
Abstract
We localized REG protein in Paneth cells and nonmature columnar cells of the human small intestinal crypts and speculated that this protein was associated with growth and/or differentiation. The aim of this study was to determine whether REG protein is present in two human colon cancer cell lines that exhibit enterocytic differentiation after confluence and to investigate changes in the level of its expression during growth and differentiation. Results were compared to those obtained on cells that remain undifferentiated. Western immunoblotting and immunofluorescence demonstrated the presence of REG protein in the three cell lines. With the antisera against human REG protein, the staining was diffusely spread throughout the cytoplasm at Day 2, and after Days 3-4 it appeared to have migrated to cell boundaries. After confluence, we observed only a punctate staining array along cell boundaries, which disappeared at Day 15. REG mRNA expression was demonstrated by RTPCR and REG mRNA hybridization until Day 13, but not after, in the three cell types. REG protein may be involved in cellular junctions. Its presence appears to be associated with the cell growth period and the protein must be downregulated when growth is achieved and differentiation is induced.
I
In the small intestine, we demonstrated the presence of REG protein (Senegas-Balas et al. 1991). We localized this protein in Paneth cells and nonmature columnar cells of the human intestinal crypts but never in functional villous epithelium. Therefore, we speculated that this topographical distribution might be associated with growth and also with differentiation.
In an attempt to verify this hypothesis, we use some human colon carcinoma cell lines (Caco-2 and HT-29 Glc-/+ cell lines) in which the occurrence of enterocytic differentiation has been reported (Pinto et al. 1983; Lesuffleur et al. 1990, 1991; Kerneis et al. 1994; Bernet-Camard et al. 1996ba,b; Bernard-Perrone et al. 1998) because no human small intestinal cell lines exist. This differentiation developed as a function of day of culture after confluence, and is characterized by the appearance of tight junctions, of well-organized brush borders, and of intestinal hydrolases in these cancer cell lines, as in the small intestine. Moreover, the REG gene was also found to be expressed ectopically in some human colon and rectal tumors but not in healthy tissues (Watanabe et al. 1990). The present study was therefore undertaken to determine whether REG protein is present in human colon cancer cell lines and to investigate changes in the level of its expression throughout the cell culture, during the exponential phase of cell growth, the beginning of differentiation, and in the differentiated cells. All results were compared to those obtained on HT-29 Glc+cells, which always remain mainly undifferentiated.
Materials and Methods
Cell Lines and Culture Conditions
Caco-2 cells and HT-29 cells derive from human colorectal adenocarcinoma (Dr. J. Fogh; Sloan-Kettering Memorial Center, Rye, NY) (Fogh and Trempe 1975). Caco-2 cells spontaneously differentiate after confluence, as is well known (Pinto et al. 1983). We used two populations of HT-29 cells. Parental HT-29 cells (HT-29 Glc+cells) were grown in a medium containing glucose and remained undifferentiated throughout cell culture (Pinto et al. 1983; Lesuffleur et al. 1990). Permanently differentiated HT-29 cells (HT-29 Glc-/+cells) were obtained from A. Zweibaum. HT-29 Glc-/+cells were a 100% enterocytic subpopulation (Kerneis et al. 1994) obtained by selection through glucose deprivation that maintains its differentiation characteristics when switched back to standard glucose-containing medium (Lesuffleur et al. 1991). HT-29 Glc-/+cells also differentiated after confluence and exhibited well-organized brush borders (Lesuffleur et al. 1991; Bernet-Camard et al. 1996b) and tight junctions (Bernard-Perrone et al. 1998). These cells expressed aminopeptidase N and DPPIV, whose levels strongly increased after confluence (Lesuffleur et al. 1991; Bernet-Camard et al. 1996ba,b; Bernard-Perrone et al. 1998).
Cells were grown in Dulbecco's modified Eagle's minimal essential medium (DMEM) (Sigma Chemical; St Louis, MO) supplemented with 10% (HT-29) or 20% (Caco-2) inactivated (56C, 30 min) fetal calf serum (FCS) (Gibco, Grand Island, NY) and 1% nonessential amino acids (Caco-2). Cells were seeded at 8 × 104 cells/ml in 35-mm Petri dishes or on glass coverslips, which were placed in six-well tissue culture plates (Corning Glass Works; Corning, NY). All the experiments and the maintenance of cells were carried out at 37C in 10% CO2-90% air. The culture medium was changed daily. In our culture conditions (Bernard-Perrone et al. 1998), growth curves showed that cell confluence was reached between Days 5 and 6 for HT-29 cells and between Days 6 and 7 for Caco-2 cells. A stationary phase was noted at Days 11, 12, and 13, respectively, for HT-29 Glc+ cells, HT-29 Glc-/+ cells, and Caco-2 cells.
Preparation of Cell Lysates
After washing the cells in PBS, cell lysates (Days 8 and 15) were recovered in PBS containing 2 mM EDTA. The cells were pelleted by centrifugation at 1000 rpm for 15 min and then resuspended in a lysis buffer [50 mM Tris-HCl (pH 7.5), 1% Triton X-100, 2 mM ethylene glycol bis (β-aminoethyl ether)-N, N, N', N'-tetraacetic acid, 10 mM EDTA, 100 mM NaF, 1 mM Na4P2O7, 2 mM Na3VO4, 1 mM PMSF, 1 μg/ml aprotinin, 1 μg/ml pepstatin A, and 1 μg/ml leupeptin]. The supernatant fractions obtained by centrifugation of the lysates at 14,000 × g for 10 min were harvested and stored at −80C for Western immunoblot analysis.
Preparation of Antibodies
Antisera against REG protein and its proteolysis product were prepared by injecting into rabbits the two protein bands (19 kD and 14 kD) obtained by SDS-gel electrophore-sis of the fractions containing these proteins, co-purified by DEAE-Trisacryl chromatography. The antibodies tested by immunoelectrophoresis against human pancreatic juice gave one single precipitating line (Amouric et al. 1987). By Western immunoblotting, they reacted exclusively with the REG protein family in nonactivated pancreatic juice and with the proteolytic product of REG protein in slightly activated pancreatic juice (Senegas-Balas et al. 1991).
To avoid false-positive reactions resulting from the rabbit blood groups, we checked that antibodies against human erythrocytes from group A and O were absent from all rabbit sera (Oriol and Dalix 1977). Because it was reported that the IgM component of rabbit serum reacts with the plasma membrane of human colon carcinoma cell lines (Hager and Tompkins 1976), the IgG of the antibodies and of the preim-mune rabbit serum was purified by chromatography on protein A-Sepharose CL-4B (Pharmacia; Uppsala, Sweden). The absence of IgM in these purified antibodies was checked by Ouchterlony double diffusion performed with goat IgG fraction to rabbit IgM (x -chain) (Organon Teknika; Durham, NC). These purified IgGs were used for immunofluorescence techniques.
Immunofluorescence Staining
Indirect immunofluorescence was performed on cell monolayers grown on glass coverslips. Cells were fixed in paraformaldehyde (3.5%) (Days 2-20) for 2 min for HT-29 cells and for 10 min for Caco-2 cells, and then were quenched in 50 mM ammonium chloride. These cells were permeabilized in various concentrations of saponin (0.1%, 0.05%, 0.075%) or Triton (0.05%, 0.1%) for 5-15 min. We obtained good staining and the best cell preservation for saponin (0.1%, 10 min). For Caco-2 at Day 15, we also used pepsin solution (0.1% in HCl 0.001 N, pH 2, 10 min) (Sigma) for retrieval of antigen masked by formalin fixation. Cells were then incubated sequentially with 3% nonimmune goat serum, rabbit IgG against human REG protein, or against its nonglyco-sylated proteolysis product (IgG against 14-kD protein) (1:20-1:100 in PBS/3% goat serum/0.2% gelatin; 12 hr, 4C) and an FITC-conjugated goat anti-rabbit IgG (1:100, 1 hr) (Dako; Carpinteria, CA).
Caco-2 cells (days 3 and 10) were also observed with a confocal laser microscope LSM 410 (Carl Zeiss; Jena, Germany) after using IgG against 14-kD protein. An argon ion laser adjusted to 488 nm was used for analysis of fluorescence. Optical sectioning was used to collect four en face images (8 × m) at Day 3. At Day 10, nuclei were labeled by propidium iodide and a series of en face views (1 μm) and lateral sections (x-y axis) (8 μm) were performed.
Several controls were performed: (a) PBS or whole preimmune rabbit serum replacing primary antisera; (b) PBS replacing FITC-conjugated goat anti-rabbit IgG; and (c) immunoadsorption tests by incubating 0.2 mg/ml of the 14-kD proteolytic product of REG protein (4C, 24 hr) with the two primary antisera.
Immunocytochemical reactions were also carried out on living Caco-2, HT-29 Glc -/+, and HT-29 Glc+ cells at Days 3 and 7. They were incubated for 1 hr with IgG against 14-kD protein (1:100) in culture medium supplemented with 10% FCS (CM). After three rinses in culture medium without FCS, cells were incubated for 30 min with an FITC-conjugated goat anti-rabbit IgG (1:100) in CM. After rinses, cells were treated in methanol/acetic acid (95/5, v/v). Fixed cells were treated in the same fashion.
Peroxidase-Anti-peroxidase Staining
Multiple human duodenum and colon biopsy specimens were taken from five consenting subjects. Fragments of duodenum, jejunum, ileum, and colon from five Wistar rats and five Swiss albino mice were removed. These specimens were fixed in Baker's formalin at 4C, embedded in cytoparaffin, and treated with PAP techniques using IgG against 14-kD protein.
Western Immunoblotting
SDS-polyacrylamide gel (10%) electrophoresis (SDS-PAGE) was performed (Laemmli 1970) with cell lysates of Caco-2 and HT-29 GLc-/+ (20 μg of protein) and HT-29 GLc+ (40 μg of protein) at Days 8 and 15 and with human nonactivated pancreatic juice (0.1 μg of protein). After transferring the proteins to PVDF membranes (250 mA; Millipore, Bedford, MA), the membranes were incubated overnight in PBS containing 5% (w/v) dry skimmed milk. Immobilized proteins were characterized by using the antiserum against 14-kD protein (1:5000) in PBS containing 10% (w/v) dry skimmed milk or whole preimmune rabbit serum under the same conditions. The blots were incubated with horseradish peroxidase-labeled anti-rabbit antibody (1 hr, 1:5000; Bio-Rad, Hercules, CA). Then the membranes were washed three times with PBS, 0.1% (v/v) Tween-20. The immunoreactive bands were detected by the Enhanced Chemi-Luminescence system (Amersham; Poole, UK). We used the standard prestained proteins (Bio-Rad prestained SDS-PAGE Standards, low range).
Reverse Transcription Polymerase Chain Reaction (RT-PCR)
After being washed twice in PBS, the Caco-2, HT-29 Glc+, and HT-29 Glc-/+ cells (Day 8) were scraped and dispersed in 4 M guanidium isothiocyanate (3 dishes/cell line). Total RNAs were purified (Chirgwin et al. 1979). Purification of mRNA from total colon cancer cell RNA was performed with an mRNA separator kit (Clontech Laboratories; Palo Alto, CA). These mRNAs (0.5 mg) and total human pancreatic RNA (1 mg) were reverse-transcribed into cDNA using Oligo (dT)15 (Boehringer; Mannheim, Germany) as primer and with or without SuperScript TM II reverse transcriptase (Life Technologies; Gaithersburg, MD) according to the manufacturer's recommendations. The mRNAs were used for REG mRNA PCR analysis only if human actin amplification was observed and none without reverse transcriptase. PCR amplifications were accomplished (Taq DNA polymerase; Life Technologies) with an MJ Research, PT-100 DNA thermal cycler. REG cDNA was produced by RT-PCR from 0.5 μg for the cells and from 1 μg of human pancreatic RNA by using the sense primer 5′-CAGCTCAGCATGGCTCAGACC-3′ (nucleotides 367-387) (exon 2) and antisense primer 5′-GTCATGGAGGCCAATCCAGAC-3′ (nucleotides 1609-1629) (exon 4). Samples were initially denatured at 95C for 1 min. Cycling parameters (35 cycles) were as follows: denaturation at 95C for 1 min, annealing at 68C for 1 min, and extension at 72C for 1 min. A final extension at 72C for 5 min was performed. Amplified products were analyzed by electrophoresis. Cross-tissue contamination was checked with primers used alone in liquid controls.
Quantitative Analysis of REG mRNA Expression by Dot-Blot Hybridization
We used total RNA of Caco-2, HT-29 Glc+, and HT-29 Glc-/+ cells of human jejunum, pancreas (positive control), and spleen (negative control; Watanabe et al. 1990) (obtained from organ donors). GAPDH cDNA (a gift from Dr. Hirsh) was used to test mRNA stability during the days of the culture before the REG mRNA hybridizations. The human REG cDNA corresponds to a 621-BP cDNA in pBS vector (Terazono et al. 1988). cDNAs were labeled by nicktranslation with [32P]-dCTP (Maniatis et al. 1975). Dot-blot assays were performed (Favarolo et al. 1980; White and Bancroft 1982). Sequential dilutions of total RNA (5-0.156 μg for the cells and 0.5-0.0156 μg for tissues) were loaded onto nitrocellulose filters and hybridized with cDNA probes (three assays). REG mRNA transcript levels were quantified by densitometry after hybridization with the REG probe. mRNA concentrations were expressed in arbitrary units per μg of total RNA (mean ± SEM). Statistical analysis was made with the analysis of variance (ANOVA) using the Stat View 512+ software (Brain Power; Calabasas, CA).
Results
Cytoimmunofluorescence Demonstration of REG Protein in Cell Lines
The presence of REG protein in the three cancer cell lines was demonstrated because, with the two primary antisera, intense labeling was seen until Day 11. The staining was diffusely spread throughout the cytoplasm at Day 2 (Figures 1A, 1G, 1J, and 1M). At Days 3 to 4, the labeling was always diffusely spread in some cells, whereas in other cells it already seemed to migrate to cellular junctions (Figure 1B. After confluence (Days 7 and 11), we observed a punctate staining array along cell boundaries (Figures 1C, 1D, 1H, 1K, 1O, and 1Q). The labeling disappeared at Day 15 (Figure 1E even after pepsin treatment (not shown). All the control tests were negative (Figures 1F, 1I, 1L, 1N, 1P, and 1R).
No staining was seen in living cells at Days 3 and 7 (not shown). On the contrary, fixed cells treated for 1 hr with the IgG against REG protein or against its proteolytic product exhibited similar but slight labeling compared to cells treated for 12 hr (not shown).
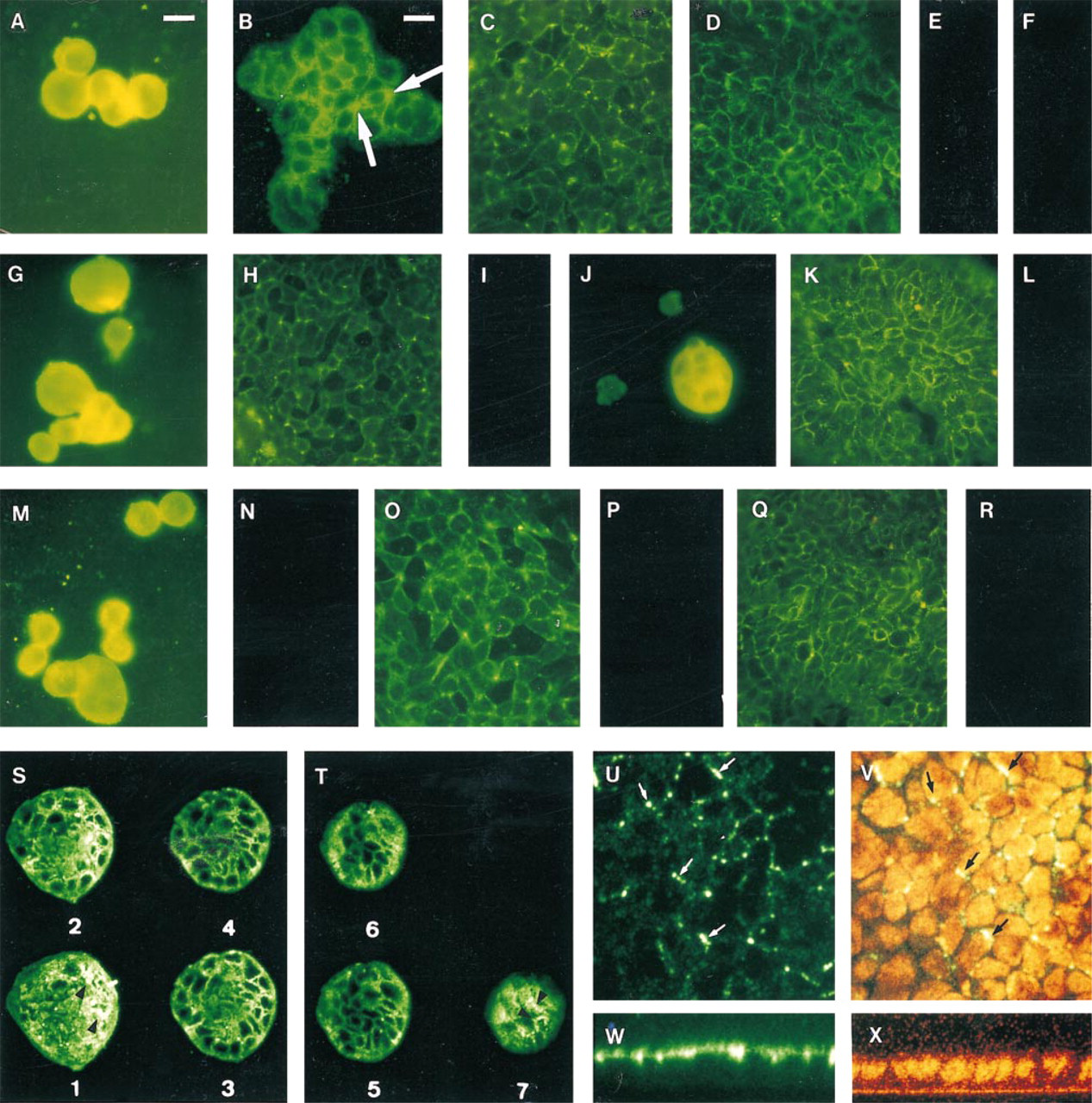
Immunocytofluorescence on colon cancer cell lines. (
This absence of staining on living cells demonstrated that the labeling seen on fixed cells is clearly localized inside these cells before confluence. After confluence, the staining observed on fixed cells could not correspond to labeling of the external part of the apical membrane because nonpermeabilized living cells were not labeled. In these living cells, we could not exclude that the antibodies might not have access to basolateral intercellular spaces where the REG protein could be present.
For Caco-2 cells at Day 3, we could see diffuse labeling from the basal part of the cells to the apical one (Figures 1S and 1T) with confocal microscopy. We observed intense staining in the first section of some cells, which could correspond to endoplasmic reticulum (Figures 1S1 and 1S2) as we have already seen in human intestine (Senegas-Balas et al. 1991). However, faint staining could also be seen near the boundaries of some cells (Figure 1T7); these cells are the tallest cells of the islet (upper sections). These results confirmed those observed by immunofluorescence.
At Day 10, lateral views clearly show that the staining is restricted to a small zone near or on the lateral plasma membrane (Figure 1U, located on a level with the upper part of the nucleus (Figure 1W; this labeling could be seen in seven successive en face sections of 1 μm (not shown).
Peroxidase-Anti-peroxidase Staining
Paneth and crypt cells of duodenal sections, but not human colon or rat and mouse small intestine and colon, were labeled (not shown). Therefore, the two antisera reacted specifically with the human small intestine, as with human pancreas (Senegas-Balas et al. 1991).
Western Immunoblotting
A 19-kD band was observed with pancreatic juice and with the three cell line cell lysates at Day 8, which is consistent with the presence of REG protein (Figure 2). On the contrary, no band could be observed at Day 15, confirming immunofluorescence studies.
REG mRNA Expression in Cell Lines at Day 8
PCR analysis of cDNA samples generated from the three cell lines indicated the presence of a specific signal with the expected length (321 BP) for REG mRNA (Figure 3, Lane c, Caco-2 cells; Lane e, HT-29 Glc-/+ cells; Lane g, HT-29 Glc+ cells) as in human pancreas (Figure 3, Lane i). Cross-tissue contamination could be excluded because no PCR product was detected when primers were used alone in liquid controls (Figure 3, Lanes b, d, f, and h). REG mRNA expression was demonstrated in the three colon cancer cell lines.

Representative Western immunoblots of REG protein isolated from the Caco-2 (Lane 4, Day 8; Lane 7, Day 15), from HT-29 Glc-/+ (Lane 5, Day 8; Lane 8, Day 15), and HT-29 Glc+ cell lysates (Lane 6, Day 8; Lane 9, Day 15) and human pancreatic juice (Lane 10), using rabbit anti-human REG protein proteolytic product antibody. Western immunoblots were performed with the same lysates of Caco-2 cells (Lane 1), HT-29 Glc-/+ cells (Lane 2), and HT-29 Glc+ cells (Lane 3) and with pancreatic juice (Lane 11) by using whole rabbit serum. A 19-kD band was observed with the three cell line lysates at Day 8 and with pancreaticjuice.
REG mRNA Levels
Dot-blot assays enabled us to show that REG mRNA was present in human jejunum and in the three cell lines from Day 3 to Day 13 and was absent at Day 21 (Figure 4). As expected, it was present in human pancreatic tissue, and no hybridization for human spleen was detected (Figure 4). Similar amounts of mRNA were detected at Day 3 and Day 13 of culture in the three colon cancer cell lines (Figure 5). Similar amounts of mRNA coding for REG protein were found in HT-29 Glc-/+ and HT-29 Glc+. The levels of Caco-2 REG mRNA were significantly higher than those of HT-29 cell lines (+25% and +29% at Days 3 and 13 compared to HT-29 Glc-/+ levels; + 35% and + 29% at Days 3 and 13 compared to HT-29 Glc+ levels; p<0.05).
Discussion
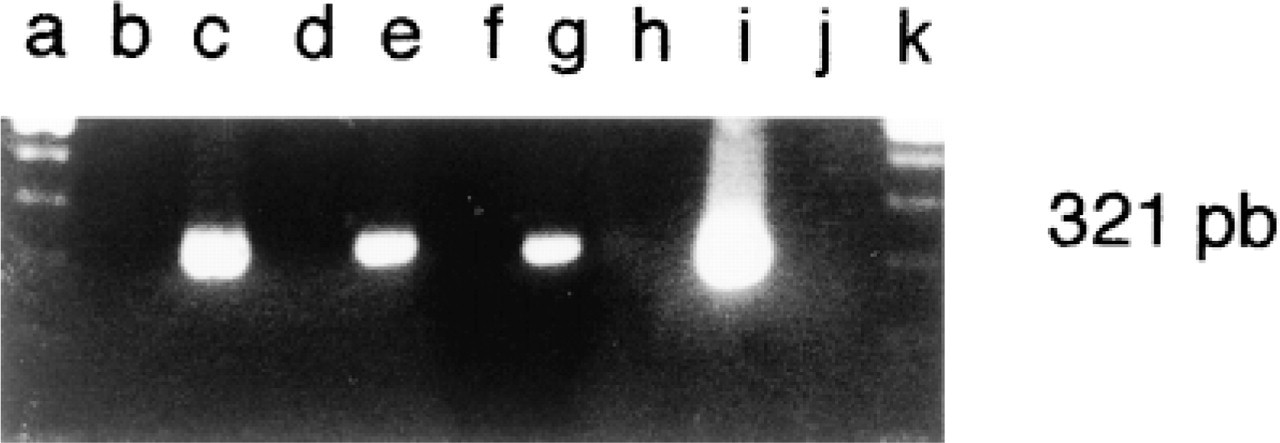
Expression of REG mRNA detected by PCR in the three cell lines at Day 8 and in the pancreas (Lane c, Caco-2 cells; Lane e, HT-29 Glc-/+ cells; Lane g, HT-29 Glc+ cells; Lane i, pancreas). PCR amplifications were also performed without cDNA sample (Lanes b, d, f, h) and the molecular size marker is found on Lanes a and j.
We confirmed our previous cytoimmunological results in human small intestine (Senegas-Balas et al. 1991) by the demonstration of REG mRNA expression in human jejunum. We also clearly demonstrated REG mRNA and REG protein in Caco-2, HT-29 Glc-/+, and HT-29 Glc+ colon cancer cell lines by Western immunoblotting, immunofluorescence, RT-PCR, and hybridization of REG cDNA with total RNA of these cells. Although REG 1a and REG 1b proteins were demonstrated in pancreas (Moriizumi et al. 1994; Miyashita et al. 1995), we could only postulate the presence of REG protein in the three cancer cell lines and in human jejunum.
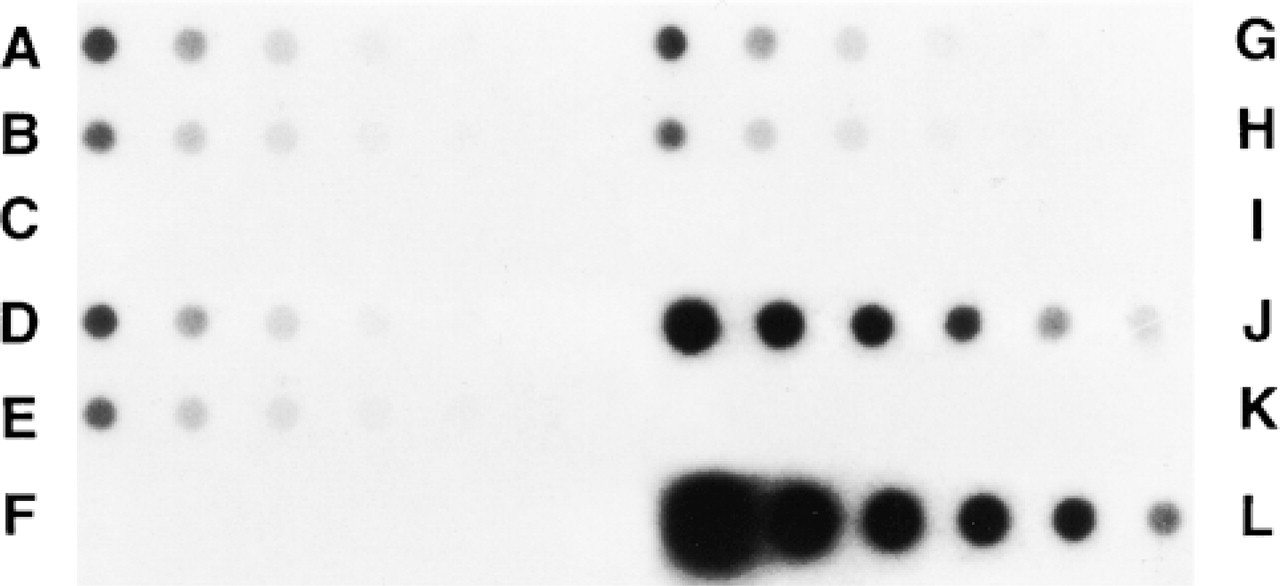
Dot-blot autoradiographs of REG mRNA. (
The presence of REG protein in colon cancer cell lines was in accordance with the hybridization of REG cDNA with RNA from some human colon and rectal tumors (Watanabe et al. 1990; Kim et al. 1997). This type of hybridization has never been shown in healthy human colon or rectum (Watanabe et al. 1990; Moriizumi et al. 1994), and this result is in agreement with the absence of labeling that we demonstrated in human healthy colon sections. Therefore, REG protein and REG mRNA expressed to be present only in tumorous colon. Some authors have proposed that REG protein is involved in the dedifferentiated states of the gastrointestinal tract (Watanabe et al. 1990; Kim et al. 1997).
The staining localization varied during the culture, with similar patterns in the three colon cancer cell lines, differentiated or not. Labeling was observed until Day 11 and had disappeared at Day 15. REG protein was detected at Day 8 but not at Day 15 by Western blot. The REG mRNA expression was observed until Day 13. Days 11-13 correspond to the first days of the stationary phase of growth. Therefore, expression of REG protein and REG mRNA was observed during the growth period of colon cancer cell lines and also during the first days of the stationary phase. This REG protein expression during the growth period agrees with the results observed in regenerating pancreatic islets, in pancreatic exocrine tissue, in stomach, and in cornea, where REG protein appears to be increased during the growth period (Terazono et al. 1988, 1990; Rouquier et al. 1991; Francis et al. 1992; Unno et al. 1993; Otonkoski et al. 1994; Watanabe et al. 1994; Rafaeloff et al. 1995; Fredj-Reygrobellet et al. 1996; Asahara et al. 1996).
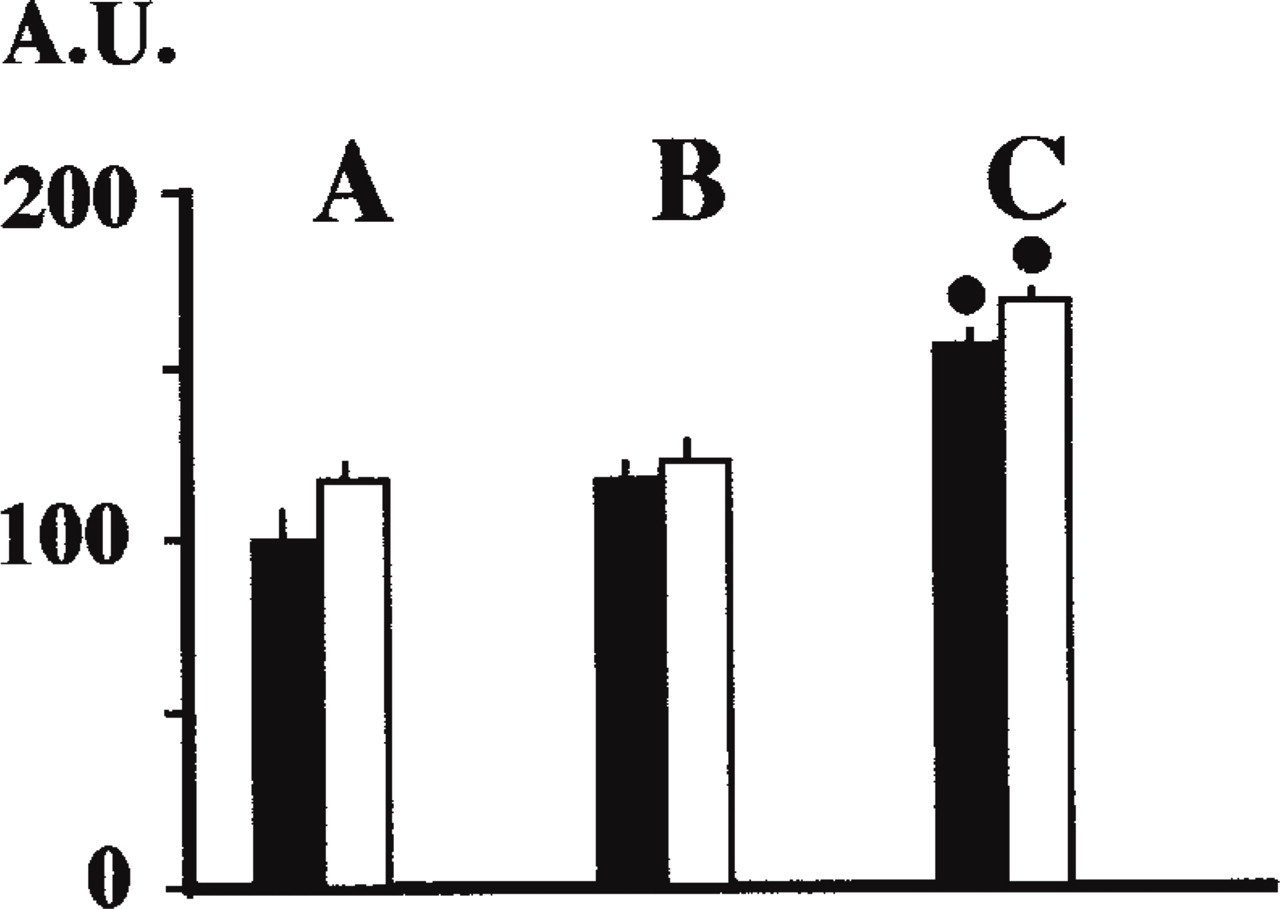
REG mRNA levels in HT-29 Glc+ cells (
In colon cancer cell lines that differentiate (Caco-2 and HT-29 Glc-/+), the beginning of differentiating processes was observed after Day 5 (Bernard-Perrone et al. 1998). Finally, REG expression disappeared when growth and the well-known burst of differentiation is achieved. We have demonstrated the same phenomenon in healthy human small intestine, in which only the cells localized in the glands of Lieberkühn expressed REG protein but not the mature and functional villous cells (Senegas-Balas et al. 1991). It is well known that the crypt base columnar cells are only proliferative cells and that the mid-crypt columnar cells proliferate and also differentiate, whereas the crypt top columnar cells never divide but continue to differentiate (Cheng and Leblond 1974). These crypt top cells were stained by our two antisera against REG protein (Senegas-Balas et al. 1991). Therefore, REG protein may be associated with growth, and our results also suggest a role early in the differentiating process of intestinal epithelium, as has been proposed for pancreas (Kim et al. 1997).
The localization of the staining varied during the culture of the three colon cancer cell lines. Labeling was observed diffusely in the cytoplasm during the first days of culture. After confluence, it was restricted to cell junction zones and presented a punctate staining array along cell boundaries. By confocal microscopy, this staining was confirmed to be near the cell boundaries and in a very specific zone on a level with the upper part of nucleus. These results suggest that REG protein is implicated in cell junctions. In this respect, it is interesting to note that REG protein was integrated into Group VII of the calcium-dependent lectin (C-type animal lectins) classification sharing a single carbohydrate recognition domain linked to an amino acid sequence corresponding to a signal peptide (Patty 1988; Drickamer 1993; Lassere et al. 1994). C-lectins participated in many cell-surface recognition events that are known to be associated with cell growth and/or differentiation (Patty 1988; Petersen 1988; Tachibana et al. 1988; Lassere et al. 1994). In addition, HIP/PAP protein, which is also a C-lectin of Group VII, induced the adhesion of rat hepatocytes and bound strongly with extracellar matrix proteins such as laminin-1 and fibronectin (Christa et al. 1996). Moreover, REG protein is closely related to invertebrate lectins (Lassere et al. 1992; Drickamer 1993), which are important in the development of these organisms (Lassere et al. 1992; Drickamer 1993).
We clearly demonstrated the presence of REG protein in the three colon cancer cell lines used, where this protein seemed to be involved in cell junctions. REG expression was concomitant with the cell growth period. We can also hypothesize that REG protein will be downregulated after the induction of differentiation.
Footnotes
Acknowledgements
We are indebted to Dr A. Servin (Laboratoire de Microbiologie; Chatenay-Malabry, France) for the kind gift of the cell lines Caco-2 and HT-29, and to Dr Hirsh (Centre d'Immunologie de Luminy; Marseille, France) for GAPDH cDNA.