wide set of legume species include the bean weevil parasitoid Dinarmus basalis (n = 10 legumes), the polyphagous egg parasitoid Trichogramma chilonis (n = 8) and the granary weevil parasitoid Anisopteromalus calandrae (5). This skewed attention to (certain groups of) parasitic hymenopterans possibly can be explained by a relatively high share of flower visitor or pollinator surveys (vs. predation records) in the GloBI database. This may also explain a heightened coverage of predaceous hoverflies (Syrphidae; n = 11 species) i.e., the world's second most important pollinators after bees (Rader et al., 2020). Though the particular morphology of legume flowers may deny non-bee visitors the access to nectar (Wäckers and Van Rijn, 2012), hoverflies and minute parasitic wasps may still access floral rewards e.g., through biting holes of nectar robbers or by accessing extrafloral nectaries. Besides hoverflies, there are only six species records for ladybeetles (Coccinellidae), seven for assassin bugs (Reduviidae), three for damsel bugs (Nabidae) and single records for three spider families i.e., Anyphaenidae, Clubionidae, Thomisidae. Meanwhile, merely one or two records are found for common natural enemies such as ants, flower bugs (Anthocoridae), thread-waisted wasps (Sphecidae) or flower beetles (Anthicidae). The paucity of predator data is surprising, as legumes’ extra-floral nectar is readily consumed by a wide variety of vespid wasps, spiders, carnivorous mites, bugs or beetles (Heil, 2015) and several of those predators also frequently visit flowers (Nyffeler et al., 2016). Ants, widely seen as the most abundant and voracious predator in tropical farming systems, only feature in interactions with two species of the legume panel i.e., common bean and soybean in GLoBI. The near-absence of predator records also points to methodological shortfalls. For instance, at least six ladybeetle species and 10 species of vespid wasps are known to feed upon herbivores of faba bean (Nuessly et al., 2004) and the predatory mirid Macrolophus pigmaeus consumes its extra-floral nectar (Portillo et al., 2012). Yet, as none of these feature in GloBI, the database likely only contains a fraction of known interaction records. Also, several popular legume intercrops bear no known (or recorded) interaction links with natural enemies. These include the pinto peanut Arachis pintoi i.e., a popular cover crop in tropical citrus orchards or palm plantations. By offering honeydew as well as plant-provided floral and extrafloral nectar (Wäckers, 2005), legumes such as A. pintoi likely increase NE activity-density in those settings. Yet, as none of these patterns can be deduced from GloBI-derived data, it is questionable if the database -in its current form- can adequately inform biological control science and practice.
For 12 legume species with most NE interaction records, only 12.9 ± 8.3% (range 0–25.0%) and 29.8 ± 16.4% (range 0–61.5%) of associated organisms are relevant to biological control in either sorghum or maize respectively. In absolute terms, soybean, pigeon pea and faba bean are associated with most organisms relevant to biological control in either cereal (Figure 2). This compares to lentil, black gram or dolichos bean, for which the associated biota have not or rarely been studied through a biological control lens. Meanwhile, chickpea and mung bean consistently harbor the highest fraction of associates with relevance for biological control in either cereal. Taxon-specific scientific attention however is shallow, with 3.4 ± 1.6 and 10.2 ± 5.3 publications addressing the contribution of a given legume-associated NE to biological control in either system (Figure 2). Chickpea and groundnut associates receive relatively more attention, while scant scientific insights have been gained on legumes such as pea, common bean or faba bean. Common bean is regularly integrated with sorghum or maize (Namatsheve et al., 2021) e.g., in drought-prone areas of Central America. Yet, only a small slice of its known NE associates have been studied from a biological control perspective – merely featuring in a respective 7.4 ± 4.9 or 7.6 ± 2.6 all-time publications. When homing in on biological control within cereal x legume intercrops in particular, there is a near-total absence of scientific output. As little as three biological control publications feature legume-associated NEs i.e., Trichogramma chilonis in cowpea x sorghum or pigeon pea x sorghum or Orius insidiosus in soybean x maize intercrops. Though legume-associated NEs such as Trichogramma ostriniae, Harmonia axyridis or Trichogramma pretiosum receive ample attention as biological control agents in sole maize or sorghum crops, whether and how they add (or not) to natural pest control within the related cereal x legume intercrops is shrouded in mystery. For legumes such as common bean, mung bean or faba bean, there is either a lack of knowledge on their association with key natural enemies of cereal pests and/or on the contribution of those biota to biological control in intercrop systems. Patterns are equally bleak when examining known legume associations among the global parasitoid community of S. frugiperda – a pest of global allure which reportedly inflicts US$ 9.4 billion losses per year in Africa alone (Kenis et al., 2022) (Figure 3). Out of a complex of 167 hymenopteran or dipteran parasitoids (as recorded at the species level), legume associations are only known for four species i.e., T. chilonis (8 associated legumes), T. pretiosum (4), Trichogramma minutum (3) and Trichogramma chilotraeae (1). Hence, for the remaining 161 species, there is no information on their eventual (trophic, non-trophic) association with any of the 25 legume species (Figure 3). Hence, whether the tens of effective egg-larval, larval and pupal S. frugiperda parasitoids interact with legumes remains utterly unknown. Considering how parasitoids such as Cotesia marginiventris, Chelonus insularis or Aleioides laphygmae provide up to 92% natural pest control in diversified maize crops (Meagher et al., 2016), it is puzzling how the potential benefits of legume integration are roundly overlooked. Predators, as prime S. frugiperda biological control agents, do not fare better. Pioneering work in the 1980s showed how S. frugiperda mortality is substantially higher in maize x bean intercrops than in maize monocrops where Ectatomma ruidum ants were excluded (Perfecto and Sediles, 1992), but whether or how legume x ant interplay mediates these dynamics remains unclear. Equally, Aphidiine parasitoids consume extrafloral nectar from legumes such as faba bean (Jamont et al., 2013), but whether this translates in enhanced aphid biological control in intercrop systems is unknown. Lastly, while polyphagous ladybeetles such as Harmonia axyridis or Coccinella septempunctata could benefit from access to alternative prey (e.g., Aphis craccivora on cowpea or groundnut; Obrycki et al., 2009), this habitat management tactic remains virtually unexplored in intercropped cereals.
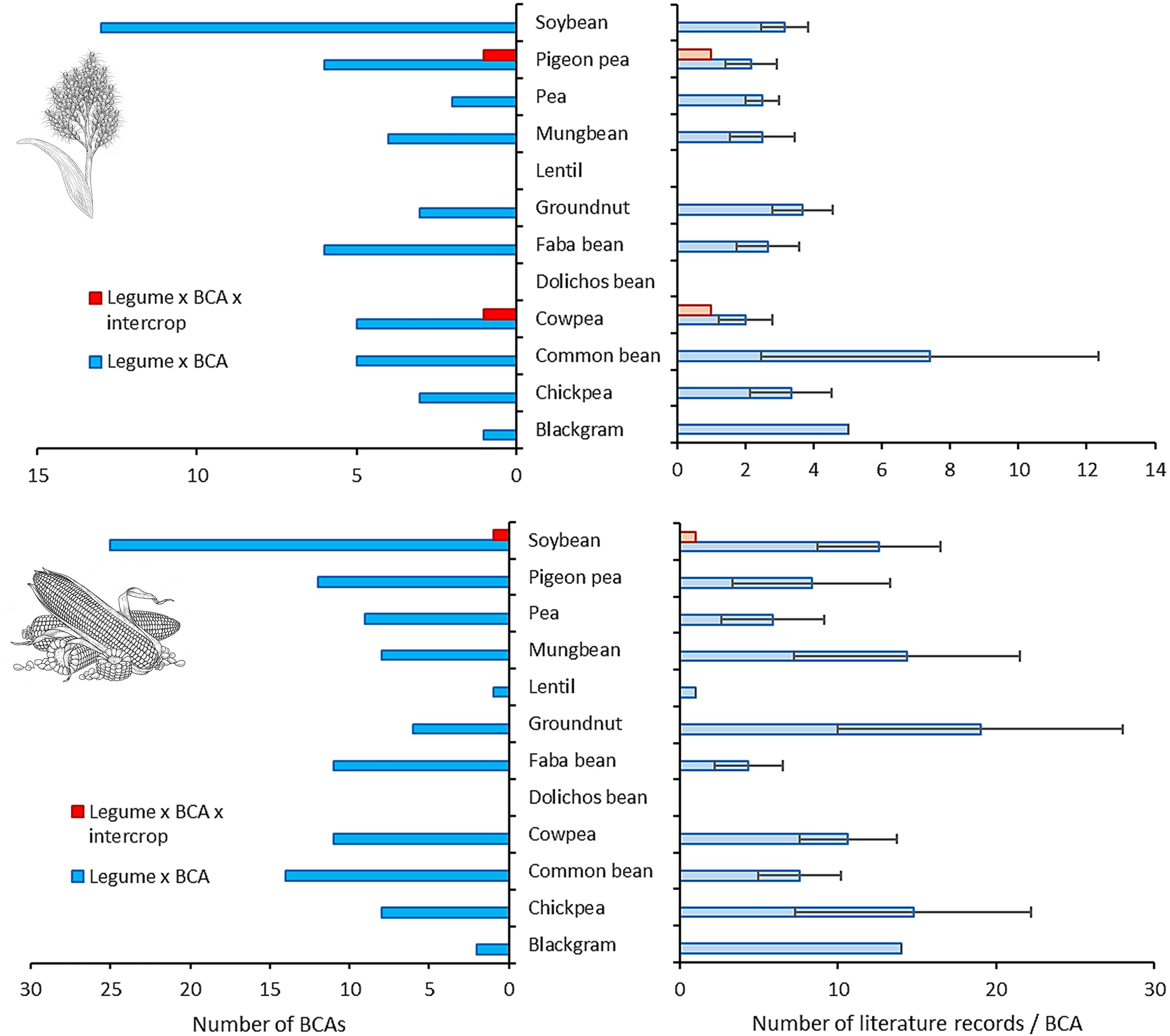
Extent of scientific attention to legume-associated biological control agents (BCAs) and their role in sorghum- or maize-based intercropping systems. Data are plotted for sorghum (top) and maize (bottom) focal crops, and reveal patterns for 12 commonly intercropped legume species. For each legume species, BCAs are identified by screening (plant x animal) species interaction records in GloBI (Table 1). The left panel shows the number of legume-associated BCAs that feature in any peer-reviewed scientific publication related to biological control in either focal crop i.e., sorghum or maize (blue bars) and the respective legume-cereal intercrops (red bars). The right panel depicts the average (mean ± SE) number of all-time scientific publications related to a given BCA in each focal crop and its respective legume-cereal intercrop. Crop drawings obtained from www.freepik.com.
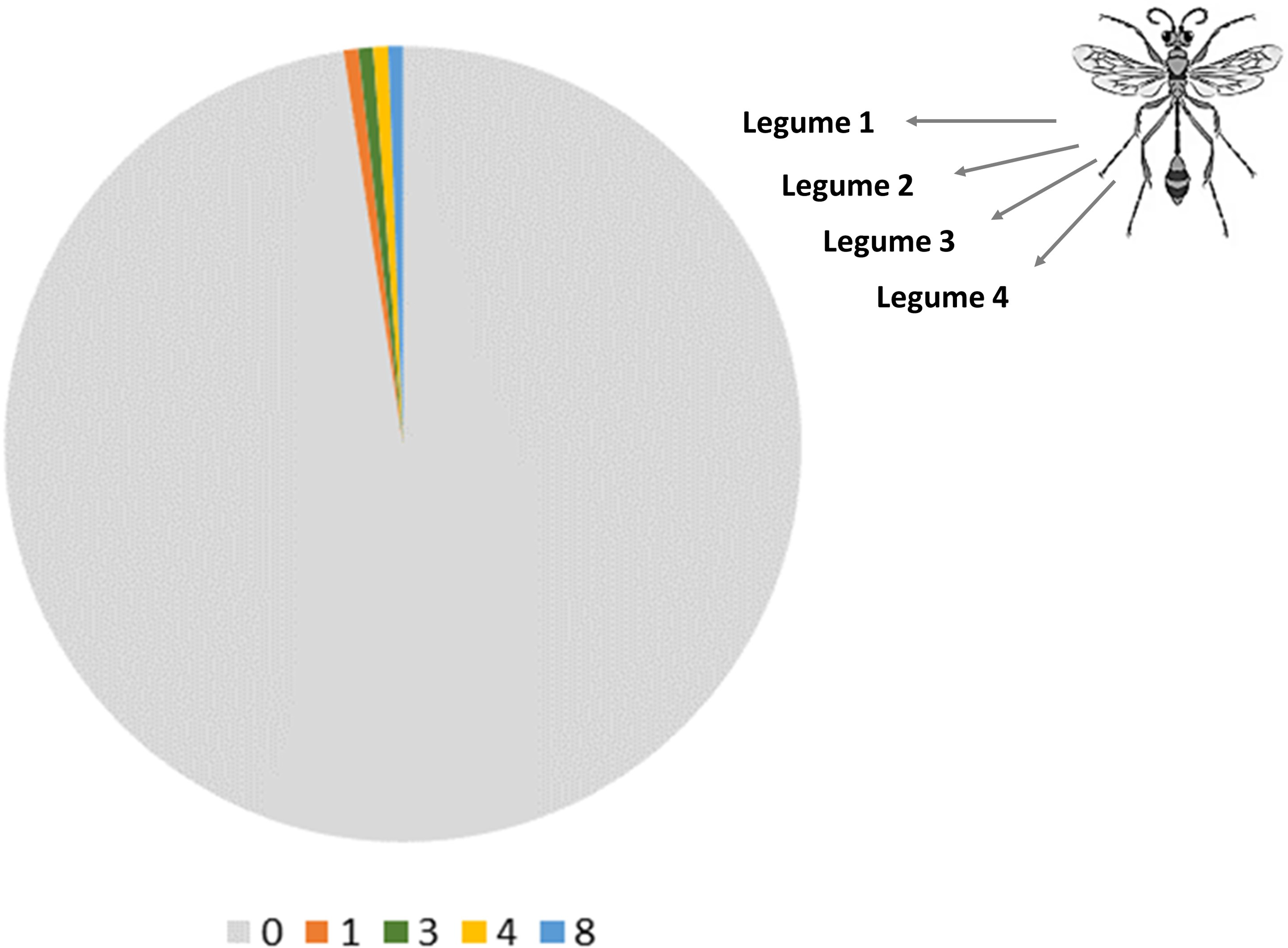
Share of the global Spodoptera frugiperda parasitoid community with known interaction linkages to a panel of 25 cultivated legumes. The pie chart depicts the relative share of 167 dipteran or hymenopteran S. frugiperda parasitoids that exhibit interaction records with zero, one or more cultivated legumes, as logged through GloBI (www.globalbioticinteractions.org). Legumes comprise 25 plant species that are extensively used for crop diversification in the tropics or sub-tropics (Table 1). The chart covers all known parasitoids of S. frugiperda in its native (i.e., Neotropics) and invasive range i.e., Africa, Middle East and Asia as per Kenis et al. (2022) and Molina-Ochoa et al. (2003). Wasp drawing from www.freepik.com.
Though maize or sorghum are globally grown on approx. 240 million ha and increasingly paired with legume intercrops, arthropod biological control in those systems evidently lacks vital scientific underpinnings. We do not argue that legume integration for biological control is not progressing; insect-mediated ecosystem services are comparatively well-studied in agricultural settings as compared to those in natural environments (Noriega et al., 2018) and habitat management has grown into a full-fledged scientific discipline (Gurr et al., 2017). Yet, emphasis of the latter is often placed on off-field features such as flower strips or beetle banks (Albrecht et al., 2020) and not necessarily tuned to field-level adaptations such as inter- or intraspecific crop diversification (González-Chang et al., 2020). Equally, as biodiversity and food web structure jointly shape biological control in farm settings (Yang et al., 2021), our inability to record the most basic inter-organismal linkages ultimately transpires in a failure to tap legumes’ potential for sustainable crop protection (Bianchi, 2022; Cappelli et al., 2022). Indeed, how do we ever intend to engineer plant communities to maximize ecosystem service delivery (Storkey et al., 2015) if we don't know how individual plant species interact with ecosystem service providers i.e., BCAs. By thus failing to uncover its ecological underpinnings, biological control scientists are prone to get stuck in its actual case-by-case empiricism (Gurr et al., 2017), prove unable to gain traction with growers and conservationists (Chaplin-Kramer et al., 2019) and ultimately find themselves unsuccessful in attaining desired social-ecological outcomes at scale (González-Chang et al., 2020). Those shortfalls are apt to be exacerbated in the Global South (Steward et al., 2014). Not only are insect biodiversity discovery and description lagging in the tropics (Eisenhauer et al., 2019; Srivathsan et al., 2023), but most plant x invertebrate functional linkages and their underlying traits most certainly wait to be uncovered (Moretti et al., 2013). Other limiting factors are the lower likelihood of actual publication and the lower visibility of scientific outputs – with the latter mirrored in the omission of popular bibliographic databases such as Scielo in WoS Core Collection (Fox et al., 2023; Gomez et al., 2022). As a result, ecological interaction data may often be stuck in the gray literature or in published manuscripts, and biological control scientists appear to (blissfully) discount their massive potential. Hence, unless this gap is filled, we remain far removed from generating scientifically-grounded mechanistic principles to guide the design of pest-suppressive systems in the Global South (Bianchi, 2022; Wyckhuys et al., 2022). Indeed, the development of more predictive e.g., trait-based approaches to anticipate biological control strength and stability under different legume intercrop arrangements seems a far-off goal. A more attainable goal is to methodically assess the biological control outcomes in intercrops that have been designed for agronomic objectives, to then use those findings to build broader momentum for ecological intensification practices. Overall, unless one opts to “ride the intercropping wave” without consciously steering biological control science towards this domain, thorough work is instantly needed to fortify its very (scientific) foundation.
Shifting gears: from blissful ignorance to remedial action
Legume integration carries extraordinary promise to attain ecological intensification of agriculture and put in place low-emission, climate-resilient agri-food production systems (Foyer et al., 2016; Jensen et al., 2020; Snapp et al., 2019). Evidently, legumes are well-appreciated from an agronomic angle as they fix nitrogen, stabilize soil or enhance carbon sequestration. As a consequence, they have widely been investigated in those disciplinary domains or “silos.” Yet, unless we can build a robust evidence base and related decision rules on how legume integration -or, more broadly, crop diversification- contributes to diverse ecosystem service “bundles” (Couëdel et al., 2019; Tamburini et al., 2020), achieving their wide-scale adoption may become a predicament. This Perspective piece argues that legume integration for arthropod biological control finds itself on a particularly weak scientific foundation. This, however, may not be different than for other insect-mediated ecosystem services such as pollination (Otieno et al., 2020). Individual case studies e.g., in Brazilian Inga sp. x coffee systems build on extensive ecological insights and show how legumes are pivotal to achieving pest-suppression in polycultures. Some precaution may be needed as these and other results can be highly contextual and their validation in distinct (social-ecological) production contexts is lagging. Though such is recognized by biological control scientists and practitioners, (globally) coordinated research thrusts to fill these taxonomic, biological and ecological knowledge gaps are few (but see Perović et al., 2018). Hence, to avoid that biological control science continues to yield serendipitous outcomes, we argue that remedial action on a number of fronts is in order.
Strengthen biodiversity and biostructure databases
Irrespective of the core role of agro-biodiversity (Dainese et al., 2019), biostructure i.e., the intricate architecture of inter-organismal interactions is what ensures proper ecosystem functioning and what underpins biological control in farming systems (e.g., Yang et al., 2021). As elegantly phrased by McCann (2007), overlooking the role of biostructure is analogous to cataloging body parts without recording the overall anatomical structure in the animal physiology domain. Biodiversity is the central pivot of biological control science (González-Chang et al., 2020), but biostructure puts biodiversity to use i.e., by anchoring ecosystem processes, functions and the ultimate service of natural pest control. Biostructure may also enable farmers to perceive and appreciate the role of biodiversity. Scientists however tend to ignore the role of biostructure, perhaps because it is comparatively easy to count species (McCann, 2007) – especially in above-ground realms (Wyckhuys et al., 2021). Clearly, as extensive as the GloBI database is, its current version contains surprisingly few relevant legume x NE interactions; its reliance upon open-access data and volunteer contributions plausibly compromises its completeness. As illustrated above, even for popular intercrops such as faba bean, tens of known NE linkages are missing. Hence, first and foremost, baseline data on the interactions between NEs and (legume) plant species either need to be gathered or consolidated in (open-access) databases. In the Global South, absence of these interaction records reflects the paucity of data on invertebrate biodiversity - which can only be filled through additional discovery and description. In the Global Biodiversity Information Facility (GBIF), only 0.17 G of the 2.16 G records i.e., 7.9% pertain to arthropods (Srivathsan et al., 2023). Though sufficient data may already be available in Europe or North America, our work demonstrates that these are not necessarily accessible at scientists’ fingertips. Hence, in the Global North, mobilizing, archiving and securing the interoperability of existing data is needed (Eisenhauer et al., 2019). The GloBI database offers a uniquely suitable portal to offer biostructure data and added funding can help to ensure the completeness of its records. Meanwhile, text mining and natural language processing can be used to build structured databases from texts in the grey literature, websites or primary journal articles (Farrell et al., 2022). Equally, tools such as iNaturalist or Web of Life are also capable of recording interactions and other interaction databases (mangal.io) can be queried through its Application Programming Interface (API). If one can thus gather sufficient data and organize them into comprehensive, unbiased large-scale public databases, these may catalyze scientific progress in a similar way as in genomic or viral discovery arenas (Edgar et al., 2022). Indeed, only when such databases are in place can we expect to see major advances in the field of biological control (Segoli et al., 2023).
Broaden the scope and organismal focus of future experiments
For the few intercrop systems where data is available (e.g., soybean and perhaps common bean), efforts are needed to move beyond the initial stages of small-scale experimentation, and to advance towards a greater understanding of biodiversity x ecosystem function (BEF) interplay globally (Gonzalez et al., 2020). Also, as adoption is slow, farmer knowledge, attitudes and perceptions are insufficiently integrated (Nicholls and Altieri, 2018). More holistic “systems” perspectives couple the social and natural facets of legume-based intercrops and may provide further opportunities to unveiling the mechanisms behind pest suppression. This possibly can entail the establishment of networked trials and an integration with remote sensing. In ongoing efforts such as FAO's Global Trial on S. frugiperda, data from diversification trials can be linked to biodiversity, landscape composition, land-use change, soil fertility or agrochemical pollution layers (Tang and Maggi, 2021). Doing so can expand the knowledge base and guide future work in other intercrop systems. Clearly, the scope of consideration and organismal focus also needs to broaden beyond parasitic Hymenoptera i.e., which constitute 85% of recorded legume associates in our assessment. Generalist predators are effective biological control agents in agricultural settings and positively benefit from diversification (see Puliga et al., 2021; Rivers et al., 2017; Symondson et al., 2002), though certain species are discouraged by the more complex crop architectures of diversified settings (Gontijo et al., 2018). As an added advantage, large bodied predators such as carabid beetles or ants can be voracious weed seed predators e.g., as demonstrated in oat x pea systems (Ward et al., 2011). Hence, for a given legume intercrop, natural enemy abundance, foraging behavior, (multi-target) biological control efficacy and traits of the focal and (legume) companion plants are to be thoroughly assessed in sync.
Seek to anticipate pest suppressiveness
Once such data are in place, a transition from a simple logging of patterns to gaining a quantitative understanding of pest suppressiveness, presents itself as the logical next step (Bianchi, 2022; Brooker et al., 2021). Though underrecognized, legume integration may confer embedded complexity and internal resiliencies in food production (Jahn et al., 2018), which could allow intercropped systems to swiftly adapt to varying (a)biotic conditions e.g., drought or pest outbreaks (Renwick et al., 2020). Trait-based approaches carry vast potential to generate universal “rules of thumb” for crop diversification (Perović et al., 2018) and to generate inferences regarding pest suppressiveness of particular arrangements. Flower traits such as blooming phenology, attractiveness, nectar availability and nutritional quality are already widely used in pollination ecology studies (but, see Fiedler and Landis, 2007). So far, trait-based ecology approaches for ecological intensification revolve around plant x plant interactions yet this should not continue to be the case. Full integration with animal traits and abiotic parameters is paramount, more so as plant traits alone act as poor predictors of biodiversity-mediated effects on ecosystem services such as biological control (Van der Plas et al., 2020). Yet, any attempt to infer ecosystem function through trait-based approaches in the absence of a comprehensive logging of biostructure i.e., by skipping a critical step in the BEF continuum may very well prove futile.
Dismantle disciplinary silos
Agriculture-related scientific disciplines cannot all orbit in distinct planes. Actually, trans- or inter-disciplinary science is crucial to resolving global change and to mitigating the externalities of the present-day food system (Vandermeer et al., 2018). Pest management professionals in particular should strive to adopt holistic, system-level perspectives (Wyckhuys et al., 2023), taking into account below-ground realms, soil fertility, farmer knowledge and attitudes, or landscape-level dynamics (Tscharntke et al., 2021; Wyckhuys et al., 2017, 2019, 2021). The societal benefits of (legume-assisted) biological control are indeed huge, yet this is rarely apprehended. In light of the bonanza of scientific outputs on legumes’ renowned agronomic benefits, we have barely scratched the surface regarding their contribution to (pest, pathogen; above- or below-ground) biological control. Meanwhile, the often excessive use of synthetic pesticides in legume-based systems across the globe hits farmers with a double whammy. By inflicting mortality in (bee, non-bee) pollinators and natural enemies alike, farmers suffer pollination deficits and degrade natural pest regulation (e.g., Saqib et al., 2020), thus deepening pest issues. Contrariwise, by cutting pesticide inputs and making smart use of natural biological control, one could restore pollinator-induced fruit set to the tune of 20–90%, 10–100% and up to 60% in cowpea, pigeon pea or common bean, respectively (Otieno et al., 2020).
Pollination ecologists have acted swiftly on the above by targeting legume mixtures. As legume intercrops have proven equally attractive to pollinators as sole crops, they offer clear “win-win” options for yield gain and pollinator conservation in diverse landscape matrices (Cole et al., 2022; Kirsch et al., 2023). Frameworks have equally been built to assess trade-offs between legume yield and pollinator conservation (Knapp et al., 2022), though the emphasis is -rather counterintuitively- placed on insecticide-based measures instead of biological control. New vistas also open up in breeding or crop improvement domains. In view of an envisioned agroecological transition, breeding legumes apt for intercropping or with facultative outcrossing plus pollinator-attractive floral traits, have all become top goals (Bailes et al., 2018; Rubiales et al., 2021). Yet, as above, while superior yield and (biotic) stress tolerance have become routine breeding targets, breeding for biological control has not (Stenberg et al., 2015). Given the massive externalities of global pesticide use, one comes to wonder whether (legume) breeding will continue to be performed under a “pesticide umbrella” with total disregard of prime ecological processes (Bernal and Medina, 2018). Or will legumes’ unique functionalities for natural pest and disease regulation finally be embraced, and (natural) biological control effectively deployed to replace chemical means of crop protection. Indeed, so much is to be gained if pollination ecologists, biological control scientists and breeders could join hands. Equally, engagement with social scientists is crucial to ensure that legumes’ contribution to biological control is known, valued and sought after by end-users. Along the same vein, biological control scientists encounter fertile ground for cooperation with agronomists or plant pathologists, in the co-design of legume intercrops. On both fronts, scientists are increasingly challenged to “reinvent and redefine” their fields, as interdisciplinary or integrative disciplines, drawing others into their orbits to work towards common goals (Struik and Kuyper, 2017). Why shouldn't biological control scientists make the first move? What might be a small step for this thriving discipline could very well alter the course of global agriculture.
Footnotes
Acknowledgements
This work was funded by the European Commission through project GCP/GLO/220/EC and executed by the United Nations Food and Agriculture Organization (FAO). We wish to thank Gabor Pozsgai for providing helpful comments on an initial draft of the manuscript.
Author contributions
KAGW led the idea generation, writing, analytics and editing process. MGC extracted and organized data from GloBI. BARH provided the funding. All authors revised the text and provided editing suggestions.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: KAGW is the chief executive officer of Chrysalis Consulting – a firm which provides tailored support to nature-friendly farming and biological control.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the FAO (grant number GCP/GLO/220/EC).