
Abstract
The outfit of husbandry facilities of, and the enrichment provided for, experimental rodents plays an important role in the animals’ welfare, and hence also for the societal acceptance of animal experiments. Whether rats and mice benefit from being provided with running wheels or plates is discussed controversially. Here we present observations from a feeding experiment, where rats were provided a running plate. As a pilot study, six identical cages, with three animals per cage, were filmed for six days, and the resulting footage was screened for the number of bouts and the time the animals spent on the plates. The main activities observed on the plate in descending order were sitting (18.5 ± 13.8 bouts or 8.0 ± 13.7 min/animal per day), standing (10.2 ± 11.6 bouts, 3.8 ± 4.2 min), running (8.2 ± 13.3 bouts, 10.1 ± 21.4 min), grooming (2.0 ± 2.8 bouts, 6.7 ± 25.7 min), sleeping (1.0 ± 2.6 bouts, 24.0 ± 61.8 min) and playing (0.5 ± 0.9 bouts, 0.1 ± 0.5 min). Most of these activities (91% of all bouts, 90% of total time) occurred at night, similar to previous studies on running wheel usage. The running plate seems well-accepted as cage enrichment, even though in further studies, the motivating triggers and the effects of long-term use could be evaluated more in-depth.
Introduction
Rats (Rattus norvegicus) are indispensable research animals and represent more than 11% of animals used in research in Switzerland in 2019. 1 Whenever the experiment allows for it, adapting the environment to meet the social and environmental needs of these animals is essential to produce good quality science. 2 Abnormal behaviours caused by insufficient welfare standards can have a detrimental impact on experimental outcomes 3 and a negative effect on reproducibility. 4
The term ‘enrichment’ has been defined many times depending on the environment it is employed in (i.e. zoo, laboratory) and the species it is applied to (i.e. wild animals in captivity, laboratory rodents).4,5 Structural enrichment is generally thought to promote species-specific cues for the display of natural behaviours, and promoting physical and psychological health in captive animals. 6 Non-suitable enrichment, on the other hand, can provoke stress reactions and potentially lead to pathological conditions. The use of running wheels as structural enrichment mainly aims to encourage laboratory rodents to exercise, and in rats has been shown to decrease blood pressure and body fat. 7 It even increased the average lifespan of female rats by more than 9%. 8 The motivation of rodents to use running wheels and the resulting effects are influenced by multiple factors both internal (e.g. genes, hormones, hunger and thirst, psychology) and external. 9
When offered access to a running wheel, wild mice with ample territories and a wide range of stimuli from living in urban and suburban Netherlands made frequent use of the wheels, even without a nutritional reward system. 10 Living in a more remote setting, their Paraguayan counterparts made little use of running wheels presented in the same manner. 11 These experiments tried to answer, among other questions, whether wheel running should be considered a stereotypy, as stereotypic behaviours are usually only assigned to captive animals. 11 Even though the Dutch animals seem to disprove wheel running as a stereotypy, Mason and Würbel 12 still insist that wheel running in laboratory mice is a stereotypy, as it corresponds with their standard definition of being ‘repetitive, unvarying and apparently goalless’. Whether or not wheel running qualifies as such, stereotypic behaviour per se is not harmful. In a sub-optimal environment in captivity, animals compensate with stereotypies for reduced welfare. For example, fewer abomasal ulcers were seen in tongue-playing calves compared with the ones that did not show any sign of repetitive oral behavioural patterns. 13 Sometimes stereotypies can even have a positive impact on general health, as known from crib-biting horses, where it lowers the heart rate or increases production of saliva that can regulate gastrointestinal pH if swallowed. 14 In mice, running in wheels decreased other behavioural patterns such as stereotypic jumping or bar-mouthing. 15 Mason and Latham 16 even described stereotypies as ‘do-it-yourself enrichment’ for coping with sub-optimal environments, and therefore the animals benefit from performing repetitive behaviour such as mice reacting with wheel-running when faced with food restrictions. 15
Wild rats are staple omnivores reflected, for example, by the stomach content analysis from different regions in Japan: the amount of animal based food items, namely molluscs, insects, slugs and fishes, was relatively higher compared with other components such as leaves, seeds, fruits and roots. 17 Different food components can have a direct influence on the animal’s behaviour. For example, rats fed a high-carbohydrate diet ran significantly more in a running wheel than rats fed a high-fat diet. 18 Not only the feed content, but also food deprivation and scheduled feeding events influence the intensity of running activity in rats9,19 as well as physical activity in general, irrespective of whether or not running wheels are provided. 20
We used an experimental setup designed for a different purpose (as part of a study on tooth wear and isotope physiology) with group-housed rats fed different diets. As part of the enclosure furniture, running plates were provided. Irrespective of diet, running on the plates is expected as a behavioural enrichment engaged in by all rats. We expected individuals from all feeding groups to make ample use of the running plates from the first day onward. We hypothesised that running would be the main activity, but also evaluated other behaviours occurring while a rat was on the running plate.
Materials and methods
This study was part of a long-term feeding experiment approved by the Cantonal Veterinary Office in Zurich, Switzerland (licence no. ZH 135/16). Eighteen female adult Wistar (RjHan:WI) rats (body mass 221 ± 16 g) were divided into six feeding groups consisting of three individuals each (identified though weekly fur marking). The rats were floor housed in compartments (0.58 m × 1.00 m × 0.50 m; width × length × height) on a 12-h light cycle (artificial light from 07:00 to 18:00, with natural light dimming until sunset around 19:00) at constant temperature of 20°C. The bedding consisted of dust-free softwood granulate, deep enough to allow digging behaviour. The structural enrichment of the cage consisted of two plastic drainpipes with different diameters, a metal climbing frame (0.25 m height), and a shelter (0.15 m height) with a 0.3 m diameter running plate (Trixie 60812 Laufteller, Tarp, Germany; Figure 1) screwed on top. Balancing on the rim of the running plate the rats could reach a maximum height of 0.4 m. The cage design was meant to encourage the animals to perform their natural range of movements, for example, grooming, stretching, hiding, playing and sleeping in every possible position.

Floor housed cage design for three Wistar female rats (Rattus norvegicus; N=18) in a long-term feeding experiment. The use of the running plate with a 30 cm diameter is analysed in this study for six days as a pilot experiment.
The rats were handled twice weekly for weighing, health checks and, if necessary, re-marking. Throughout the filming period, the cages were not cleaned completely; only soiled granulate was replaced to ensured that behaviour was not altered due to territorial stress. Water was provided for ad libitum consumption from two bowls which were cleaned and refilled daily. In accordance with their experimental group, the animals received a specific diet consisting of either mixed vegetables, mixed fruit, mixed grains, crickets, black soldier fly larvae or day-old chicks. The dietary composition can be found in Supplementary material Table S1 online. Due to the short duration of this experiment, no nutritional deficiency was anticipated, but animals additionally received a small portion of curd and sunflower seed oil mixed with a mineral-vitamin supplement calculated to match the composition of the respective diet.
Digital cameras (Metal mini array 1080P AHD-N HD CCTV Camera SONY IMX323 Full Digital 2mp Outdoor Waterproof ip66 Infrared) with dark cycle infrared recording enabled filming of the animals’ behaviour without disturbance. Filming started at 18:00 on the day of arrival (beginning of the dark cycle) and ended after six 24-h periods. The video sequences were saved, and after termination of the experiment, only the activities on the running plates were analysed in this explorative study. The video time stamps when a specific rat placed all four limbs on the running plate and when all four limbs were off the plate again were recorded. The time between those two time stamps was defined as one bout. If one rat was already on the plate and another one joined the first one, this was recorded as ‘in company’, but each rat’s activity on the plate was recorded separately. The activities were divided into seven specific categories, namely running clockwise, running anti-clockwise, sitting, standing, sleeping, grooming, playing (pin and pounce behaviour according to Panksepp and Beatty 21 ) or other (Supplementary Table S2). Descriptive statistics were performed using Microsoft Excel (Microsoft Excel® for Mac, v.16.36, Zurich, Switzerland). Although the experimental unit was the cage, we present data on the basis of all animals evaluated together (the average of all 18 animals, rather than an average of the mean animal value per cage). This procedure does not affect the calculated means, but leads to higher standard deviations compared with giving the latter only for the cage means. To explore a possible correlation between body mass and running, the Pearson’s correlation test was used in R (v.4.0.2; https://www.r-project.org). The impact of the diet group on the behaviour was not analysed statistically due to the limited sample size.
Results
During the observation period of six days, the rats spent 3.7% of their time on the running plates, during 4435 individual bouts.
Activities on the running plate
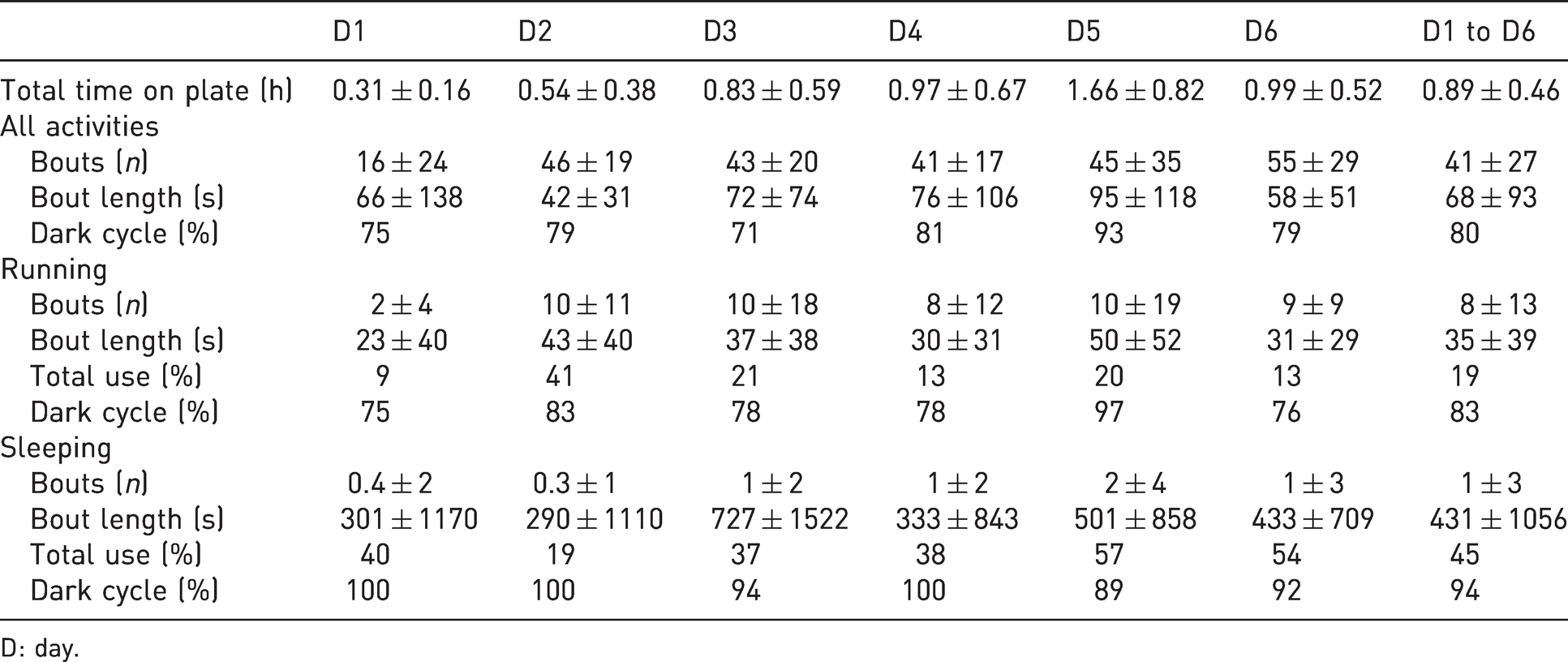
The mean bout length spent on the plate was 1.3 ± 5.2 min. Sleeping had the longest bouts (mean 23.7 ± 5.3 min), followed by grooming (mean 3.3 ± 5.2 min), running (mean 1.2 ± 5.2 min), sitting (mean 0.4 ± 5.2 min) and standing (mean 0.4 ± 5.2 min). The shortest bout length was during playing (mean 0.2 ± 5.0 min).
The main activity performed on the plate was sitting, with an overall percentage of 46% of total bouts (mean 18.5 ± 8.8 bouts/animal per day; Figure 2(a)), followed by standing with 25% (mean 10.1 ± 7.9), running 19% (mean 8.2 ± 9.0), grooming 5% (mean 2.0 ± 2.0), sleeping 2% (mean 1.0 ± 1.8) and playing 1% (mean 0.5 ± 0.6) of total bouts. Sitting and standing were performed by all rats, whereas only seven out of 18 rats (39%) slept on the plate. One rat spent 14% of its bouts on the plate sleeping (80% of this animal’s time spent on the plate). Other observed but not further defined activities on the plate included eating, jumping up to the cage brim or urinating. On several occasions (32% of total bouts), more than one rat was using the plate at the same time for sitting, sleeping, standing, grooming and running (12% of bouts in company) either next to or behind each other.
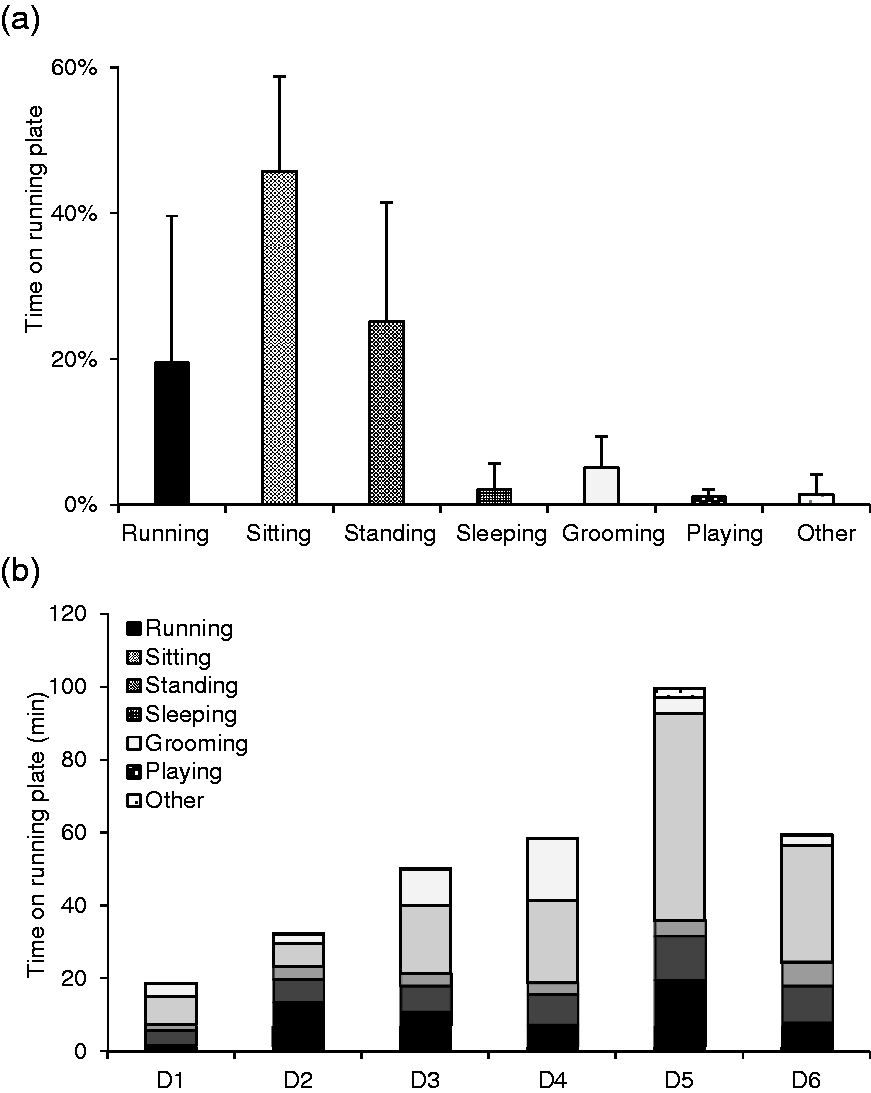
Individual mean (+SD) percentages of time that Wistar rats (Rattus norvegicus; N = 18) spent on running plates when they were offered access for a six-day period. (a) Different activities in per cent of the total time spent on the running plates and (b) the total daily time spent on the running plates in various activities. D: day.
Change in running plate use over time
Every day an average of 41.1 ± 27.2 bouts per rat occurred. Bouts increased from day 1 (D1; mean 16.3 ± 24.0 bouts per rat) of the experiment to D2 (D2 46.3 ± 19.4 bouts) and remained more or less constant from then on (D3 42.9 ± 19.6 bouts; D4 40.9 ± 17.4 bouts; D5 45.3 ± 35.4 bouts; D6 54.6 ± 29.5 bouts). The time of running plate use increased from D1 to D3 (D1 18.7 ± 19.2 min; D2 32.2 ± 1.4 min; D3 50.1 ± 2.7 min per rat). The increase in time between D2 and D3 was mainly due to increased sleeping time (Figure 2(b)). The time of running plate use then remained relatively constant (D4 58.5 ± 4.7 min; D6 59.5 ± 8.5 min), with the exception of D5, when a particularly high amount of time was spent sleeping (D5 99.6 ± 8.5 min). This was not due to one long sleeping bout by one animal, but rather due to several sleeping bouts lasting more than 1 h of three rats from different groups.
Running on the running plate
The average number of running bouts per day and rat was 8.2 ± 13.3. On average, every rat spent 10.1 ± 21.4 min per day on the running plate with actual running. The mean bouts as well as time were lowest on D1 (D1 mean 1.8 ± 3.7 bouts, 1.7 ± 3.2 min per animal). After an increase on D2 (D2 10.4 ± 11.5 bouts, 13.3 ± 17.3 min), the number of running bouts stayed relatively constant (D3 9.9 ± 18.3 bouts; D4 8.2 ± 12.1 bouts; D5 10.2 ± 18.8 bouts; D6 8.7 ± 8.8 bouts). However, the time per rat per day from D3 on was still very variable, with a remarkable peak on D5 (D3 10.7 ± 20.4 min; D4 7.4 ± 11.4 min; D5 19.4 ± 41.8 min; D6 8.0 ± 9.3 min)
For 55% of running events, the rats ran clockwise and for 45% anticlockwise, corresponding to 60% of total running time clockwise and 40% anticlockwise. For one animal, a mean of 20.8 bouts clockwise per day was recorded compared with a mean of 0.2 bouts anticlockwise, while other animals displayed nearly equal numbers, or the opposite preference. Subjectively, the rats appeared to run mainly on the bottom quarter of the running plate. Total running time across individuals was negatively correlated with body mass at the start of the experiment (Pearson’s rho = –0.55; p = 0.019; Figure 3(a); Supplementary Table S3) but not with the body mass gain during this short pilot phase (Pearson’s rho = –0.15; p = 0.542; Figure 3(b); Table S3).
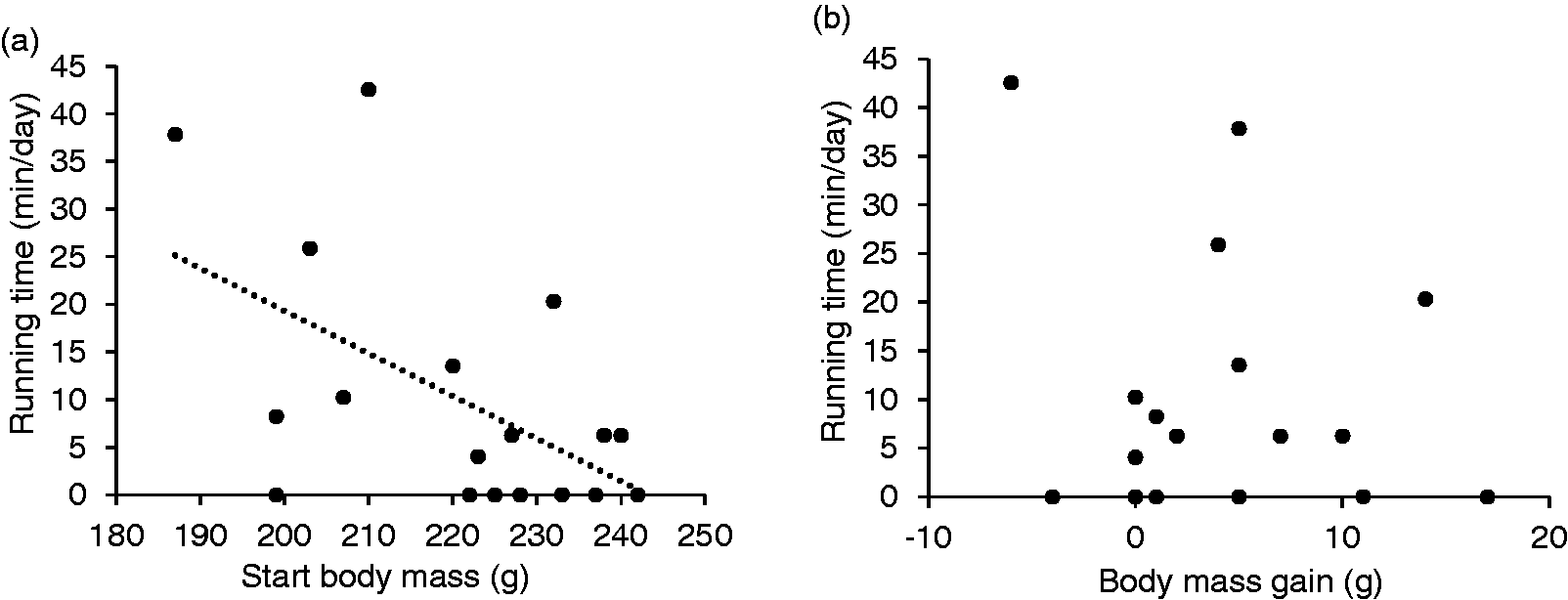
(a) Start body mass and (b) body mass gain of Wistar rats (Rattus norvegicus; N= 18) when offered access to a running plate for six days in relation to the mean running time on the plate per day. The dotted line in (a) represents a negative correlation between mean running time per day in relation the body mass at the start of the experiment (Pearson’s rho = –0.55; p = 0.019).
Diurnality
The rats’ main activity was during the dark cycle, when most bouts (mean 90% ± 9%) as well as most time spent on the running plate occurred (mean 95% ± 5%). It was also clearly visible that as soon as lights switched from dark to bright and vice versa, activity bouts decreased and increased, respectively (Figure 4). Similarly, most of the running bouts (83%) as well as most time running (90%) happened during the dark cycle. During the light cycle, sitting (43% of all bouts happened during light phase) and standing (34%) were observed most frequently, followed by running (17%), grooming (4%) and sleeping (1%). Sleeping bouts occurred also mainly during dark (94%), with mean 0.9 ± 1.6 bouts/animal per day during dark and mean 0.1 ± 0.2 bouts during the light cycle.
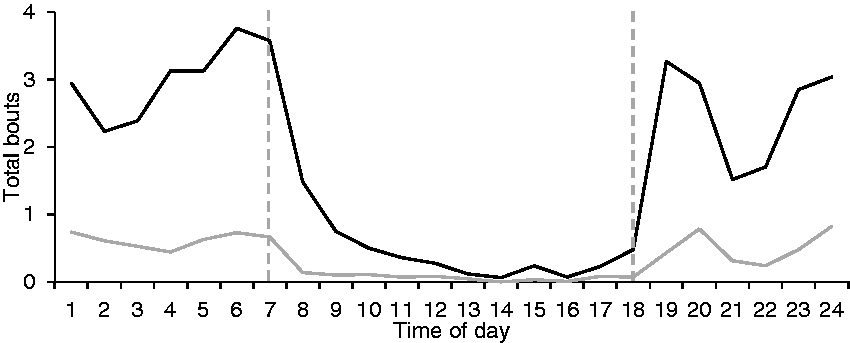
The use of a running plate by Wistar rats (Rattus norvegicus; N= 18) over six days. Data are given as mean per animal for all activities (black line) and for running (grey line). The dashed vertical lines indicate the switch between light phases (artificial light from 07:00 to 18:00, dark from 18:00 to 07:00).
Discussion
As hypothesised, the rats in this experiment made frequent use of the running plates from the first day onwards. Single events ranged from 1 s up to more than 90 min (mean 1.29 ± 5.22 min), during which time a wide range of different activities could be observed on the plate. Apart from its intended purpose (‘running’), the animals used the plate for sitting, standing, grooming, playing and much more. Sticking to their normal night-cycle behaviour, the rats mainly accessed the running plate during the night.
Study limitations
This short pilot experiment had several limitations: comparison of our results is limited to published literature on running wheels. Most of these studies measured speed and wheel turns,15,22–25 resulting in ‘distance run’ as a proxy for activity, and not number of bouts or durations, or did not have the same definition for bouts.26,27 The absence of rotation counting facilities in the present study prevented a comparison on that level. Even though it is thought that running wheels appear to be mainly used for the purpose of running, 9 because running wheel activity is usually recorded by the number of rotations, other activities observed in the present study (e.g. grooming, sleeping, sitting) are typically not documented. Additionally, the presence of other enclosure furniture may vary between studies. For the setup of our own study, we cannot say whether a less furnished enclosure, without an additional climbing structure and a pipe, would have led to a different use of the running plates. Because the main purpose of the experimental setup was not related to running plate usage, no control group was instigated.
Differences between rats in this short pilot experiment were more likely due to individual differences than to the different diet groups. To actually test a diet effect, larger groups would have to be observed over longer periods of time. Further, social compatibility between individuals could have an influence, as a low-ranked or a more anxious rat might make fewer entries on the plate, especially if a rat with a higher rank is restricting access; alternatively, those rats might have chosen to sleep on the plate rather than in the provided shelter if they wanted to distance themselves from conspecifics. Analysing the hierarchy, or the relationship between cage mates, was beyond the means of the present study but might be a valuable addition in the future. The Wistar (RjHan:WI) rat used here might not be representative for all rat strains since the use of running plates might be similarly dependent on strain characteristics as documented for running wheels. 27 As all individuals in this experiment were females, no comment can be made on potential gender differences as, for example, shown in mice. 28
Running plate use
The running plate offered the rats the opportunity to display a wide range of activities (Figure 2) and therefore could be classified as structural enrichment. The benefits of enriched housing designs have been shown repeatedly. For example, in young rats an enriched environment had a positive impact on structural brain development, 29 better performance in experimental mazes 30 and improved learning when confronted with new skills such as driving a rodent operated vehicle. 31 In older animals, previously housed in standard laboratory conditions, the animals showed slower response times in maze experiments after they were rehoused in enriched cages. 32 However, they needed fewer attempts and made fewer mistakes than the control animals to complete a task. Additionally, the mortality rate in dynamically enriched cages with changing furniture compared with normal laboratory cages and impoverished cages (isolated) was significantly lower. In further studies it would be interesting to test whether there is a preference for running plates with specific diameters, similarly to a study with wheels. 33
Running was not the most common activity performed by the rats (19% of bouts and time) on the running plate. The main activity was sitting, which might be because it often occurred between different activities. The running plate was the highest point in the cage, providing the best overview. For example, after running on the plate, the rat was sitting before it started grooming. As one might expect, sleeping had the longest bouts of all observed activities (Table 1). Playing, on the other hand, had the shortest bouts, as it contained the usual rough and tumble play with the running plate just being one of many obstacles in the cage.
The use of running plates by Wistar rats (Rattus norvegicus; N=18) for six days. The total time on plate (mean per animal ± SD), the bouts and bout length (mean per animal ± SD), the percentage of time spent running or sleeping per total time on plate, and the percentage of the respective activity during the dark cycle are given. D: day.
D: day.
Total plate use per animal increased progressively during this short pilot experiment. Time spent running remained more or less constant across days 2–6. This indicates that already after a single day, rats were comfortable using the running plate, and running had the highest proportion of the total plate use on that day (Figure 2). We did not observe any problems of coordination, where the rat would fall off the plate accidently while running. In several studies it was shown that running increased over several days and only decreased again after the animals reached a certain age.8,34 The stability of this behaviour might encourage studies to evaluate voluntary plate or wheel running as a method for determining the severity of an experimental procedure, as done in mice. 35
The most time spent running on the plate was during the dark period, as already described in a previous study performed with running wheels 26 where female rats performed more than 3500 wheel turns in the first three hours of the night, with a constantly decreasing number of turns until the light phase started again. In our study, the number of running events was more or less stable all through the night (Figure 4). A clockwise running direction was preferred, with 55% compared to 45% of events in anticlockwise direction. Despite the relatively balanced number of total bouts in both directions, it was obvious that some rats had a preferred running direction. Apart from environmental distractors (i.e. ventilation outlet, proximity of other animals, light sources), motor handedness might play a role for the directionality of running. This phenomenon, well known in humans, has also been described in rats 36 and many other animal species.37,38
Our data suggest that rats of lower body mass were more prone to running activity than rats of higher body mass (Figure 3). Voluntary wheel running was previously shown to decrease the body mass of rats regardless of the caloric intake, clearly separating the runners and non-runners. 18 As the experiment’s duration was only six days and we did not measure individual food intake, no definitive interpretation on body mass development, food intake and total running time can be made. Differences in plate use can conveniently be ascribed to differences in animal personality. 20 One potential correlate is a general propensity for activity and exploration, which could be indicated by the negative correlation of initial body mass and running time.
Considerations on running plates versus running wheels
Running in a running wheel has been described as an activity of its own that should be interpreted separately from other physical activities. 20 Compared with wheel running, running on plates is performed with a lateral directional axis and should theoretically be a more difficult task, as the animal must find a right spot to balance and perform without moving its centre of gravity. However, no coordination problems were observed in our animals. By running in wheels, animals can develop physical transformation due to running in a lordosis-like position, and even tails bent upwards might occur in some mice. 15 Whether a similar transformation might occur in animals that use plates only in a single direction remains to be investigated.
As there are no further studies with running plates, it is unclear to what degree plates are comparable to wheels. One important difference might be that the field of vision for the running animal is different. In a closed running wheel, the runner only sees the upcoming, more or less evenly downward-moving track. As this is hard to focus on visually, this might lead to a trance-like state and therefore allows the rat to exit the ‘real world’, creating an experience apart from other activities. With a running plate, this effect should not exist, as the fixed, unmoving surroundings of the enclosure can be observed at all times. The optical impression is likely one with a small part of moving substrate with the main field of vision consisting of static objects. In anthropomorphising words, an animal might use a running wheel for both the physical exercise and a sensory experience that is different (but remains to be objectively qualified) from any other experience it can get in its cage. By contrast, it may use a running plate only for the physical activity. To test whether this interpretation is valid, several experiments could be conceived: preference tests could determine to what degree animals prefer self-driven running wheels to a similar sensual experience that does not involve activity (e.g. a wheel that starts rotating once the animal occupies a fixed platform in the wheel); or the effect of conventional running plates compared with partially walled running plates, where the animal also creates a specific visual experience by its own activity.
Conclusion
Running plates are a readily accepted structural enrichment for laboratory rats. In this experiment, they were used by the rats not only for running but also for a range of other activities, e.g. grooming, sitting, standing, playing and even sleeping, with inter-individual differences. Whether they have more effect as structural enclosure furniture than as facilitators of running exercise, and to what degree they differ from running wheels, requires further investigation.
Supplemental Material
sj-pdf-1-lan-10.1177_00236772211036572 - Supplemental material for Use of running plates by floor housed rats: A pilot study
Supplemental material, sj-pdf-1-lan-10.1177_00236772211036572 for Use of running plates by floor housed rats: A pilot study by Jessica Frei, Marcus Clauss, Daniela E Winkler, Thomas Tütken and Louise F Martin in Laboratory Animals
Footnotes
Acknowledgements
We thank Lukas Sprenger for setting up the video equipment and lending technical support; Sandra Heldstab, Kathrin Zbinden and Annelies De Cuyper for support in animal husbandry; and Maria José Duque-Correa for the photographs.
Author contribution
MC and TT designed the experiment and obtained funding. DW and MC performed the animal experiment. JF analysed the video footage. LFM and JF wrote the manuscript with input from all co-authors.
Data accessibility statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The long-term feeding experiment was funded by the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (ERC CoG grant agreement No 681450) to TT.
Supplemental material
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.