
Abstract
To visualize cell surface V1a vasopressin receptors in rat hepatocytes in the absence of receptor-mediated endocytosis, we used a high-affinity fluorescent linear antagonist, Rhm8 -PVA. Epifluorescence microscopy (3CCD camera) and fluorescence spectroscopy were used. Rhm8 -PVA alone did not stimulate Ca2+ signals and competitively blocked Ca2+ signals (Kinact of 3.0 nM) evoked by arginine vasopressin (vasopressin). When rat hepatocytes were incubated with 10 nM of Rhm8 -PVA for 30 min at 4C, the fluorescent antagonist bound to the surface of cells, presumably the plasma membrane. The V1a receptor specificity of Rhm8 -PVA binding was confirmed by its displacement by the nonfluorescent antagonist V4253 and by the natural hormone vasopressin at 4C. Prior vasopressin-mediated endocytosis of V1a receptors at 37C abolished binding of the labeled antagonist, whereas in non-preincubated cells, Rhm8 -PVA labeled the cell surface of rat hepatocytes. When cells labeled with Rhm8 -PVA at 4C were warmed to 37C to initiate receptor-mediated internalization of the fluorescent complex, Rhm8 -PVA remained at the cell surface. Incubation temperature at 4C or 37C had little effect on binding of Rhm8 -PVA. We conclude that Rhm8 -PVA is unable to evoke receptor-mediated endocytosis and can readily be used to visualize cell surface receptors in living cells.
VASOPRESSIN V1a receptors are found in a variety of tissues, including liver, blood vessels, kidney, choroid plexus, and central nervous system (for reviews see Jard and Thibonnier 1993; Howl and Wheatley 1995). The rat hepatocyte, a cell relatively rich in V1a receptors, expresses 200,000 binding sites (Cantau et al. 1980). Vasopressin acts via V1a receptors to elicit [Ca2+]i oscillations and intracellular Ca2+ waves in single rat hepatocytes (Woods et al. 1986; Rooney et al. 1990). The hormone also evokes Ca2+ waves and synchronized oscillations that travel from cell to cell in hepatocyte doublets (Nathanson and Burgstahler 1992a,b; see Boyer 1997), in multicellular systems (Combettes et al. 1994; Tordjmann et al. 1997a, 1998), and in the intact perfused liver (Nathanson et al. 1995; Robb-Gaspers and Thomas 1995). The V1a receptor is associated with Gα11/Gαq proteins (Morel et al. 1992; Exton 1996). Like most G-protein-coupled receptors already studied (Lefkowitz 1993), the V1a receptor undergoes desensitization when stimulated by its agonist. Such a desensitization is due to PKC-mediated uncoupling of the receptor from its G-proteins within 1 min and to internalization of cell surface V1a receptors within 10 min. Internalization of V1a receptors is an agonist-mediated endocytotic process because it is blocked by V1a vasopressin antagonists (Lutz et al. 1992). Both rapid desensitization and delayed endocytosis have been demonstrated for the V1a receptor in native tissues (Breant et al. 1981; Fishman et al. 1985; Caramelo et al. 1991), cell lines (Cantau et al. 1988; Grier et al. 1989; Lutz et al. 1990b,1993), and in cells expressing the cloned receptor gene (Nathanson et al. 1994b; Ancellin et al. 1997).
V1a vasopressin receptors have been visualized in several tissues by different methods (for reviews see Carmichael et al. 1993; Jard and Thibonnier 1993; Howl and Wheatley 1995). In most studies, localization was investigated by autoradiography of radiolabeled vasopressin ligands (Barberis et al. 1995; for references see Serradeil Le Gal 1996) or by immunolocalization with specific antibodies raised against the receptor (Nathanson et al. 1995). These methods have significant disadvantages. Autoradiography can be used only with whole organs and gives poor spatial definition, and specific antibodies are not commercially available for immunolocalizing the V1a vasopressin receptor in rat hepatocytes. Fluorescent V1a vasopressin antagonists and agonists have been synthesized (Jans et al. 1990; Lutz et al. 1990a; Guillon et al. 1992) to visualize, respectively, the cell surface receptors or the development of receptor-mediated endocytosis of V1a vasopressin receptors in living cells (Lutz et al. 1990b,1993). The objective was to produce high-affinity ligands that would remain fluorescent when bound to the receptor and would dissociate slowly when the free molecule was washed away (McGrath et al. 1996). These fluorescent ligands are relatively easy to handle and produce easily interpreted cell images. However, synthesizing them is laborious and they are therefore no longer produced and commercially available (Carmichael et al. 1993).
To visualize cell surface V1a vasopressin receptors in rat hepatocytes in the absence of receptor-mediated endocytosis (Lutz et al. 1992), we used a high-affinity fluorescent linear antagonist, Rhm8 -PVA previously described (Durroux et al. in press), which could be used as an irreversible ligand (McGrath et al. 1996). We used an epifluorescence microscope equipped with a high-resolution and sensitive 3CCD camera to detect the binding of Rhm8 -PVA to V1a receptors and to assess the ability of the antagonist to inhibit receptor-mediated endocytosis. The experiments were performed in single cells and in multicellular systems of rat hepatocytes. The potency of the fluorescent vasopressin antagonist in blocking vasopressin-mediated Ca2+ signals was examined by fluorescence spectroscopy.
Materials and Methods
Preparation of Rat Hepatocytes
Single cells (70%) and multicellular systems (30%), called multiplets (doublets and triplets), were prepared from fed female Wistar rats by limited collagenase (Boehringer Mannheim; Meylan, France) digestion of rat liver, as previously described (Combettes et al. 1994). After isolation, rat hepatocytes were maintained (2.106 cells/ml) at 22C in modified Eagle's medium (MEM) (Gibco BRL Life Technologies; Paisley, UK) supplemented with 1.5% gelatin (Labosi; Elancourt, France), and continuously gassed with O2/CO2 (19/1) at pH 7.4. Cell viability, assessed by trypan blue exclusion, remained greater than 96% for 4-5 hr.
Synthesis of the Fluorescent Vasopressin Antagonist
A fluorescent vasopressin antagonist, 4-HO-Ph(CH2)2CO-DTyr(Me)-Phe-Gln-Asn-Arg-Pro-Lys(5-carboxytetramethylrhodamyl)-NH2 (Rhm8 -PVA), which binds to vasopressin V1a receptors, has been recently synthesized. The synthetic strategy involved Rhm8 -PVA by coupling 5-carboxy tetramethyl rhodamine to a Lys8 linear vasopressin antagonist (Durroux et al. in press).
Cell Loading with Quin2 for Measuring [Ca2+]i in Suspensions of Rat Hepatocytes
Suspensions of rat hepatocytes were loaded with quin2 by incubation with the permeant acetoxymethyl ester form of the dye (Molecular Probes; Leiden, The Netherlands) and [Ca2+]i was measured as previously described (Tordjmann et al. 1997b). Competition inhibition assays were performed to establish the ability of Rhm8 -PVA to activate hepatic V1a receptors. Except for the experiment illustrated in Figure 1, rat hepatocytes were preincubated with 20 nM Rhm8 -PVA and then 1 min later with one concentration (0.1-10 nM) of arginine vasopressin (Sigma Chemical; St Louis, MO). In another series of experiments, cells were preincubated with one concentration (0.1-100 nM) of Rhm8 -PVA and then 1 min later with 0.4 nM vasopressin.
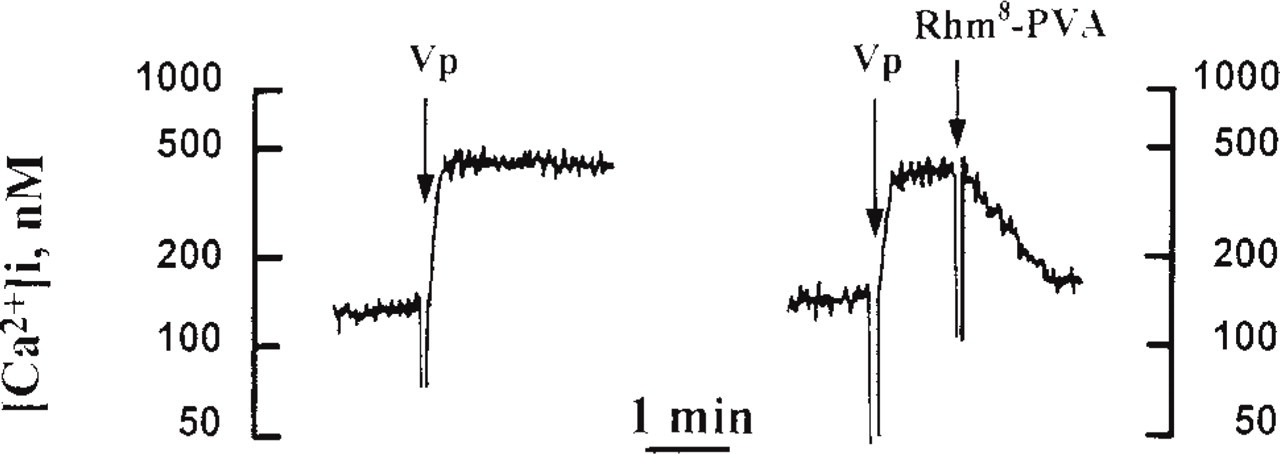
Rhm8 -PVA inhibits vasopressin-mediated [Ca2+]i rise in suspensions of rat hepatocyte (typical traces). Left panel illustrates a typical trace of the effects of 1 nM vasopressin (near the kD of V1a receptors) on [Ca2+]i (nM) measured in quin2-loaded rat hepatocytes. The hormone increased [Ca2+]i within a fraction of a minute from 120 nM to a sustained plateau close to 500 nM in suspensions of rat hepatocytes. Right panel shows that addition of 20 nM Rhm8 -PVA 1 min after 1 nM vasopressin reversed the [Ca2+]i rise induced by the hormone within 1 min.
Visualization of Cell Surface V1a Receptors with Rhm8 -PVA in Rat Hepatocytes
Single cells and multiplets were suspended in Williams' medium E (5 × 105 cells/2 ml), supplemented with 10% fetal calf serum, glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin (Gibco BRL; Gaithersburg, MD) and plated on collagen-coated glass coverslips for 2 hr at 37C under an atmosphere containing 5% CO2. Cells were rinsed twice with MEM and preincubated with vasopressin or with the non-fluorescent vasopressin antagonist V4253 ([β-Mercapto- β-β-cyclopenta-methylenepropionyl1, O-Et-Tyr2, Val4, Arg8]-vasopressin) (Sigma) at concentrations, temperatures, and time periods of incubation indicated in the figure legends. At the end of these preincubation periods, cells were rinsed twice with MEM, then further incubated for 30 min at the same temperature with 10 nM Rhm8 -PVA in MEM supplemented with 1 mg/ml bovine serum albumin (Miles Laboratories; Kankakee, IL). Cells were washed twice at 4C with MEM, fixed by incubation for 15 min in 4% formaldehyde (FA), and mounted in buffered glycerin (Sanofi Diagnostics Pasteur; Marnes-la-Coquette, France). In some experiments, fixed cells were further incubated with FITC-phalloidin (Sigma) (1:100 for 15 min at 22C) to label actin.
All images were observed with an epifluorescent photomicroscope Axioskop (Carl Zeiss; Le Pecq, France) equipped with a high-resolution (752 × 582 pixels) and sensitive 3CCD cooled camera LH750RC3 (Lhesa; Cergy Pontoise, France). The excitation wavelengths λex were 546 nm for Rhm8 -PVA and 490 nm for FITC-phalloidin. Throughout the study, and for longitudinal studies over longer time periods, the 3CCD camera had its mode “auto mode” turned off and manual controls were used to adjust the image intensity and the acquisition time to 4 sec. These controls were kept at an identical level during the studies. Standardized controls of fluorescence were used every day, with proper calibrations of the microscope and 3CCD camera to yield comparable conditions for image collections and analyses. Red and green fluorescence controls were obtained using Inspeck Red (580/ 605) and Green (505/515) Microscope image intensity calibration standards (Molecular Probes). Digitalized images were acquired with Photomat software and were stored on an external Magneto Optical Disk Device (Sony; Tokyo, Japan).
All results are expressed as means ± SEM.
Results and Discussion
Synthesis and Characterization of Rhm8 -PVA
The synthesis and characterization of the fluorescent vasopressin antagonist Rhm8 -PVA are described elsewhere (Durroux et al. in press; see Materials and Methods). The dissociation constant (Ki) of Rhm8 -PVA was 70 pM, as assessed by competition experiments on CHO cells expressing V1a receptors. The nonspecific binding does not represent more than 20% of total binding for a concentration at Rhm8 -PVA near the Ki value. Its Ki for V1b, V2, and oxytocin receptors was 27 nM, 7490 nM, and 1.23 nM, respectively. The ability of Rhm8 -PVA to block functional responses of V1a receptors was also investigated. Rhm8 -PVA alone was unable to stimulate inositol 1,4,5-trisphosphate (InsP3) synthesis. It competively inhibited vasopressin-mediated synthesis of InsP3 in CHO cells expressing V1a receptors with an inactivation constant (Kinact) of 70 pM. We investigated the binding and internalization properties of Rhm8 -PVA in rat hepatocytes, in which it binds to vasopressin receptors but does not cause their activation.
Rhm8 -PVA Does Not Stimulate Ca2+ Signals and Competitively Blocks Vasopressin-mediated Ca2+ Signals
The ability of Rhm8 -PVA to activate hepatocyte V1a receptors was assessed by fluorescence spectroscopy to evaluate its ability to evoke Ca2+ signals in cell suspensions. The presence of a rhodamine-labeled antagonist did not affect [Ca2+]i signals, as measured using the fluorescent indicator quin2. Addition of 20 nM Rhm8 -PVA abolished Ca2+ signals elicited by 1 nM vasopressin within 1 min (Figure 1). Rhm8 -PVA itself, at concentrations from 0.1 nM to 100 nM, was unable to evoke Ca2+ signals (see the control curve in Figure 2). These concentrations are above its Ki for binding to V1a vasopressin receptors and its Kinact for inhibiting vasopressin-mediated synthesis of InsP3 in the CHO cell line. The fluorescent probe did not prevent the [Ca2+]i rises generated by the αl-adrener-gic agonist norepinephrine (not shown). Because vasopressin and norepinephrine use different hormone receptors (Morel et al. 1992; Exton 1996), these data confirm the primary antagonistic action of Rhm8 -PVA at the level of V1a vasopressin receptors.
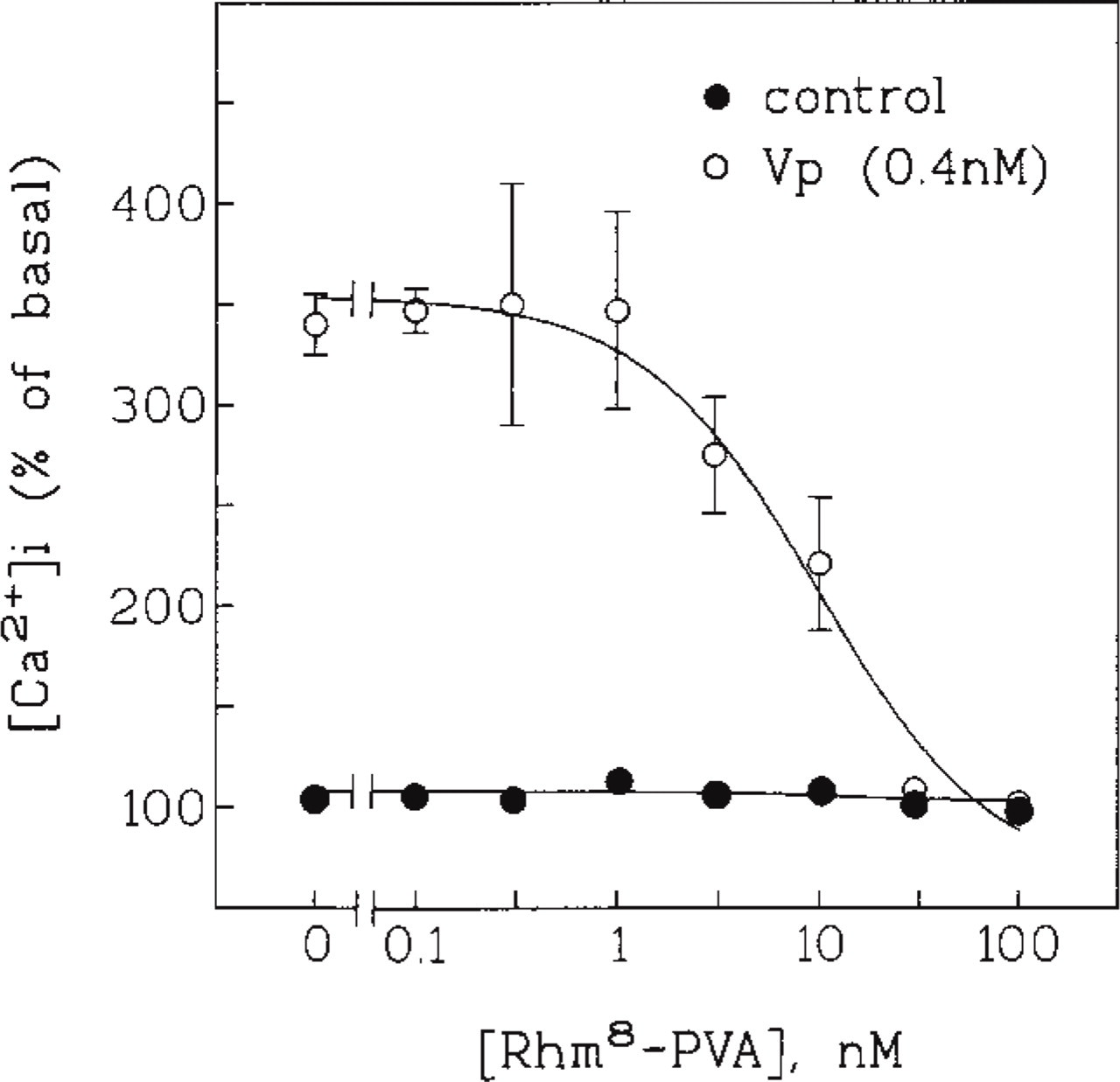
Competitive inhibition of vasopressin-elicited Ca2+ signals by Rhm8 -PVA. Effects of various concentrations of Rhm8 -PVA on the [Ca2+]i rise elicited by 0.4 nM vasopressin. Rhm8 -PVA was added 1 min before vasopressin. Values shown are [Ca2+]i measured after stimulation relative to their basal [Ca2+]i (171 ± 6 nM). They are the mean ± SEM of 18 separate experiments.
Competition inhibition assays were used to establish the inactivation constant of Rhm8 -PVA in suspensions of rat hepatocytes. The inhibition by 20 nM Rhm8 -PVA of increases in [Ca2+]i caused by various concentrations of vasopressin (not shown), and the effects of various concentrations of Rhm8 -PVA on Ca2+ signals elicited by a concentration of vasopressin near the Kinact (0.4 nM), were determined (Figure 2). The control curve shows that Rhm8 -PVA at concentrations of 0.1-100 nM did not alter basal [Ca2+]i. The concentration of Rhm8 -PVA that resulted in half-maximal inhibition of Ca2+ signals induced by 0.4 nM vasopressin was 7 nM. The Kinact for Rhm8 -PVA inactivation of V1a receptors in rat hepatocytes was calculated to be 3.0 nM. These results show that, as in CHO cells expressing V1a vasopressin receptors (Durroux et al. in press), Rhm8 -PVA is unable to activate V1a vasopressin receptors in suspensions of rat hepatocytes.
Visualization of Rhm8 -PVA Binding to Cell Surface of Rat Hepatocytes
The ability of Rhm8 -PVA to bind and to be internalized was evaluated by epifluorescence microscopy with a high resolution and sensitive 3CCD camera to improve the resolution of the image and highlight the binding of the fluorescent vasopressin analogue Rhm8 -PVA to V1a receptors (see Materials and Methods). The experiments were carried out with single cells and also with doublets and triplets of rat hepatocytes that retain certain aspects of structural and functional epithelial polarity, particularly the expression of basal, lateral and canalicular domains found in the intact liver (Gautam et al. 1987; Graf and Boyer 1990; Watanabe et al. 1991b; Spray et al. 1994; Boyer 1997).
The 3CCD camera revealed the fluorescent signal at the periphery of cells. In cells incubated with 10 nM Rhm8 -PVA for 30 min at 4C, the fluorescent antagonist bound to the surface of cells, presumably the plasma membrane of rat hepatocytes (compare the peripheral labeling of the single cell and the triplet observed in Figure 3A to that seen in Figure 3B). Significant fluorescent signal was also observed intracellularly, close to nuclei. Because this labeling was present in cells incubated without Rhm8 -PVA (Figure 3B), it may have been due to the high autofluorescence signal consistently found in rat hepatocytes (Berthon et al. 1984).
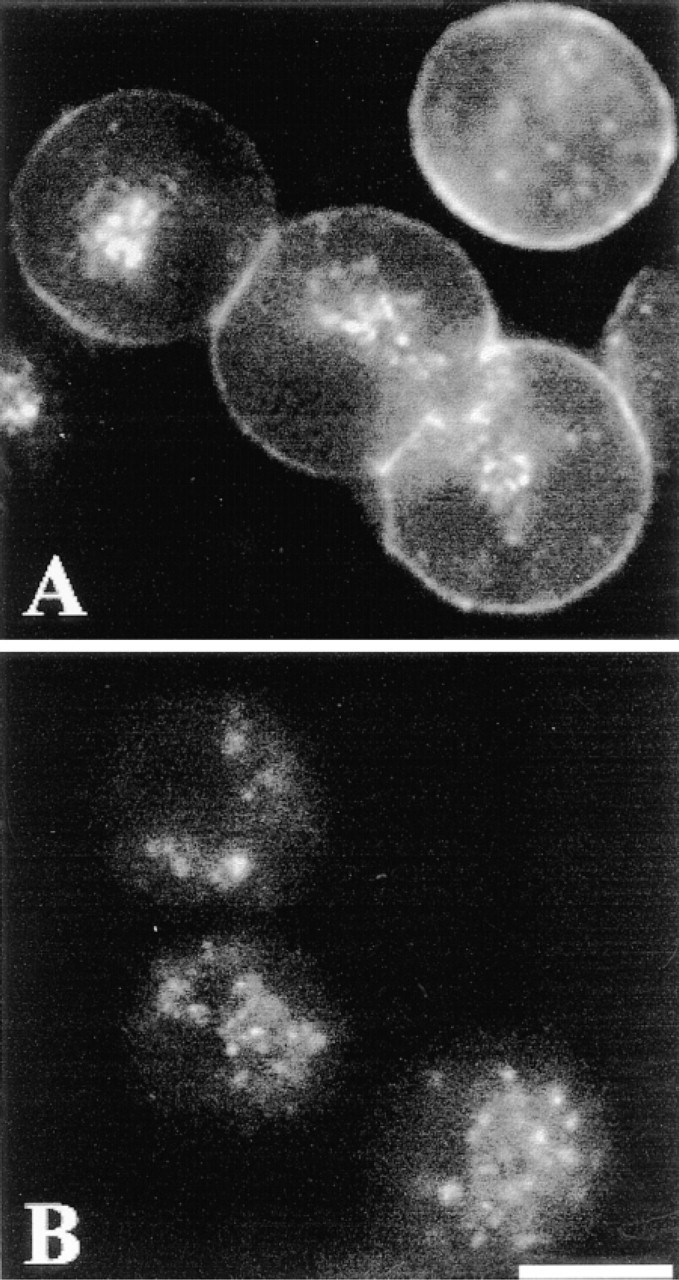
Visualization of Rhm8 -PVA labeling of rat hepatocytes. Cells plated on collagen-coated glass coverslips were incubated in the presence (A) or absence (B, autofluorescence) of 10 nM Rhm8 -PVA for 30 min at 4C. The fluorescent antagonist bound to the periphery of rat hepatocytes. The image is representative of 122 single cells, 100 doublets, and 30 triplets observed from seven cell preparations. Bar = 10 μm.
The cell doublet system has been widely used to investigate a variety of cytoskeletal-dependent functions because the networks of actin and tubulin remain polarized in these preparations (Boyer 1997). For example, the actin cytoskeleton is responsible for apical canalicular contraction in hepatocytes (Watanabe et al. 1991a), an event that is elicited by vasopressin (Yamaguchi et al. 1991; Dufour et al. 1995), increased [Ca2+]i (Oshio and Phillips 1981), and cytosolic ATP (Watanabe et al. 1991a). The position of V1a vasopressin receptors at the cell periphery was compared to that of actin in doublets or triplets of rat hepatocytes by double labeling experiments (rhodamine-labeled V1a receptors and fluorescein-labeled actin).
Rat hepatocyte doublets were incubated (as described for Figure 3) with 10 nM Rhm8 -PVA for 30 min at 4C to label V1a receptors, washed, fixed, and then incubated with FITC-phalloidin (see Materials and Methods) for labeling actin. Both Rhm8 -PVA (Figure 4A) and FITC-phalloidin (Figure 4B) labeled a very thin layer at the periphery of the cells, corresponding to the basolateral domain of the plasma membrane. The fluorescent V1a receptor antagonist was not observed in the apical area, a domain strongly labeled by FITC-phalloidin in the hepatocyte doublet (Nathanson et al. 1994a).
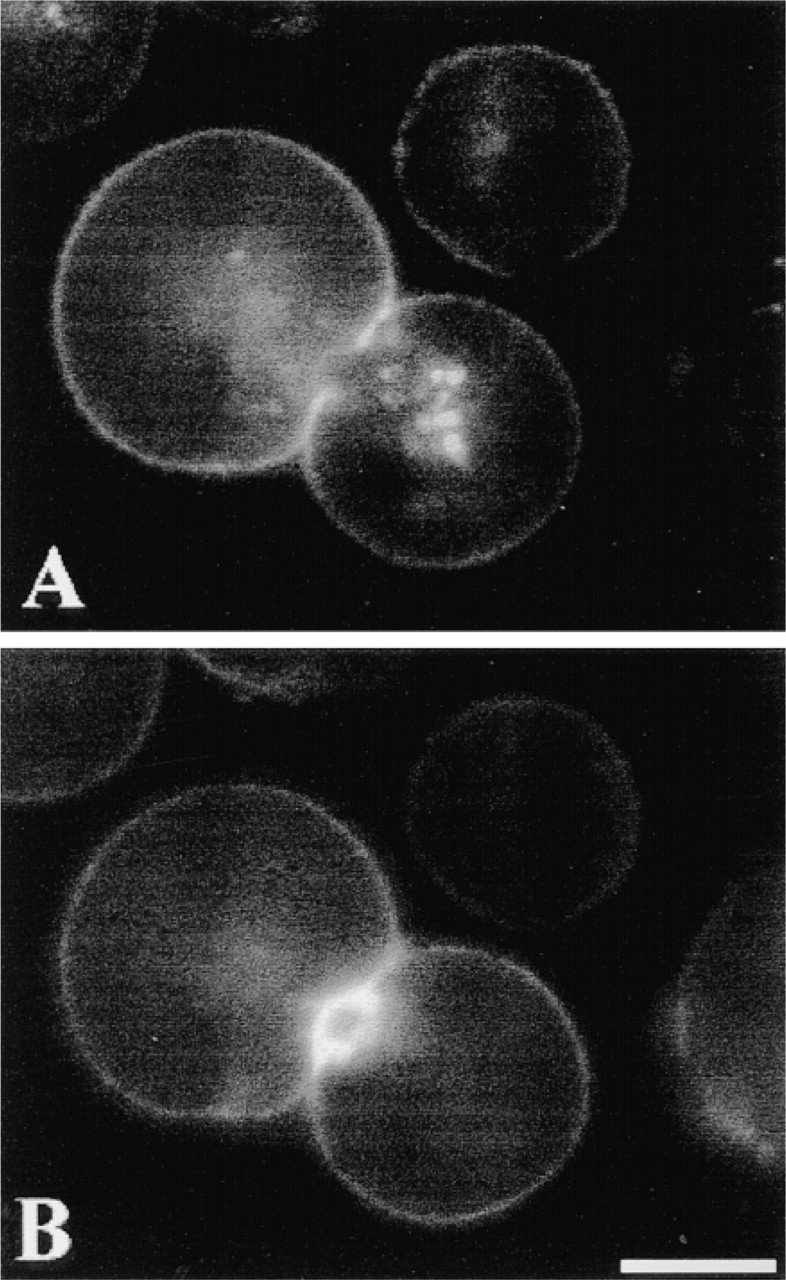
Localization of Rhm8 -PVA (red fluorescence) and FITC-phalloidin (green fluorescence) probes by double labeling of rat hepatocytes. The cell doublet was first incubated with 10 nM Rhm8 -PVA for 30 min at 4C to label V1a receptors, then with FITC-phalloidin to label actin (see Materials and Methods). Both Rhm8 -PVA (A) and FITC-phalloidin (B) labeled the basolateral domain of the plasma membrane, but the fluorescent V1a receptor antagonist was excluded from apical area. The image is representative of 151 single cells and 92 doublets from five cell preparations. Bar = 10 μm.
Specificity of Rhm8 -PVA Binding to V1a Receptors
The V1a receptor specificity of Rhm8 -PVA binding was assessed by investigating its displacement by the nonfluorescent antagonist V4253 and by the natural hormone vasopressin. The experiment was performed at 4C throughout to prevent any receptor-mediated endocytosis (Lutz et al. 1992). V1a receptors were saturated by preincubation with 5 μ? of the nonfluorescent antagonist V4253 for 15 min, then labeled by incubation with 10 nM Rhm8 -PVA for 30 min (Figures 5A and 5B). Homologous competition of the fluorescence by the nonfluorescent antagonist abolished the labeling of V1a receptors at the surface of the cells. Specificity of Rhm8 -PVA binding to V1a receptors was also illustrated by its heterologous displacement by 100 nM of vasopressin (Figures 5C and 5D). The suppression of labeling at the cell surface confirmed the high selectivity of the novel fluorescent indicator Rhm8 -PVA for V1a receptors in rat hepatocytes.
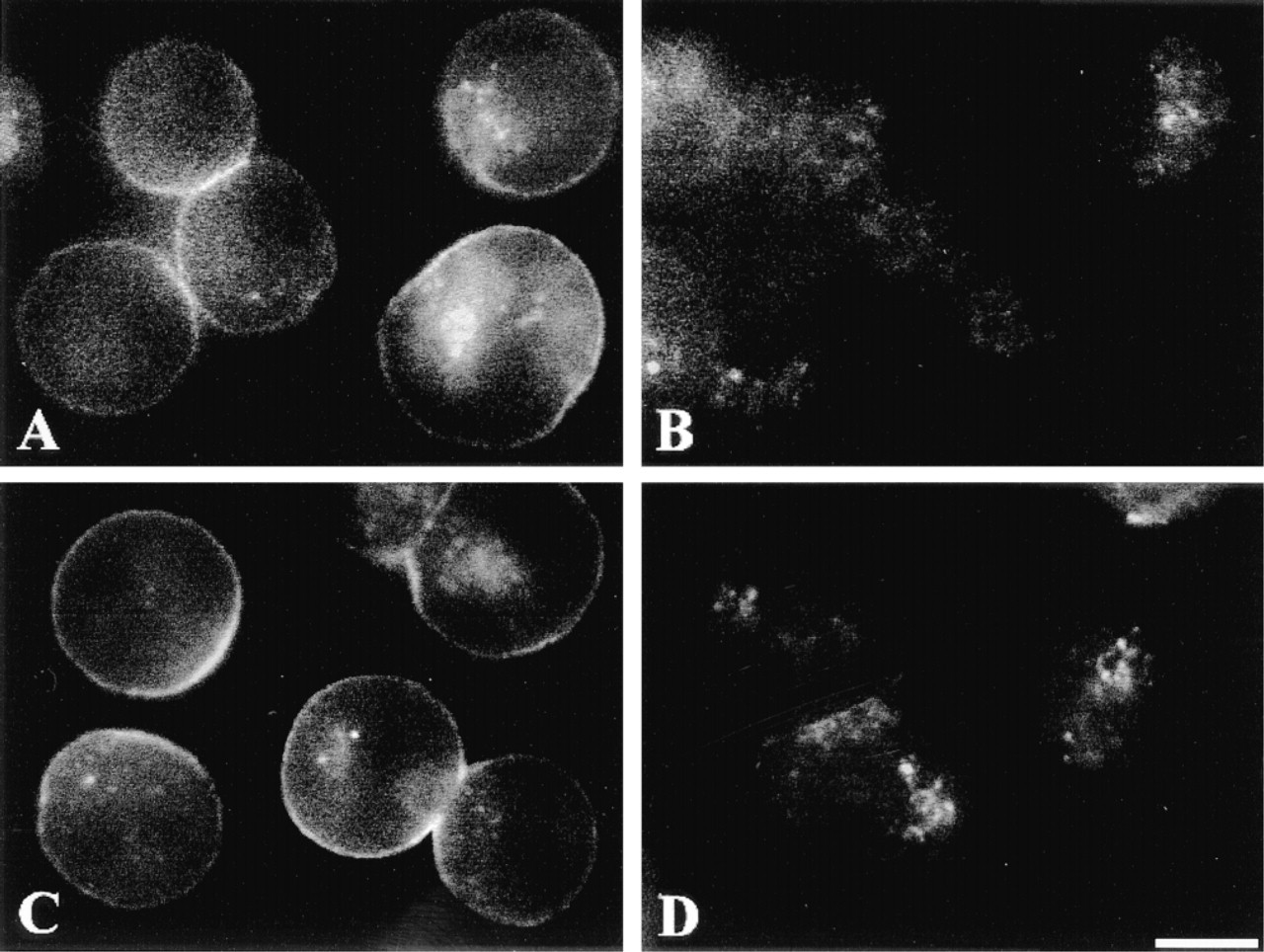
Specificity of Rhm8 -PVA binding to V1a receptors in rat hepatocytes. Cells were incubated in the presence of 10 nM Rhm8 -PVA for 30 min at 4C (A) and after prior incubation with 5 μM of the nonfluorescent antagonist V4253 at 4C for 15 min to saturate V1a vasopressin receptors (B). The image is representative of 98 single cells and 35 doublets observed from two cell preparations. Cells were equilibrated with 10 nM Rhm8 -PVA for 30 min at 4C (C) and after prior incubation with 100 nM vasopressin at 4C for 15 min (D). The image is representative of 81 single cells, 28 doublets, and 10 triplets observed from two cell preparations. Bar = 10 μm.
Vasopressin Induces Internalization of V1a Receptors
We compared the internalization of Rhm8 -PVA and vasopressin in rat hepatocytes. We investigated whether Rhm8 -PVA binding to the cell surface of rat hepatocytes was abolished by prior endocytosis of the V1a receptors. V1a receptor-mediated endocytosis was initiated by preincubation for 5-60 min with 100 nM vasopressin at 37C, and then cells were cooled at 4C to examine binding of Rhm8 -PVA (10 nM for 30 min). For all periods of preincubation, prior vasopressin-mediated endocytosis of V1a receptors abolished antagonist binding to the cell surface (Figures 6B-6D) whereas in non-preincubated cells, Rhm8 -PVA bound to the cell surface of rat hepatocytes (Figure 6A). The V1a receptor of rat hepatocytes is therefore specifically internalized by the natural agonist vasopressin. In addition, the development of vasopressin-mediated receptor endocytosis detected in fluorescence microscopy is as rapid (less than 5 min) as that previously observed using tritiated vasopressin in suspensions of rat hepatocytes (Fishman et al. 1985).
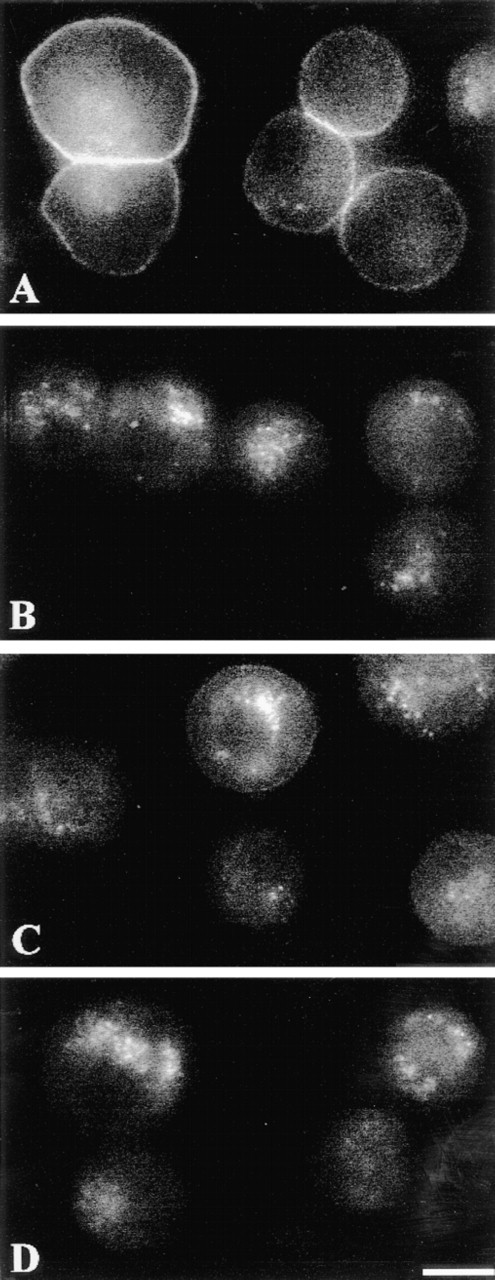
Internalization of V1a receptors by vasopressin. Cells were incubated with 10 nM Rhm8 -PVA for 30 min at 4C (A), and after prior incubation with 100 nM of the natural hormone vasopressin for 5 min (B), 30 min (C), or 60 min (D) at 37C, to initiate receptor-mediated endocytosis of V1a receptors in rat hepatocytes. The image is representative of at least 65 single cells, 26 doublets, and 12 triplets per condition from two cell preparations. Bar = 10 μm.
Rhm8 -PVA Does Not Induce Internalization of V1a Receptors
Because Rhm8 -PVA is a specific antagonist of V1a receptors in rat hepatocytes, we investigated whether incubation of cells at 37C can elicit endocytosis of the Rhm8 -PVA/V1a receptor complex. Rat hepatocytes were equilibrated with 10 nM Rhm8 -PVA for 30 min at 4C to label V1a receptors (Figure 7A) and then warmed to 37C to initiate receptor-mediated internalization of the fluorescent complex for 1, 3, 5, 15, and 30 min (Figures 7B-7D). The Rhm8 -PVA label remained at the cell surface even after 30 min. Therefore, unlike vasopressin, Rhm8 -PVA is unable to evoke receptormediated endocytosis of V1a receptors in rat hepatocytes.
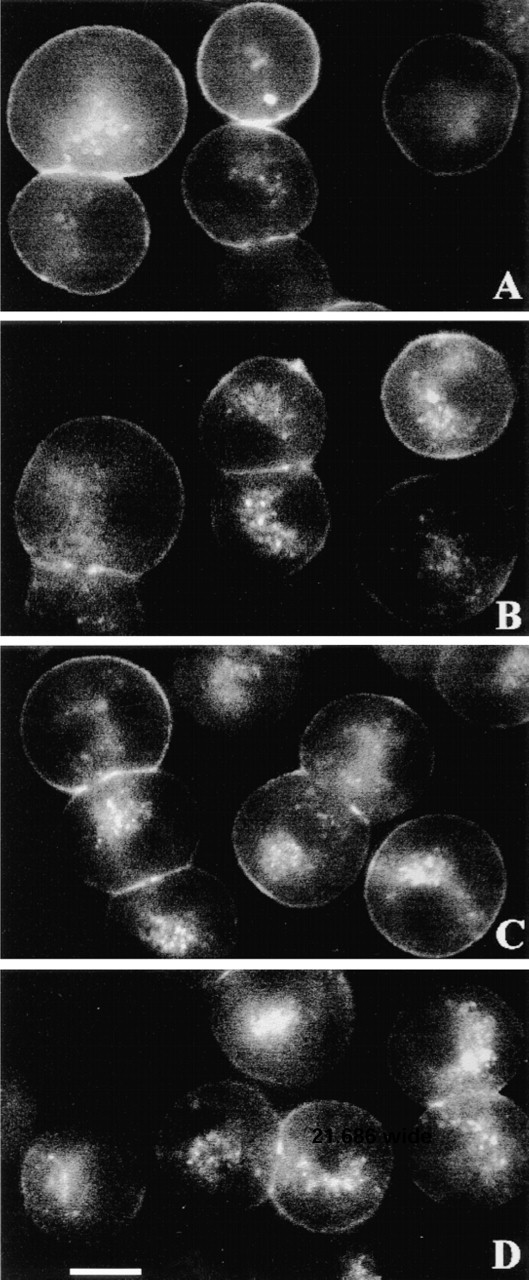
V1a receptors are not internalized by Rhm8 -PVA. Rat hepatocytes were incubated with 10 nM Rhm8 -PVA for 30 min at 4C to label V1a vasopressin receptors (A) and then warmed for 5 min (B), 15 min (C), or 30 min (D) at 37C to initiate receptor-mediated internalization of the fluorescent complex. The shortest periods of rewarming (1 and 3 min) at 37C are not illustrated, but they show that, as in B-D, the fluorescent complex remained bound to the cell surface. The image is representative of at least 69 single cells, 25 doublets, and 15 triplets per condition from two cell preparations. Bar = 10 μm.
Effect of Temperature on Rhm8 -PVA Binding
The incubation temperatures at 4C and 37C had little effect on binding and noninternalization of 10 nM Rhm8 -PVA by rat hepatocytes. At the two incubation temperatures, Rhm8 -PVA remained bound to the cell surface for 15 and 30 min (not shown), whereas vasopressin-mediated endocytosis of V1a receptors is complete by that time (see Figure 7).
Conclusion
In summary, the vasopressin fluorescent linear antagonist Rhm8 -PVA binds V1a receptors with a high affinity and does not elicit cellular responses in single cells and multiplets of rat hepatocytes. The inability of Rhm8 -PVA bound to the cell surface to be internalized indicates that occupancy of the hepatocyte vasopressin Via receptor in the absence of receptor activation is insufficient to elicit receptor-mediated endocytosis, as previously observed with another fluorescent antagonist in the A10 rat smooth muscle cell line expressing V1a vasopressin receptors (Lutz et al. 1992). Rhm8 -PVA is therefore a useful analogue for investigating the distribution of cell surface receptors in a variety of living cells that express V1a receptors.
Footnotes
Acknowledgements
Supported by the Association pour la Recherche sur le Cancer, la Fondation pour la Recherche Médicale, and the Association Française Contre les Myopathies.
We thank Raymonde Leuillet and Denise Reuter for excellent technical assistance and A. Edelman for help in editing the manuscript.