
Abstract
Application of immunogold cytochemistry revealed polar (asymmetric) distribution of GLUT-1 in mouse brain microvascular endothelia, representing the anatomic site of the blood-brain barrier (BBB). This polarity was manifested by an approximately threefold higher immunolabeling density of the abluminal than the luminal plasma membrane of the endothelial cells. The immunoreaction for GLUT-1 in nonbarrier continuous (skeletal muscle) or fenestrated (brain circumventricular organs) microvascular endothelial cells was absent. In the choroid plexus, the basolateral plasmalemma of the epithelial cells was labeled more intensely than the vascular fenestrated endothelium. Addition of morphometry to the applied immunogold technique makes it possible for even subtle differences to be revealed in the density of immunolabeling for GLUT-1 in blood microvessels located in four brain regions. We found that the density of immunosignals in the microvessels supplying the cerebral cortex, hippocampus, and cerebellum was essentially similar, whereas in the olfactory bulb it was significantly lower. Asymmetric distribution of GLUT-1 in the endothelial plasma membranes presumably leads to a reduced concentration of glucose molecules in the endothelial cells compared to blood plasma and also secures their more rapid transport across the abluminal plasmalemma to the brain parenchyma. (J Histochem Cytochem 47:1021–1029, 1999)
Keywords
G
Evidence indicates that GLUT-1 is associated mainly with the microvascular endothelial lining (Goldstein et al. 1977; Hawkins et al. 1983; Harik et al. 1990; Farrell and Pardridge 1991). Detailed electron microscopic examination showed that immunoreactive GLUT-1, revealed by gold particles, is scattered irregularly throughout the cytoplasm of the brain blood microvascular endothelial cells (ECs) and is distributed asymmetrically in their plasma membranes (Dick and Harik 1986; Farrell and Pardridge 1991). This asymmetry is evidenced by the higher density of specific immunolabeling of abluminal plasma membrane than luminal plasma membrane of the vascular ECs (Carruthers 1990; Crone 1965).
The problem of the distribution of GLUT-1 in the brain microvascular network, however, appears to be rather complex in view of several reports revealing regional variations in glucose utilization by brain parenchyma. These differences in the utilization of glucose are believed to reflect different degrees of glucose availability, as suggested by some authors (Sharp 1976; Bagley et al. 1989; Gould and Holman 1993; Cornford et al. 1995). Therefore, it was postulated that the physiological heterogeneity of the microvascular bed (with respect to glucose transport) could affect the uptake of glucose and, ultimately, the metabolic and functional activity of different regions of the brain (Robinson and Rapoport 1986; Kasanicki et al. 1989). It is noteworthy that the regional heterogeneity of the brain microvascular network in its barrier function to endogenous albumin was also noted in the course of our quantitative immunocytochemical studies (Vorbrodt et al. 1995; Ueno et al. 1996).
Taking these data into consideration, we decided to perform an immunocytochemical study, the main objective of which was to clarify the issue of whether or not any differences exist in the distribution of GLUT-1 in the vascular bed of four different mouse brain regions: cerebral cortex, hippocampus, cerebellum, and olfactory bulb. The choice of these brain regions and the application of quantitative immunocytochemical procedures were based on our studies on the BBB function to endogenous albumin (Vorbrodt et al. 1995, 1997).
Materials and Methods
Animals
Twelve adult (6–8-month-old) Balb/c mice of both sexes, weighing 25–30 g, were used. Under sodium pentobarbital anesthesia, mice were perfused through the heart with freshly prepared fixatives for approximately 15 min. After perfusion, brains were removed, sectioned on a coronal plane, and samples of the parietal cerebral cortex, hippocampus (medial and rostral regions), cerebellum, and olfactory bulb were dissected and immersion-fixed in the same fixative at room temperature (RT; 22C). Similar tissue samples from three animals were immersion-fixed only for comparison.
After fixation (up to 2 hr), the tissue blocks were washed overnight in ice-cold 7.5% sucrose, dehydrated in ethanol with concomitant lowering of temperature to −35C, and finally embedded in hydrophilic resin Lowicryl K4M. After polymerization under a UV lamp in the low-temperature embedding apparatus TTP 010 (Balzers Union; Balzers, Liechtenstein), the samples were cut with a diamond knife on an ultramicrotome (Sorvall MT-5000; DuPont Boston, MA).
For comparative purposes, samples of skeletal muscle (diaphragm) and brain regions supplied with nonbarrier type blood vasculature (circumventricular organs), such as choroid plexus, subfornical organ, and hypophysis (adenohy-pophysis), were also used.
Because, according to the opinions of several authors, the antigenic properties and immunoreactivity of GLUT-1 can be compromised by glutaraldehyde even when used at low concentration (Gerhart et al. 1989; Farrell and Pardridge 1991; Cornford et al. 1993), the following fixative solutions were used in the course of our study.
Fixative PLP was prepared according to McLean and Nakane (1974), containing 0.01 M sodium meta-periodate, 0.075 M lysine, and 3% formaldehyde (freshly prepared from paraformaldehyde) in 0.07 M phosphate buffer, pH 6.6.
Fixative 3P0.03G contained 3% formaldehyde, 0.03% glutaraldehyde, 0.002 M CaCl2, and 0.1 M sucrose in 0.05 M cacodylate buffer, pH 7.2.
Fixative 2P0.1G contained 2% formaldehyde, 0.1% glutaraldehyde, 0.002 M CaCl2, and 0.15 M sucrose in 0.05 M phosphate buffer, pH 7.2.
Immunocytochemical Procedure
Ultrathin sections attached to Formvar-carbon-coated nickel grids were placed in a moist chamber at RT on a drop of 0.1 M glycine for 10 min to quench free aldehyde groups. The sections were incubated in a refrigerator (4C) overnight (approximately 16 hr) in a drop of solution of rabbit anti-GLUT-1 antiserum (Chemicon; Temecula, CA) diluted 1:200 with PBS containing 0.5% ovalbumin. After washing with PBS, the sections were exposed for 1 hr at RT to goat-anti-rabbit antiserum (AuroProbe EM; Amersham Life Science, Arlington Heights, IL) complexed with two different colloidal gold particles of 10-nm or 15-nm diameter (GAR-G10 or GAR-G15, respectively) diluted with PBS (1:25). After several washings in PBS and distilled water, and staining with 4% uranyl acetate (3 min) and Reynold's lead citrate (1 min), the sections were examined in a Hitachi 7000 electron microscope.
Controls for the specificity of the immunostaining consisted of sections incubated with normal rabbit preimmune serum or with secondary antibodies (GAR-G10 or GARG15) only.
Quantitative Analysis
Randomly sampled capillary profiles were examined in representative sections of brain regions chosen for this study. Sections labeled with 15-nm gold particles, i.e., after incubation with GAR-G15, were used for morphometric analysis. The density of labeling associated with luminal and abluminal plasma membranes (LPM and APM, respectively) of the microvascular ECs was expressed as the number of gold particles (GPs) per μm of EC plasmalemmal length.
Morphometry was performed with Sigma Scan software packages on a PC 486 DX equipped with a Jandel JS-2 tablet. Mean values, standard deviation (SD), and standard error (SEM) were calculated. Analysis of variance (ANOVA) and post hoc comparisons of means (Bonferroni procedure) were performed for statistical evaluation of the results (CSS program). Significance levels for differences in mean values between all experimental groups were accepted as p <0.05.
Results
Effect of Fixation on Density of Immunolabeling for GLUT-1 in Walls of Microvessels
Before starting our comparative study of different brain regions, we decided to determine the optimal conditions for immunolabeling of brain microvascular endothelium. For this purpose, the sections of cerebral cortex samples fixed in three different fixatives were processed for immunodetection of GLUT-1 using the same incubation media and the same steps of the entire immunocytochemical procedure.
In the samples of cerebral cortex fixed in PLP, the preservation of antigenicity of GLUT-1 appeared to be the best. It was manifested by relatively uniform and dense immunolabeling of both luminal and abluminal plasma membranes and the cytoplasm of the vascular ECs. Because the preservation of ultrastructural details of the components of the vascular wall was poor, the endothelial plasma membranes were hardly visible and the outlines of the ECs were fuzzy, as shown in Figure 1.
After fixation in 3P0.03G fixative, the immunolabeling was slightly less intense but the preservation of ultrastructural details was better, especially of membranous structures, including the EC plasma membranes (Figure 2). The brain samples fixed in 2P0.1G fixative showed good preservation of ultrastructural details, but some segments of the vascular endothelial lining were weakly immunolabeled (Figure 3).
Relatively strong labeling, irrespective of the fixative used, was observed in the area of interendothelial junctional complexes. Whereas the labeling of the EC cytoplasm in all specimens was not uniform, the distribution of immunosignals for GLUT-1 in the EC plasma membranes was always characterized by markedly higher density of immunolabeling of the abluminal than the luminal plasmalemma. No notable differences in the preservation of ultrastructural details or in the intensity of immunolabeling for GLUT-1 were observed in brain samples fixed by perfusion or by immersion.
The morphological observations described above were supplemented with morphometric (quantitative) evaluation of the immunolabeling density distribution in the capillary profiles. The results of this introductory quantitative and statistical analysis are presented in Table 1.
It appears that the application of different fixatives did not affect the typical asymmetric distribution of GLUT-1 in the vascular endothelial lining. In all specimens of cerebral cortex examined, the ratio of the labeling density of the luminal to abluminal plasma membranes was approximately 1:3.
The results of these pilot experiments prompted us to use in our further study the tissue samples fixed in 3P0.03G fixative only. Because of great variations and absence of uniformity in the distribution of immunosignals in the EC cytoplasm with high labeling of the interendothelial junctional complexes, morphometric analysis was performed on the endothelial plasma membranes (luminal and abluminal plasmalemma) only.
Immunolabeling of Endothelial Cells of Nonbarrier Blood Microvessels
The immunosignals for GLUT-1 in the choroid plexus were mainly associated with the undulated basolateral surface of the epithelial cells. The intercellular junctions and apical microvilli of these cells were not labeled. The fenestrated endothelium of the blood vesels was weakly labeled, with a few irregularly scattered gold particles over the luminal front of the thick portions of the ECs (Figure 4). In general, more immunosignals were associated with the surface of epithelial than of endothelial cells.
In the blood microvessels of the hypophysis, lined with fenestrated endothelium, no immunolabeling was present (Figure 5). Scattered single gold particles were occasionally attached to the surface of perivascular cells.
The fenestrated endothelial lining of blood vessels located in the subfornical organ also did not show any immunolabeling for GLUT-1, but several gold particles were scattered on the perivascular membranous cell processes (Figure 6).
The continuous endothelium of the skeletal muscle capillaries remained unlabeled, whereas solitary gold particles appeared occasionally over the muscle fibers (Figure 7).
Comparison of the Labeling Density Distribution in the BBB Vasculature in Different Brain Regions
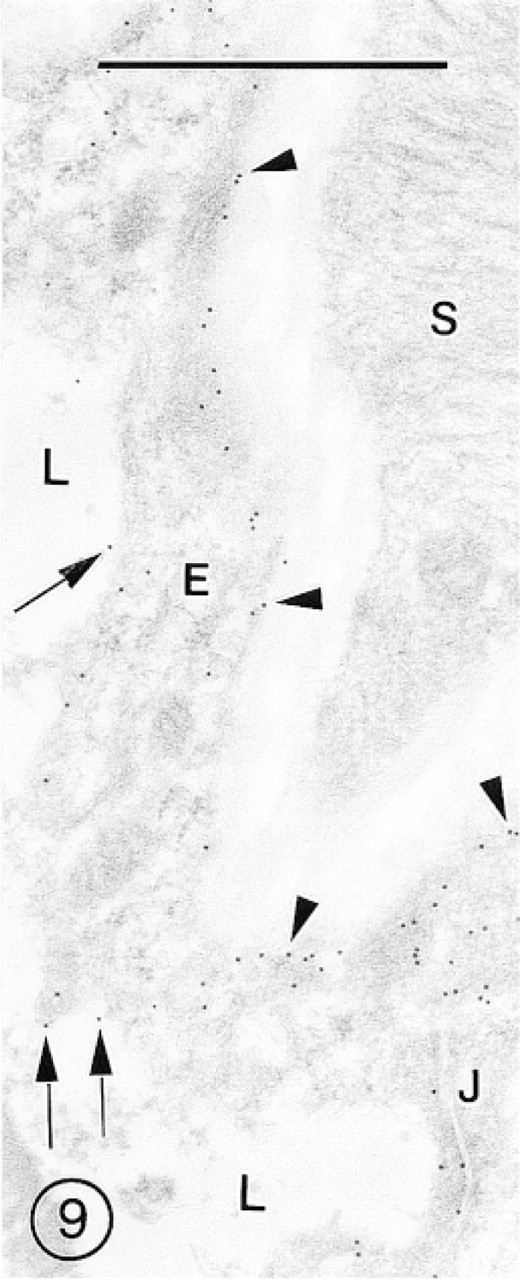
Cerebral Cortex. The distribution of gold particles in the blood microvessels of the cerebral cortex is shown in Figures 1–3. The most reproducible feature of the endothelial lining of these vessels was higher labeling of the abluminal than of the luminal plasmalemma. The interendothelial tight junctions were always decorated with several gold particles (Figures 3 and 8). The labeling of the EC cytoplasm was irregular, i.e., some segments showed lack of label, whereas others were immunostained (Figures 1, 3, and 8). A similar pattern was observed in postcapillary venules (Figure 2). In the arterioles, immunolabeling for GLUT-1 was associated with the endothelium but was absent in the subendothelial basement membrane and in smooth muscle cells (Figure 9).
Morphometric analysis of electron micrographs revealed that the labeling density of the EC abluminal plasmalemma was approximately three times higher than that of the luminal plasmalemma (Table 2).
Quantitative evaluation of the labeling density did not reveal any significant differences between blood microvessels from the cerebral cortex and those from the hippocampus (Table 2).
Although quantitative analysis did not show significant differences in immunodensity between cerebellar vasculature and other brain regions, it did show slightly increased labeling of the EC abluminal plasmalemma (Table 2).
Comparison of the labeling density distribution for GLUT-1 in the capillaries of mouse brain cortex after fixation in three different fixatives
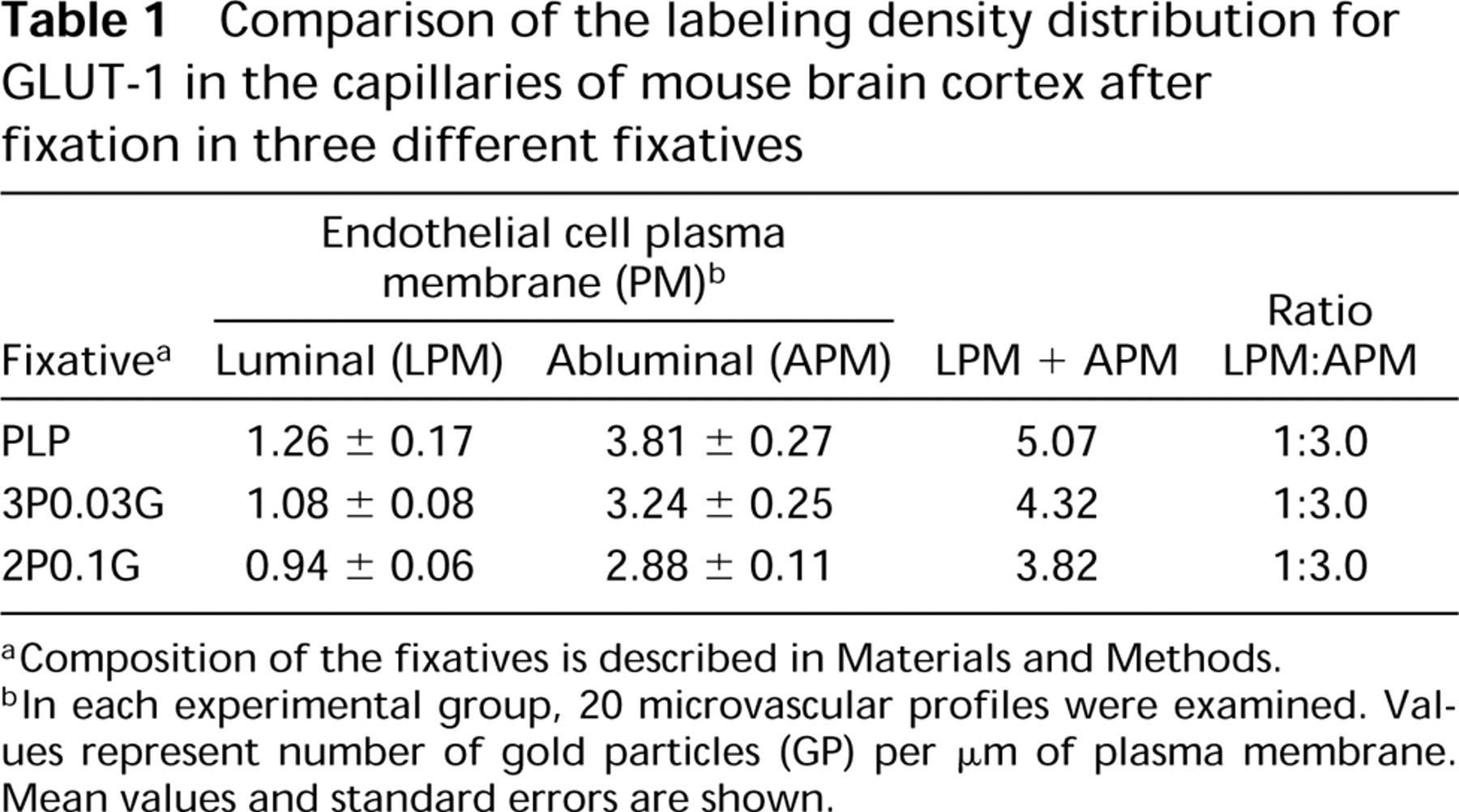
Composition of the fixatives is described in Materials and Methods.
In each experimental group, 20 microvascular profiles were examined. Values represent number of gold particles (GP) per μm of plasma membrane. Mean values and standard errors are shown.
Morphometric analysis revealed that the mean value of the labeling density for GLUT-1 was significantly lower in the vascular profiles of the olfactory bulb than in other brain regions under study (Table 2).
Discussion
The main findings of these studies were as follows. (a) A short fixation time (up to 2 hr) and a low concentration of glutaraldehyde (up to 0.03%) in paraformaldehyde-containing fixative, followed by embedding of tissue samples at low temperature in hydrophilic resin, are important factors for preservation of both GLUT-1 antigenicity and structural details of the components of the vascular wall. (b) In the mouse brain, the distribution of immunoreactivity for GLUT-1 in BBB-type vasculature is asymmetric, as manifested by approximately threefold higher immunolabeling of abluminal than of luminal plasmalemma of the EC. In non-BBB-type vasculature (brain cirumventricular organs and skeletal muscle), immunoreactivity for GLUT-1 in the vascular endothelium is negligible or totally absent. (c) Despite the existence of slight variations, the density of immunosignals for GLUT-1 in the BBB microvessels supplying cerebral cortex, hippocampus, and cerebellum was essentially similar, whereas in the olfactory bulb it was significantly lower. These points deserve additional comments and discussion.
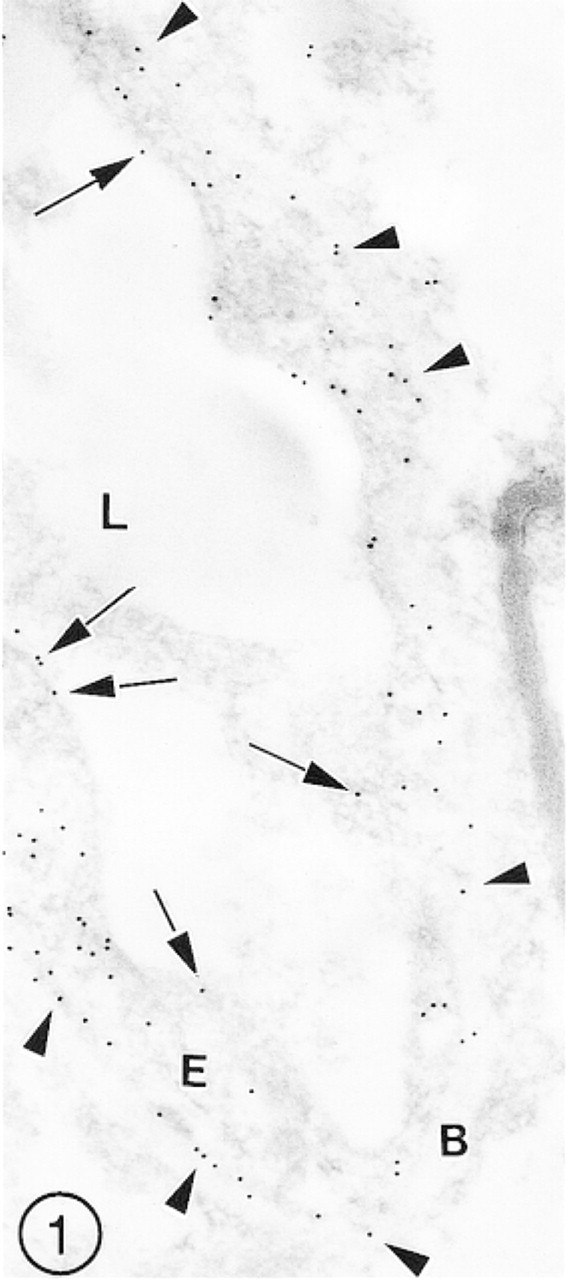
A cross-sectioned capillary in mouse cerebral cortex fixed in PLP fixative. Several immunosignals (gold particles) for GLUT-1 are scattered over the endothelial luminal plasmalemma (arrows), and even more are associated with the barely visible abluminal plasmalemma (arrowheads). Several gold particles are also scattered irregularly over the endothelial cell (E) cytoplasm.
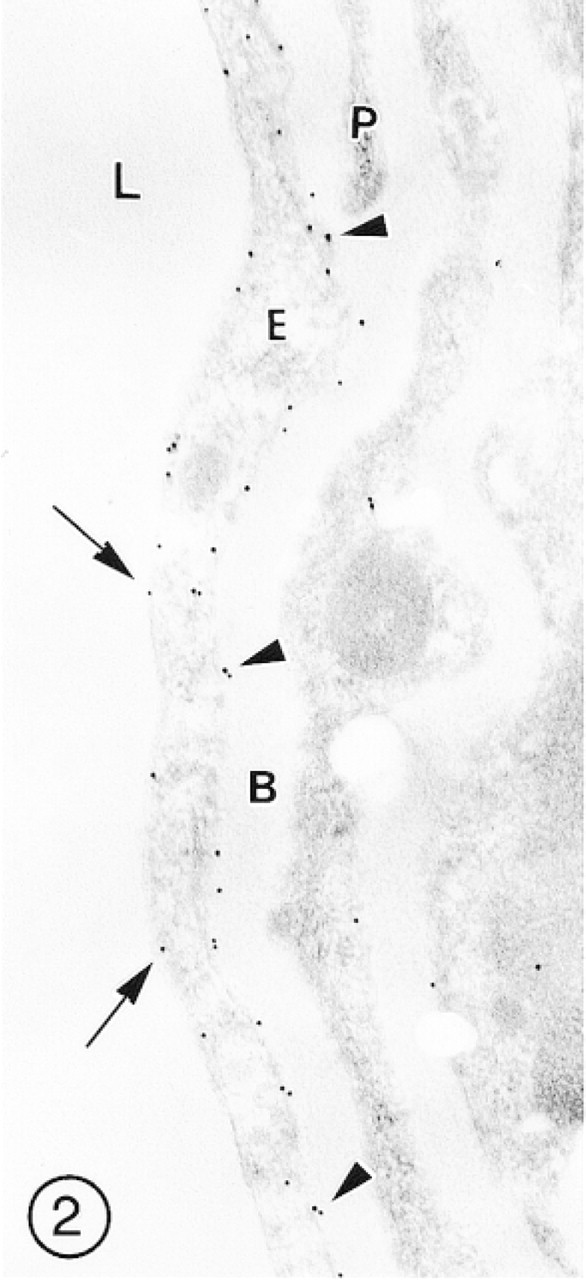
A longitudinal section of the wall of a blood microvessel (presumably a precapillary venule) in the cerebral cortex fixed in 3P0.03G fixative. Immunosignals for GLUT-1 are more numerous in clearly visible abluminal (arrowheads) than in luminal (arrows) plasmalemma of the endothelial cell (E). Pericytes (P) and the vascular basement membrane (B) remain unlabeled.
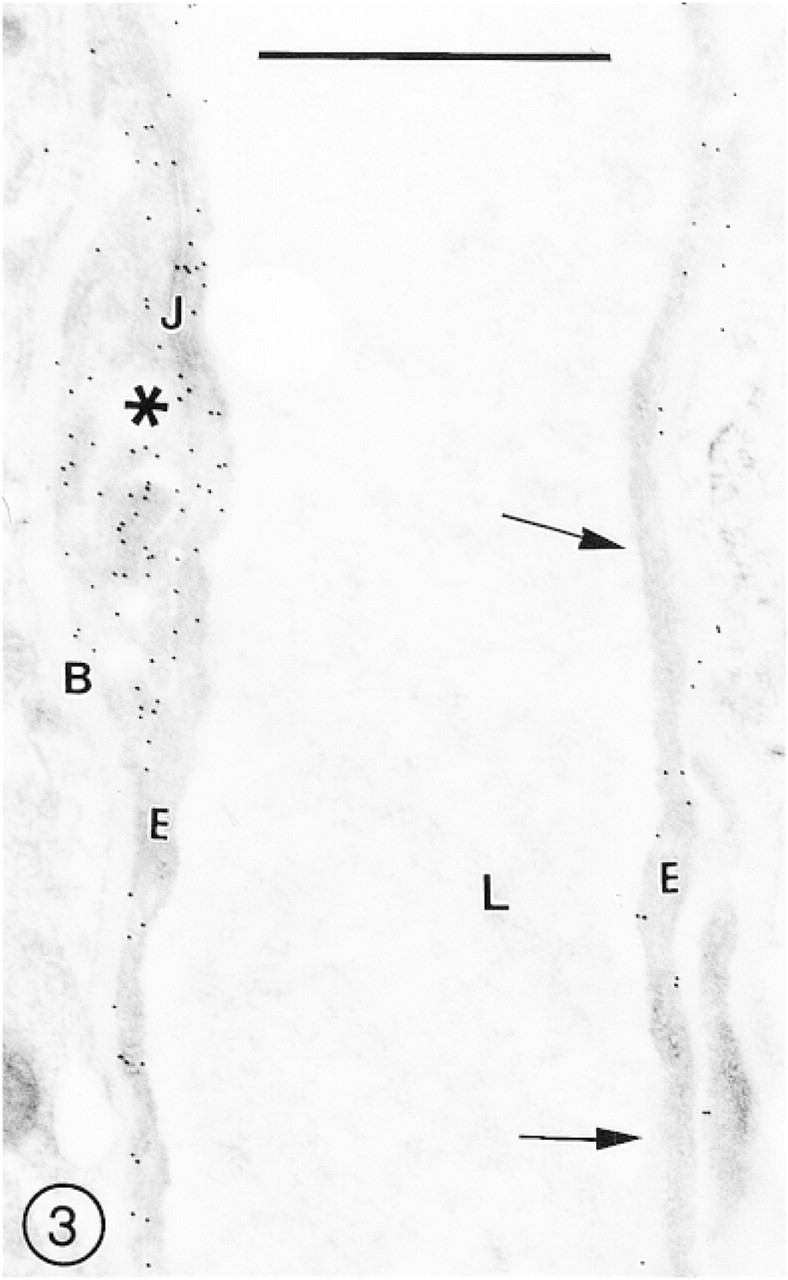
This longitudinally sectioned capillary of the cerebral cortex fixed in 2P0.1G fixative is a good example of uneven immunolabeling for GLUT-1 of the microvascular endothelium. At left, a high density of immunosignals over the EC body (asterisk) is presumably associated with the interendothelial junctional complex (J). In contrast, at right, long segments of the endothelial cell (E) remain unlabeled (arrows).
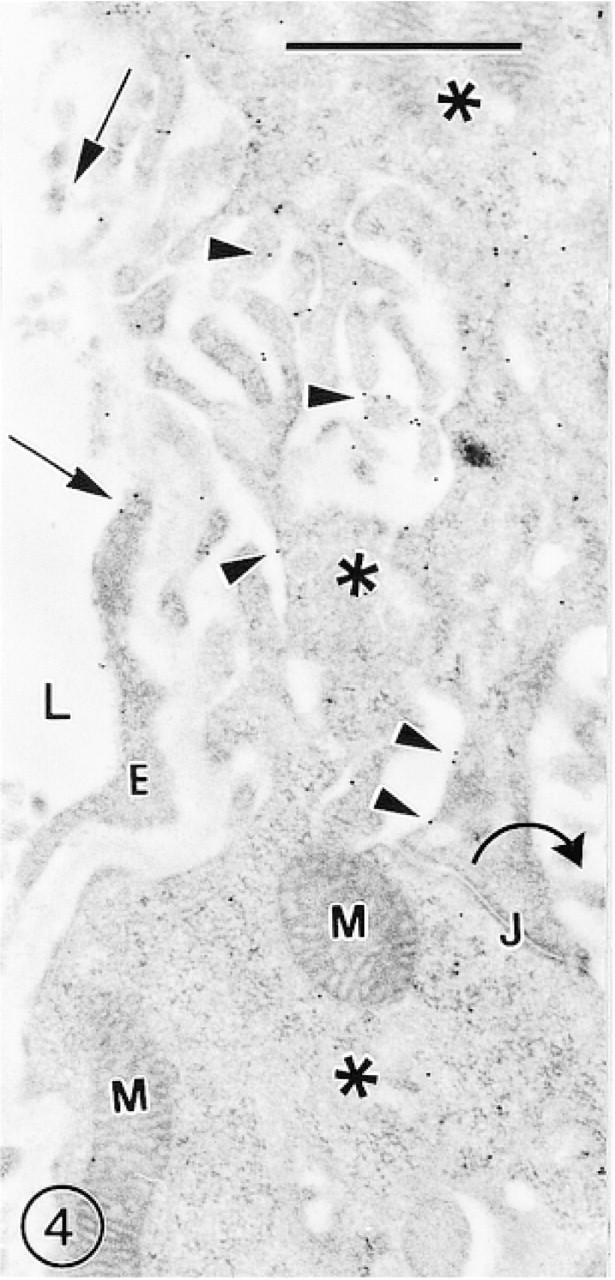
A section of the mouse choroid plexus showing many immunosignals for GLUT-1 (arrowheads) scattered over the undulated basolateral surface of the epithelial cells (asterisks). The apical surface with many microvilli (curved arrow) remains unlabeled. The tangentially sectioned vascular fenestrated endothelium is occasionally labeled with a few gold particles (arrows).
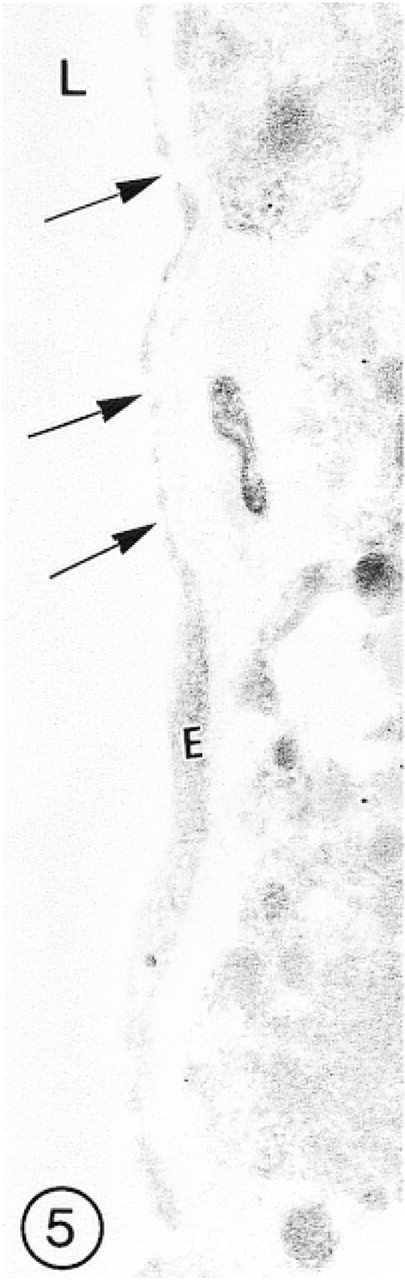
Fenestrated endothelium (E) of the capillary of hypophysis, showing absence of immunoreaction for GLUT-1 (arrows).
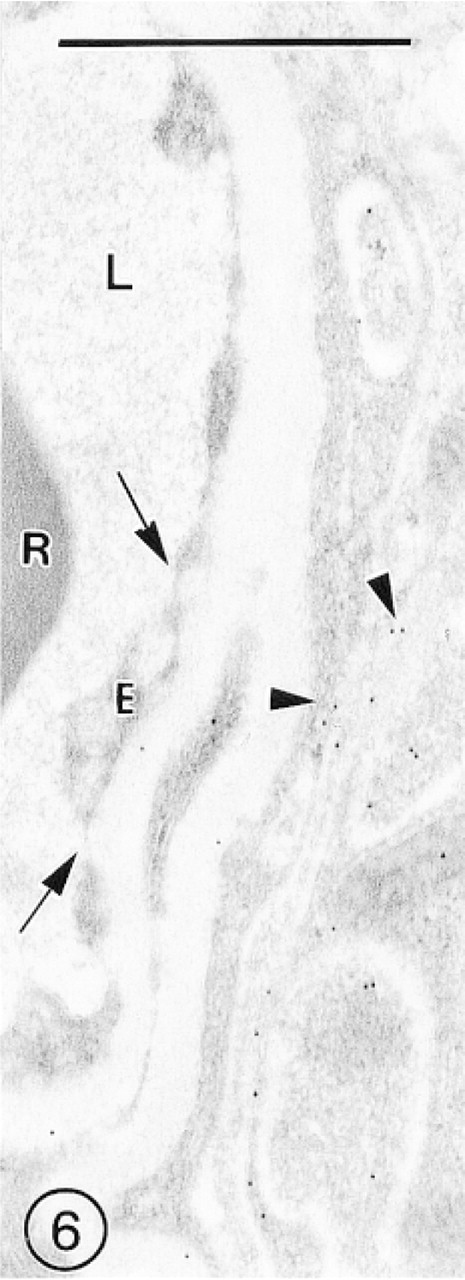
A capillary located in the area of the subfornical organ lined with the fenestrated endothelium (E) shows complete absence of immunolabeling for GLUT-1 (arrows). A few immunosignals are associated with the membranes of perivascular cells (arrowheads).
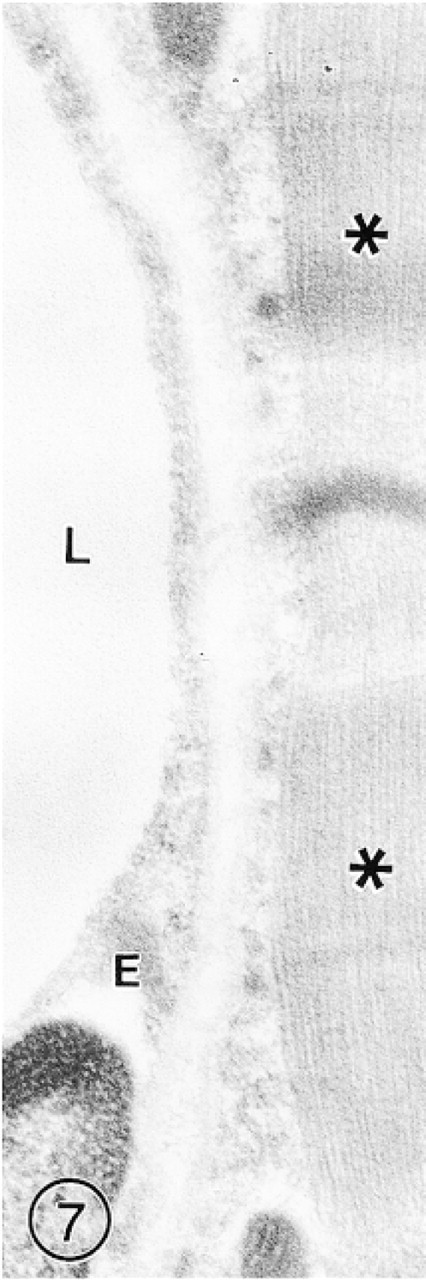
In a section of skeletal muscle, immunoreaction for GLUT-1 is absent in the capillary endothelium (E). Only solitary gold particles are occasionally present over the muscle fibers (asterisks). Bars = 1 μm.
The short fixation time (by perfusion or immersion) of tissue samples in the gentle fixatives, with concomitant omission of osmium tetroxide, is not optimal for good preservation of ultrastructural details of cellular components, especially those of membranous character, such as plasmalemma, plasmalemmal vesicles, and vacuoles. The use of such fixatives, based mainly on paraformaldehyde, for immunocytochemical detection of various antigens with electron microscopy was postulated long ago (McLean and Nakane 1974). The addition of glutaraldehyde usually significantly improves the quality of fixation but appears to be harmful to sensitive antigens. For immunodetection of GLUT-1, Farrell and Pardridge (1991) suggested the use of as low a concentration of glutaraldehyde as 0.01% which, in their opinion, appeared to be better than PLP. Cornford et al. (1993, 1995, 1998) applied concentrations between 0.01% and 0.05%, and Gerhart et al. (1989) used only glutaraldehyde-free PLP fixative. Taking all these data into consideration, three different fixatives were applied during the first stage of this study. 3P0.03G fixative appeared to be the best, securing antigenic properties of GLUT-1 and sufficiently preserving the cytoplasmic membranous structures, making the outlines of the ECs clearly visible. Therefore, all our observations presented in Figures 4–15, as well as morphometric analysis (Table 2), relate to tissue samples fixed in 3P0.03G fixative.
Both morphological and morphometric analysis demonstrated the asymmetry of distribution of GLUT-1 in the endothelium of BBB-type mouse brain microvessels, expressed by the ratio of the labeling density of luminal to abluminal plasmalemma equaling about 1:3. During pilot observations, this ratio was not changed by the application of three different fixatives, as shown in Table 1. In the four brain regions examined, the ratio ranged from 1:2.5 in the hippocampus to 1:3.2 in the cerebellum. These values are roughly similar to those obtained by Cornford et al. (1993) in newborn and adult rabbits (ratio approximately 1:3). They are also similar to, although slightly lower than, those presented by Farrell and Pardridge (1991), which were obtained in rat brains fixed by perfusion (ratio about 1:3) or by immersion (ratio about 1:4).
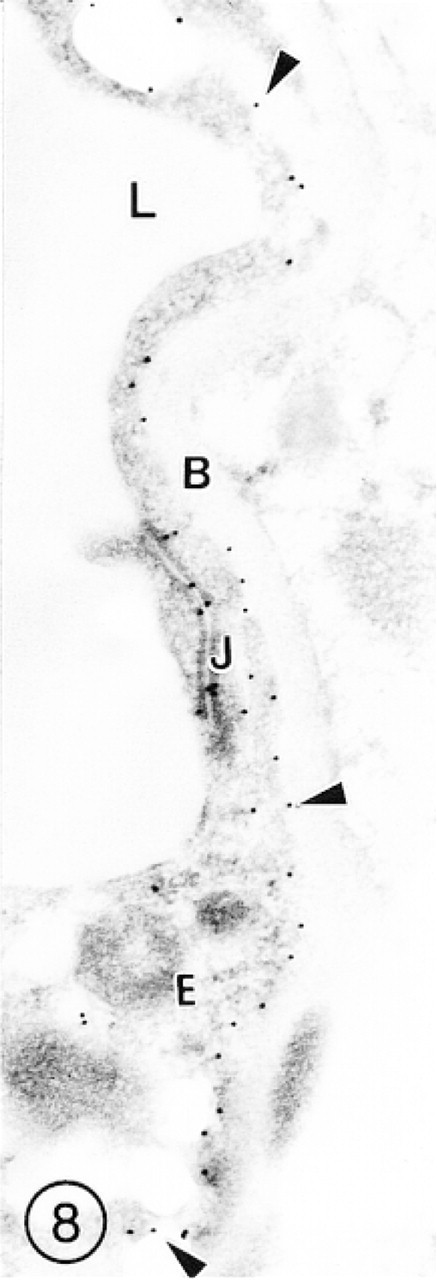
In this capillary from the mouse cerebral cortex, immunosignals for GLUT-1 are scattered mainly along the endothelial cell abluminal plasmalemma (arrowheads). Several gold particles are also associated with the interendothelial tight junction (J). Neither the basement membrane (B) nor the perivascular neuropil is immunolabeled.
In this cross-sectioned arteriole from cerebral cortex, both luminal (arrows) and abluminal (arrowheads) plasma membranes of the endothelial cell (E) are intensely labeled with small (GAR-G10) gold particles. Both subendothelial basal lamina and smooth muscle cells (S) remain unlabeled.
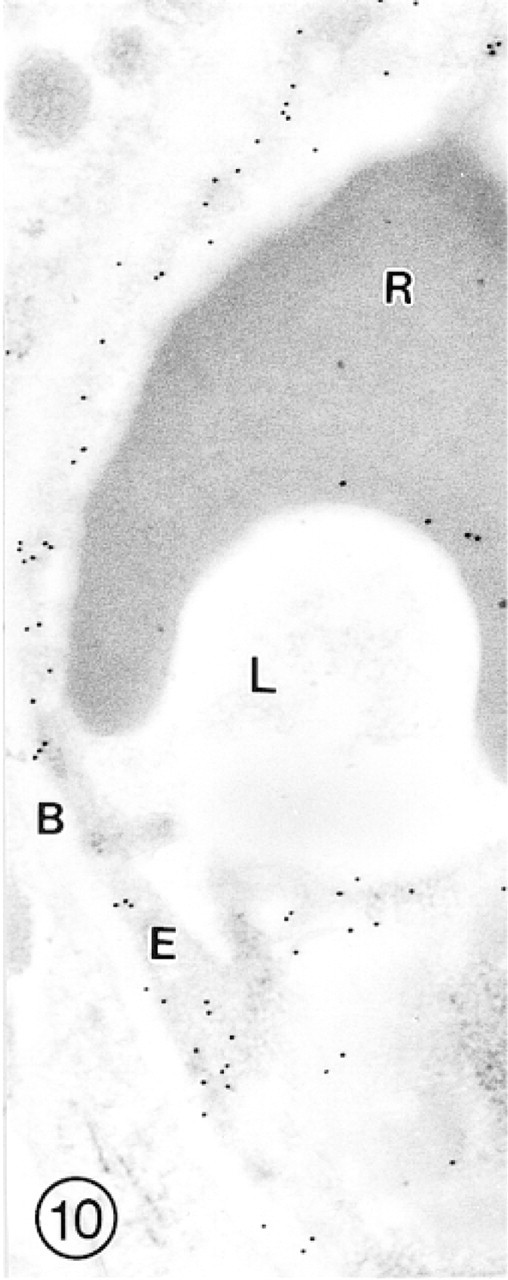
In this cross-sectioned capillary from hippocampus, immunosignals are scattered throughout the endothelial lining (E) and also over the red blood cell (R).
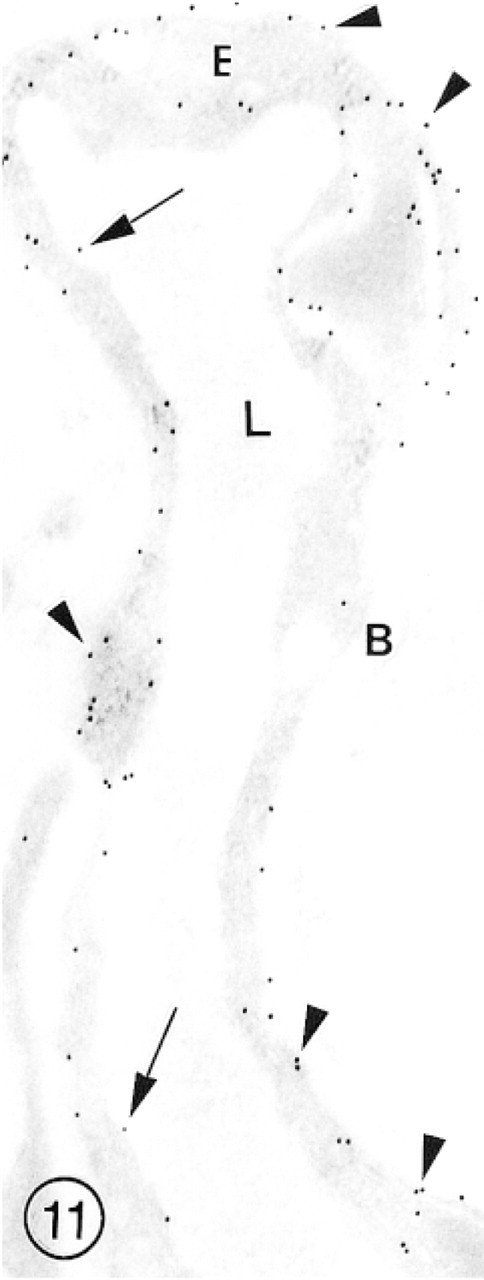
In this slightly collapsed capillary from hippocampus, intense immunolabeling for GLUT-1 is associated with luminal (arrows) and abluminal (arrowheads) plasma membranes of the endothelial cell. A few gold particles are also scattered throughout the endothelial cell (E) cytoplasm.
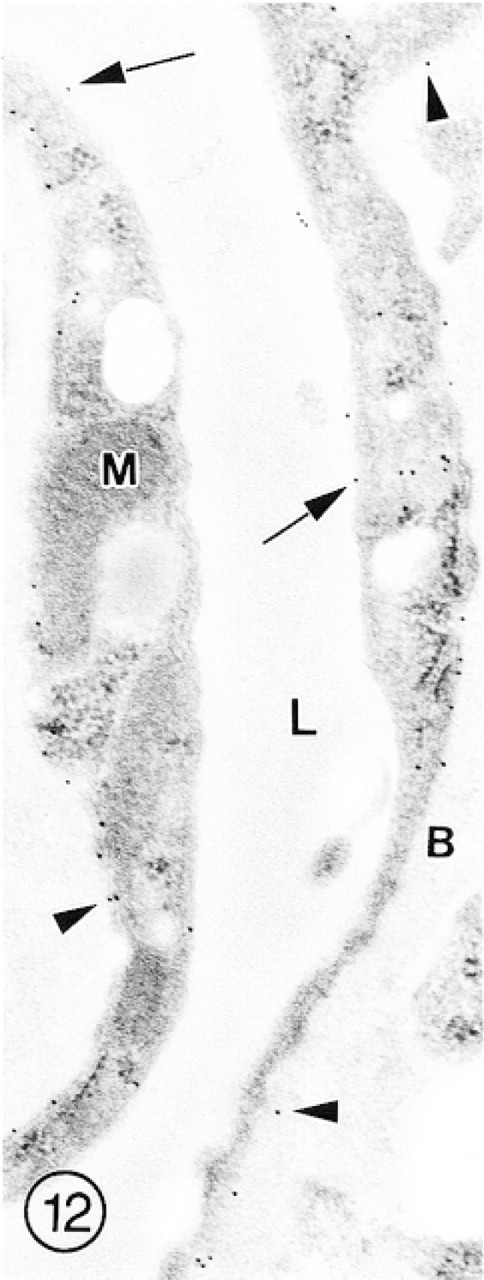
In this longitudinally sectioned capillary from the molecular layer of the cerebellum, immunolabeling for GLUT-1 is of rather low intensity on both luminal (arrows) and abluminal (arrowheads) plasma membranes of the endothelial cell.
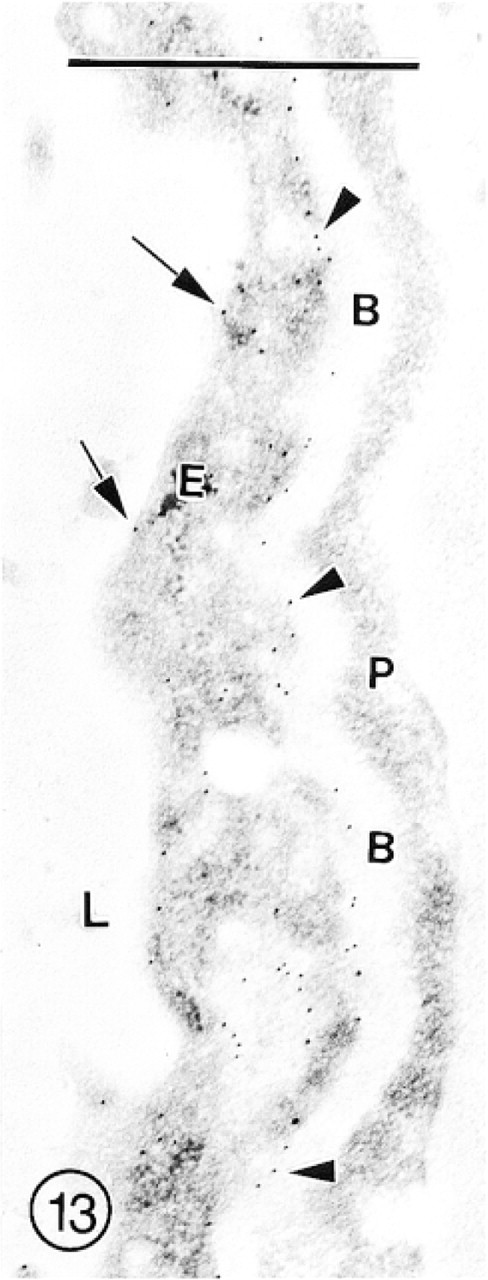
A portion of the capillary from the granule cell layer of cerebellum, showing rather dense labeling with GAR-G10 associated with the endothelial cell abluminal plasmalemma (arrowheads), and less intense labeling of the luminal (arrows) plasmalemma. In the endothelial cell cytoplasm, a few lucid vacuoles, some of which are labeled with several gold particles, are present. Subendothelial basement membrane (B) and pericytes (P) are not labeled.
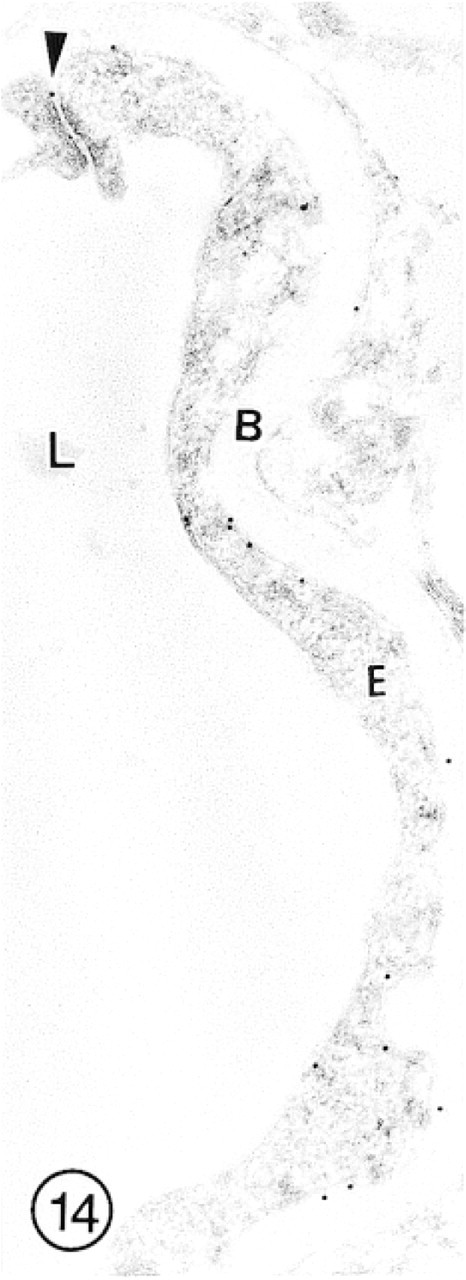
In this capillary from the internal granular layer of olfactory bulb, immunolabeling for GLUT-1 is of low intensity. Even the interendothelial tight junction is labeled with only a single gold particle (arrowhead).
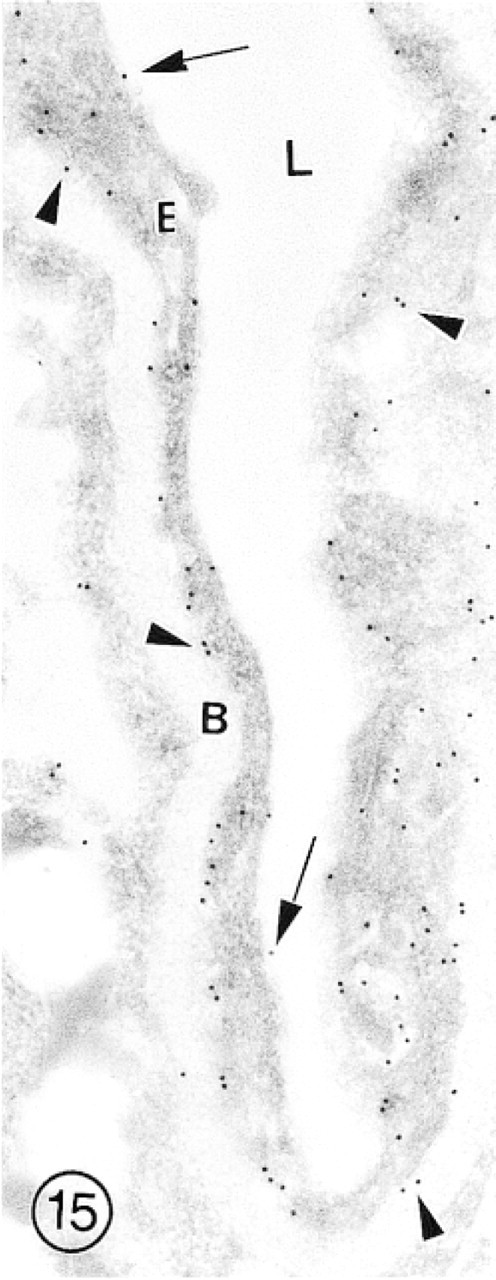
This slightly collapsed capillary from the external granular layer of olfactory bulb shows relatively intense immunolabeling for GLUT-1 associated with the EC luminal (arrows) and abluminal (arrowheads) plasma membranes. Several gold particles are also scattered throughout the cytoplasm of the endothelial cell (E). Bars = 1 μm.
The functional meaning of asymmetric distribution of GLUT-1 molecules in the brain vascular endothelium is not yet fully known. It is assumed that, when transported from the blood plasma to the brain interstitium, glucose at first faces the EC luminal plasma membrane which, because of the lower number of transporter molecules, limits the intensity of this influx of glucose. According to Farrell and Pardridge (1991), a higher number of GLUT-1 molecules in the abluminal plasmalemma may reduce the concentration of glucose in the ECs compared to blood plasma. One can expect that the greater concentration of transporters secures a faster transport of glucose from the EC body (efflux) across the abluminal plasmalemma to the adjacent neuropil. This structural asymmetry is paralleled by the kinetic asymmetry of the transporter molecules, as suggested by Gould and Holman (1993). According to these authors, the net influx of glucose is lower than the net efflux, thus allowing GLUT-1 to function effectively as a unidirectional transporter. The observed asymmetry appears to be one more piece of morphological evidence of the previously proposed functional polarity of the brain endothelium representing the BBB (Vorbrodt 1993). It appears that, according to data presented by Gerhart et al. (1989), the immunolabeling density for GLUT-1 in the brains of dog and human is equally distributed in the EC luminal and abluminal plasma membranes, with some accumulation in the area of tight junctions. This discrepancy requires experimental elucidation with application of the newest quantitative immunogold techniques.
Comparison of labeling density distribution for GLUT-1 in four brain regions of adult mice
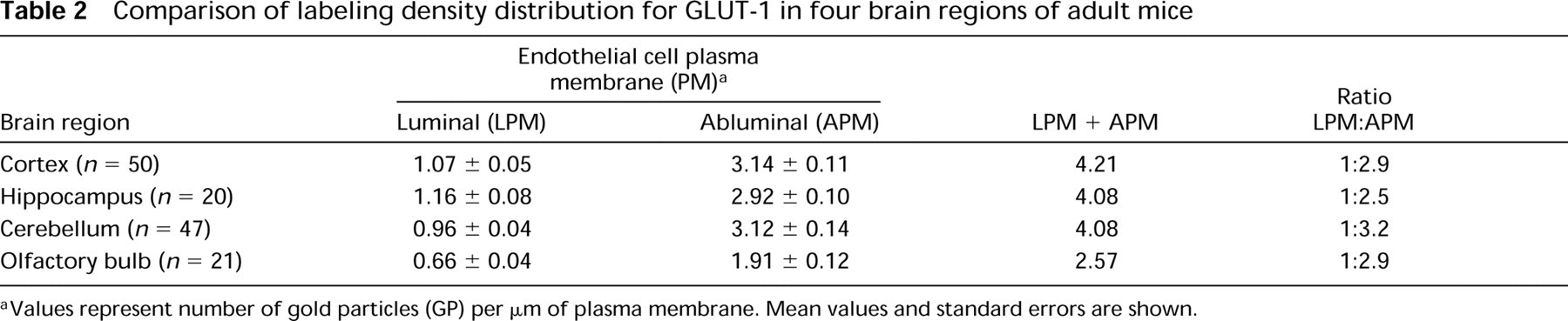
Values represent number of gold particles (GP) per μm of plasma membrane. Mean values and standard errors are shown.
It is believed that the intraendothelial immunosignals indicate the presence of GLUT-1 molecules in the cytoplasmic compartment, presumably serving as a reserve that can be recruited to the plasma membranes on stimulation of glucose transport to the brain (Carruthers 1990; Farrell and Pardridge 1991; Yu and Ding 1998). However, no convincing and reasonable explanation has yet been provided for the functional significance of GLUT-1 associated with interendothelial tight junctional complexes.
The absence of GLUT-1 immunoreactivity in the ECs in non-BBB-type blood microvessels lined with fenestrated (hypophysis and subfornical organ) or continuous (skeletal muscle) endothelium is not surprising in view of earlier findings indicating the association of GLUT-1 with the barrier function and its absence in the vasculature of circumventricular organs (Gerhart et al. 1989; Harik et al. 1990). More complex, however, is the problem of the localization of GLUT-1 in the choroid plexus. Our observations indicating negligible immunoreactivity of the vascular fenestrated endothelium and distinct labeling of the undulated basolateral membrane of the epithelial cells, with absence of immunolabeling of apical plasmalemma and intercellular junctions, are compatible with those obtained in mice, rats, and rabbits by other authors (Kalaria et al. 1988; Harik et al. 1990; Farrell et al. 1992; Cornford et al. 1995; Bolz et al. 1996). Nevertheless, they differ from recent observations of Cornford et al. (1998) indicating an abundance of GLUT-1 immunoreactivity in the vascular fenestrated endothelium and in the basolateral membranes of the epithelial cells of human (biopsy specimens) and nonhuman primate choroid plexi. These observations suggest the existence of species differences as well as differences between primates, including humans, and laboratory animals.
Our morphometric analysis of the density of immunolabeling for GLUT-1 in microvessels supplying cerebral cortex, hippocampus, and cerebellum revealed similarity among these brain regions. Despite the roughly similar mean values shown in Table 2, in all of these regions some differences and nonuniformity of the labeling density in particular vascular profiles and segments were observed. The uneven distribution of immunosignals suggests the existence of local differences in glucose transport from blood to brain parenchyma. Similar results using confocal laser scanning microscopy, suggesting uneven distribution of glucose-transporting molecules in the mouse hippocampus, were reported by Shimada et al. (1994).
More pronounced fluctuations in the labeling density observed in the blood microvessels of the cerebellum were probably associated with their anatomic location in different histological layers of the cerebellar gray matter. When radiolabeled cytochalasin B, which binds specifically to glucose transporter molecules, was used as a ligand, the existence of variations in the density of labeling of cerebellar microvessels was also reported (Dick and Harik 1986; Tucker and Cunningham 1988). Dick and Harik (1986) observed that the labeling density with cytochalasin B was slightly lower in the rat cerebellar than in cerebral microvessels, whereas in the pig, this lowering was more pronounced (almost to 50%). According to these authors, the differences in density of GLUT-1 molecules in the microvascular wall are related to different utilization of glucose in a given brain region: higher density of transporter molecules means higher glucose utilization by neurons.
Our observations of statistically significant lower immunodetection of GLUT-1 in the olfactory bulb microvasculature suggest lower utilization of glucose in this region. It is also possible that a high concentration of GLUT-1 in the vascular endothelium is associated with its barrier function. This notion is supported by our actual and previous observations indicating that the microvessels located in the internal granular layer show both low immunolabeling for GLUT-1 and leakage of the endogenous albumin. The leakage suggests that the BBB in the olfactory bulb is inefficient or, at least, is not as tight as it is in the cerebral cortex or other brain regions (Ueno et al. 1996).
The problem of the relation between the density of GLUT-1 molecules in the brain vascular endothelium and its barrier function is of great importance for better understanding of BBB physiology and pathology, and requires experimental elucidation.
Footnotes
Acknowledgments
Supported by funds from the New York State Office of Mental Retardation and Developmental Disabilities.
We wish to express our appreciation to Ms M. Stoddard Marlow for meticulous editorial revisions and Ms J. Kay for secretarial assistance.