
Abstract
While the world’s attention is focused on controlling COVID-19, evidence points at the biodiversity crisis as a leading factor in its emergence, and the outbreak of many past emerging infectious diseases. Agriculture is a major driver of biodiversity loss globally. Feeding a growing human population in ways that minimize harm to biodiversity is thus imperative to prevent the next COVID-19. Solutions exist, but the burden of implementing them should not be left to farmers alone, who are mainly small-scale family farmers. Supportive policies and markets are needed, but unlikely to bring about the required changes alone. A global concerted effort similar to the Paris Agreement for climate is probably required.
Viewpoint
Infectious diseases are caused by pathogenic microorganisms, such as bacteria, viruses, parasites or fungi (Anderson et al., 1991). The most recent, COVID-19, is caused by the SARS-CoV-2, a virus that belongs to the Coronaviridae family, and has spread across the globe in about 2 months from its origin in China, infecting people and claiming lives at an exponential rate, and leading to measures that are disrupting the global economy in an attempt to contain it (Hellewell et al., 2020; McKibbin and Fernando, 2020; Ramelli and Wagner, 2020; Sohrabi et al., 2020). Outbreaks of infectious diseases are on the rise (Smith et al., 2014). Most are zoonotic (60% of 335 infectious disease outbreaks that occurred between 1940 and 2004), meaning that they are spread from animals to humans, and the majority of these zoonoses (72% of the zoonoses that occurred between 1940 and 2004) originates from wildlife (Jones et al., 2008). COVID-19 is just the last in a long list of zoonoses originating from wildlife species (Zhou et al., 2020). In the past 20 years only, humanity was hit by three coronaviruses (SARS-CoV-1, 2003, Li et al., 2005; MERS-CoV, 2012, Zumla et al., 2015, SARS-CoV-2, 2019, Sohrabi et al., 2020), one influenza virus (Swine flu, 2009, Borkenhagen et al., 2019), two arboviruses (Sam et al., 2015; Zika virus, 2015, Metsky et al., 2017) and one filovirus (Ebola virus, 2014 and 2018, Bourgarel and Liégeois, 2019). When in 2017, it was demonstrated that SARS-CoV-1 may have emerged through recombination among different virus strains in a single bat population in a cave in Southern China, the authors of the paper warned that “the risk of spillover into people and emergence of a disease similar to SARS is possible” (Hu et al., 2017). This indeed reads like a forecast of SARS-CoV-2 and the current COVID-19 pandemic….
So why do we see more cases of pathogens crossing from wildlife to humans? Several lines of evidence point to the current biodiversity crisis—which can be qualified as mass extinction (Barnosky et al., 2011)—as one of the primary causes. Land use change has been the leading driver in the emergence of zoonoses caused by a pathogen with a wildlife origin between 1940 and 2004 (Figure 1). The link between deforestation and these events has been established both theoretically (Faust et al., 2018), as well as empirically in a number of cases, including the emergence of Mycobacterium ulcerans and the recent outbreak of Ebola (Morris et al., 2016; Olivero et al., 2017). First, deforestation brings wildlife and people (and their livestock) into greater contact, increasing the risk of spillover (Morse et al., 2012). The fact that 70% of forests are within 1 km of a forest edge (Haddad et al., 2015) illustrates how pervasive ecotones, where wildlife, people and livestock interact, are. Second, species that survive (or thrive) during deforestation tend to be less sensitive to human disturbance (opportunistic/generalist species), and thus the ones most able to transmit pathogens to humans (or their livestock) (Johnson et al., 2020; Keesing et al., 2010).
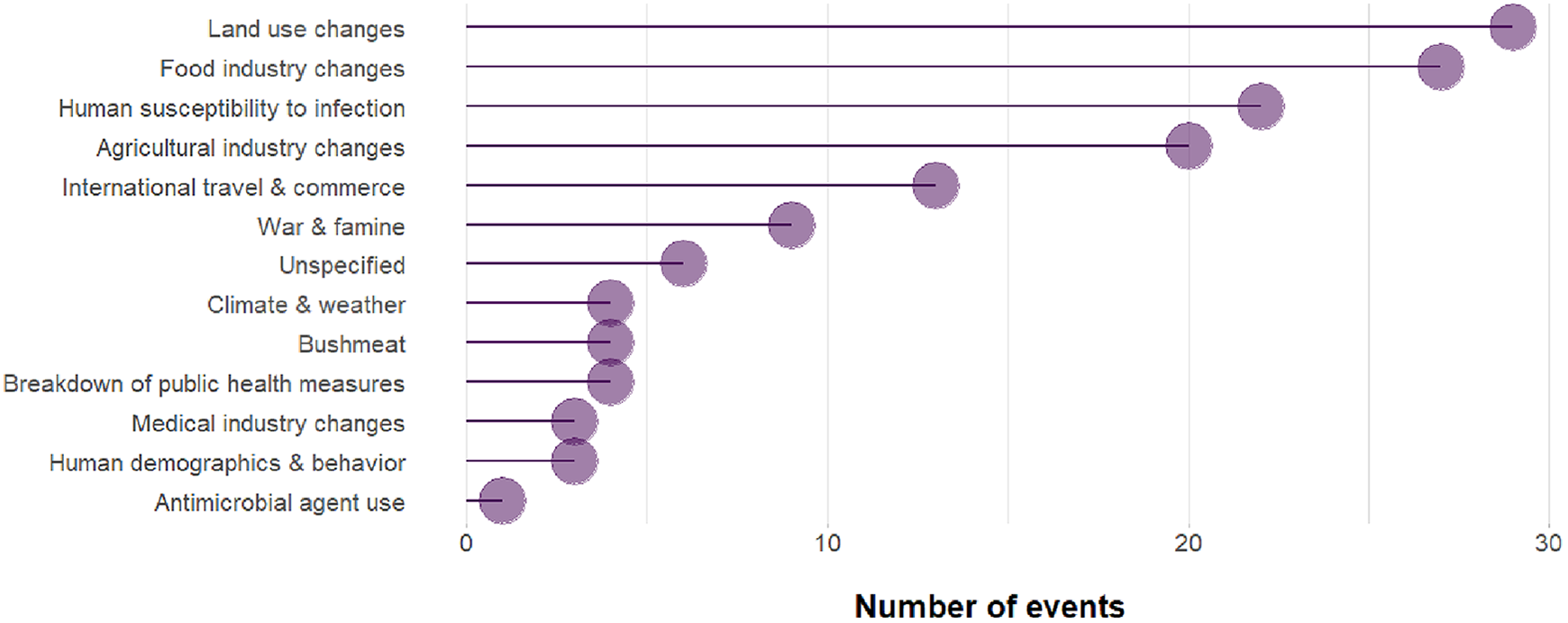
Most commonly cited driver associated with 145 events of zoonotic emerging infectious diseases caused by a pathogen with a wildlife origin between 1940 and 2004 (from data published by Jones et al., 2008). “Land use changes” generally refer to changes in cropping practices and “agricultural industry changes” generally refers to changes in livestock husbandry. Together, they represent more than a third of all events.
Finally, pathogens may be more prevalent in animal communities with reduced diversity. Diverse host communities tend to include less competent reservoir species for any given pathogen, lowering prevalence and risk of spillover (to humans or livestock), a phenomenon known as the “dilution effect” (Ostfeld, 2009). Trophic downgrading—the extirpation of apex consumers through human activities (Estes et al., 2011)—is less visible and quantifiable than deforestation, but can lead to similar changes in wildlife communities with ripple effects on a number of ecosystem processes including disease dynamics (Dirzo et al., 2014). For example, the continuing emergence of Lyme disease in North America has been attributed to the decline of the red fox, leading to an increase in the abundance of small mammal hosts of the pathogen (Levi et al., 2012). Extirpation of lions and leopards in parts of Ghana led to an increased abundance of baboons coming more frequently into contact with people, with high transmission risk of intestinal parasites (Ryan et al., 2012; Taylor et al., 2016). In India, the increasing number of human cases of rabies has been linked to the decline of vultures (Markandya et al., 2008).
Once biodiversity changes have altered the dynamics of pathogen transmission, the likelihood of spillover to humans is high. Indeed, humans are one of the most abundant vertebrates worldwide, representing 36% of the global biomass of mammals (Bar-On et al., 2018), and we interact with almost every ecosystem around the world (Ellis and Ramankutty, 2008). This risk is, of course, increased by practices such as bush meat consumption (Wolfe et al., 2005). Yet at the same time, bushmeat represents a critical source of quality protein and readily available micronutrients with few alternatives for millions of people in tropical countries (Fa et al., 2003; Golden et al., 2011). Rodents and bats are often the origin of zoonoses caused by a pathogen with a wildlife origin (Figure 2; bats are for example known reservoirs of SARS-like coronaviruses; Hu et al., 2017), simply because these two orders represent more than 60% of all mammal species (Burgin et al., 2018) while pathogen richness tends to correlate with host species richness (Mollentze and Streicker, 2020). Many zoonotic pathogens may then be transmitted to humans directly from these reservoirs (e.g., Ebola, Olivero et al., 2017) or through another intermediate wild host (e.g., SARS, for which the palm masked civet was likely an intermediate host between bats and humans; Li et al., 2005). For many others, however, livestock is an intermediate host (Figure 2). Livestock and poultry are indeed very abundant vertebrates worldwide, representing 60% of the global biomass of mammals and 70% of the global biomass of birds, respectively (Bar-On et al., 2018), and have accompanied human in most ecosystems around the world (Gilbert et al., 2018). Livestock—particularly when kept in industrial operations where large numbers of animals of low genetic diversity are confined—may amplify pathogens and provide them with an opportunity to mutate and become transmissible to humans (Jones et al., 2013). Change in livestock husbandry has been an important driver in the emergence of zoonoses caused by a pathogen with a wildlife origin between 1940 and 2004 (Figure 1). Industrial poultry farming played a major role in the outbreak of the H5N1 avian influenza (Graham et al., 2008). The recent outbreak of Middle East respiratory syndrome (which is caused by a coronavirus, similarly to COVID-19) is thought to have crossed from bats to humans, with camels as an intermediate “stepping stone” (Zumla et al., 2015).
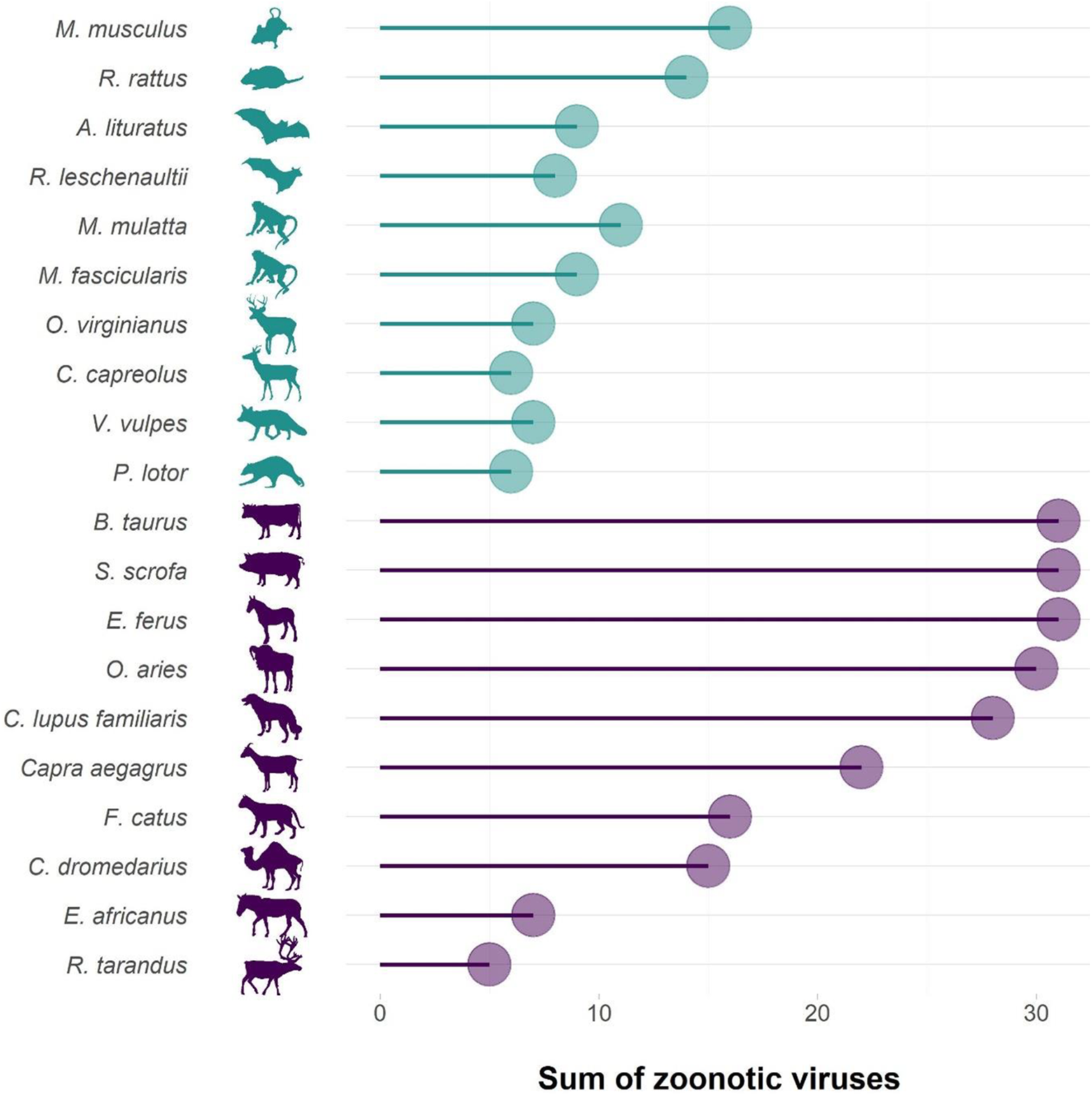
Sum of zoonotic viruses carried by most common livestock and pet mammal species compared to the wild mammal species with the highest sums in the orders Rodentia, Chiroptera, Primates, Artiodactyla and Carnivora (two species per order, from data published by Johnson et al., 2020).
Safeguarding biodiversity thus appears essential to prevent future pandemic zoonotic diseases. In a 2017 article of Nature that followed the publication of evidence suggesting that SARS-CoV-1 emerged from a bat colony in Southern China, Professor Kwok-Yung Yuen—one of the co-discoverer of the SARS coronavirus—warned that to prevent the emergence of another SARS-like zoonosis “we should not disturb wildlife habitats and never put wild animals into markets” (Cyranoski, 2017). If overexploitation remains a leading threat, agriculture was found to be one of the “biggest killers” for 62% of the species listed as threatened or nearly threatened (Maxwell et al., 2016) (crop farming was found to threaten 54% of these species and livestock farming 26%). About 22% of the land area represented by Biodiversity Hotspots is threatened by agricultural expansion (Veach et al., 2017). Furthermore, these areas often overlap with emerging disease “hotspots” (Jones et al., 2008). Agricultural expansion, as well as agricultural intensification, are major drivers of biodiversity loss (Baudron and Giller, 2014; Foley et al., 2011; Kehoe et al., 2017). Feeding a growing human population in ways that minimize harm to biodiversity is thus imperative, and core to achieving the “World we Want” (UN General Assembly, 2015).
Norman Borlaug—wheat scientist, “Father of the Green Revolution” and recipient of the Nobel Peace Prize in 1970—argued that agricultural intensification does just that: sparing land for nature (Borlaug, 2007). If crop yields had remained at their 1961 levels, India would have required 19.9 million ha of additional land to reach the amount of maize produced in 2018 and 87.6 million ha of additional land for its wheat production in 2018 (Figure 3). Mexico would have similarly required a substantial amount of additional land, likely through conversion of nature to farmland (22.2 million ha for maize and 1.2 million ha for wheat; Figure 3). Spared areas are smaller in countries largely bypassed by the Green Revolution, such as Kenya, but still substantial (1.1 million ha for maize, and 0.2 million ha for wheat; Figure 3). Several publications have demonstrated that maximizing crop yields and thereby limiting the total area under agriculture—an approach known as “the Borlaug hypothesis” or “land sparing”—could be our best option to minimize trade-offs between agriculture and biodiversity (Balmford et al., 2019; Phalan et al., 2011). Recommending to segregate biodiversity and agriculture (and other human activities) is indeed tempting to reduce the risk of emerging zoonoses (Marco et al., 2020).
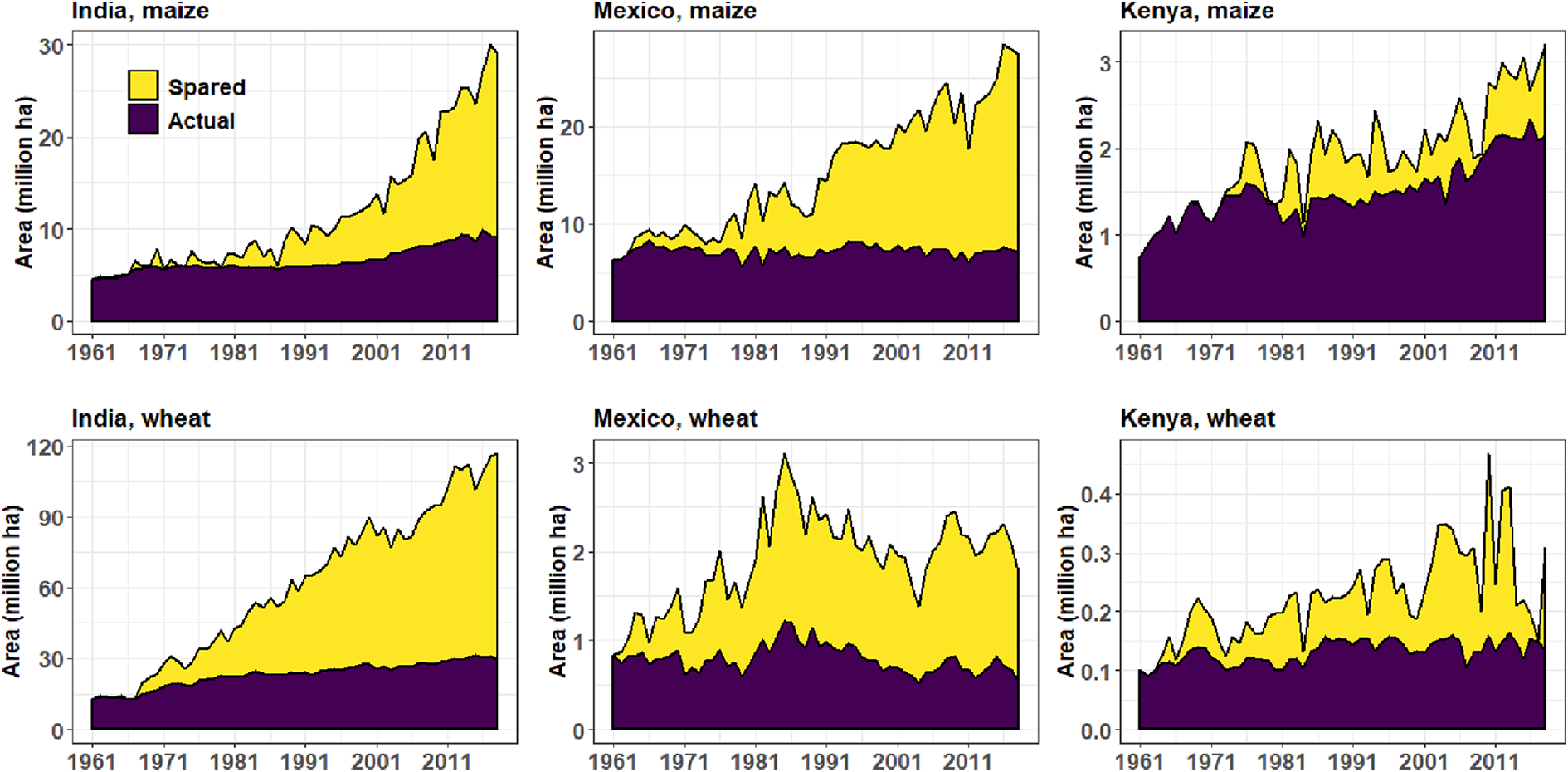
Comparison of actual land and land that has been theoretically spared by yield increase (i.e., additional land that would have been needed if yields had remained at their 1961 levels) for maize and wheat in India, Mexico and Kenya (source: FAOSTAT).
Land sparing however, suffers from at least two major weaknesses. First, maximizing yields implies the use of large quantities of agricultural inputs, often impacting a larger area than merely the land farmed intensively. When large rates of nutrients (e.g., nitrates and phosphates) and pesticides are used, they may be transported for thousands of kilometers downstream and downwind (Carpenter et al., 1998; Gordon et al., 2008; Hoferkamp et al., 2010; Smith et al., 1999). Second, maximizing yields is often accompanied by extreme simplification of agroecosystems and loss of farmland biodiversity. This may reduce ecosystem disservices—such as the risk of zoonosis emergence—but it also cuts farmers from vital ecosystem services, including soil fertility maintenance, pest control and regulation of microclimate (Kebede et al., 2018; Sida et al., 2019; Yang et al., 2020). Ecosystem services provided by biodiversity may have far reaching impacts on human well-being, including improved diets (Baudron et al., 2019b). Indeed, such services are particularly crucial to family farmers in developing countries (as in the examples given), as they use insignificant quantities of external input and often cannot afford them. They are often qualified as being “organic by default”—but still produce more than half of the food consumed globally through processes supported by nature (Tittonell et al., 2016). Ecosystem services are also important for the sustainability of other—more intensive—production systems as well, as they also depend on ecosystem services as highlighted by a recent FAO report (FAO, 2019). The urgency to maintain ecosystem services to sustain the global food system is epitomized by the increase in soil degradation and in pesticide use (illustrating the decline of pest biocontrol), and by the decline of pollinators worldwide (Garibaldi et al., 2009; Oldeman, 1994; Wilson and Tisdell, 2001).
In response to these issues, and at the opposite side of the spectrum of solutions to minimize trade-offs between agriculture and biodiversity is “land sharing” (Clough et al., 2011; Perfecto and Vandermeer, 2010; Wright et al., 2012). It is based on a land-use model that integrates (rather than segregates) production and biodiversity on the same land units, by minimizing the use of external inputs and retaining patches of natural habitat within farmlands. Land sharing, however, also suffers from at least two major weaknesses. First, low-input agriculture generally achieves yields inferior to those achieved by intensive farming, and thus consumes more space—likely taken away from nature—to meet any production target (Green et al., 2005). Second, agriculture can rarely be sustained without some degree of external inputs. For instance, some nutrient inputs are needed to at least compensate nutrients exported in harvest (Vitousek et al., 2009). In addition, crop losses due to pests tend to be severe and pesticide-free farming can only be achieved in rare situations (Lucchi and Benelli, 2018; Oerke, 2006).
Ultimately, principles from both land sparing and land sharing will be part of the required solution: inputs are needed, but their spill-over needs to be managed (through e.g., conservation agriculture and precision farming); efforts to increase productivity need to be balanced with the need to retain ecosystem services; and using a landscape approach, as ecological flows of energy, materials (e.g., soil, water) and organisms (e.g., natural enemies, pollinators, species of concern to conservation) are best understood and managed at landscape-level, not at plot-level (Baudron and Giller, 2014). A landscape approach to reduce tradeoffs between farming and biodiversity involves “targeted sparing,” in biodiversity hotspots only, and only to the degree necessary for maintaining critical wildlife habitats (e.g., habitat of species with restricted range; Folberth et al., 2020). In areas where the opportunity cost of taking land away from production is high, such targeted sparing should center on the establishment of networks of small natural or semi-natural areas across the landscape rather than large contiguous conservation areas (Maes et al., 2015). These networks should however cover the range of abiotic conditions in the landscape, including some highly productive land, for biodiversity to be effectively conserved (Fischer et al., 2006). It is also beneficiary for the network to include critical transition zones (e.g., riparian areas, mangroves), due to the critical role they play as conduits for materials and organisms between adjacent ecosystems (Ewel et al., 2001). In addition to be feeding and breeding grounds, as well as dispersal corridors, for many species, they provide critical ecosystem services including the regulation of water flow, and the trapping and transformation of sediments, nutrients and pollutants (Levin et al., 2001; Richardson et al., 2007). The role multifunctional landscapes can play in both conserving biodiversity and feeding people now and in the future is increasingly recognized (Kremen and Merenlender, 2018). Southern Ethiopia offers an example of such a multifunctional landscape that hosts high biodiversity while being more productive than simpler landscapes (Baudron et al., 2019a). This multifunctional landscape of Southern Ethiopia is also more sustainable and resilient (Duriaux Chavarría et al., 2018), provide more diverse diets (Baudron et al., 2017) and produce staple food with higher nutritional content (Wood et al., 2018).
Current policies and markets, however, tend to impede multifunctional landscapes, and to promote instead simplified agricultural landscapes depleted in biodiversity, highly dependent on external inputs and susceptible to shocks and stresses. Promoting “landscapes that work for nature and people” (Kremen and Merenlender, 2018) requires the removal of perverse policies such as subsidies and distortions, and their replacement by incentives that promote production systems that are less demanding of land and other natural resources (Mooney et al., 2005). Agricultural landscapes are also shaped by our consumption patterns. Consumers in developed countries (in particular) export biodiversity threats through their consumption of coffee, tea, chocolate, sugar, textiles, fish, etc (Lenzen et al., 2012). In order to promote consumption patterns that reduce impact on biodiversity, proper pricing, certification and labeling are required. The true costs and benefits to nature of agricultural practices need to be embodied in the prices of the corresponding commodities (Balmford et al., 2002). Environmental labels have a long history (e.g., organic produce, Protected Designation of Origin, Gracia and de-Magistris, 2016) and could be expanded to communicate impacts and benefits on nature to consumers, as an incentive to purchase more environmentally-friendly products. Consumption patterns that cause biodiversity loss tend to also be those causing unhealthy diets, one of today’s leading risk factors for mortality globally (Willett et al., 2019).
Yet supportive policies and markets alone are unlikely to bring about the required changes. A large share of the global food produced does not enter the market, but is consumed by the small-scale family farmers who produce it (Frelat et al., 2016). Reducing the negative impact of our agricultural systems on biodiversity will thus require a global, concerted effort similar to the Paris Agreements for climate. Although several high-profile reports have recently highlighted the importance of biodiversity for our very existence (FAO, 2019; HLPE, 2017; IPBES, 2019), raising public awareness of the consequences of the biodiversity crisis to the same intensity as awareness of the consequences of the climate crisis is critical (Legagneux et al., 2018). In a more-and-more connected world, we share joint responsibility for our global future. We must shoulder the burden of feeding humanity whilst maintaining the very biodiversity that confers resilience on our increasingly cultivated planet against shocks such as the global pandemic of COVID-19.
Footnotes
Acknowledgements
We are grateful to Ken E. Giller and Bruno Gérard for their comments on an earlier version of the manuscript. We also thank an anonymous reviewer for his/her critical and constructive comments.
Author contributions
FB conceived the paper and wrote the first draft of the manuscript. FL contributed to the writing of the final manuscript
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.