
Abstract
The beagle dog with naturally occurring periodontal disease is one of the most widely used animal models in periodontal research for histological studies on disease pathogenesis and on the effect of potential therapeutic regimens. However, previous studies were restricted to morphological assessment of immunocompetent cells because of the lack of available cell-specific markers. In this study we systematically characterized the specificity and immunoreactivity of a panel of anti-human antibodies for identification (ABC method) of immunocompetent cells in formalin-fixed, EDTA-decalcified, paraffin-embedded inflamed periodontal tissues obtained from six beagle dogs. Canine lymph nodes and a panel of different human tissues served as positive controls. Polyclonal anti-CD3 immunolabeled canine T-lymphocytes specifically. Anti-CD79 α (clone HM57) reacted with B-lymphocytes and plasma cells, and CD79 α (clone JCP117) showed no staining in canine tissues. Neutrophils, monocytes, small macrophages, and keratinocytes reacted with an anti-myeloid/histiocyte antibody (clone MAC387). Anti-CD68 (clones PG-M1 and EBM11) immunolabeled large macrophages and plasma cells. Clone EBM11 also stained osteoclasts and cementoclasts. With the exception of JCB117, all antibodies revealed similarly favorable immunolabeling of canine and human immunocompetent cells. Long-term EDTA decalcification appeared to weaken immunostaining of plasma cells with HM57. MAC387 and CD68 can be used to distinguish macrophages in different differentiation stages in canine periodontal tissues.
Keywords
P
Because of the extremely limited availability of human periodontal tissues, systematic analysis of the pathogenesis of periodontal disease and new therapeutic methods requires reproducible, well-characterized animal models. The beagle dog is one of the most widely used animal models for histological studies in periodontal research (Page and Schroeder 1982; Giannobile et al. 1994). However, owing to the lack of available cell-specific markers for canine tissues, previous studies were restricted to conventional histological evaluation and morphological assessment of inflammatory reactions.
Whereas many monoclonal and polyclonal antibodies are available for the different types of inflammatory cells in human tissues, there are no specific anti-canine antibodies. Some studies have shown crossreactivity of an anti-human CD3 and an anti-human myeloid/histiocyte antibody with T-cells, neutrophils, and certain macrophages in canine extraoral soft tissues (Ferrer et al. 1992,1993; Day 1996,1997; Pérez et al. 1997). However, little is known about their sitespecific immunoreactivity in periodontal tissues and the effect of decalcification procedures critical for the routine histological evaluation of the densely mineralized tissues of teeth and alveolar bone. Furthermore, the usefulness of cell-specific markers for B-lymphocytes, plasma cells, and different subsets of macrophages in canine tissues remains to be explored. Because the specific nature of immunocompetent cells involved in periodontitis is not well understood, such baseline information is necessary for future studies aimed at evaluating the molecular mechanisms underlying the progress and resolution of periodontal disease. Therefore, the aim of the present study was to systematically characterize the specifity and immunoreactivity of a panel of anti-human antibodies in the identification of immunocompetent cells in diseased canine periodontal tissues.
Materials and Methods
Tissue Preparation
Canine tissues were obtained from six 5-7-year-old female beagle dogs which were purchased from federally licensed dealers (Harland Sprague-Dawley, Indianapolis, IN, and Marshall Farms, Northrose, NY) and were subsets of an ongoing periodontal therapy study. All animals showed advanced, naturally occurring periodontal disease. On the day of sacrifice, the dogs were anesthesized with 3% isoflurane/O2 gas (Iso-thesia; Burns Veterinary Supply, Rockville Centre, NY) and sacrificed by exsanguination and perfusion with 10% neutral buffered formalin (VWR Scientific Products; West Chester, PA) through the carotid arteries. Mandibular and maxillary segments including the second, third, and fourth premolars were dissected and further fixed by immersion in 10% neutral buffered formalin for 48 hr at 4C. After the initial 24 hr, each jaw segment was separated into three smaller segments containing only one tooth with its surrounding periodontal tissues (alveolar bone, gingiva, periodontal ligament fibers) to improve tissue fixation. After 48 hr of fixation, the specimens were washed and then decalcified in 5% neutral buffered EDTA using a magnetic stirrer at 4C for 7-8 months. The endpoint of decalcification was radiographically determined.
Gingival soft tissue biopsy specimens and submandibular lymph nodes were also included in the study. These specimens were fixed in 10% neutral buffered formalin for only 24 hr at 4C.
All tissues were dehydrated through a graded series of ethanols and chloroform and then embeddded in low melting point (56C) paraffin (Paraplast; Oxford Labware, St Louis, MO). Serial sections at a thickness of 5-8 μm were prepared and collected on commercially available positively charged glass slides (Superfrost Plus; Fisher Scientific, Fair Lawn, NJ). All canine teeth were cut in a buccolingual direction. The sections were dried on a heating plate (Fisher Scientific) at 45C for 48 hr to increase tissue adherence to the slides. Representative sections were stained with hematoxylin and eosin (H&E) for conventional histological assessment (Figure 1).
Immunohistochemical Procedures
Immunohistochemical labeling of immunocompetent cells was carried out using the avidin-biotin complex (ABC) method (Hsu et al. 1981) as described previously (D'Souza et al. 1992). Representative sections were deparaffinized in xylene (EM Science; Gibbstown, NJ) and rehydrated through a descending series of ethanol concentrations. Unless otherwise mentioned, each incubation step during the immunohistochemical procedure was performed in a humidified chamber at room temperature (24C) and was followed by thorough rinsing in 0.01 M Dulbecco's phosphate-buffered saline (PBS; Irvine Scientific, Santa Ana, CA) three times for 5 min.
Endogeneous peroxidase activity was blocked by treating the sections with 0.3% hydrogen peroxide (Sigma; St Louis, MO) in methanol for 30 min. Then one or a combination of the following pretreatment methods was carried out to expose the antigen structures under investigation: (a) enzymatic digestion with hyaluronidase (4000 U/ml; bovine testicular, Sigma) for 30 min; (b) enzymatic digestion with proteinase K (Dako; Carpinteria, CA) for 6-8 min; (c) antigen retrieval using 1% sodium dodecyl sulfate (SDS, Sigma) in PBS for 5 min at room temperature (Brown et al. 1996); (d) antigen retrieval by boiling (95-100C) the sections in 0.01 M citrate buffer (pH 6) for 10 min. Afterwards, nonspecific protein binding was blocked by incubation with 0.01 M PBS containing 10% normal goat serum (Vector Laboratories; Burlingame, CA) for 30 min. Before one of the following commercially available primary antibodies was applied (Table 1), excess buffer was only blotted and rinsing with PBS was avoided: (a) affinity-isolated polyclonal rabbit anti-human CD3 antibody (1:200) as pan-T-lymphocyte marker; (b, c) monoclonal mouse anti-human CD79α antibodies, clones HM57 (1:20) and JCB117 (1:20) as B-cell marker; (d) monoclonal mouse anti-human myeloid/histiocyte antibody, clone MAC387 (1:200); (e, f) monoclonal mouse anti-human CD68 antibodies, clones PG-M1 (1:50-1:100) and EBM11 (1:10-1:50) as macrophage marker. All antibody dilutions were prepared in 0.01 M PBS containing 5% normal goat serum. The incubation time was 1 hr for the polyclonal rabbit antibody (CD3) and 2 hr for the monoclonal mouse antibodies (HM57, JCB117, MAC387, PG-M1, EBM11).
Thereafter, the sections were treated sequentially with biotinylated goat anti-rabbit IgG (1:200) or goat anti-mouse IgG (1:200) (Vector Laboratories) and the avidin-enzyme complex (Vectastain Elite; Vector Laboratories). These treatments were followed by standardized development in 3,3′-diaminobenzidine (DAB; Vector Laboratories). The sections were lightly counterstained with Harris's modified hema toxylin (Fisher Scientific). Immunohistochemical staining and morphological features of inflammatory cells were assessed using a light microscope (Olympus BX40F-3, Olympus Optical, Tokyo, Japan). The immunohistological staining patterns in periodontal tissues were further compared with those of immunocompetent cells in canine lymph nodes.
Panel of anti-human antibodies tested in canine tissues
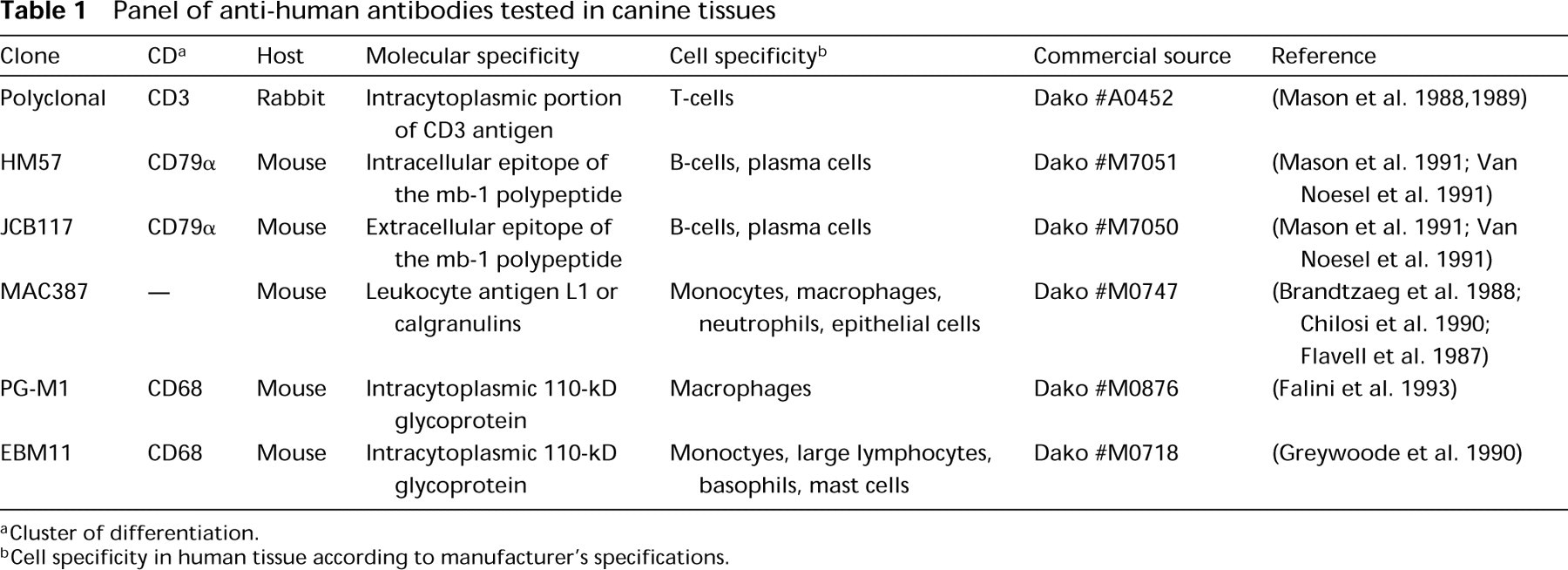
Cluster of differentiation.
Cell specificity in human tissue according to manufacturer's specifications.
For comparison of immunoreactivities of the individual antibodies, the labeling intensity of cells was scored on the following scale of 0 to ++++ using subjective criteria, similar to that described previously (Chilosi et al. 1990; Pulford et al. 1990; Ackermann et al. 1994): 0, no staining; +, weak staining (minimally detectable); ++, moderate staining; +++, strong staining; and ++++, very strong staining (dark brown, almost black).
Controls for Immunohistochemical Staining
Negative control sections were treated by replacement of the primary antibodies either with preimmune rabbit IgG (1:8000) (Dako), preimmune mouse IgG1/2 (1:36) (Dako), or 0.01 M PBS alone. In addition to canine lymph nodes, the following panel of formalin-fixed, paraffin-embedded human tissues was used as positive control for the anti-human antibodies: palatine tonsil, reactive lymph node, periodontal abscess, periapical granuloma, radicular cyst, peripheral giant-cell granuloma, and ossifying fibroma obtained from the archive collection of the Division of Oral Pathology, Department of Stomatology, UTHSC.
Animal Care
All animals used in this study were maintained in accordance with policies established by the Animal Welfare Committee at the University of Texas Health Science Center at Houston and with those of the National Institutes of Health.
Results
Tables 2 and 3 show the immunohistochemical reactivities and subjectively determined staining intensities of the tested antibodies with relevant immunocompetent cells in canine and human tissues. In comparing decalcified hard tissue and nondecalcified soft tissue sections, it appeared that long-term EDTA decalcification did not negatively influence the stability and accessibility of most antigens tested. However, anti-CD79α (clone HM57) showed weaker plasma cell staining in decalcified tissue. Best immunolabeling with anti-CD3 and anti-MAC387 was obtained after gentle enzymatic predigestion using bovine hyaluronidase (4000 U/ml) for 30 min. Both anti-CD68 antibodies (EBM11, PG-M1) required pretreatment with proteinase K for 6-8 min. Hyaluronidase preserved histological structures better than proteinase K. Both CD79α antibodies (HM57, JCB117) showed best immunostaining when enzymatic digestion with hyaluronidase (30 min) was followed by an additional antigen retrieval procedure using 1% SDS for 5 min, as recently described for cryostat sections (Brown et al. 1996). Antigen retrieval by boiling with 0.01 M citrate buffer (pH 6) for 10 min provided comparable staining results in soft tissue sections to those of antigen retrieval using SDS, but caused adherence problems of hard tissue sections to the slides. None of the negative control sections revealed any specific immunohistochemical staining (Figure 2).
Reactivities and staining intensities a of tested anti-human antibodies in canine tissues
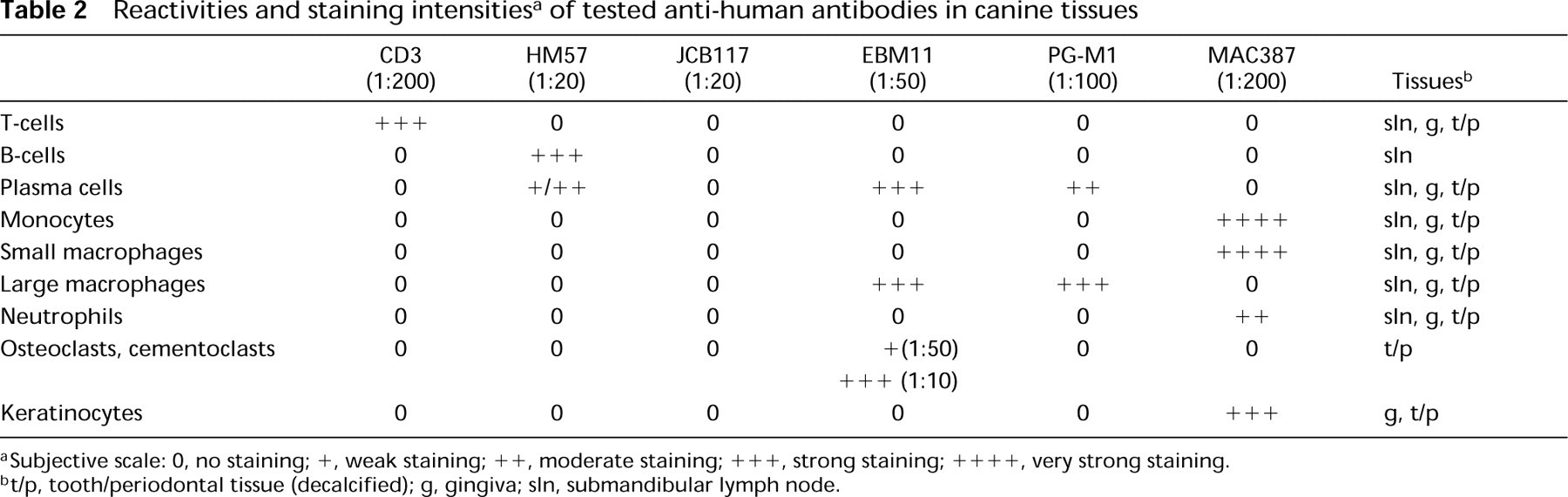
Subjective scale: 0, no staining;+, weak staining;++, moderate staining;+++, strong staining;++++, very strong staining.
t/p, tooth/periodontal tissue (decalcified); g, gingiva; sln, submandibular lymph node.
Reactivities and staining intensities a of tested antibodies in human control tissues
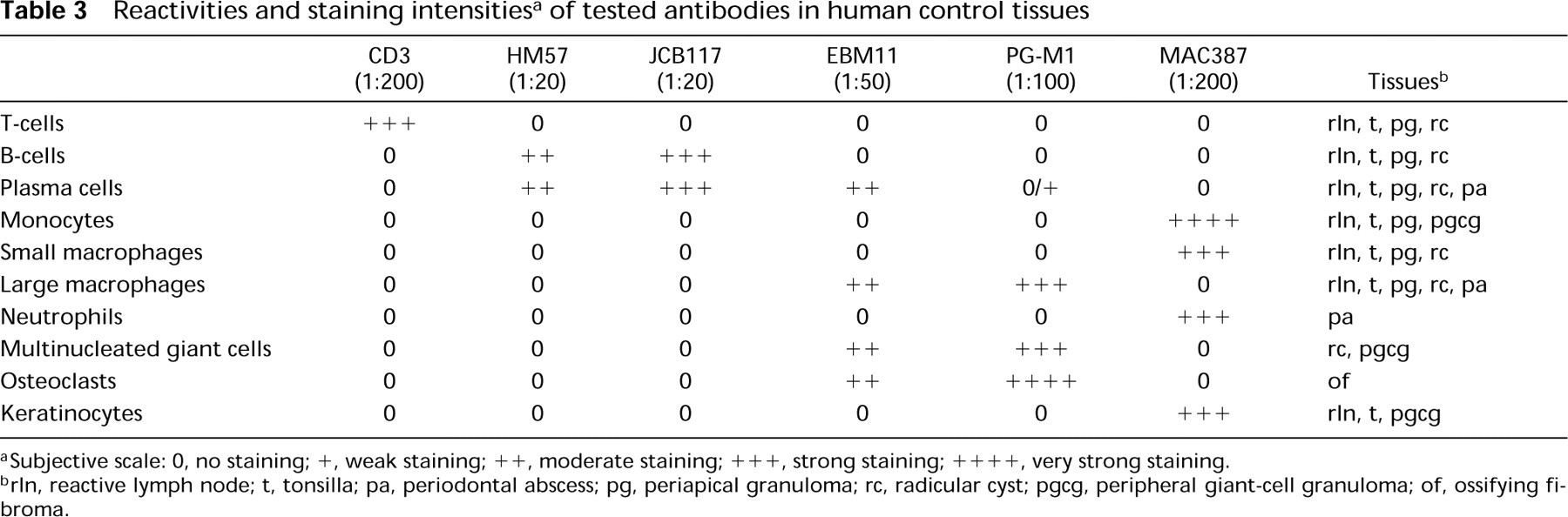
Subjective scale: 0, no staining;+, weak staining;++, moderate staining;++++, strong staining;++++, very strong staining.
rln, reactive lymph node; t, tonsilla; pa, periodontal abscess; pg, periapical granuloma; rc, radicular cyst; pgcg, peripheral giant-cell granuloma; of, ossifying fibroma.
Polyclonal Rabbit Anti-human CD3 Antibody (Pan-T-lymphocyte Marker)
The anti-human CD3 antibody, which was used as a pan-T-lymphocyte marker, stained lymphocytes mainly in the connective tissue subjacent to the oral and sulcus epithelium of chronically inflamed canine gingiva (Figure 3A). Immunostaining was evident in the connective tissue stroma located between elongated epithelial rete pegs (Figure 3B) and in perivascular tissue (Figure 4). In more central parts of the lamina propria, only very few lymphocytes reacted with this antibody. Plasma cells, neutrophils, and macrophages were clearly not stained. Comparison of decalcified and nondecalcified sections revealed no negative influence of longterm EDTA decalcification on the immunoreactivity.
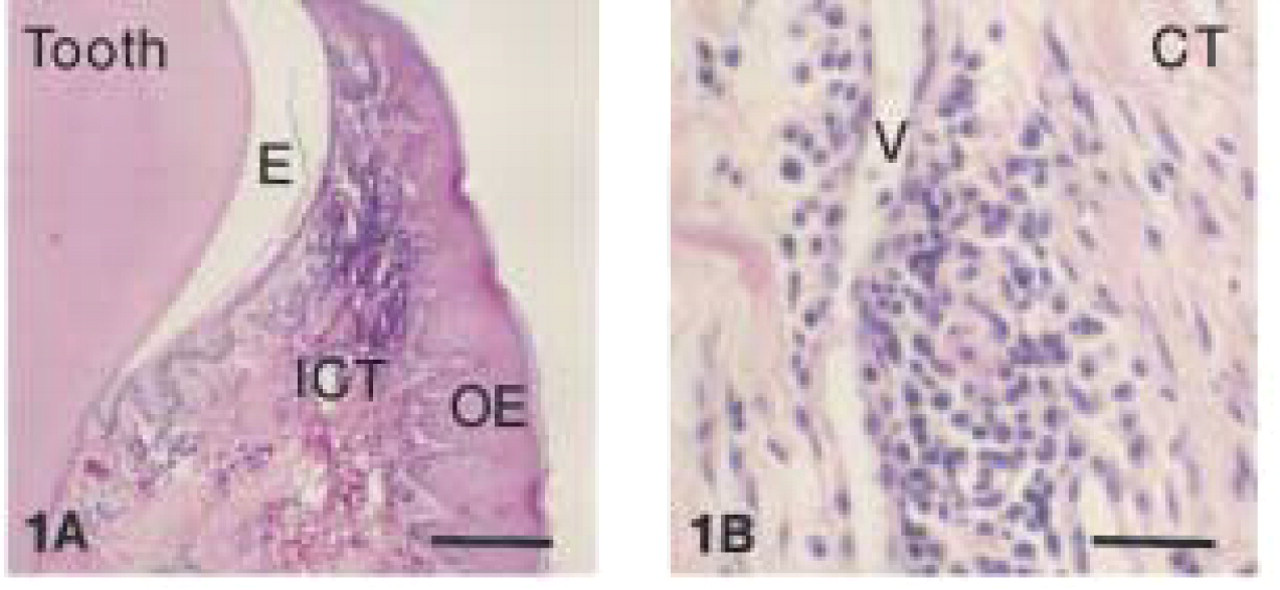
Inflamed marginal gingiva of beagle dog (H&E staining). (

Canine marginal gingiva: negative staining with preimmune mouse IgG. Counterstaining with Harris hematoxylin. Bar = 282 μm.
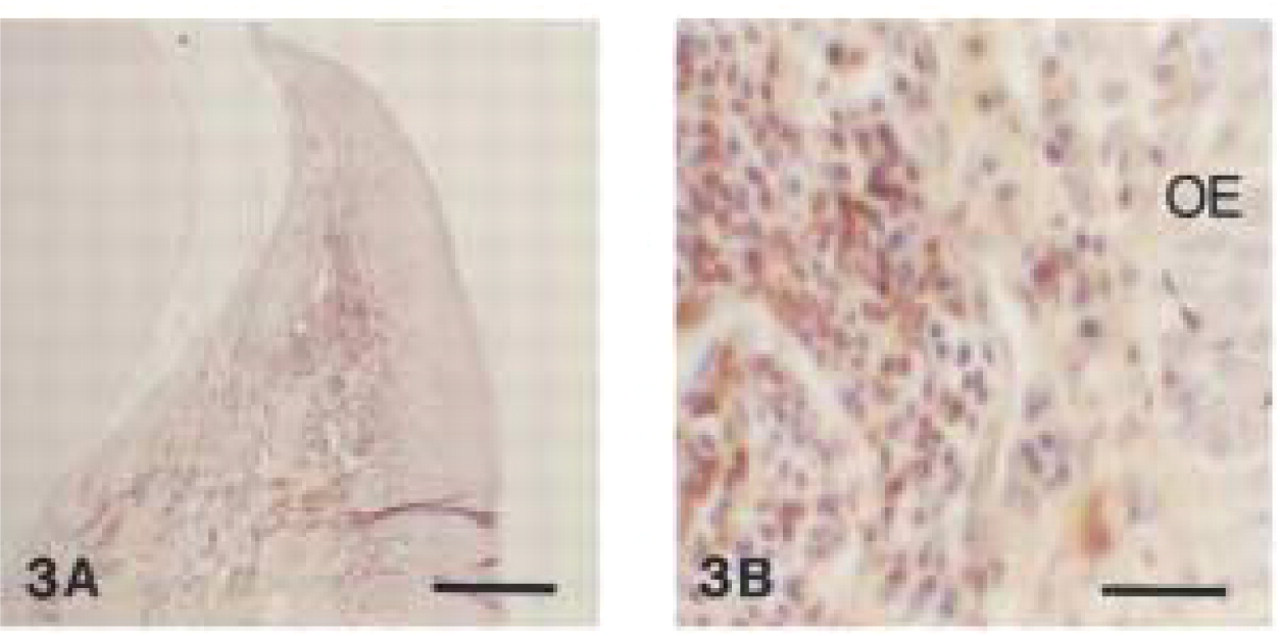
Polyclonal anti-human CD3. (
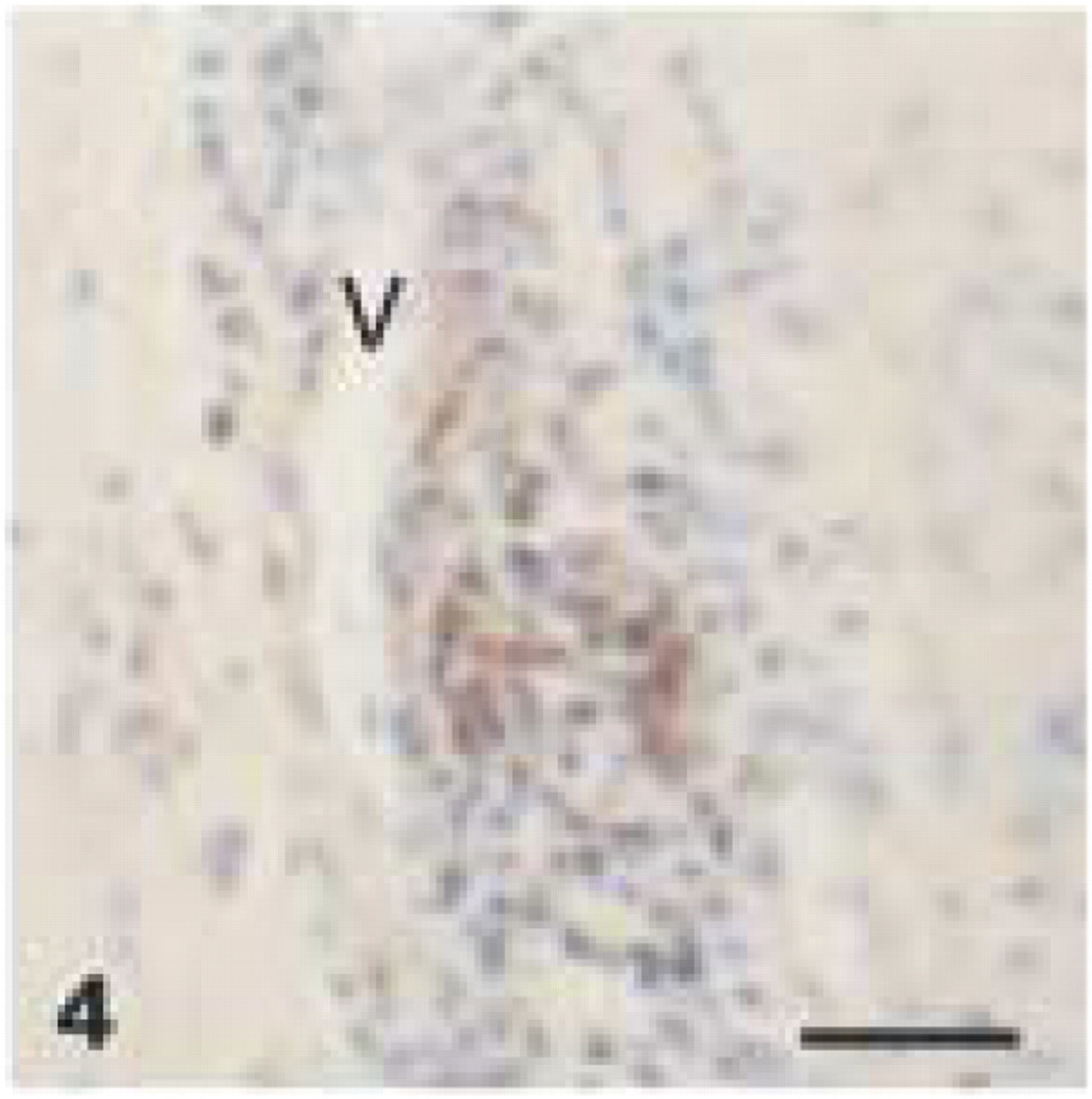
Polyclonal anti-human CD3. Accumulation of CD3+ T-cells around dilated blood vessel (V) in subepithelial gingival connective tissue (EDTA-decalcified specimen). Bar = 28 μm.
Polyclonal anti-human CD3. (
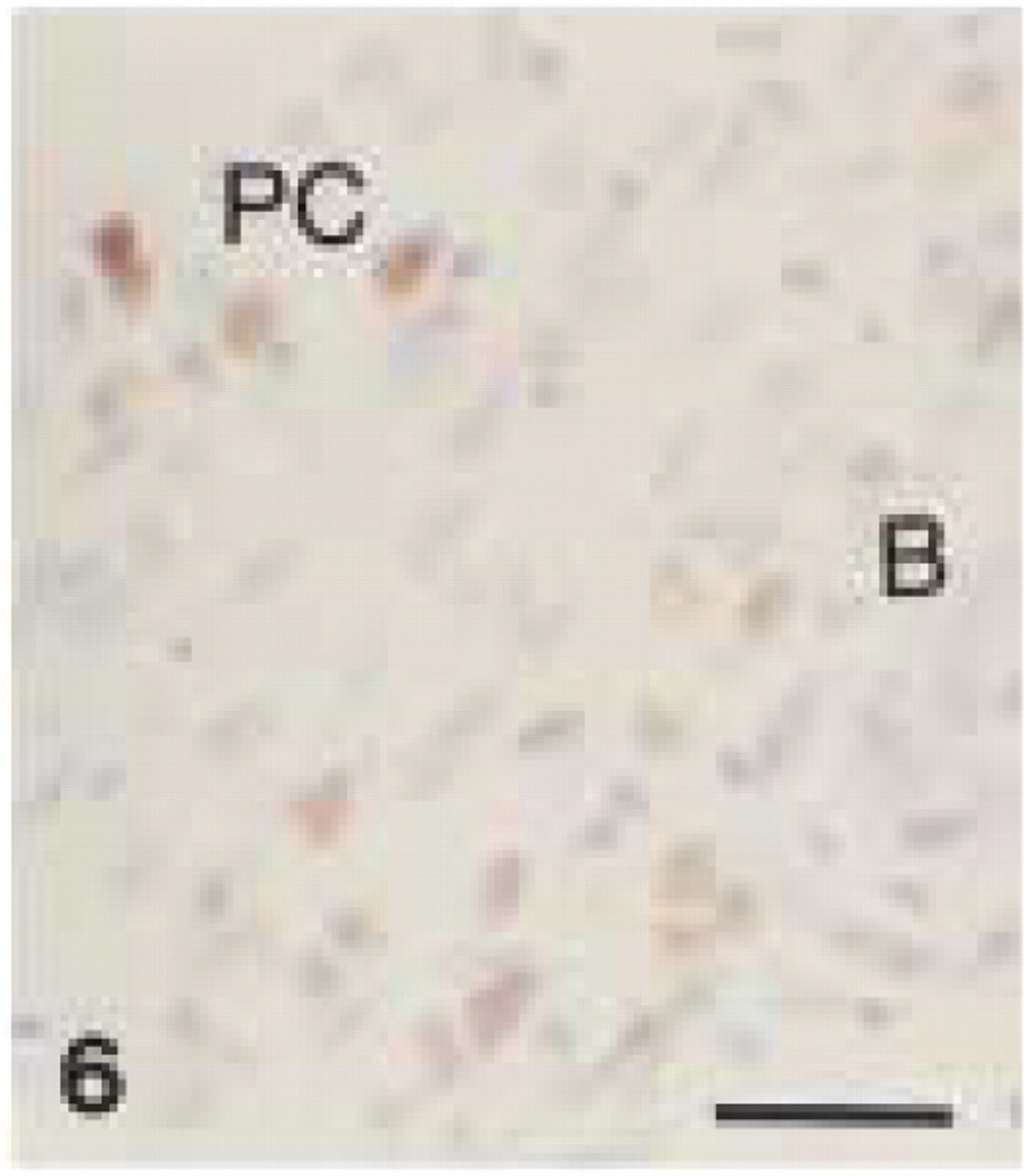
Monoclonal anti-human CD79α (clone HM57). Moderate CD79α+ staining of plasma cells (PC) and B-lymphocytes (B) in lamina propria of canine gingival biopsy. Bar = 28 μm.
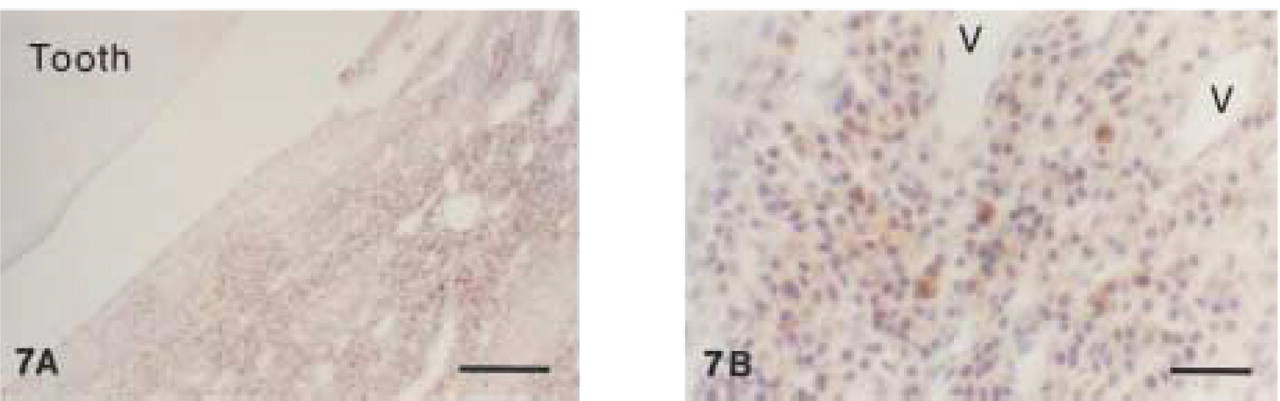
Monoclonal anti-human CD79α (clone HM57). (
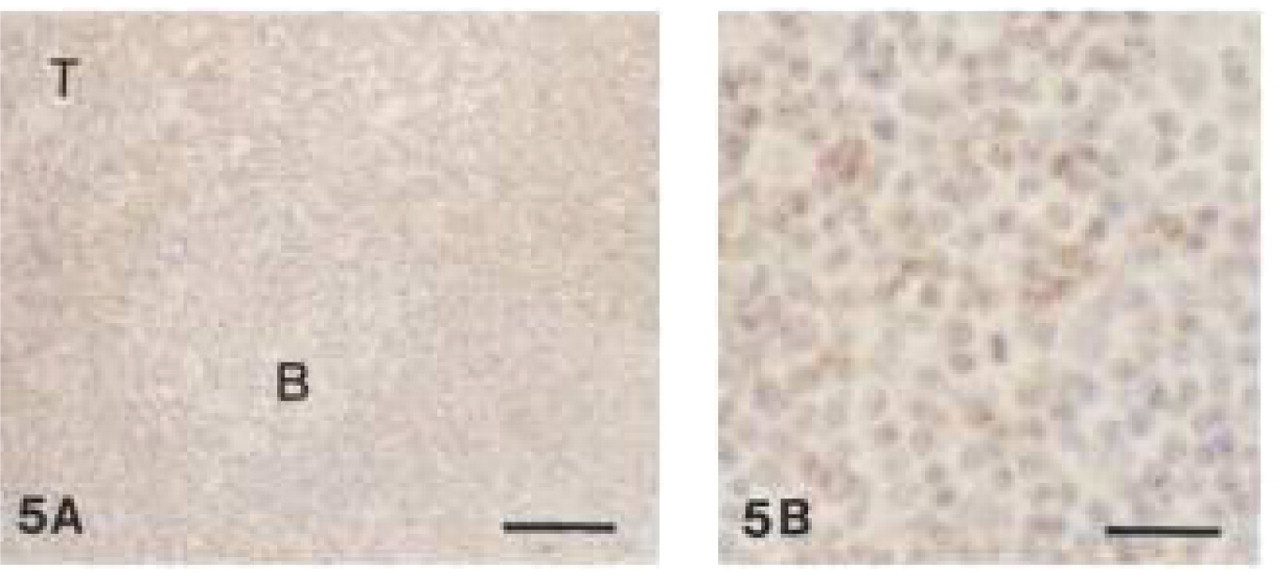
As a positive tissue control, canine lymph nodes treated with anti-CD3 showed immunoreactive lymphocytes in the interfollicular region of the paracortex, whereas follicular cells were negative (Figure 5). Anti-CD3 stained the cytoplasm and cell membrane of T-lymphocytes.
Monoclonal Mouse Anti-human CD79α Antibodies, Clones HM57 and JCB117 (B-cell Marker)
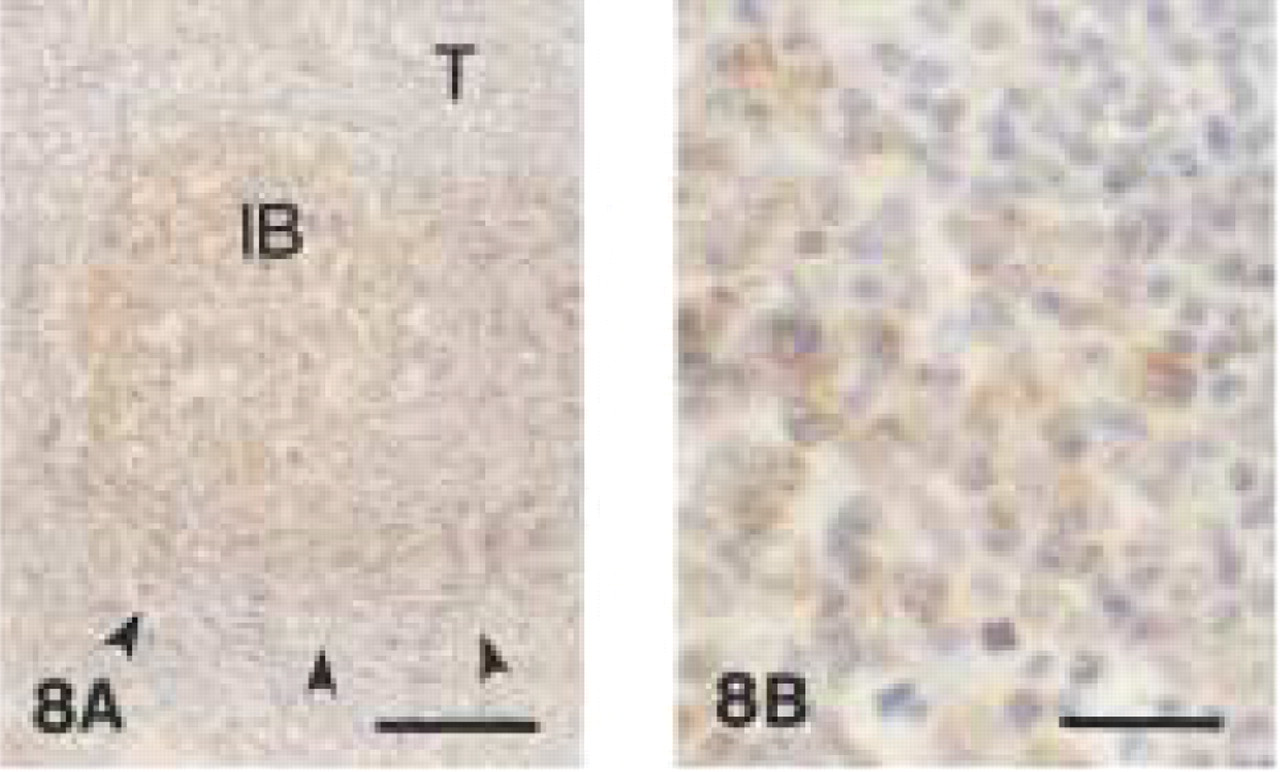
Clone JCB117 showed inconsistent and weak immunoreactivity in canine tissues (data not shown). In contrast, clone HM57 stained plasma cells and B-lymphocytes in sections obtained from gingival biopsies (Figure 6). In decalcified periodontal tissues, immunostaining of plasma cells with HM57 was weaker and more variable than in undecalcified tissues (Figure 7). Plasma cell staining was found mainly in central parts of the gingival lamina propria and in perivascular connective tissue. Clone HM57 reacted with follicular B-lymphocytes and immunoblasts as well as with interfollicular and medullary plasma cells in control canine lymph nodes (Figure 8). However, immunoblasts in the germinal centers of secondary follicles often showed weaker immunoreactivity than the follicular B-cells (data not shown). In canine tissue, anti-CD79α stained the cytoplasm of B- and plasma cells.
Monoclonal Mouse Anti-human Myeloid/Histiocyte Antibody, Clone MAC387 (Monocyte/Neutrophil Marker)
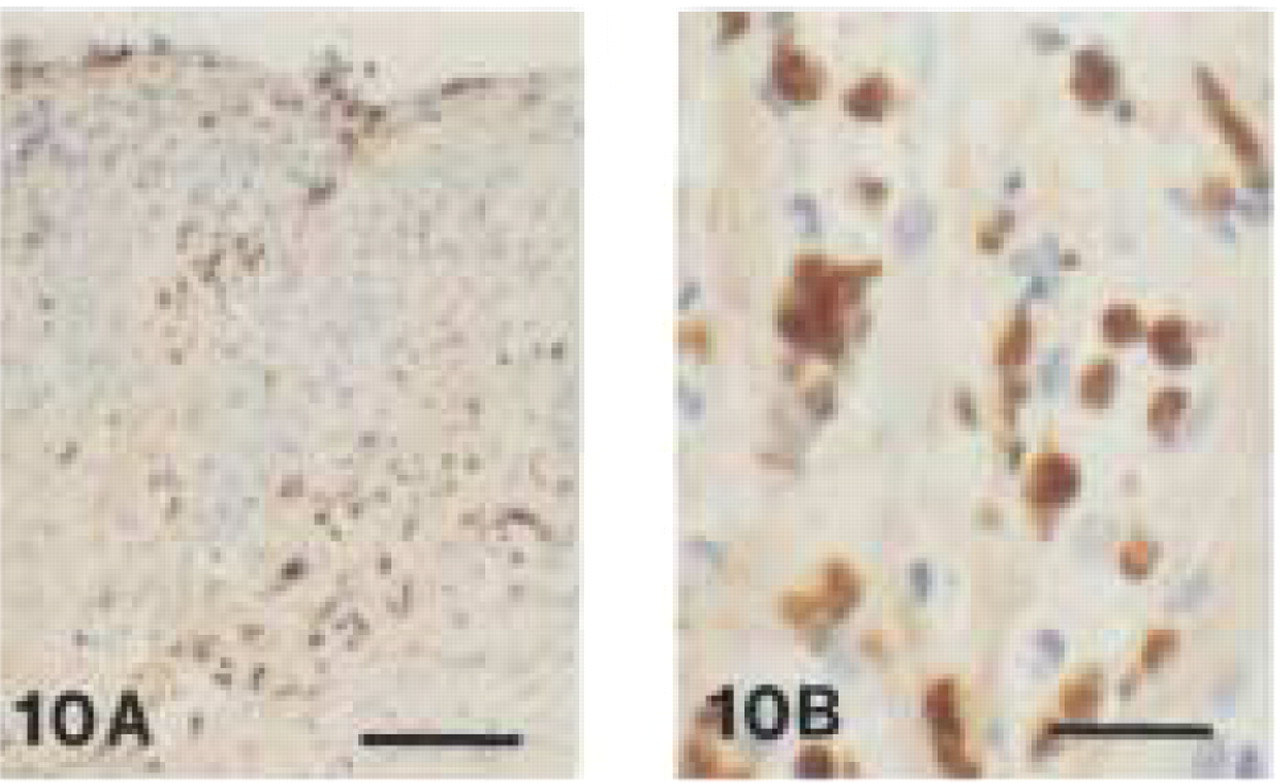
MAC387 reacted strongly with the cytoplasm of monocytes, small macrophages, and neutrophils of acute and chronic inflammatory infiltrates in canine gingival tissues. Both cell types were mainly found in intraand subepithelial layers (Figures 9 and 10). Monocytes in blood vessels were highly immunoreactive with MAC387. Furthermore, immunolabeling was found in the cytoplasm of keratinocytes of the oral epithelium (Figure 9). EDTA decalcification did not appear to impair the degree of immunoreactivity. In canine lymph nodes, small macrophages and some neutrophils in the medulla and paracortical areas showed immunoreactivity with MAC387 (Figure 11). This antibody did not react with large macrophages, lymphocytes, plasma cells, or osteoclasts. The cytoplasmic staining pattern was granular in neutrophils and homogeneously dark brown in monocytes and small macrophages.
Monoclonal anti-human CD79α (clone HM57).
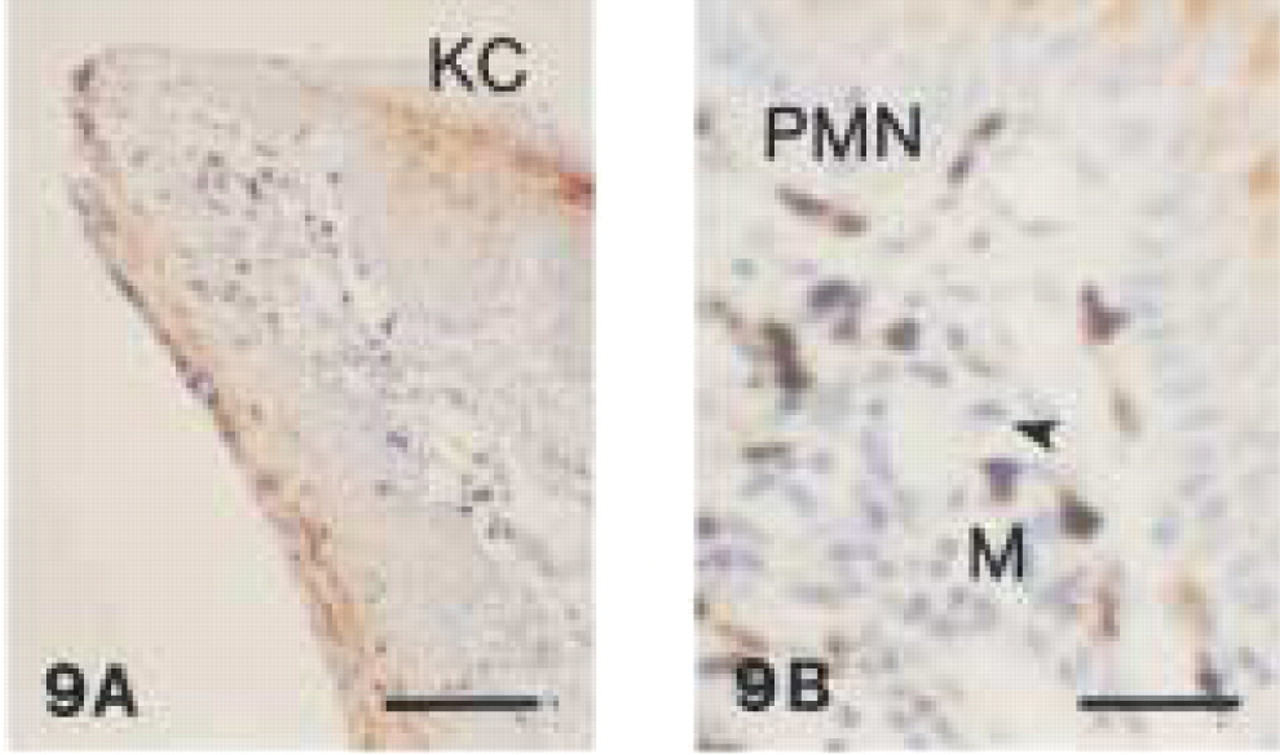
Monoclonal anti-human myeloid/histiocyte antibody (clone MAC 387).
Monoclonal anti-human myeloid/histiocyte antibody (clone MAC 387).
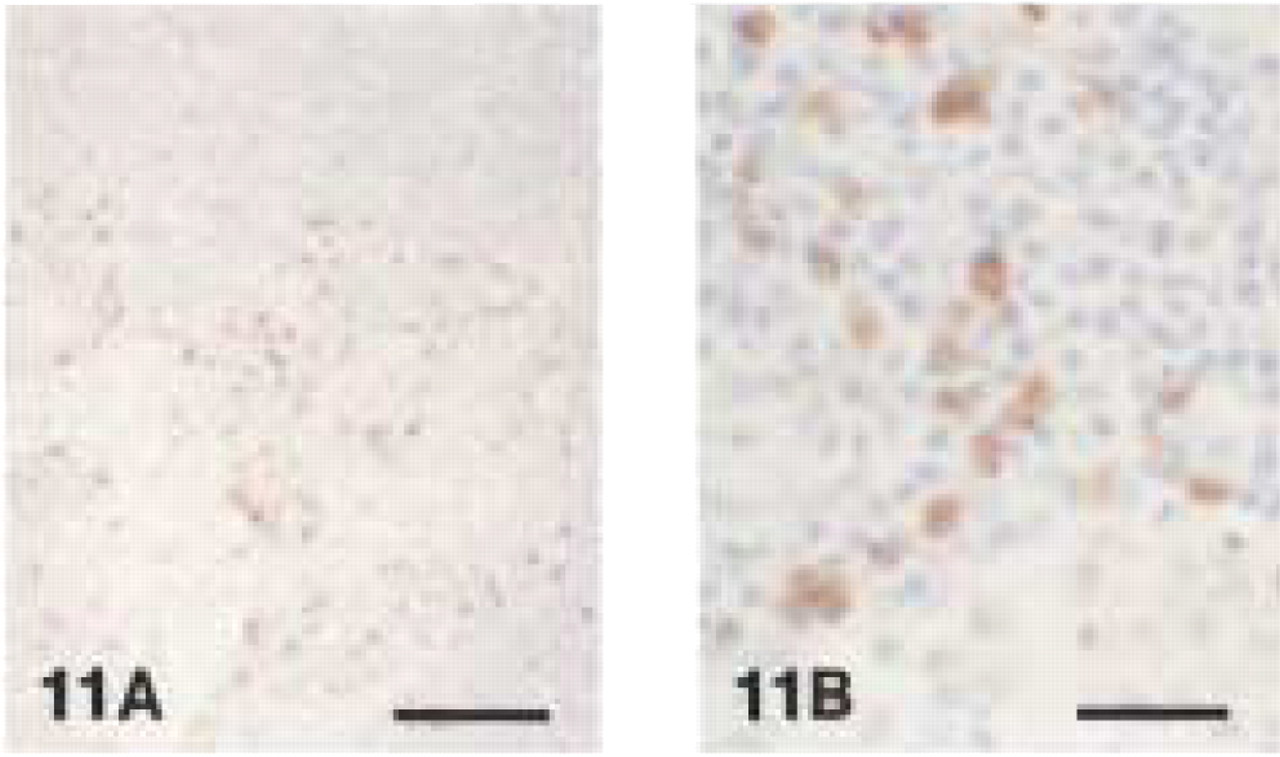
Monoclonal anti-human myeloid/histiocyte antibody (clone MAC 387).
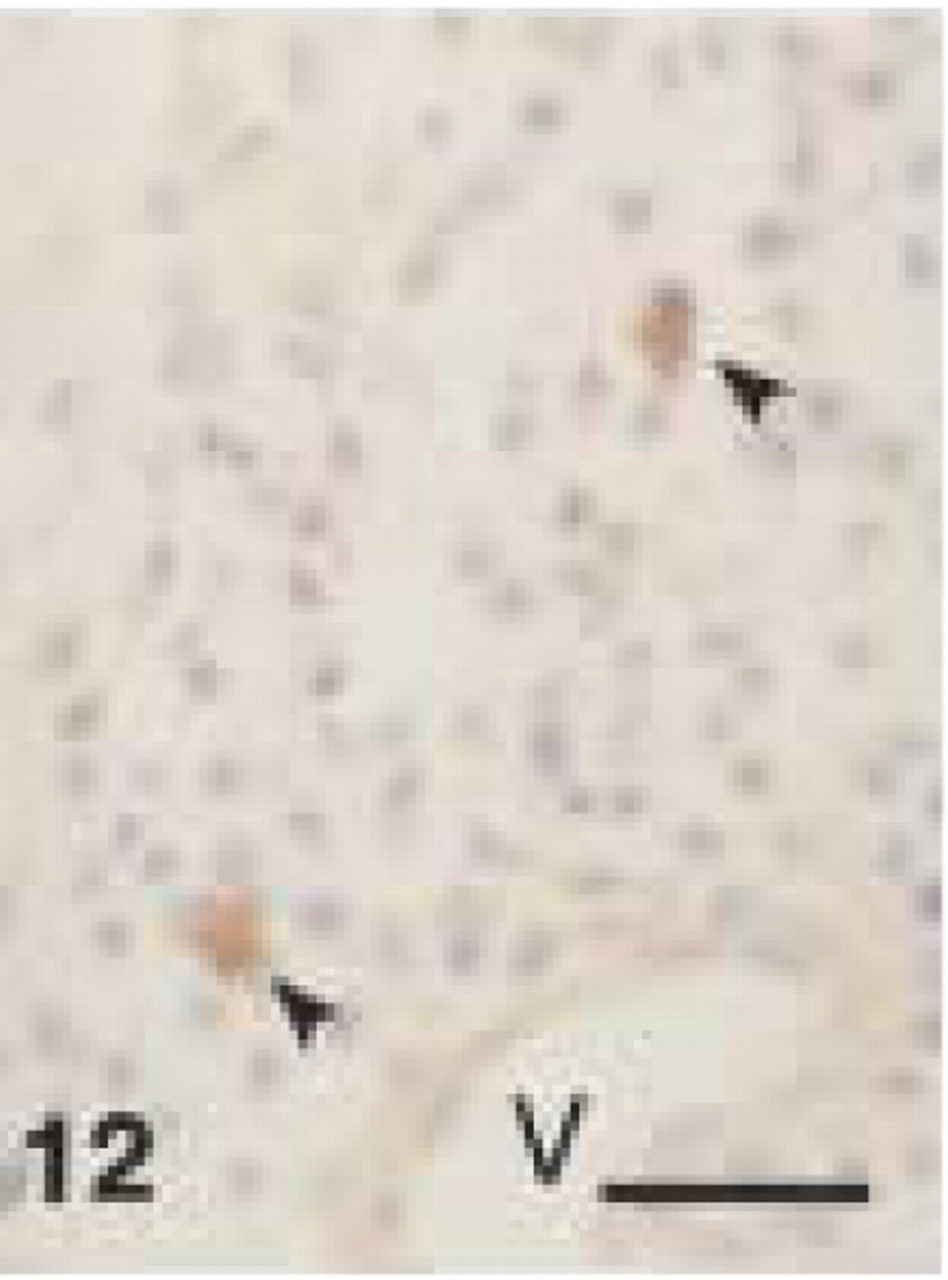
Monoclonal anti-human CD68 (clone PG-M1). CD68+ staining of macrophages (arrowheads) in connective tissue of canine gingiva. V, blood vessel (EDTA decalcified specimen). Bar = 28 μm.
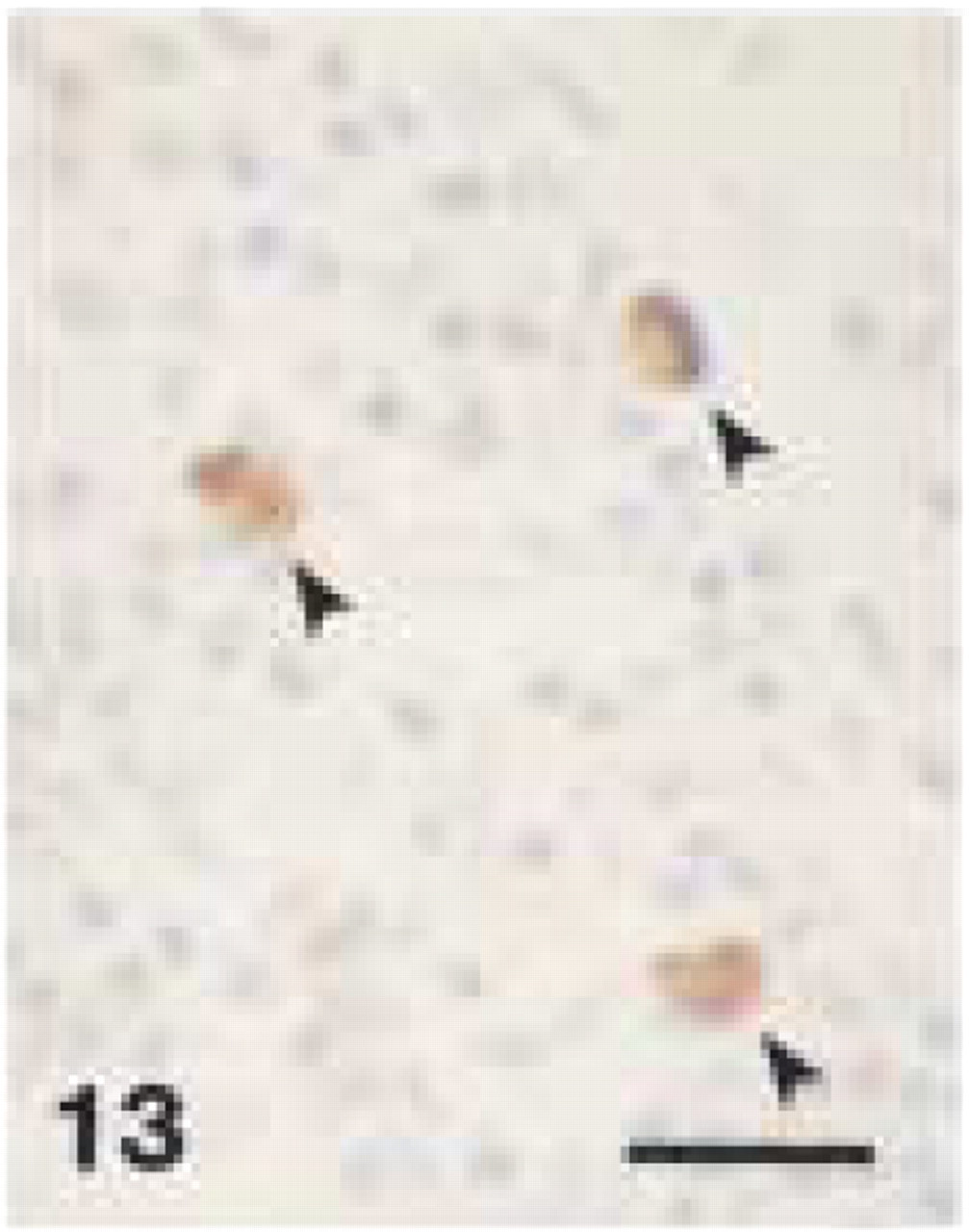
Monoclonal anti-human CD68 (clone PG-M1). CD68+ staining of macrophages (arrowheads) in canine gingival biopsy. Bar = 28 μm.
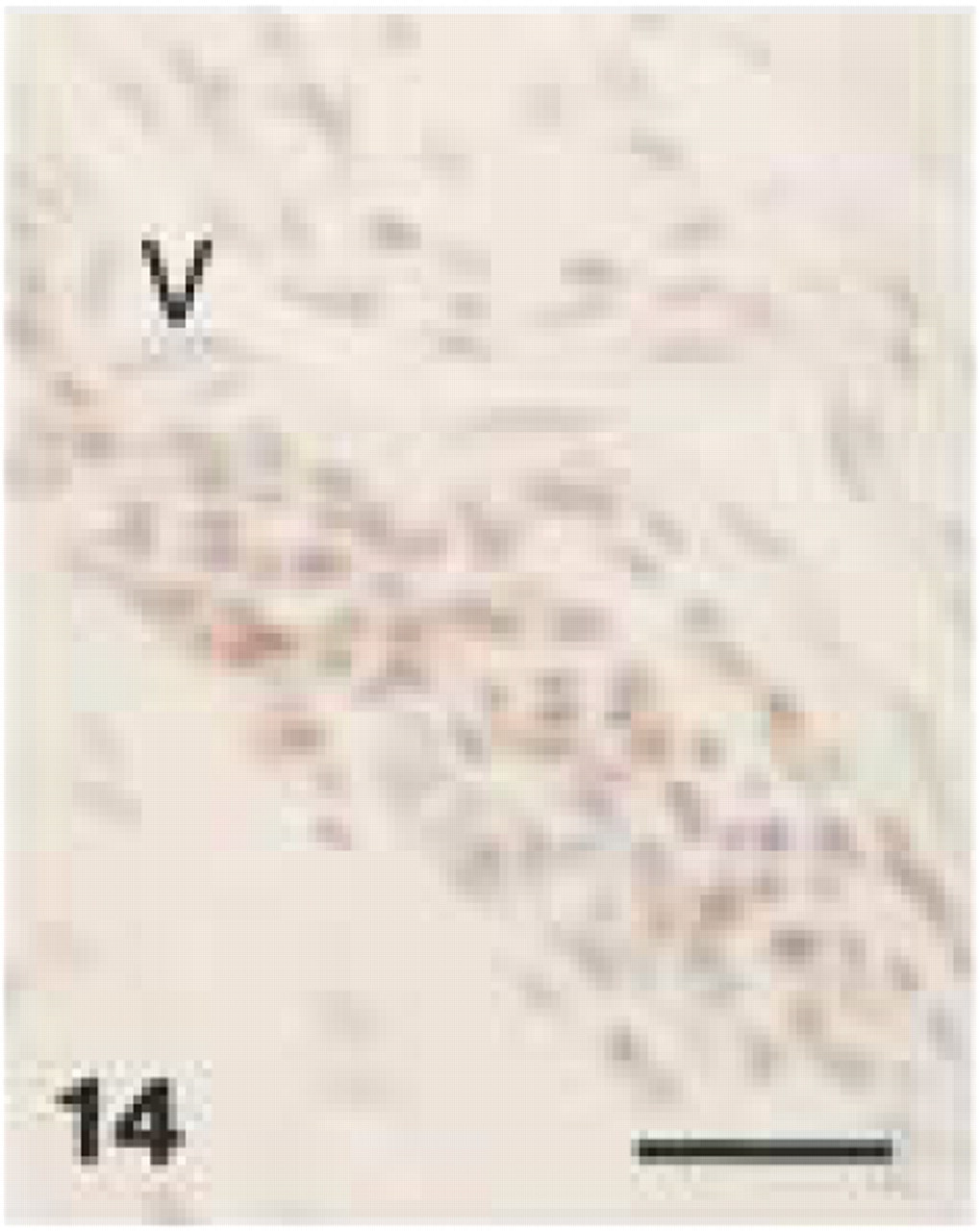
Monoclonal anti-human CD68 (clone PG-M1). CD68+ staining of plasma cells in perivascular connective tissue of canine gingiva. V, blood vessel (EDTA decalcified specimen). Bar = 28 μm.

Monoclonal anti-human CD68 (clone PG-M1). CD68+ staining of plasma cells in lamina propria of canine gingival biopsy. Note typical spoke-like appearance of nuclei. Bar = 28 μm.
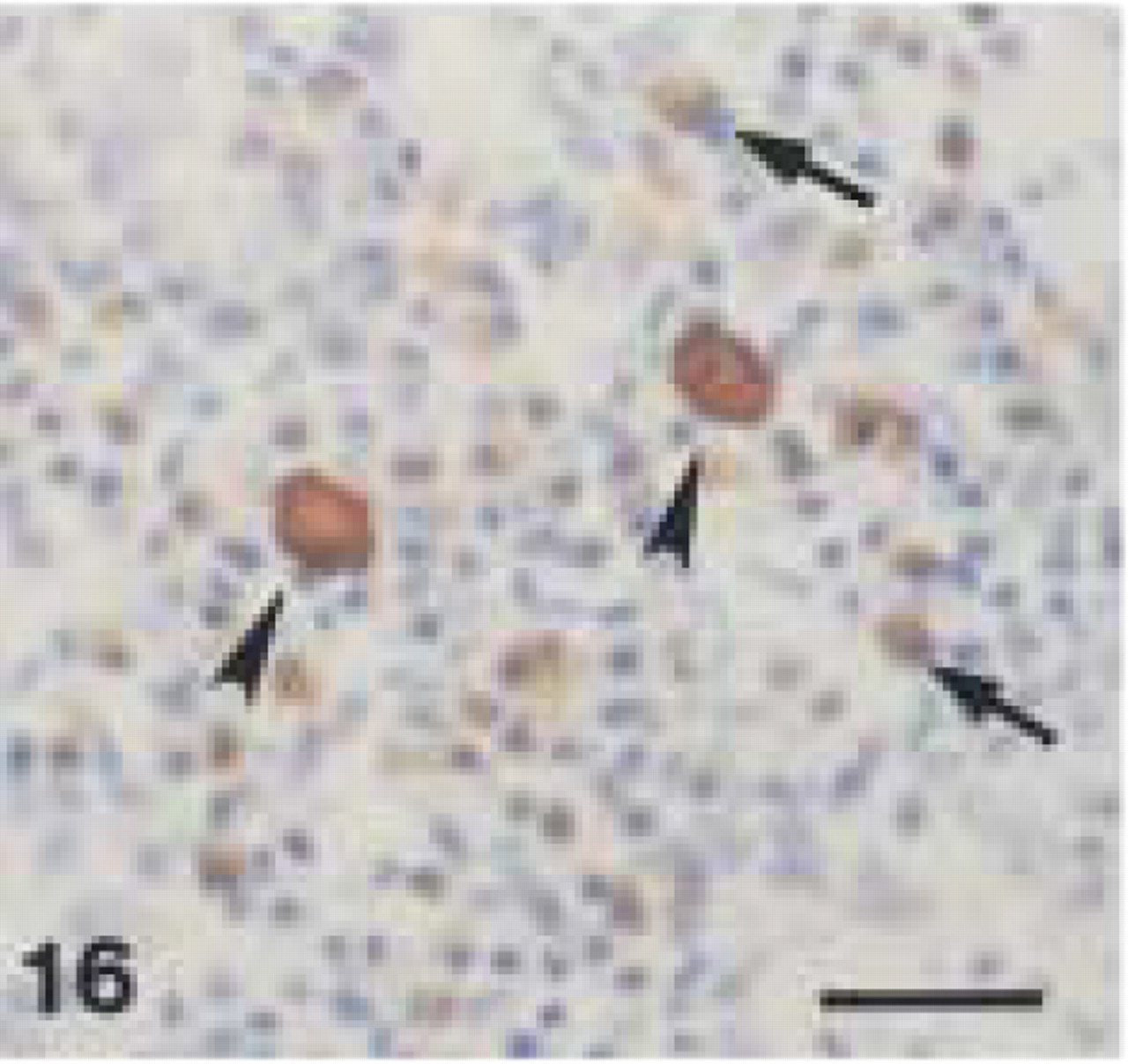
Monoclonal anti-human CD68 (clone PG-M1). Canine lymph node: intense CD68+ staining of large macrophages (arrowheads) and weaker immunolabeling of plasma cells (arrows). Bar = 28 μm.
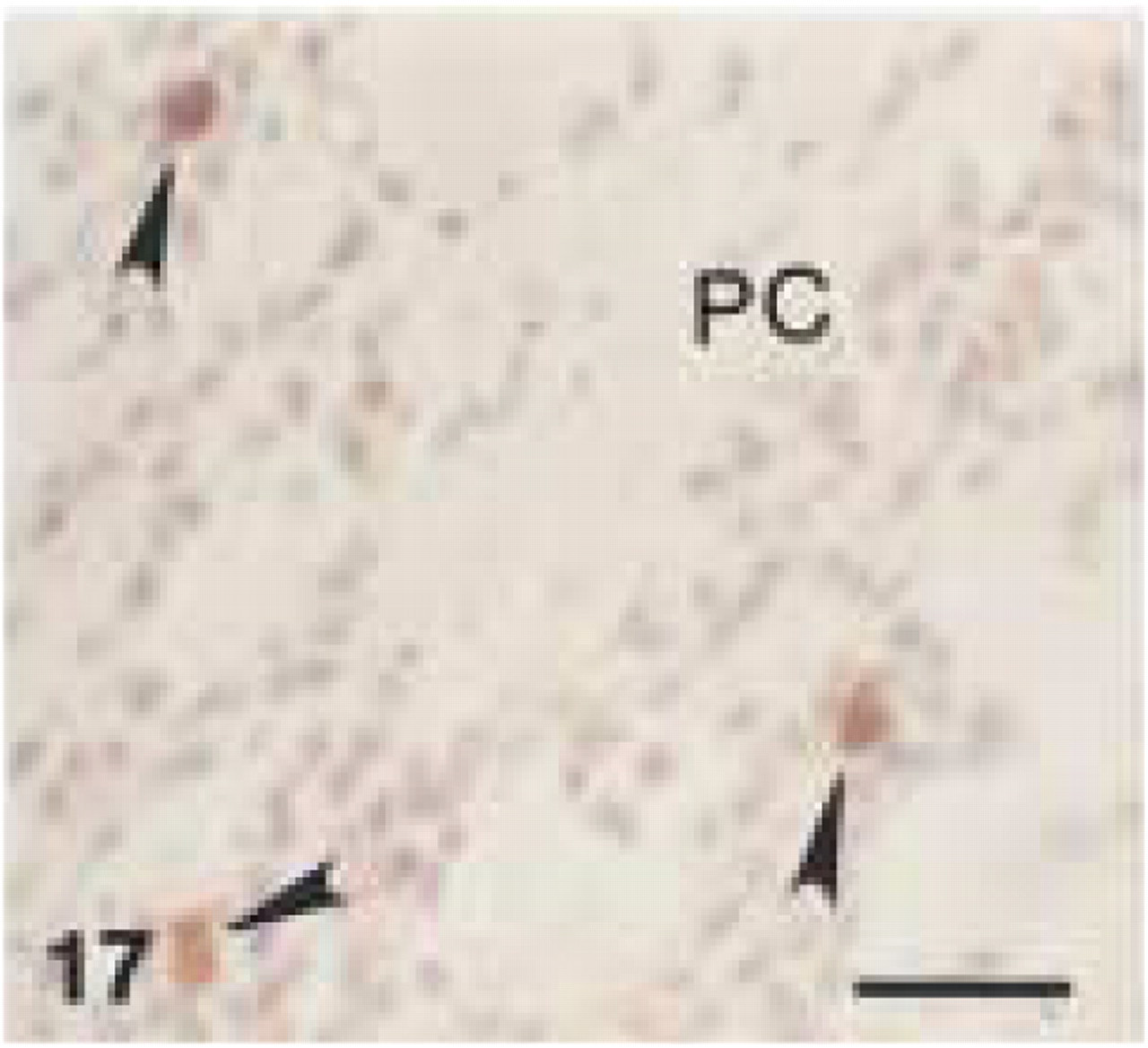
Monoclonal anti-human CD68 (clone EBM11). Chronic inflammatory infiltrate in central part of gingival lamina propria. CD68+ staining of macrophages (arrowheads) and weaker staining of plasma cells (PC). (EDTA-decalcified specimen.) Bar = 28 μm.
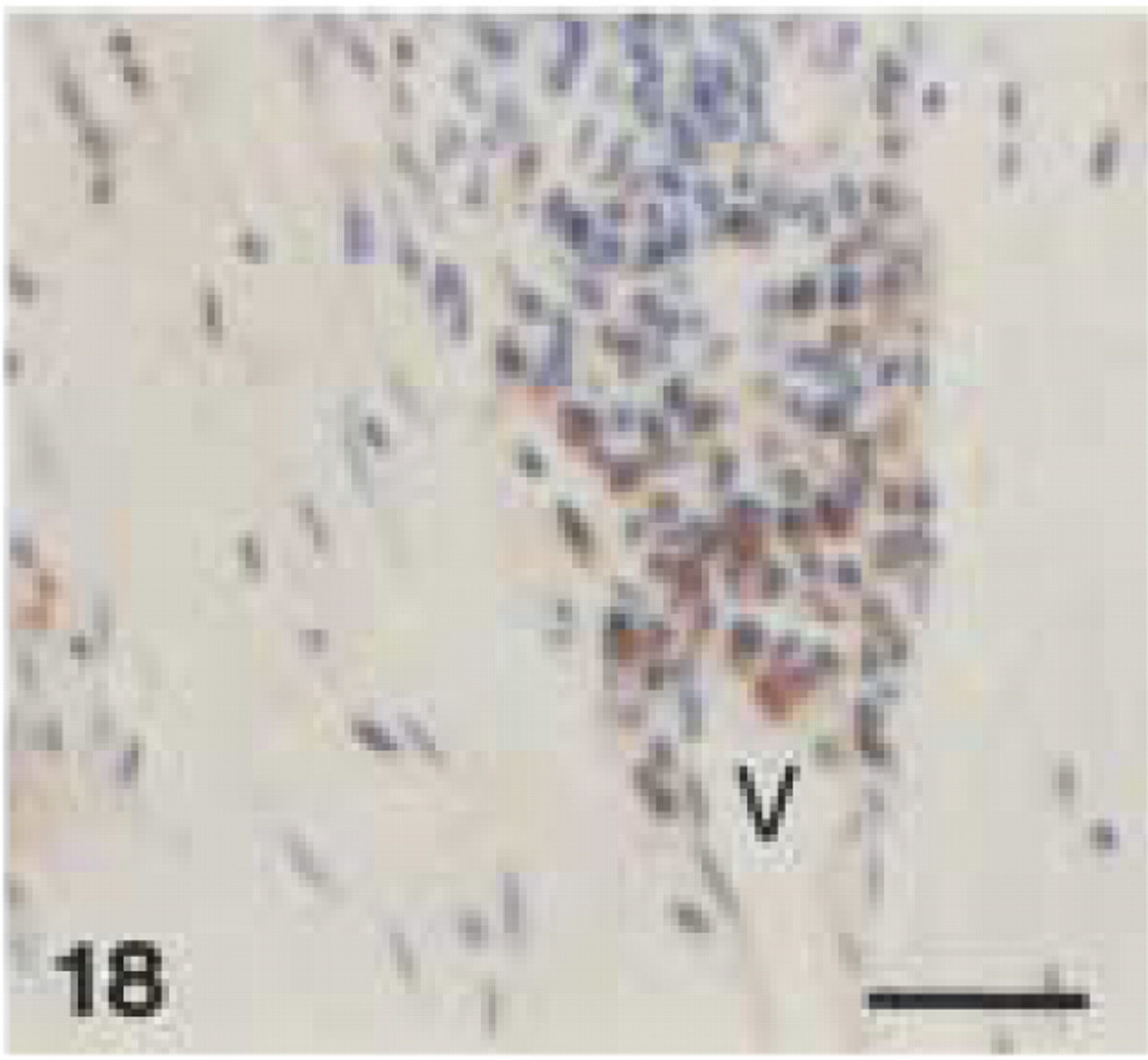
Monoclonal anti-human CD68 (clone EBM11). Perivascular inflammatory infiltrate in gingival subepithelial connective tissue. Note intense CD68+ cytoplasmic staining of plasma cells around blood vessel (V) (EDTA-decalcified specimen). Bar = 28 μm.
Monoclonal Mouse Anti-human CD68 Antibodies, Clones EBM11 and PG-M1 (Macrophage Marker)
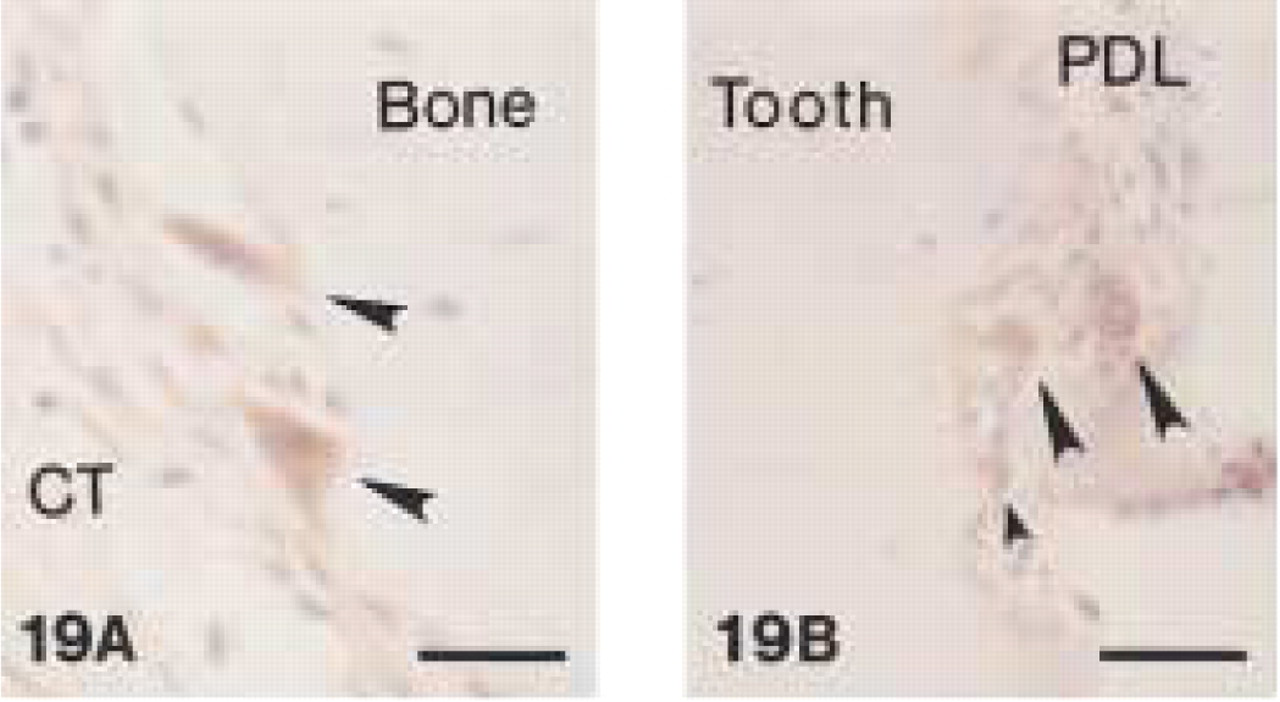
In canine gingival and periodontal tissues, both clones stained medium-sized and larger macrophages (Figures 12, 13, and 17) located in the central zone of the lamina propria of the gingiva and adjacent to the alveolar bone. Small macrophages and monocytes remained unstained. Interestingly, both anti-CD68 antibodies showed reproducible strong immunolabeling of plasma cells in canine tissues (Figures 14, 15, and 18). Anti-CD68 reacted more strongly and with more plasma cells than CD79α. Furthermore, canine multinucleated cells, e.g., either osteoclasts (Figure 19A) or cementoclasts (Figure 19B) in areas of bone and root resorption, showed weak staining with clone EBM11. This staining could be improved by increasing the antibody concentration from 1:50 to 1:10, although high unspecific background reactions were seen at this concentration. PG-M1 and EBM11 stained macrophages and plasma cells in medullar and paracortical regions of canine lymph nodes (Figures 16 and 20). Immunolabeling with both antibodies resulted in cytoplasmic staining that showed a foamy or granular pattern in macrophages and a more homogeneous staining in plasma cells.
Immunoreactivity of All Antibodies in Human Control Tissues
In human control tissues, anti-CD3 stained specifically mononuclear cells in the typical T-cell regions of a reactive lymph node and a tonsil (Figure 21), as well as variable numbers of lymphocytes in tissue sections obtained from a periapical granuloma and a radicular cyst.
Anti-CD79α antibodies (clones HM57 and JCB117) showed strong staining of lymphocytes in the follicular mantle zone of reactive lymph nodes. In the germinal centers, immunoblasts were weakly stained (Figure 22A). Plasma cells were intensively labeled (Figure 22B). In all human tissues, clone JCB117 reacted distinctly more strongly than clone HM57.
The anti-human myeloid/histiocyte antibody (clone MAC387) stained intravascular monocytes (Figure 23), small macrophages in the medullar region of a reactive lymph node (Figure 24) and a tonsil, as well as a periapical granuloma and radicular cyst. Neutrophils in a periodontal abscess showed strong immunoreactivity with MAC387.
Monoclonal anti-human CD68 (clone EBM11).
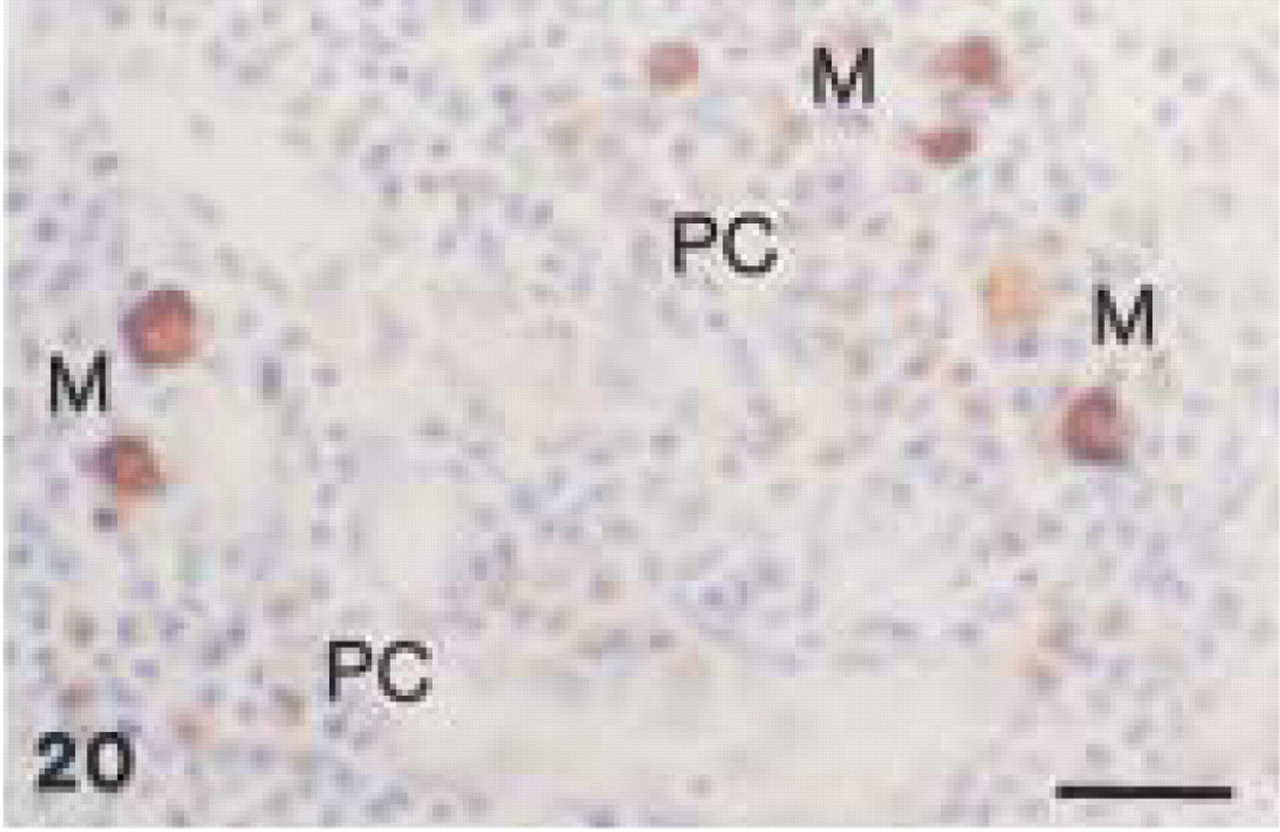
Monoclonal anti-human CD68 (clone EBM11). Canine lymph node: intense CD68+ staining of large macrophages (M) and weaker immunolabeling of multiple plasma cells (PC). (Similar staining pattern as with PG-M1 in Figure 16). Bar = 28 μm.
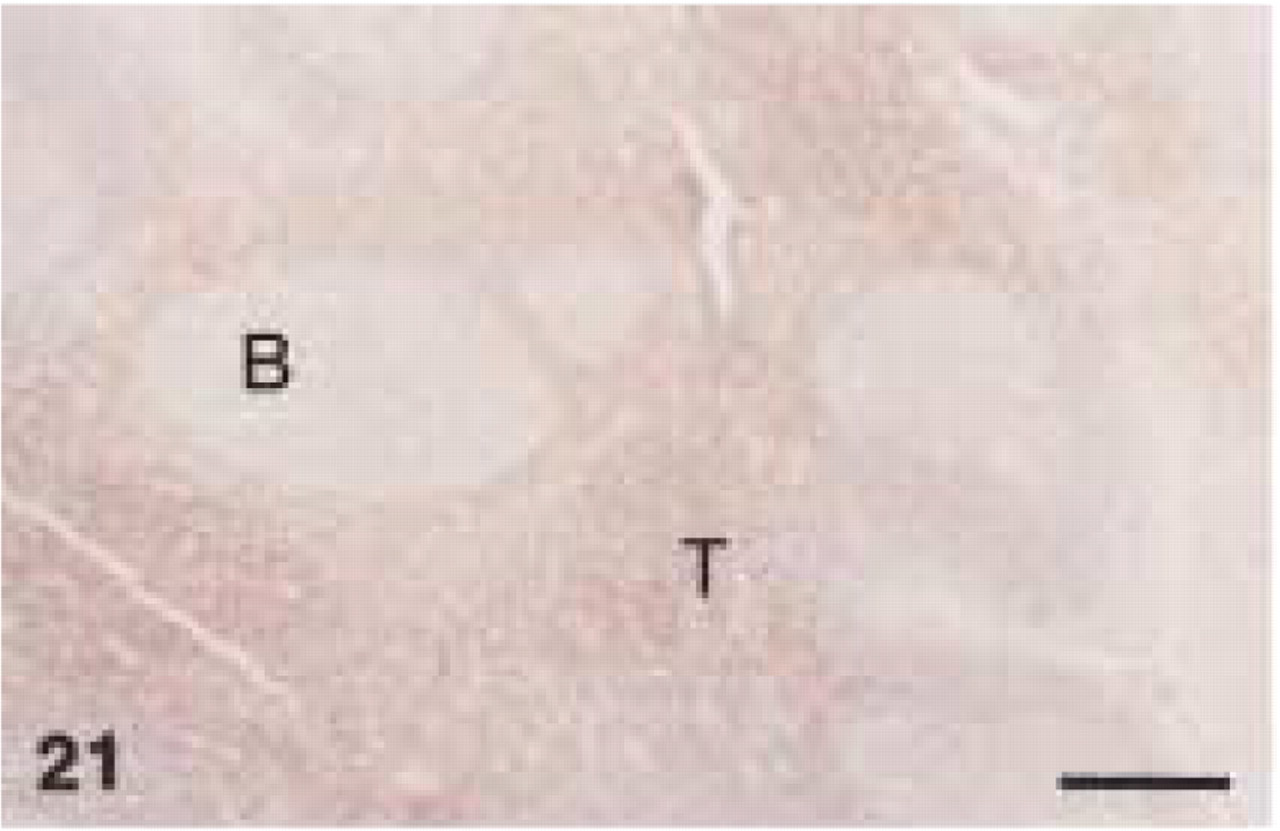
Polyclonal anti-human CD3. Human tonsil: note intense CD3+ staining of T-cells (T) in interfollicular area, whereas follicular B-Cells (B) are negative. Bar = 282 μm.
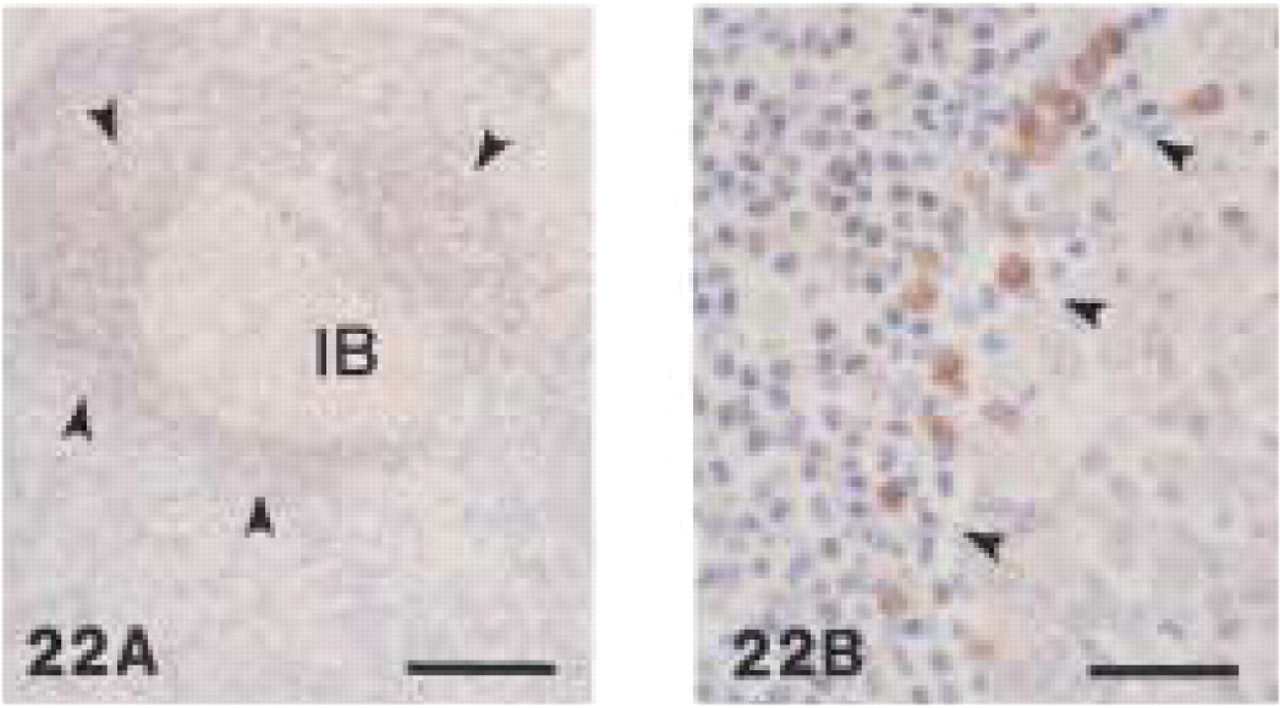
Monoclonal anti-human CD79α (clone HM57). Human reactive lymph node
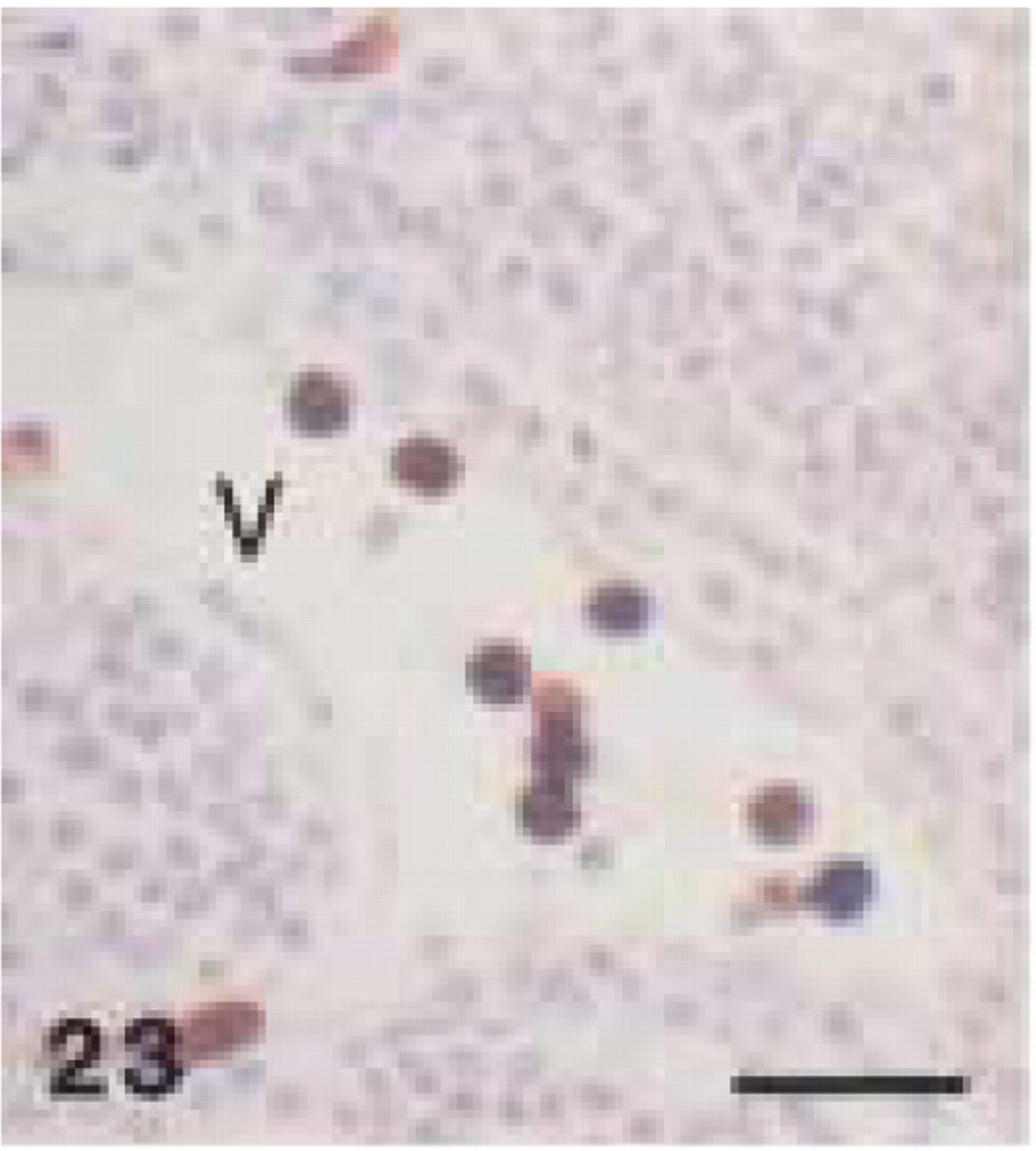
Monoclonal anti-human myeloid/histiocyte antibody (clone MAC 387). Human tonsil: intense immunolabeling of intravascular monocytes. V, blood vessel. Bar = 28 μm.
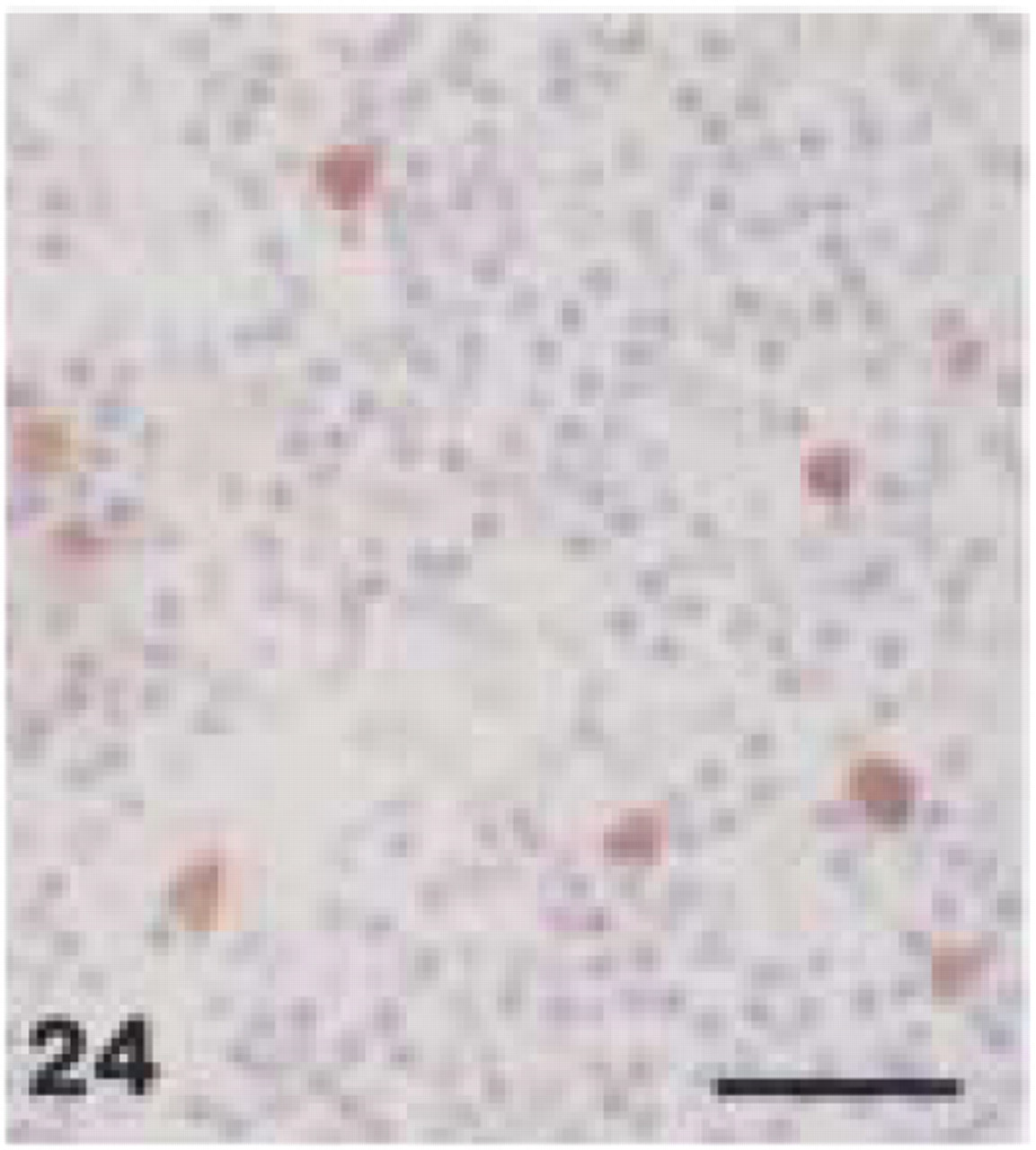
Monoclonal anti-human myeloid/histiocyte antibody (clone MAC 387). Human reactive lymph node: immunolabeling of small macrophages. Bar = 28 μm.
Monoclonal anti-human CD68 (clone PG-M1). Human periodontal abscess: strong immunolabeling of medium-sized and large macrophages. Note negative staining of neutrophils (arrowheads). Bar = 28 μm.
Monoclonal anti-human CD68 (clone PG-M1). Human ossifying fibroma: intense CD68+ staining of multinucleated osteoclasts in resorption lacunae. B, bone. Bar = 57 μm.
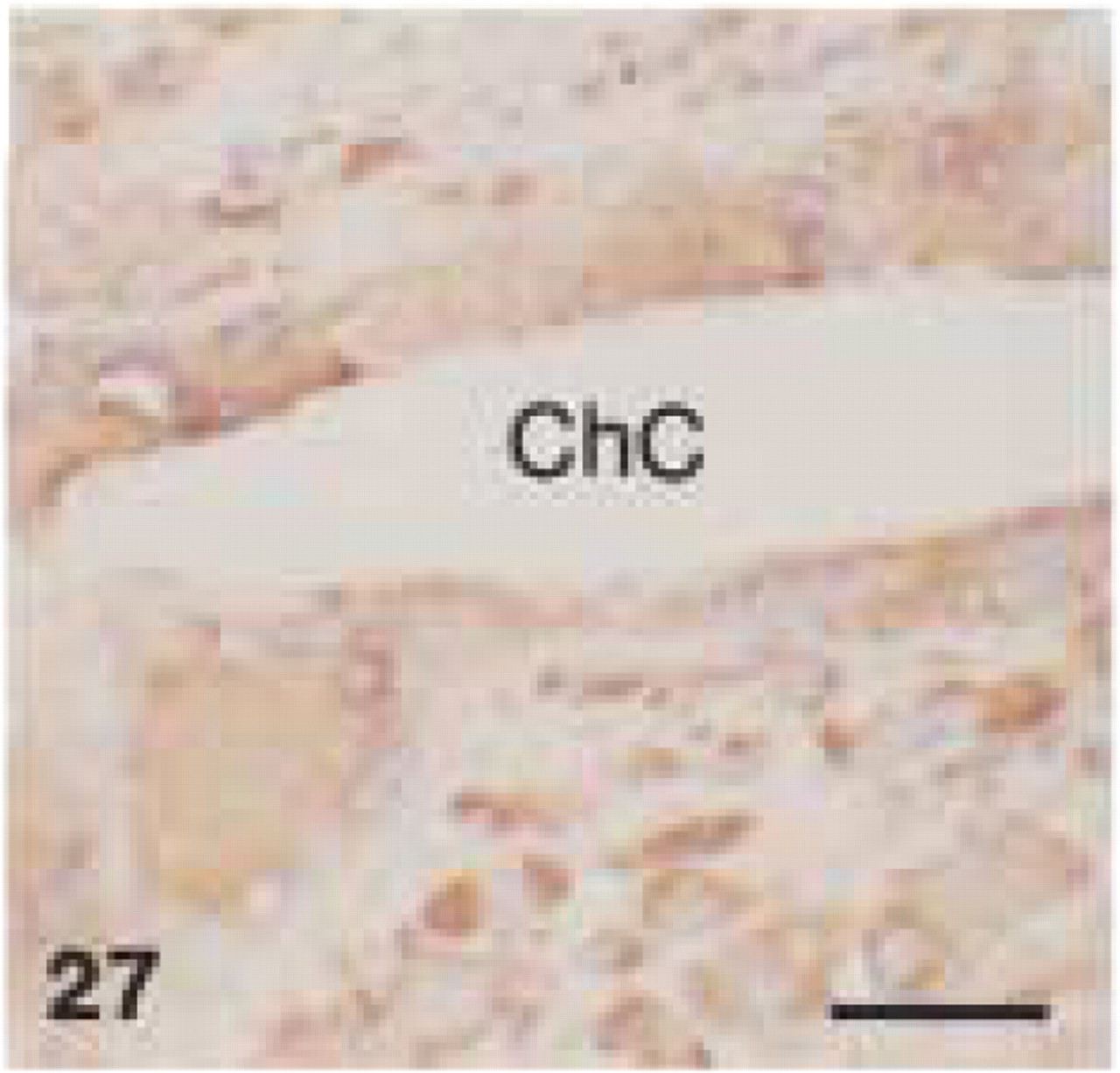
Monoclonal anti-human CD68 (clone PG-M1). Human radicular cyst: CD68+ staining of multinucleated giant cells along cholesterol crystals (ChC). Bar = 28 μm.
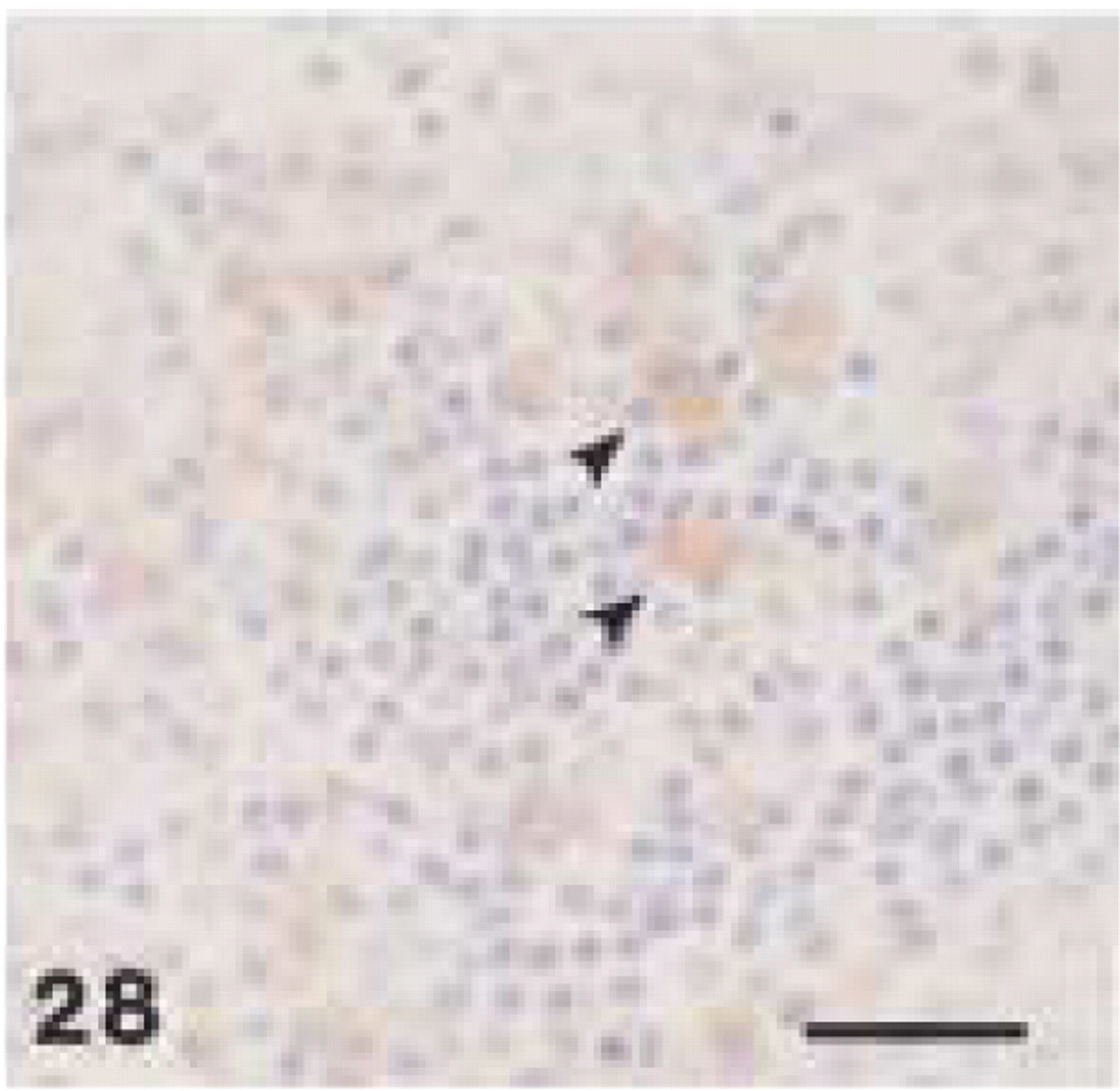
Monoclonal anti-human CD68 (clone EBM11). Human tonsil: CD68+ cytoplasmic staining of plasma cells (arrowheads). Note typical spoke-like appearance of nuclei. Bar = 28 μm.
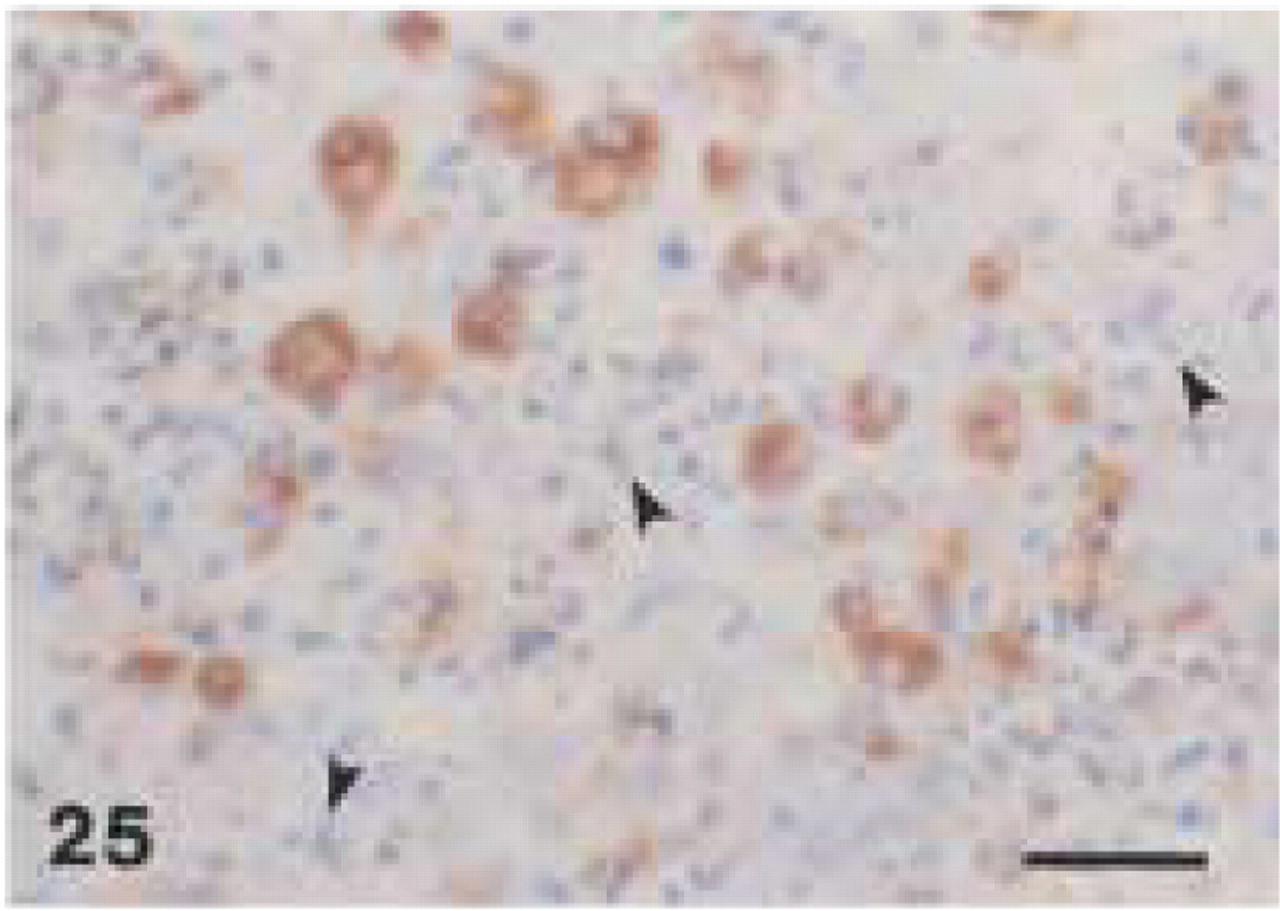
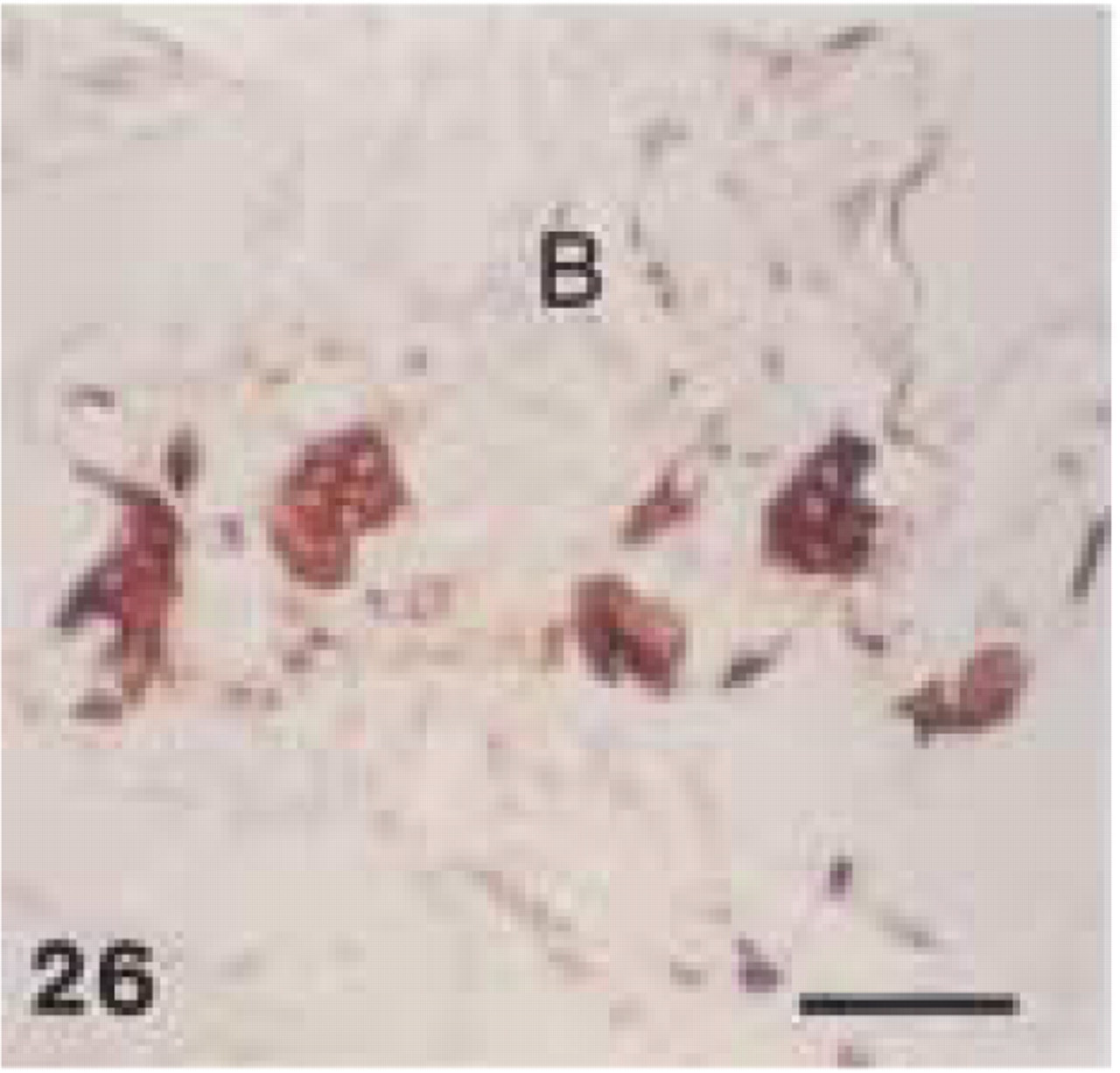
In human tissue, anti-CD68 (clone EBM11) showed distinctly weaker staining than anti-CD68 (clone PG-M1). The latter caused a foamy staining of multiple macrophages in the periodontal abscess (Figure 25) and in the medullary and paracortical region of a reactive lymph node, whereas only some large macrophages revealed granular staining with EBM11. In the periodontal abscess and in the periapical granuloma, cells, that were positively stained by MAC387 or PG-M1, did not react with EBM11. In human tissues, both anti-CD68 antibodies showed strong immunoreactivity with osteoclasts (Figure 26). Furthermore, EBM11 and PG-M1 reacted strongly with the cytoplasm of multinucleated giant cells that were found in the peripheral giant-cell granuloma and along the cholesterol crystals in the radicular cyst (Figure 27). In human tissues, only EBM11 clearly stained plasma cells, although the staining intensity was weaker than with the anti-CD79α antibodies (Figure 28).
Discussion
The beagle dog is one of the most widely used animal models for histological studies on the pathogenesis and therapy of periodontitis (Giannobile et al. 1994; Page and Schroeder 1982). However, there have been no cell-specific markers for the characterization of inflammatory cell infiltrates in canine periodontal tissues. Such baseline information is necessary for future studies aimed at evaluating molecular mechanisms underlying the progress and resolution of periodontal disease. In this study we were able to establish experimental conditions for the immunohistochemical identification of key immunocompetent cells in canine periodontal tissues using anti-human antibodies. Our results indicate that each antibody required its own individual pretreatment conditions for optimal immunoreactivity.
Macrophages play a central role in the pathogenesis and resolution of periodontitis. The release of proin-flammatory mediators induces and amplifies the inflammation, the specific immune response, and the ensuing tissue destruction (Dennison and Van Dyke 1997). On the other hand, macrophages are also an important source of key growth factors that appear to initiate periodontal wound healing (Van Dyke et al. 1993). Identification of this cell type is therefore crucial for the exploration of cellular and molecular mechanisms in disease and tissue repair. In the present study we were able to immunolabel macrophages in decalcified and nondecalcified canine periodontal tissues using two different anti-human-CD68 antibodies. Both clones, EBM11 and PG-M1, showed similar staining characteristics, although only EBM11 appeared to react with canine osteoclasts and cementoclasts. These cells, which are most probably formed by fusion of mononuclear cells of hemopoietic origin (Connor et al. 1995), play an essential role in the tissue destruction associated with periodontal disease. To the best of our knowledge, this is the first report of successful identification of canine macrophages using antihuman CD68 antibodies. Previous studies (Ackermann et al. 1994; Zeng et al. 1996) failed to find an immunoreactivity of clone EBM11 with macrophages in canine tissues obtained from lung, spleen, and lymph nodes. These differences might be explained by different experimental conditions and, in part, by different subsets of macrophages. Antibodies raised against CD68, a 110-kD glycoprotein found in the cytoplasm of human macrophages and monocytes, have been used by several laboratories as selective markers for this cell type (Kelly et al. 1988; Greywoode et al. 1990; Falini et al. 1993). In the present study, interestingly, both anti-CD68 antibodies also reproducibly stained canine plasma cells. In our human tissues, this cell type appeared to react only with clone EBM11. These findings indicate either that the antibodies used are not completely specific for CD68 or that the CD68 antigen is also present in plasma cells. Pulford et al. (1990) already suspected CD68 to be a more heterogeneous entity when they found CD68+ staining in large lymphocytes. However, until a more specific antibody becomes available, canine macrophages can be easily distinguished from plasma cells by their characteristic morphology and staining pattern.
Anti-CD68 selectively immunolabeled larger, i.e., resident, macrophages. Canine monocytes and small macrophages remained unstained by anti-CD68. We were able to stain these cells by applying an anti-myeloid/histiocyte antibody (MAC 387) raised against calgranulins (Flavell et al. 1987; Brandtzaeg et al. 1988; Chilosi et al. 1990). These are two calcium binding myeloid-associated proteins, which are related to migration inhibitory factor (MIF) and are lost within a few days from monocytes differentiated in vitro. Consistent with previous findings in human (Flavell et al. 1987; Brandtzaeg et al. 1988; Chilosi et al. 1990) and canine tissues (Pérez et al. 1997), this antibody stained monocytes, neutrophils, and keratinocytes in our canine periodontal tissues. Large macrophages were not stained.
Because they stain different subsets of the mononuclear phagocyte lineage, anti-CD68 and anti-MAC387 antibodies are excellent markers to distinguish between acute and chronic episodes in the progress of periodontal disease. Anti-CD68 labels large, probably fully differentiated macrophages, osteoclasts, and cementoclasts, which are mostly found in the chronically inflamed periodontium. In contrast, anti-MAC387 reacts with emigrating monocytes and young macrophages, indicating a more acute lesion with a high cell turnover (Chilosi et al. 1990; Poston and Hussain 1993).
As the two major cell types of the specific immune response, T- and B-lymphocytes play distinctive roles in the progress of chronic periodontal disease. Although controversial, there are indications that T-cells are the predominant lymphocytes in the stable lesion, whereas increased proportions of B-cells and plasma cells are found in the progressive lesion (Seymour, 1991; Gemmell and Seymour 1998). In the present study, canine periodontal T-lymphocytes were specifically immunolabeled with a polyclonal antibody raised against a highly conserved intracellular peptide sequence of the human CD3 antigen (Mason et al. 1989; Jones et al. 1993). These results are consistent with previous findings (Ferrer et al. 1992,1993; Day 1996, 1997; Pérez et al. 1996,1997) in which the use of this antibody was documented for the successful detection of canine T-lymphocytes in different inflamed extraoral soft tissue specimens. In our study, CD3+ staining was mainly found in the connective tissue subjacent to the sulcus epithelium of the inflamed beagle periodontium. This predominant localization is in accordance with previous immunohistological findings in human gingival tissues (Celenligil et al. 1990).
There has been no report in the literature describing specific markers for canine B-lymphocytes and plasma cells, except using polyclonal antibodies directed against immunoglobulins (Jergens et al. 1996; Day 1997; Pérez et al. 1997). In this study we were able to identify both cell types in beagle tissues by specifically applying a monoclonal antibody (clone HM57) raised against the intracellular epitope of the human CD79α antigen. In humans, this protein appears early in B-cell maturation and persists until the plasma cell stage (Mason et al. 1991; Astsaturov et al. 1996). The specifity of clone HM57 for canine B-cells was confirmed by immunolabeling of follicular lymphocytes and absence of staining in T-cell-enriched zones of canine lymph nodes. This staining pattern was similar to that found in human control tissue in the present and previous studies (Mason et al. 1991; Astsaturov et al. 1996; Chuang and Li 1997). Clone HM57 clearly reacted with plasma cells in nondecalcified canine gingival tissues. Interestingly, in decalcified tissue specimens this antibody labeled only a proportion of cells, which were recognized as plasma cells by their H&E morphology. It is feasible to speculate that long-term decalcification or species-related differences among various plasma cell subsets associated with post-translational modifications contributed to differences in CD79α staining patterns. Although B-cells reacted consistently with anti-CD79α (clone HM57), plasma cell staining was more intense and more complete with anti-CD68 (clones EBM11 and PG-M1) in canine periodontal tissues. Similar to previous findings in humans (Celenligil et al. 1990), canine gingival plasma cells were found predominantly in the central part of the lamina propria. The second anti-CD79α antibody used in this study, clone JCB117, showed no immunoreactivity in canine tissues. This clone is raised against an extracellular epitope of the human CD79α antigen, which appears to be different or not accessible in canines.
In conclusion, this study reported the use of five different antibodies (CD3, HM57, MAC387, PG-M1, EBM11) for specific immunohistochemical identification of T- and B-lymphocytes, plasma cells, macrophages in different differentiation stages, osteoclasts, and cementoclasts in inflamed canine periodontal tissues. These antibodies, all of which were raised against intracytoplasmic epitopes, showed similar staining patterns in canine and human immunocompetent cells. They can be used for characterizing the nature of the inflammatory cell infiltrate in future studies evaluating the molecular mechanisms underlying the progress and resolution of periodontal disease. Furthermore, these findings might be also of interest to other fields of medical research that use a canine animal model and for diagnostic purposes in veterinary pathology.
Footnotes
Acknowledgments
Supported in part by the DFG grant Ch 192/1-1 from the Deutsche Forschungsgemeinschaft (DFG).
We are grateful to Dr Peggy A. O'Neill (Department of Stomatology, Divison of Periodontics) and Dr Joel S. Gaikwad (Department of Basis Sciences) for helpful comments. V.M. Hubbard and G. Rittman are gratefully acknowledged for their help in tissue processing. Antibodies were provided in part by Dako (Carpinteria, CA).