
Abstract
There is increasing demand for farmers to reduce their use of anthropogenic inputs such as fertilizers and pesticides. It is often suggested that nature offers alternative ways to suppress pests, but what might these be? One prominent difference between most natural and agricultural systems is the diversity of plants and associated biota such as arthropod herbivores, predators and parasites, both above- and below-ground. It is often claimed that a move away from monocultures and towards greater field, farm and landscape diversity could be an important and obvious first step towards more natural pest control. This paper draws together strands of evidence on what a move to more diverse crop systems can and cannot bring to pest control. We conclude that diversification has great potential, but is not a silver bullet. Adopting a wider definition of IPM and including the environmental stewardship role of agriculture would enable a greater use of natural mechanisms for pest control, including a greater role for diversity, but would also require an economic mechanism to reward stewardship that serves wider societal needs while discouraging management that does not.
Keywords
Introduction
Modern agriculture relies strongly on anthropogenic inputs: fossil fuels, industrially fixed nitrogen fertilizer, and manufactured chemicals such as pesticides and plant growth regulators. The use of these inputs has resulted in tremendous increases in agricultural productivity, and has been strongly promoted by policies aiming to improve food security (Connor et al., 2011). The success of agricultural intensification in increasing productivity is overwhelming and has been well documented (Tilman et al., 2002). The uptake of intensive management practices has also resulted in scale enlargement in farming and changes to farming landscapes including lower crop diversity, removal of semi-natural habitats, increased crop density, and an increase in the number of crops grown per season.
The manufacture and use of anthropogenic inputs such as fertilizers result in spill-overs and costs, both in- and off-field including emission of CO2 into the atmosphere, loading of the geosphere with additional reactive nitrogen (i.e. N in other forms than N2), emissions of the greenhouse gas N2O, soil acidifying NH3 (Fangmeier et al., 1994) and nitrate (NO3−) runoff into surface water (Kaushal et al., 2011). The use of pesticides results in on-field and off-field effects on non-target organisms, including a very wide range of insects, with downstream consequences for the whole farmland agro-ecosystem and food web (Barmentlo et al., 2021; Hallmann et al., 2014). As such, pesticide use has been implicated in the demise of insects (Wagner et al., 2021), and may pose health risks for farmers (Jokanovic, 2018) and consumers (Forkuoh et al., 2018), while the profitability of their use is being questioned (Mourtzinis et al., 2019; Zou et al., 2019).
Organic agriculture (OA) has its roots in the late 19th century and aims to use natural means for managing crops (though fossil fuels and some pesticides of natural origin are acceptable). OA can be productive, but there is a consensus that it is less productive than conventional agriculture (Seufert and Foley, 2012). There is active debate whether OA, if adopted worldwide, would be able to meet world food demand (Connor, 2021), depending on human diets and the use of crop output for animal feed (Badgley and Perfecto, 2007). Nevertheless, OA has a strong foothold, and consumers who pursue a healthier diet with less meat and more pulses may further stimulate its growth. Here we do not focus specifically on OA, but rather on which ‘ecological intensification measures’ (Bommarco et al., 2013) can be taken at crop and landscape (farming system) level, in both organic and conventional systems, to reduce the impacts of pests and diseases on crop production.
First of all, we should ask whether diseases and pests impact plant communities in natural systems. They certainly do. For instance, soil borne pathogens have been shown to be key drivers of plant succession and productivity in grassland biodiversity experiments (Ampt et al., 2019; Mommer et al., 2018; Van Ruijven et al., 2020). With a greater number of species, average productivity increases. The connection between diversity and productivity has also been made in the Cedar Creek experiments reported by Tilman et al. (2014) and in the Jena experiment (Buchmann et al., 2018). Biodiversity effects on productivity increase over time (Thakur et al., 2021). Within mixed species plots, the abundance of different species varies over time and the decline of species is associated with increases in their species-specific pathogens (van Ruijven et al., 2020). Thus, the biodiversity effect in these trials is predominantly related to the accumulation of species-specific plant pathogens, particularly if the original stand is comprised of only a single species.
Like plant pathogens, arthropod herbivores may alter plant communities by reducing the biomass and reproduction potential of the plants on which they feed. Herbivory may promote plant species coexistence by weakening strongly competing plant species (Mortensen et al., 2018), while specific herbivore feeding preferences can also lead to increased dominance and decreased plant diversity (Olff and Ritchie, 1998). Indeed, even though natural ecosystems are often composed of diverse plant communities, there are notable exceptions, including natural stands of the wild ancestors of rice, wheat, and sorghum, which can be considered natural monocultures (Wood and Lenné, 2001; Wood and Lenné, this volume). Whether or not results obtained in natural systems can be mimicked in agricultural systems is contested (Denison and McGuire, 2015; McGuire, 2021).
It was established long ago that crop rotation can be used to avoid the build-up of species-specific plant pathogens. This understanding preceded the awareness among ecologists of the importance of negative plant-soil feedback in natural systems. Along the same lines, the large-scale planting of a single crop can be conducive to the spread of pathogens and pests (Thomine et al., 2021). This can give rise to trade-offs because good growing conditions or high crop prices of may stimulate farmers to increase the acreage and/or shorten crop rotations, which makes these cropping systems more susceptible to pests and disease (Meehan et al., 2011; Skelsey et al., 2010).
Vulnerability of cropping systems has often been addressed by the use of pesticides, but the negative side effects of frequent pesticide use are increasingly recognized and there is a strong call for more sustainable pest management alternatives (Bakker et al., 2020; Jepson et al., 2014). This call prompts the question: what we can learn from nature to reduce the use of and reliance on pesticides? Here we first discuss disease and insect pest suppression in diversified cropping systems at the field and landscape scale. We then discuss why farmers use insecticides and why agricultural scale enlargement and intensification has led to lock-in situations. We end with a discussion on options for the future.
Disease suppression in intercrops and variety mixtures
Diseases, pests and weeds may be suppressed by mixing different species in an intercrop (Trenbath, 1993; Boudreau, 2013; Gu et al., 2021a,b; Liebman and Dyck, 1993; Zhang et al., 2019) and by combining several varieties of a single species (Kiaer et al., 2009; Kiaer et al., 2012; Reiss and Drinkwater, 2018; Kristoffersen et al., 2020; Tooker and Frank, 2012). Reviews published before 2000 on the suppressive effect of species and variety mixtures on pests, diseases and weeds are most often narrative reviews, which can be critiqued as ‘potentially biased’, as the selection of sources is at the discretion of the author(s) (although see Smithson and Lenné (1996) for comprehensive tables with effect sizes). ‘Vote counting’ is a type of analysis that provides information on the frequency of suppressing effects across studies, but it does not quantify the average size or variability of effects. This approach has been used to tally how many studies report significant reductions in the abundance or impact of pests, diseases or weeds (Liebman and Dyck, 1993; Stomph et al., 2020).
The gold standard for overarching synthesis is generally considered to be meta-analysis, which includes a systematic survey of the literature to identify all relevant papers on a topic, followed by a statistical analysis of the effect sizes reported in those papers that meet inclusion criteria for relevance and quality (Koricheva et al., 2012; Makowski et al., 2019). Meta-analyses confirm that pest and disease reduction is common in intercrops and variety mixtures. For example, Zhang et al. (2019) estimated that in mixtures of wheat and faba bean, diseases were suppressed by on average by 33%, with the greatest reduction in the early stages of outbreaks. The incidence of mildew on wheat increased with N fertilizer, but intercropping provided consistent levels of disease suppression across all levels of fertilizer. Intercropping and crop rotation of grain and legumes may also reduce damage from parasitic weeds (Rusinamhodzi et al., 2012).
Ongoing meta-analyses look further into the relationship between crop diversity and disease control. One ongoing study 1 provides a quantitative synthesis of data on the suppression of rice diseases in variety mixtures in which the component varieties have different levels of disease resistance. This study is almost exclusively based on data collected in China because of the large research effort there on disease control by crop diversity, following a landmark paper by Zhu et al. (2000). Mixing varieties reduces incidence of diseases by 57% on average, ranging from 46% in the case of leaf blast to 66% in the case of panicle blast (both caused by Magnaporthe grisea). The average control effect of mixture was greater for rice blast than for two other diseases (caused by Rhizoctonia solani or Xanthomonas oryzae pv. Oryzae). The incidence of rice blast increased with nitrogen fertilizer input in both pure and intercropped stands. Resistant and susceptible rice varieties differed greatly in disease incidence at low N input, but at high N input, this difference disappeared: in other words, resistant rice varieties became susceptible to rice blast at high N input. The disease control effect of variety mixtures was consistent across N fertilizer levels.
Another on-going meta-analysis 2 aims to syntheses worldwide data on the suppression of plant diseases in intercrops across a number of crop combinations. Identified studies include 14 crop species. The average level of disease reduction in species mixtures was 50%, with no difference across continents, and there were substantial differences in disease control between pathosystems (crop species-pathogen combinations). Disease suppression was greatest when the focal species was grown in single rows while the companion species was grown in strips, suggesting an important role for host dilution as a mechanism for disease control in mixtures. The lowest level of suppression was obtained when both species were in strips.
Grain yields under intercropping are on average 1.5 t/ha greater than the average of component crop yields when grown as monocultures (Li et al., 2020). These yield benefits are primarily due to improved resource capture (complementarity) but also to pest, disease or weed control if these are not otherwise controlled with pesticides. The magnitude of the effects of crop diversity on the severity and yield effects of pests, diseases and weeds depends on the production situation. Further work is needed to elucidate the contribution to yield increases of suppression of biotic constraints (pests, diseases and weeds) in intercrops and variety mixtures.
The field studies on disease control in intercrops and variety mixtures show that the suppressive effect is substantial and could make a practically relevant contribution to disease management. This may be so even if the effect on yield is smaller than that on disease incidence or severity, due to the non-linear relationship between disease severity or incidence and yield. Intimate mixture of the different species is not always required: Ditzler et al. (2021) show effective disease control with crops species grown in alternating strips 3 m in width. Studies also show that disease control is not complete, and in many cases substantially less than the level of disease control that farmers have become accustomed to when using chemical fungicides. Hence, crop diversification by intercropping and use of variety mixtures provides a useful element in the toolbox of IPM (integrated pest management), but it is not a silver bullet.
Several mechanisms account for the disease control effect of intercropping and variety mixtures. These include: host dilution, barrier effects, changes in microclimate and induced resistance, as well as physiological or morphological changes in intercropped plants as compared to plants growing in a pure stand. Host dilution results from reducing the density of susceptible host plants, such that spores dispersing from a host have a reduced chance of landing on another host plant (Skelsey et al., 2005). For as yet uninfected plants, host dilution means that there are fewer conspecific plants in their surroundings, which should dilute the number of disease propagules to which they are exposed. Barrier effects occur when a tall non-host plant in a mixture intercepts spores that would otherwise be captured by, and potentially infect, shorter plants. Changes in microclimate can be of critical importance for the ability of a pathogen to infect. In rice variety mixtures, single rows of a tall glutinous rice variety are alternated with narrow strips of hybrid varieties (Zhu et al., 2000). The glutinous rice is susceptible to rice blast disease caused by M. grisea¸ but in an intercrop canopy, the taller glutinous plants dry more quickly than in a pure stand due to higher exposure to radiation and wind. As a result, the period of leaf wetness can drop below the minimum needed for infection by the pathogen, resulting in a major reduction in the chance of infection. On the other hand, glutinous rice can act as a barrier for the hybrid rice pathogens. Thus, both varieties can benefit from disease control in this mixture (Zhu et al., 2000). Induced resistance occurs when an incompatible pathogen tries to infect a crop plant. Due to molecular detection mechanisms, the host plant may trigger defences (e.g. cell apoptosis) to prevent pathogen entry in a living cell, but this also triggers a systemic reaction in the plant not only to this incompatible pathogen but also to compatible pathogen types (Finckh et al., 2000). Finally, intercropping results in changes in the resource environment experienced by the plant as compared to the environment experienced in a pure stand. Due to developmental plasticity, morphological or biochemical changes occur, which can affect infection by pathogens.
Arthropod pest suppression in intercrops
The size of arthropod herbivore populations in agroecosystems can be influenced by the properties of plant communities (bottom-up effects) and by their natural enemies (top-down effects). At small spatial scales, plant diversity can influence herbivore populations via a suite of responses, including disruption of (specialist) herbivores by visual or olfactory masking of the presence of host plants, the emission of insect repellent cues by associated plants, or attraction of insects to other non-crop host plants (Finch and Collier, 2000; Root, 1973). Plant diversity can also influence the natural enemies of arthropod herbivores, such as predators and parasitoids. Companion plants of crop species may offer alternate prey or hosts, pollen, nectar, refugia, and microhabitats that may not be offered by monocultures (Landis et al., 2000). On the other hand, the more complex vegetation and olfactory composition of intercrops may influence the ability of natural enemies to find their prey or hosts on the crop plants (Aartsma et al., 2020).
There is a mature body of literature reporting on effects on crop diversification on arthropod herbivore and natural enemy abundance, pest suppression, and to a lesser degree on impacts on yield. In a meta-analysis of studies reporting field-scale experiments on vegetation diversification, Letourneau et al. (2011) showed that diversification resulted in 44% higher natural enemy populations, 54% higher herbivore mortality and 23% lower herbivore populations. These effects gave rise to a 23% reduction in crop damage and translated to significant yield benefits when additive designs were used in which a companion crop or non-crop plant was added to the main crop (but not when a substitutive design was used where crop plants were replaced by non-crop plants). These findings are to a large extent echoed in a more recent meta-analysis of field scale studies, which reported that generalist predators had a strong positive response to plant diversity. However, specialist herbivores had a negative response to plant diversity, while generalist herbivores showed no significant response (Dassou and Tixier, 2016). While these and other analyses indicate that crop diversity at the field scale has clear potential to suppress herbivores via bottom-up and top-down mechanisms (Tooker and Frank, 2012), it may also have additional benefits in terms of biodiversity, nutrient cycling and water regulation (Tamburini et al., 2020). On the other hand, the management of intercrops can be challenging in heavily mechanized farming systems because of financial, time or logistic constraints (Lin, 2011). To address these constraints, the effectiveness and feasibility of strip cropping systems with strip widths varying from 3 to 24 m are studied (Juventia et al., 2021).
Landscape diversification for pest and disease control
Natural enemies provide a potentially powerful mechanism for regulation of arthropod herbivores in crops, but they require resources at landscape level to maintain viable populations at sufficiently high densities to be effective (Landis et al., 2000). The importance of the broader landscape in supporting natural enemies has been recognized since the 1960s (e.g. Galecka, 1962). However, work on the landscape effects on pest control has accelerated tremendously since Thies and Tscharntke (1999) published a statistically convincing empirical approach to demonstrating these effects (Box 1).
Spatially-indexed regression analysis to explore landscape effects on local observations of pest control.
In short, the approach used by Thies and Tscharntke (1999) entails measuring biological control activity across a series of landscape replicates. Usually measurements in each landscape are made at a single site, using, for instance cage studies or sentinel prey to measure the activity of predators and hymenopteran parasitoids. Sentinel prey can consists of paper cards with eggs of a pest species, which are then retrieved after a short period of exposure in the field. Land use in each landscape is observed by using satellite images in combination with ground truthing and online information systems. Covariables are also recorded at each sentinel site, e.g. the type of background vegetation, or if it is in a crop field, the management in the crop field. Finally, relationships between land use, crop management, and biological control activity in the field is studied using regression. The approach is illustrated in Figure 1.
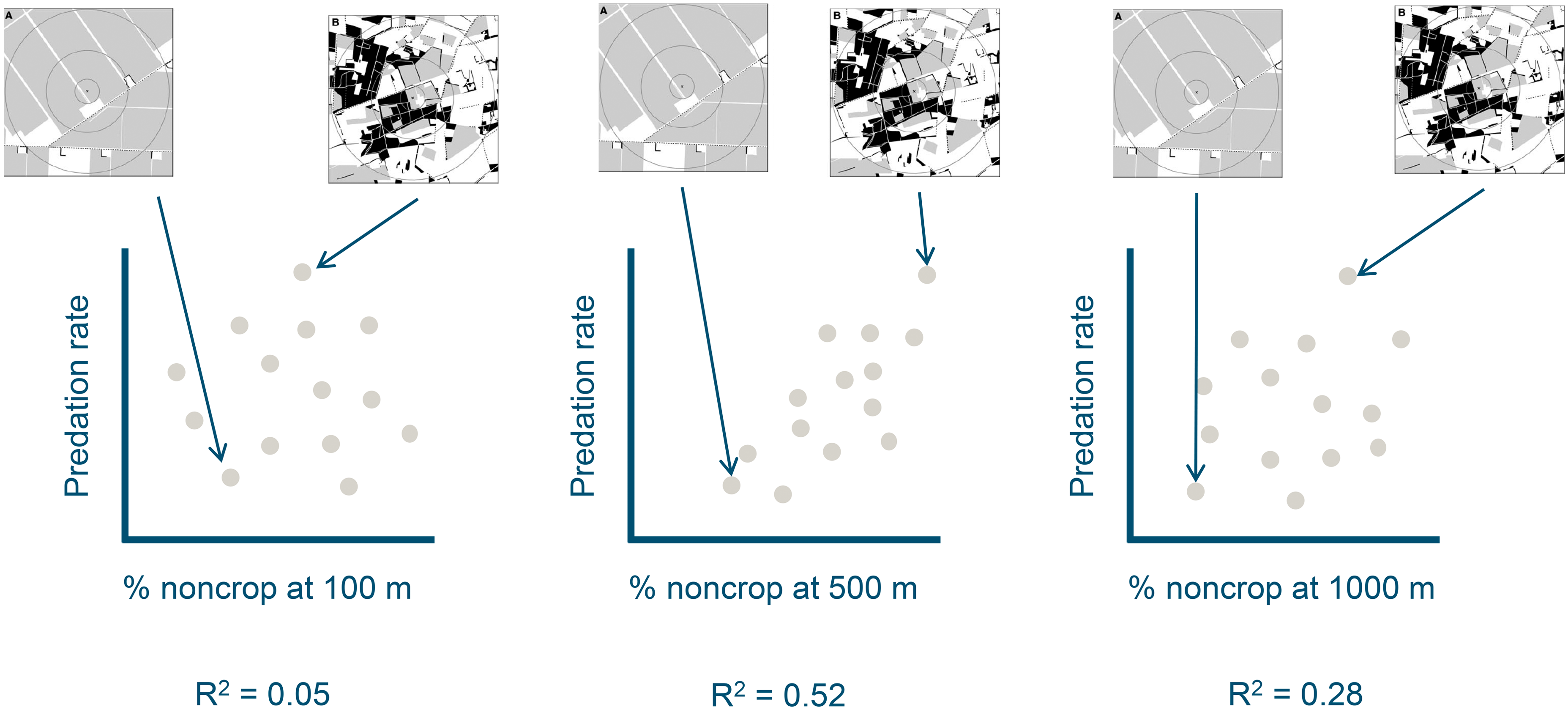
Illustration of spatially-indexed regression analysis approach based on a hypothetical dataset of 14 landscapes. Measurement of pest control are conducted at the center of each landscape and regressed on land use (% non-crop habitat) in circles of 100, 500 and 1000 m around the point of observation.
Since early papers demonstrated that diversity and semi-natural habitats in agricultural landscapes affect pest control in field crops (Landis and Haas, 1992, Thies and Tscharntke, 1999), hundreds of studies have been done globally. Recent syntheses of these demonstrate on the one hand that landscape usually has significant effects on pest control variables, but these analyses also highlight a high diversity in outcomes across studies (Karp et al., 2018). As a result, it is difficult to draw a simple conclusion such as ‘structurally diverse landscapes provide better pest control than simple landscapes’. On the other hand, Dainese et al. (2019) show that the richness of natural enemies positively influences pest control. This analysis also revealed cascading effects of landscape simplification, mediated through the impoverishment of the natural enemy and pollinator communities and associated biocontrol and pollination ecosystem services, which reduce crop productivity. While these findings highlight the importance of maintaining effective and diverse communities of natural enemies and pollinators in agricultural landscapes, species responses to semi-natural habitat can be variable and context specific (Karp et al., 2018). This variation can be explained by various mechanisms, including limited colonization of beneficial arthropods because of in-field disturbances imposed by farm management practices (Tooker et al., 2020) or species diversity and food web structure (Yang et al., 2021). Tscharntke et al. (2016) summarize the resulting conundrum through five hypotheses:
- Pest populations have no effective natural enemies in the region; - Natural habitat is a greater source of pests than natural enemies; - Crops provide more important resources for natural enemies than does natural habitat; - Natural habitat is insufficient in amount, proximity, composition or configuration to provide large enough enemy populations for pest control; - Agricultural practices counteract natural enemy establishment and biocontrol provided by natural habitat.
These hypotheses may all be at play to varying degrees in different contexts. This highlights the fact that the outcome of diversification practices based on semi-natural habitat depend on species-specific interactions between plants, arthropod herbivores and their natural enemies, and management practices that take place in these landscapes, prohibiting large-scale generalizations for habitat management.
While earlier work on landscape diversification and pest control focused on the role of (semi)-natural habitats (Thies and Tscharntke, 1999), crop diversity can also be an important component of landscape diversity. For instance, Redlich et al. (2018) found that predator and parasitoid densities increased with crop diversity at the landscape scale, and that pest control in fields with high landscape-level crop diversity was 8 to 33% greater than in low-diversity landscapes. Similarly, heterogeneous landscapes characterized by small fields and longer rotation regimes in Sweden show high levels of aphid pest control (Rusch et al., 2013). In addition to the diversification in terms of the number of (crop) habitat types (compositional heterogeneity), the spatial arrangement and size of fields may also influence pest control (configurational heterogeneity; Fahrig et al., 2011). For instance, using meta-analysis, Martin et al. (2019) showed that the abundance of 70% of pollinator and 44% of natural enemy species were positively related to edge density, and pollination and pest control was 1.7- and 1.4-fold higher in fine-grained than in coarse-grained landscapes, respectively. High yields were achieved in arable-dominated landscapes with high edge densities. This conclusion is reinforced by findings of Gagic et al. (2021) who showed that landscapes with large cotton fields and a low proportion semi-natural habitat had higher pest colonization rates, higher insecticide application rates and lower cotton yields. This suggests that enhancing edge density can promote functional biodiversity and yield-enhancing ecosystem services.
Why do farmers use pesticides?
Pesticides are easy to use and usually effective for their intended purpose, particularly in the short term. Some provide a knock-down effect that is highly appealing for farmers. However, pesticides also have serious drawbacks, including risks of side-effects on the environment and humans. Poisoning of farmers has been recorded over and over, and while precautions reduce exposure substantially, negative effects on workers and their families have been recorded, and not only when farmers have few options with which to protect themselves (Jokanovic, 2018; Pretty, 2005). Moreover, the effectiveness of insecticides in the longer term is questioned because they can trigger pest resurgence after natural enemy populations have been decimated. Based on model simulations and a meta-analysis, Janssen and van Rijn (2021) conclude that pesticide applications do not reduce pest densities significantly when natural enemies were present, which covered the vast majority of the reviewed cases. Nevertheless, rather than a last resort, pesticides are often used in conventional farming as a prophylaxis, as with neonicotinoid seed dressings (Bakker et al., 2020).
Zou et al. (2019) assessed biological control of rice pests across a landscape gradient and found high levels of biological control, irrespective of landscape composition. Despite this, all farmers routinely used pesticides. When analyzing yields in split plots, one half pesticide free and the other half managed according to ‘farmer practice’, the economic value of yield loss (on average 20%) equaled the cost of the pesticides. Zou et al. (2019) concluded that farmers used the pesticides to reduce their risks and be more certain that they would attain a certain yield level. However, on average, pesticides did not pay off from a purely cost-benefit point of view in which the impacts of pesticides on the environment and health were not considered.
Lock-in and path dependency
Another phenomenon driving continued pesticide use is known as ‘lock in’ or ‘path dependency’ (Vanloqueren and Baret, 2009). Path dependency occurs when complex systems are evolving with evolution of entities, interests and methodologies in different sub-compartments of a system dependent on each other. These entities and interest groups in different sub-compartments will try to “maintain business as usual” or develop further on their current path in order to obtain continuity and ensure long term viability and retain return on investment. In the case of farming, Bakker et al. (2020) distinguish four sub-compartments relevant to pest control: farm, landscape, society, and technology. Thus, in the face of changes in scale of operation (farm), field enlargement (landscape), government policy (society) and focus on chemical-based pest control solutions (technology), evolution of farm systems to maintain continuity most often means farmers find it difficult to get out of the habit of using pesticides (Bakker et al., 2021). They are effectively locked-into production methods that have significant environmental side effects, but are financially the best option and nearly impossible to escape from (Vermunt et al., 2021). Moreover, the use of insecticides may result in a deskilling of farmers while farmers with limited pest and natural enemy identification skills tend to be more reliant on insecticides than farmers with a higher level of “ecoliteracy” (Wyckhuys et al., 2019). Parts of the agricultural industry have a stake in perpetuating farmers’ dependency on pesticides (Pretty, 2005; Van den Bosch, 1978).
A way forward?
Most scholars in the field of plant protection have been exposed to the idea of IPM. Several definitions exist, but a typical one is ‘a decision-based process involving coordinated use of multiple tactics for optimizing the control of all classes of pests (arthropods, pathogens, weeds, vertebrates) in an ecologically and economically sound manner’ (Ehler, 2006; Prokopy, 2003). This definition acknowledges both the ecological and economic dimensions of pest control, but it does not recognize that pest control has societal ramifications as pesticides have been found deep into nature areas (Hallmann et al., 2017) and the consequences of pesticide use likely play out at a landscape level, thus transcending the farm (Bakker et al., 2021). Furthermore, pesticides used in crop fields sometimes spill over to nearby villages, resulting in exposure of citizens, who rightfully protest against this exposure. Product quality standards and pesticide regulations are meant to minimize these negative spill-overs, but challenges remain.
Common understandings of IPM emphasize the integration of different methods, but from another perspective, the integration of objectives might also be sought (Vereijken, 1999). If society were to insist on additional objectives from pest control, such as healthy produce and a healthy environment, and just rewards for farmers and others working in the value chain, it might result in a new appreciation of the relative merits of ecological and pesticidal solutions. This would have the potential to fundamentally change the notion of the damage threshold as it was defined 60 years ago (Higley and Pedigo, 1997; Stern et al., 1959), which only considers the rather narrow scope of costs and benefits to farmers who are the direct financial beneficiaries of crop yield. While pesticidal solutions are strong on the side of efficiency and control efficacy, and on short-term benefits to farmers, they do not score well on the broader societal goals such as a healthy environment.
What then can we learn from nature? There is general agreement that biodiversity is essential for agriculture and food production as it provides genetic resources for crop and livestock improvement and can strengthen the resilience of agricultural systems (FAO, 2019). For instance, biodiversity in the form of genetic resources is indispensable for resistance breeding, one of the cornerstones of IPM. Moreover, diversity at field, farm and landscape level can strengthen multiple ecosystem services (Tamburini et al., 2020), and there is growing evidence that abundant and diverse communities of natural enemies and pollinators can improve yield (Dainese et al., 2019). However, pest control depends on the action of natural enemies, which rely on resources in their environment and may be disrupted by intensive management practices. Therefore, natural pest suppression is less reliable than the use of pesticides, and trade-offs between natural pest suppression and yield may occur (Tamburini et al., 2020). This reinforces the notion that the effects of diversification are contextual and need to be ascertained for particular production systems. The social fabric of the production environment must to be factored in to weigh the benefits and costs of control solutions modeled after natural systems (Thomine et al., 2021).
In conclusion, while there is no doubt that diversity has multiple benefits for the functioning of agricultural systems and the services provided by them, we must be realistic about the potential for nature-based pest control solutions. These come with benefits, but also costs, and if their use is to be prioritized, society must be willing to compensate farmers financially for the additional losses they may incur. Therefore, a greater use of nature-based solutions hinges on a paradigm shift in agriculture, a shift from farmers meeting their own financial needs while supplying food to the market, to one where they are rewarded for being stewards of the environment while producing healthy food. A broad transition towards wider adoption of nature-based solutions in agriculture requires an economic mechanism that rewards stewardship that serves wider societal needs while discouraging management that does not.
Footnotes
Acknowledgements
We are grateful to Jim Sumberg, Ken Giller and two anonymous reviewers for their helpful comments on earlier versions of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.