
Abstract
R
HNF-3β and/or TTF-1 regulate genes critical to perinatal surfactant homeostasis and host defense, including surfactant proteins A, B, and C and CCSP (Sawaya et al. 1993; Bohinski et al. 1994; Bruno et al. 1995; Kelly et al. 1996). Surfactant homeostasis and respiratory cell differentiation are incompletely developed in premature infants with hyaline membrane disease (HMD). HMD is often followed by an abnormal repair process termed bronchopulmonary dysplasia (BPD), a common cause of morbidity and mortality in premature infants. At present, knowledge of the temporal-spatial distribution of HNF-3β is confined to the mouse, and there is no information regarding its presence in human tissues. The present study was designed to localize HNF-3β as a marker for progenitor cells in developing human foregut derivatives and to correlate its expression with that of other respiratory epithelial cell markers during lung development and after injury. The localization of HNF-3β was compared with immunohistochemical data, a part of which was generated previously for the TTF-1 peptide, surfactant proteins, and CCSP in fetuses and newborn infants (Khoor et al. 1993,1994,1996; Stahlman et al. 1996).
Materials and Methods
This study was approved by the Committee for the Protection of Human Subjects, Health Sciences of Vanderbilt University Review Board. Lung tissue was available from 31 fetuses between 10 and 40 weeks of gestation, from six infants dying with HMD between 2.5 and 10 postnatal days, from 14 infants of 12-35 days after HMD (early BPD), of which two were lobectomy specimens, and from four infants between 5 and 7 months of postnatal age with late BPD. Tissues were obtained with parental consent between 1979 and 1991 at hysterotomy abortion, hysterectomy abortion, postmortem biopsy, or autopsy. Tracheas were available from 13 fetuses between 10 and 23 weeks of gestation, four for immunostaining also with TTF-1. Esophagi were also available from eight of the same fetuses. A limited number of other fetal and neonatal tissues were also available for study including eight pancreases, three stomachs, four duodenums, two gallbladders, four livers, two testes and epididymes, one skin with hair follicles and sebaceous glands, four large intestines, four kidneys, one adrenal, eight thyroids and two umbilical cords. Tissue from some older subjects was used for comparison, including four lungs, three pancreases, one adrenal, one submandibular gland, one liver, one brain, two esophagi, two stomachs, two large intestines, two kidneys, one thyroid, two vaginas, and one skin, including sweat and sebaceous glands. Fetal tissue was obtained within 30 min and autopsy specimens were usually obtained within 2 hr after death. Tissues were fixed in 10% phosphate-buffered formalin, dehydrated through ethanols, and embedded in paraffin. Four μm-thick sections were cut and mounted on Superfrost Plus (Fisher; Norcross, GA) glass slides.
Immunohistochemistry
The specificity of the HNF-3β antiserum has been described previously (Jacob et al. 1994; Ikeda et al. 1995; Zhou et al. 1996). Briefly, a rabbit polyclonal antibody (kindly provided by Dr. Robert Costa, University of Chicago, Chicago, IL) was generated against recombinant rat HNF-3β and the antibody was purified by affinity chromatography. Tissues were processed, sectioned, deparaffinized, quenched in 0.3% H2O2 in methanol, and rehydrated as described previously (Stahlman et al. 1992). Immunostaining of HNF-3β was enhanced by antigen retrieval. Slides were subjected to a 25-min microwave treatment in a 1 × Antigen Retrieval Citra buffer (BioGenex Laboratories; San Ramon, CA). After removal from the microwave, slides were allowed to remain in the 1 × Antigen Retrieval Citra buffer for an additional 20 min, after which they were rinsed and placed in PBS. Nonspecific staining was blocked by exposing slides for 30 min to prediluted normal goat serum (BioGenex Laboratories). The appropriately diluted rabbit anti-HNF-3β serum was then applied and incubated overnight at 4C. After washing, a biotinylated goat anti-rabbit immunoglobulin reagent was applied to the slides for 30 min (BioGenex SS Kit). After washing, a peroxidase-conjugated streptavidin was added and incubated for 30 min. Peroxidase activity was localized by reaction in a solution containing 95 mg DAB, 1.6 g NaCl, 2 g NiSO4 in a total of 200 ml 0.1 M acetate buffer, pH 6.0. Immediately before use, 25 ml of 30% H2O2 was added. Specific staining was enhanced by a 4-min incubation in 0.05 M Tris-0.5% cobalt chloride, pH 7.2. Slides were then rinsed, counterstained with Nuclear Fast Red, and dehydrated. TTF-1, surfactant protein A, surfactant protein B and C precursors, and CCSP immunohistochemistry and their ablation were carried out as described previously (Stahlman et al. 1992,1996; Khoor et al. 1994,1996).
Normal rabbit serum was substituted for the primary antiserum as a negative control. An anti-human cytokeratin antibody to CAM 5.2 (Becton-Dickinson; San Jose, CA) was used as a ubiquitous protein to demonstrate that the distribution of antigen was not an artifact of antibody accessibility. HNF-3β labeling was ablated by substituting HNF-3β antibody preabsorbed with affinity purified GST- HNF-3β (aa 7-86 fusion protein) for the primary antibody.
The surface of each section was examined microscopically by two of the authors independently, and the relative number and distribution of labeled cell nuclei were agreed on.
Results
Localization of HNF-3β and TTF-1 in Fetal Human Respiratory Tract
HNF-3β and TTF-1 immunolabeling was most evident in the nuclei of nonciliated tracheal epithelial cells from the earliest fetuses. Expression of both TTF-1 and HNF-3β decreased in tracheas with increasing gestational age, and no reactivity was detected for either protein after 19 weeks of gestation. In two fetuses of 18 and 19 weeks of gestation, labeling was confined to a few basal cells. The temporal-spatial distribution of nuclear immunolabeling for HNF-3β and TTF-1 was similar in intrapulmonary bronchi to that seen in the tracheas. Nuclear staining for both proteins was intense in cells lining conducting airways in the younger fetuses and in terminal airways (Figures 1 and Figures 2). In intrapulmonary bronchi, only basal cells were stained for either antibody after 19 weeks. Nuclei of columnar nonciliated cells were the predominantly immunolabeled cell type in bronchioles of the HNF-3β in Fetal Lung and Foregut Derivatives youngest fetuses. After 19 weeks of gestation, immunolabeling in large bronchioles was restricted to a few basal cells. Nuclear labeling of cuboidal cells in bronchioles and terminal airways was similar at all gestational ages (Figure 3).
In general, the intensity of HNF-3β and TTF-1 staining of terminal airways and cuboidal bronchiolar epithelium, especially those at the periphery of lobules and abutting septa, was more pronounced than in more proximal airways, especially in older fetuses. By 36 weeks of gestation, minimal labeling for HNF-3β and TTF-1 was observed in the conducting airways at all levels, whereas nuclei of Type II cells in terminal airways remained well stained (Figure 4). In lungs from older subjects, HNF-3β was detected only in some cells lining bronchiolo-alveolar portals and in Type II cells. Immunolabeling of the lung with TTF-1 closely matched that of HNF-3β, the distribution of labeled nuclei being remarkably similar at all stages of gestation (Stahlman et al. 1996).
Localization of HNF-3β and TTF-1 in Regenerating Epithelium in HMD
We did not examine tissues from infants dying with acute HMD within the first 48 hr after birth, because the extent of the pathology made interpretation impossible. However, six infants with HMD of 2.5 to 10 postnatal days were examined. There were no intact tracheas. In sections of intrapulmonary bronchi from two of these postnatal infants, immunolabeled nuclei were detected with both antibodies. As in the fetuses, there was an increasing gradient of the numbers of immunolabeled nuclei from large bronchioles to cuboidal epithelium-lined bronchioles to terminal airways. Nuclei of terminal airways with regenerated/regenerating epithelium were immunolabeled, but not always intensely. Immunolabeling was generally most intense in parenchyma near pleura and septa, as well as near connective tissue surrounding large airways or blood vessels. In areas with extensive pathology, including hemorrhage, infection, edema, or residual hyaline membranes as in acute HMD, staining for HNF-3β and TTF-1 was absent or difficult to interpret.
Localization of HNF-3β and TTF-1 in Respiratory Epithelium in BPD
Lung tissue was available from 13 infants between 12 and 35 days after the onset of HMD, two of which consisted of lobectomy specimens obtained at 4 weeks of age for lobar emphysema. None of the tracheas was examined because of extensive cell damage and/or squamous metaplasia. Bronchi of two infants had rare nuclei immunolabeled with both antibodies. Large bronchioles of two infants were denuded, and squamous metaplasia was noted in three. Tissues from six of the seven remaining infants demonstrated nuclear labeling for TTF-1 and HNF-3β in cuboidal bronchioles, and all had some immunolabeling of nuclei in terminal airways, often dysplastic, that was occasionally widespread, especially near pleura and septa (Figures 5 and Figures 6). In the two specimens obtained at lobectomy, widespread nuclear staining was absent with both antibodies at all levels of conducting airways but was readily detected in nuclei of terminal airways.
Tissue was available from four infants with BPD obtained between 5 and 7 months of postnatal age. Lungs from these infants were scarred extensively and the airways were lined by dysplastic cells. Infection and widespread interstitial collagen deposition were noted in all samples. Only rare basal cells labeled with TTF-1 and HNF-3β antisera were seen in large bronchioles in these infants. Some labeled cells were noted in cuboidal bronchioles and in terminal airways lined with dysplastic Type II cells. In the lungs of sick infants the prevalence and distribution of nuclear labeling for TTF-1 closely matched those for HNF-3β.
Substitution of preabsorbed antibody for the primary antibody resulted in ablation of the label for HNF-3β in fetuses and BPD infants (Figures 7a and 7b).
Localization of HNF-3β and TTF-1 in Nonpulmonary Organs and in Older Subjects
Although immunolabeling was never widespread in the esophagus, TTF-1 and HNF-3β were detected in basally located nonciliated, nonsquamous cells. By 20 weeks of gestation most of the ciliated cells had disappeared from the esophagus as it became lined by stratified squamous epithelium and, at this time, TTF-1 and/or HNF-3β immunolabeling had disappeared. Most nuclei of pancreatic acini from fetuses of 12, 16, and 18 weeks of gestation labeled for HNF-3β. Islet cell nuclei were not labeled for HNF-3β. In these same fetuses, nuclei of many cells of the crypts of Lieberkühn of the duodenum were immunolabeled, with a few scattered labeled nuclei being detected in the epithelium of villi (Figure 8). In the 12-week and the 16-week fetuses, many nuclei of the epithelium of the stomach labeled for HNF-3β. Gallbladder epithelium was labeled in two fetuses. Other fetal and neonatal organs examined lacked detectable HNF-3β. In lungs of older children and adults, HNF-3β was detected only in nuclei of some Type II cells and was not seen in organs other than lung. None of these latter tissues immunolabeled for TTF-1, with the exception of the thyroid.
Immunolabeling of HNF-3β, TTF-1, SP-A, proSP-B, proSP-C, and CCSP
Trachea. Six serial or near serial sections were immunostained for HNF-3β, TTF-1, SP-A, proSP-B, proSP-C, and CCSP in seven fetuses of 10 to 20 weeks' gestation and in four infants with BPD of approximately 1 month of age. In the 10-week fetus, in one of the 12-week fetuses, and the 15-week fetus in whom tracheas were available, some epithelial cells, thought to be nonciliated, and some basal cells were immunolabeled with these six antisera (Figures 9–12). Cells containing CCSP were found more often flanking folds or putative necks of tracheal glands. In the youngest fetus, cells immunolabeling for SP-A were also localized to folds. Immunolabeling of tracheal epithelium from the younger fetuses for HNF-3β and TTF-1 was more widespread than in older fetuses. Immunolabeling of tracheal lining cells with each of these antibodies diminished progressively by 20 weeks of gestation as epithelial cell differentiation progressed.
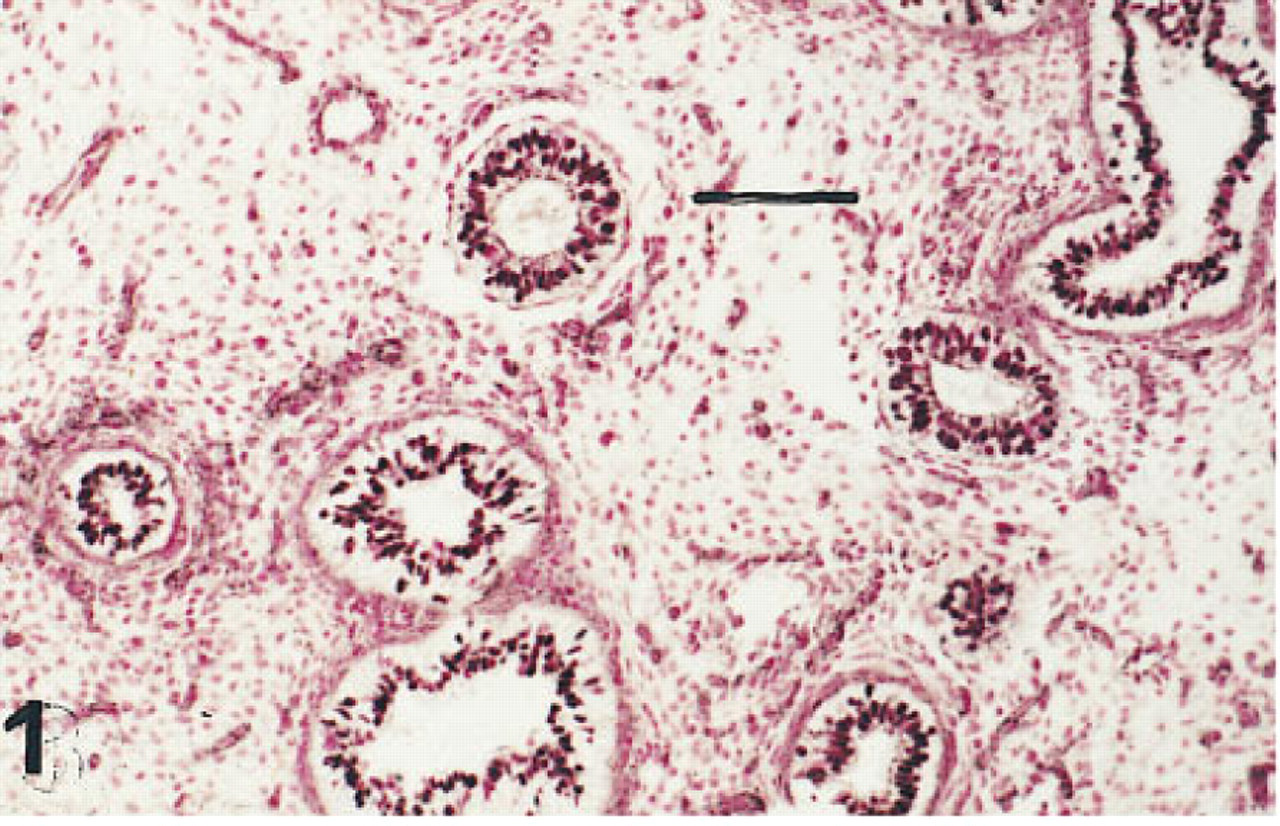
Lung of a fetus of 10 weeks' gestation immunostained for HNF-3β. The nuclei of a majority of the cells lining conducting and terminal airways are immunolabeled. Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 80 μm.
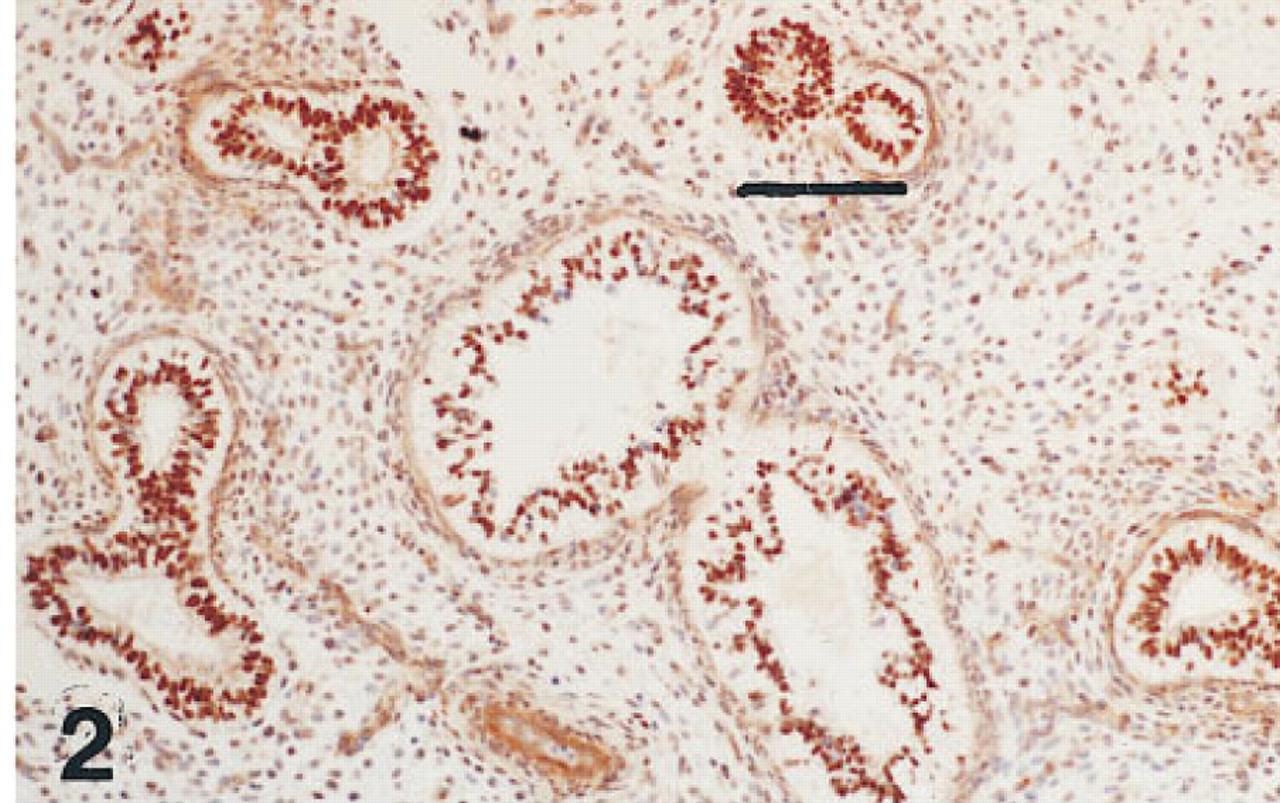
Lung of the same fetus as shown in Figure 1 immunostained for TTF-1. Nuclei in cells of the same generations of airways as those immunolabeled for HNF-3β are also immunolabeled for TTF-1. Peroxidase-conjugated streptavidin and hematoxylin. Bar = 80 μm.
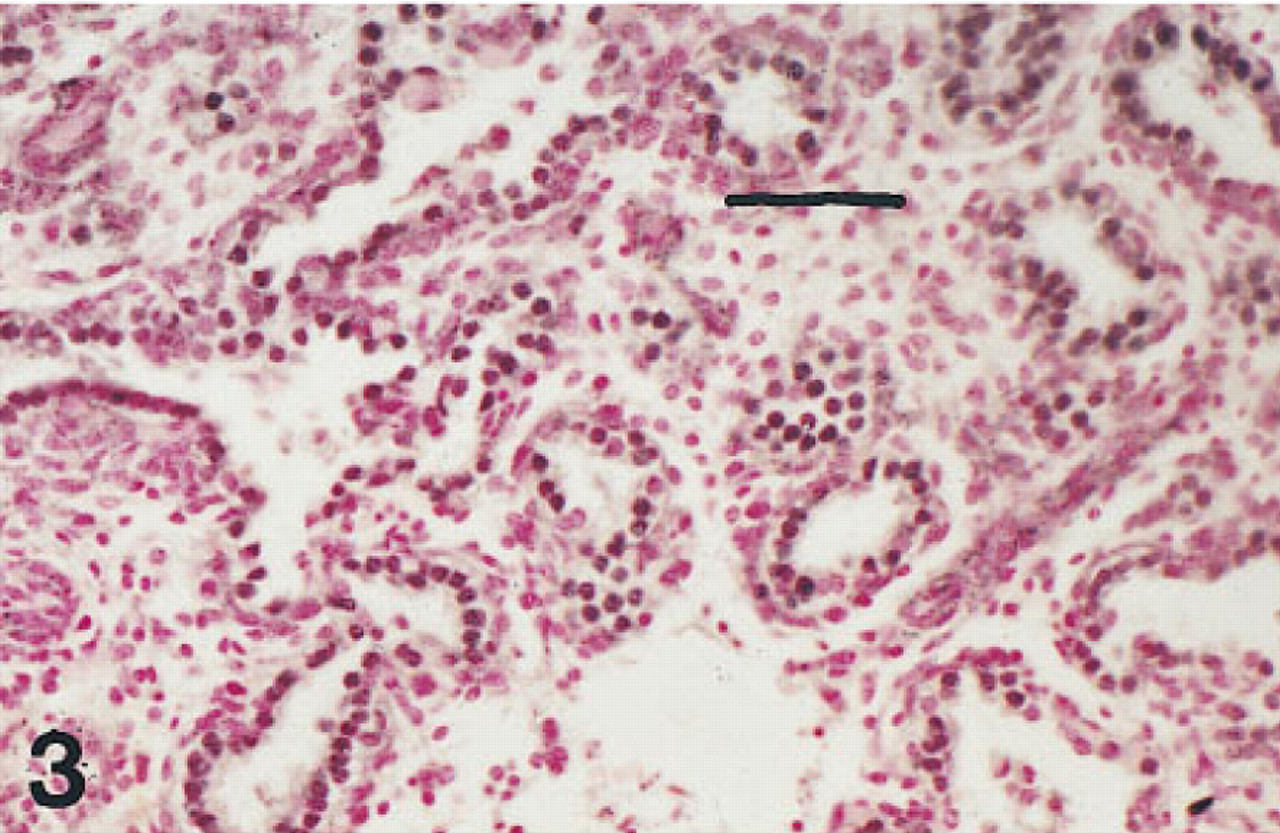
Lung of a fetus of 20 weeks' gestation immunostained for HNF-3β. The nuclei of cells lining airways are immunolabeled. Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 50 μm.
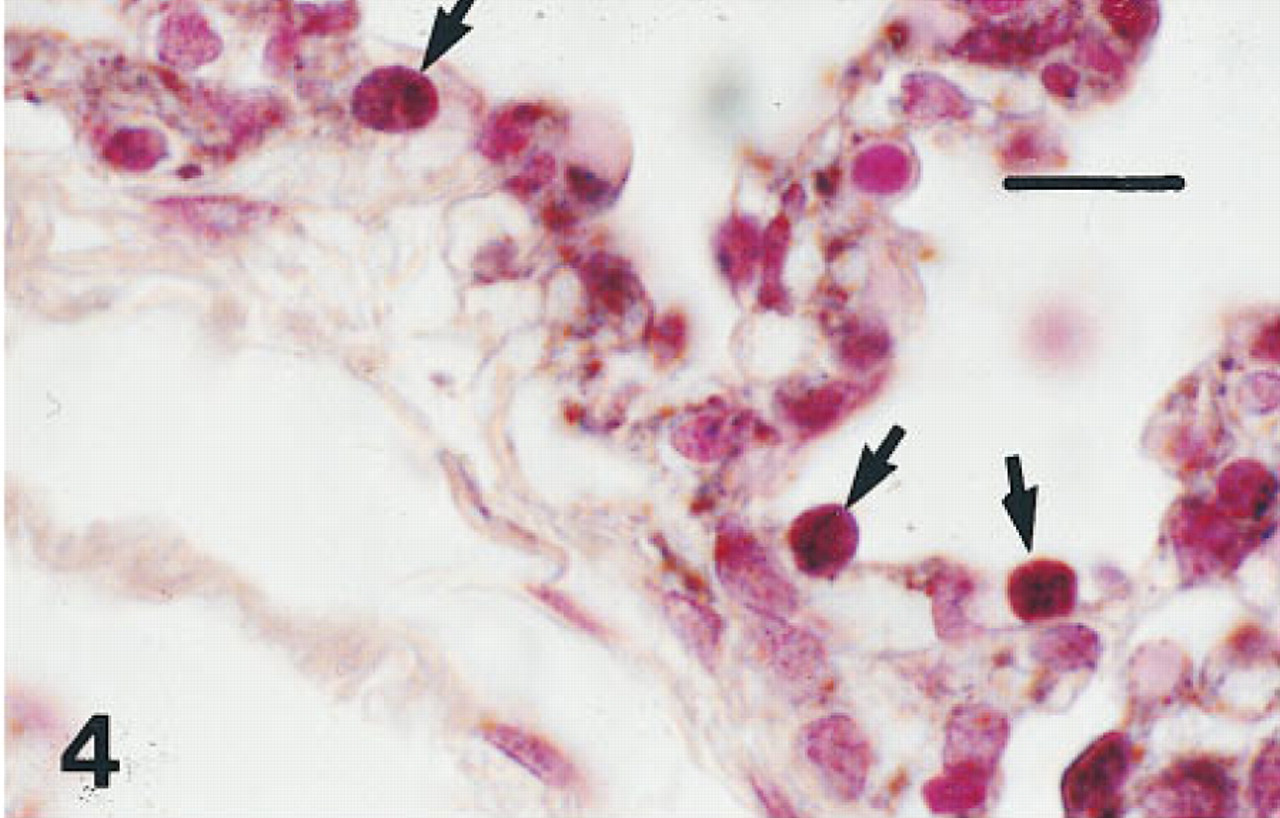
Lung of a fetus of 36 weeks' gestation immunostained for HNF-34bT. Nuclei of Type II cells (arrows) are immunolabeled. Peroxidaseconjugated streptavidin and nuclear fast red. Bar = 12 μm.
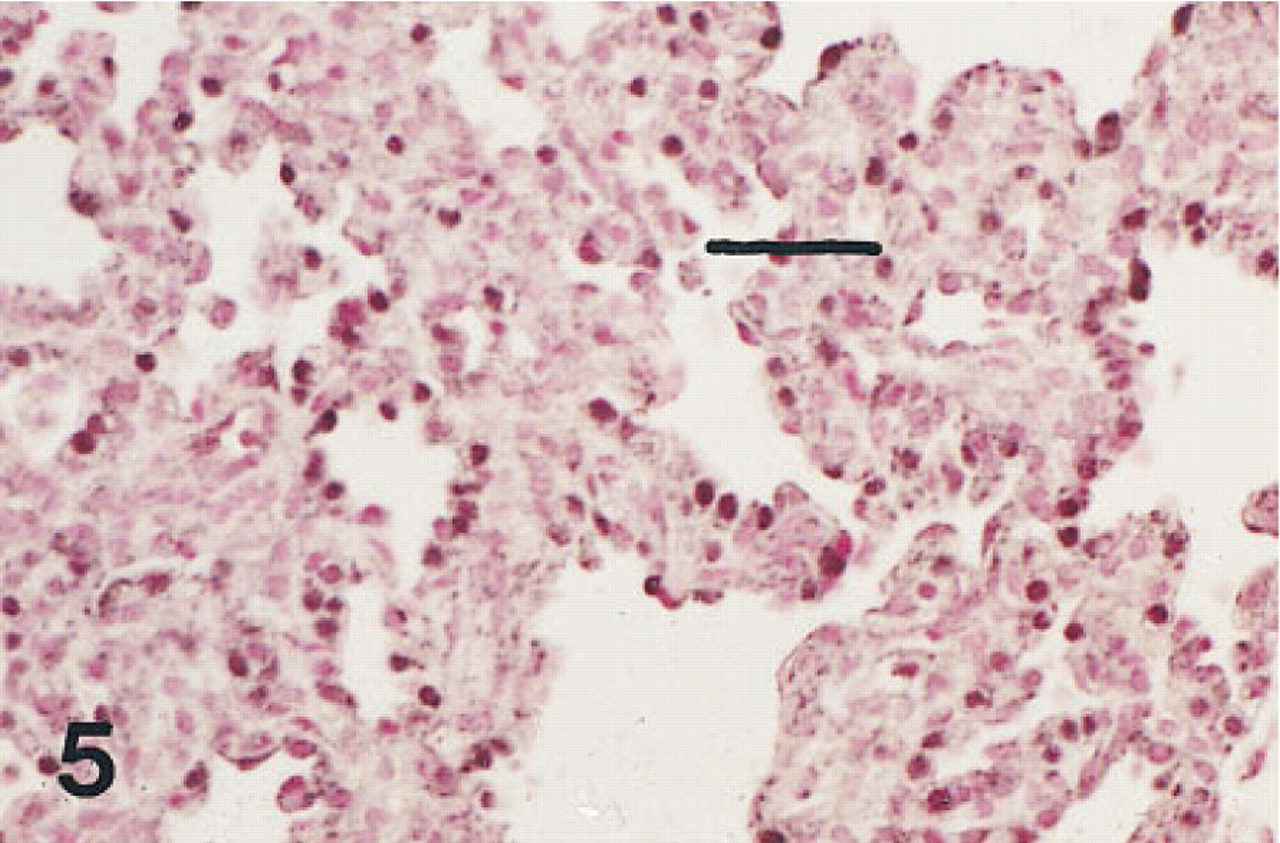
Lung of an infant of 28 weeks' gestation who survived 15 days with developing BPD immunostained for HNF-3β. The nuclei of many of the cells lining the thickened alveolar walls are immunolabeled. Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 30 μm.
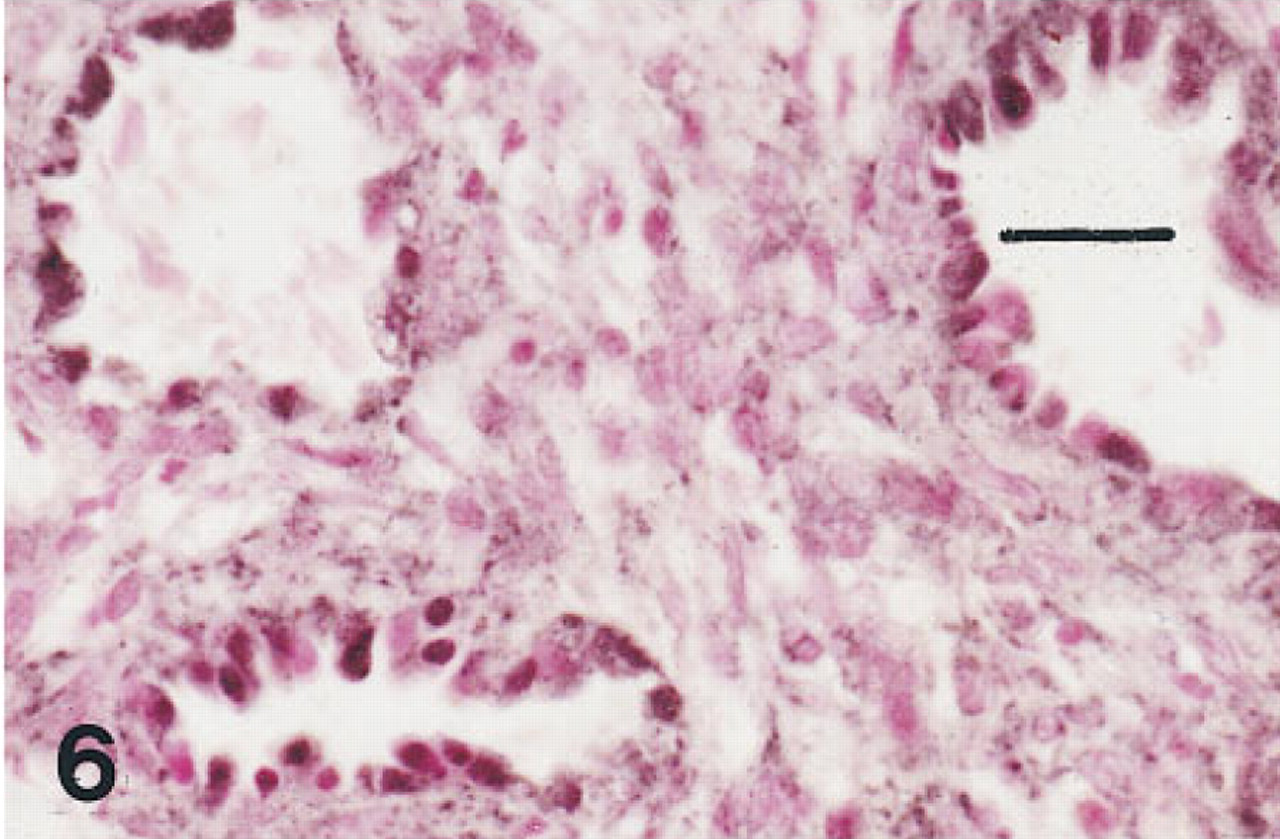
Lung of an infant of 28 weeks' gestation who survived for 30 days with BPD immunostained for HNF-3β. Nuclei of many tall dysplastic cells lining open airways are immunolabeled. Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 30 μm.
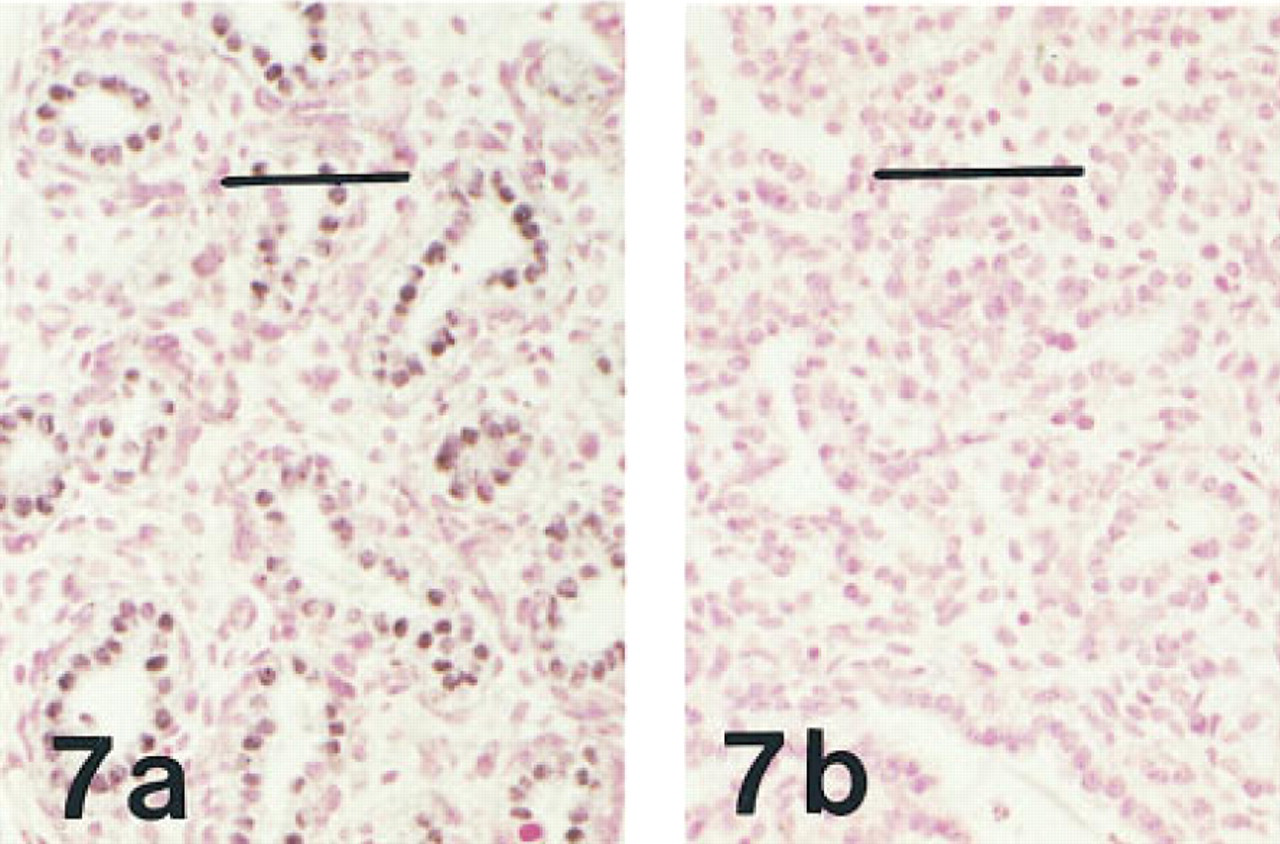
(a) Lung of a fetus of 20 weeks' gestation immunostained for HNF-3β, showing widespread immunolabeling of nuclei in terminal airways. Peroxidase-conjugated streptavidin and nuclear fast red. (b) Lung of the same fetus showing ablation of nuclear labeling when preabsorbed antibody is substituted for the primary antibody. Bar = 50 μm.
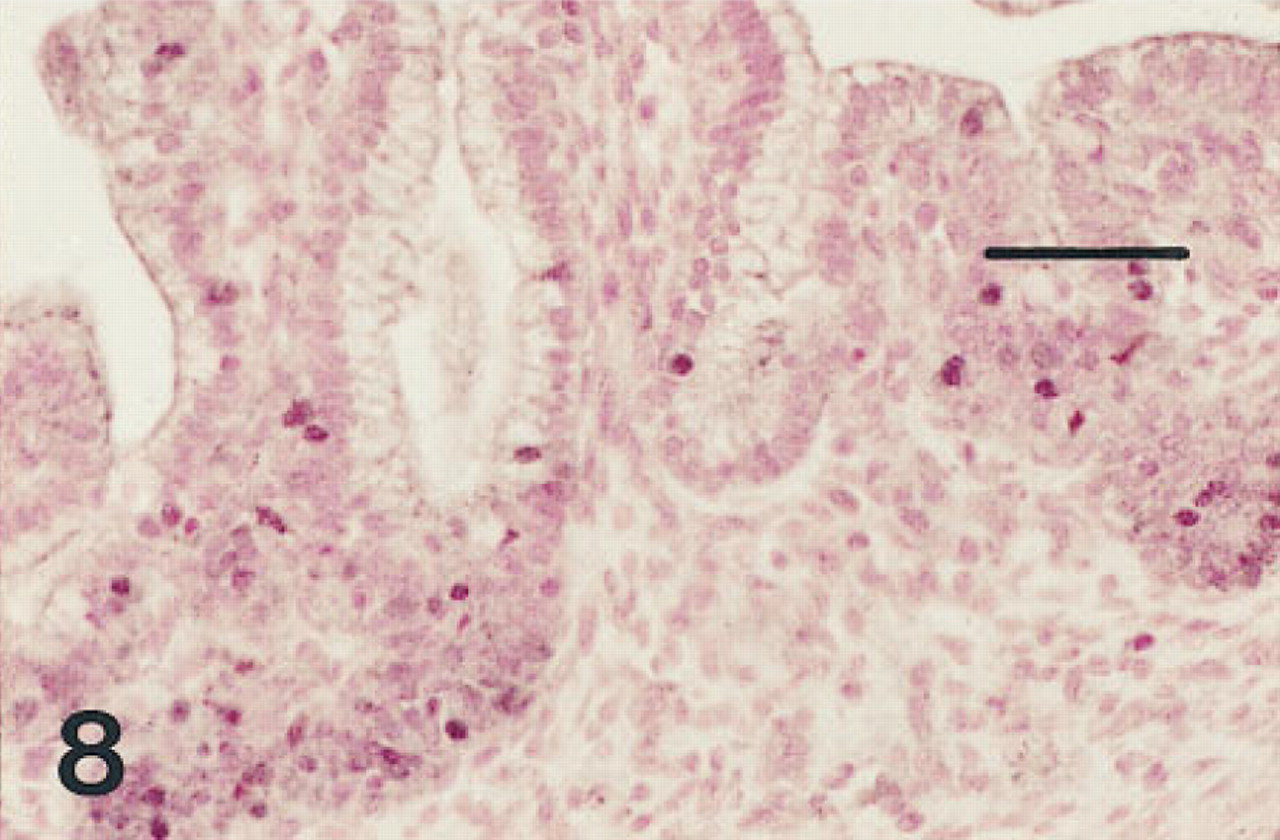
Small intestine of a fetus of 12 weeks' gestation immunostained for HNF-3β. Nuclei of many of the cells in the crypts of Lieberkühn are immunolabeled. Nuclei of some columnar cells of the villi are also immunolabeled. Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 50 μm.
Esophagus. One 12-week fetus and the 15-week fetus in whom the esophagus was available had some scattered cells immunolabeled for all six antisera, but these were more sparse than those labeled in the trachea (Figures 13–16). Esophageal epithelial cells labeled for HNF-3β, TTF-1, and proSP-B were located in basal regions. Those labeled for CCSP were primarily cells flanking folds. By 20 weeks of gestation, the esophagus was lined with stratified squamous epithelium and immunolabeling with these antibodies was no longer seen.
Conducting and Terminal Airways. In the conducting airways of younger fetuses, CCSP was seen only as “crowns” of cuboidal cells adjacent to or over the surface of neuroepithelial bodies (NEBs). TTF-1 and HNF-3β were also detected in these locations and in other epithelial lining cells. NEB cells were not immunolabeled in this series of fetuses by these antisera. In older fetuses, CCSP was seen in subsets of isolated cells in the conducting airways but was not seen in terminal airways at any gestational age (Khoor et al. 1996). In all these fetuses, TTF-1-, HNF-3β-, SP-A-, and proSP-B-labeled cells were found in decreasing numbers along the conducting airways as gestation advanced but, along with proSP-C, were detected in cuboidal pre-Type II cells in many terminal airways. In term infants, mature Type II cells were labeled for TTF-1, HNF-3β, SP-A, proSP-B, and proSP-C.
In the lungs of infants with early BPD, a few isolated cells were immunolabeled for CCSP, while in airways lined by columnar epithelium, clusters of CCSP labeled cells were observed in association with unlabeled NEBs. In bronchioles and in terminal airways, dysplastic relining cells were immunolabeled with HNF-3β and TTF-1, fewer lining cells immunolabeled for proSP-B, and rare cells for SP-A.
Discussion
The temporal-spatial distribution of HNF-3β was determined in developing human tissues, including the lung. The localization and developmental changes in HNF-3β and TTF-1 are consistent with their interacting roles in commitment of foregut endodermal cell differentiation. Later in development, these transcriptional proteins are likely involved in the regulation of target genes, including those of surfactant proteins and CCSP, critical to postnatal lung function. Both transcriptional factors are expressed in subsets of cells believed to be nonterminally differentiated or potential progenitor cells and are excluded during the process of respiratory epithelial cell divergence and terminal differentiation. The overlapping distribution of HNF-3β and TTF-1 in early fetal trachea and esophagus is consistent with a role of these transcription factors as markers for common progenitor cells, contributing to the function of both organs.
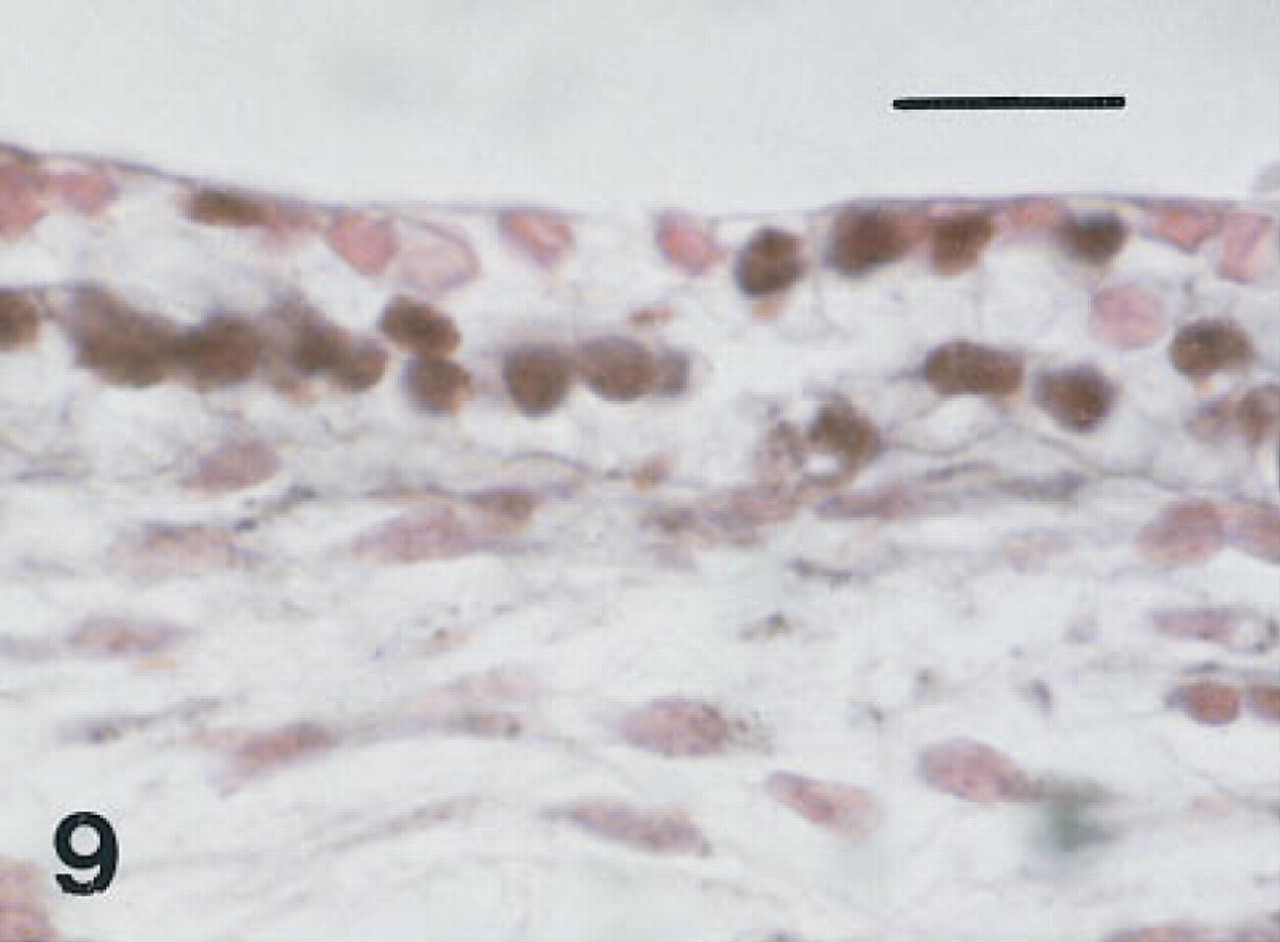
Trachea of a fetus of 10 weeks' gestation immunostained for HNF-3β. Basal nuclei and scattered nuclei in cells at the luminal surface are immunolabeled (black). Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 16 μm.
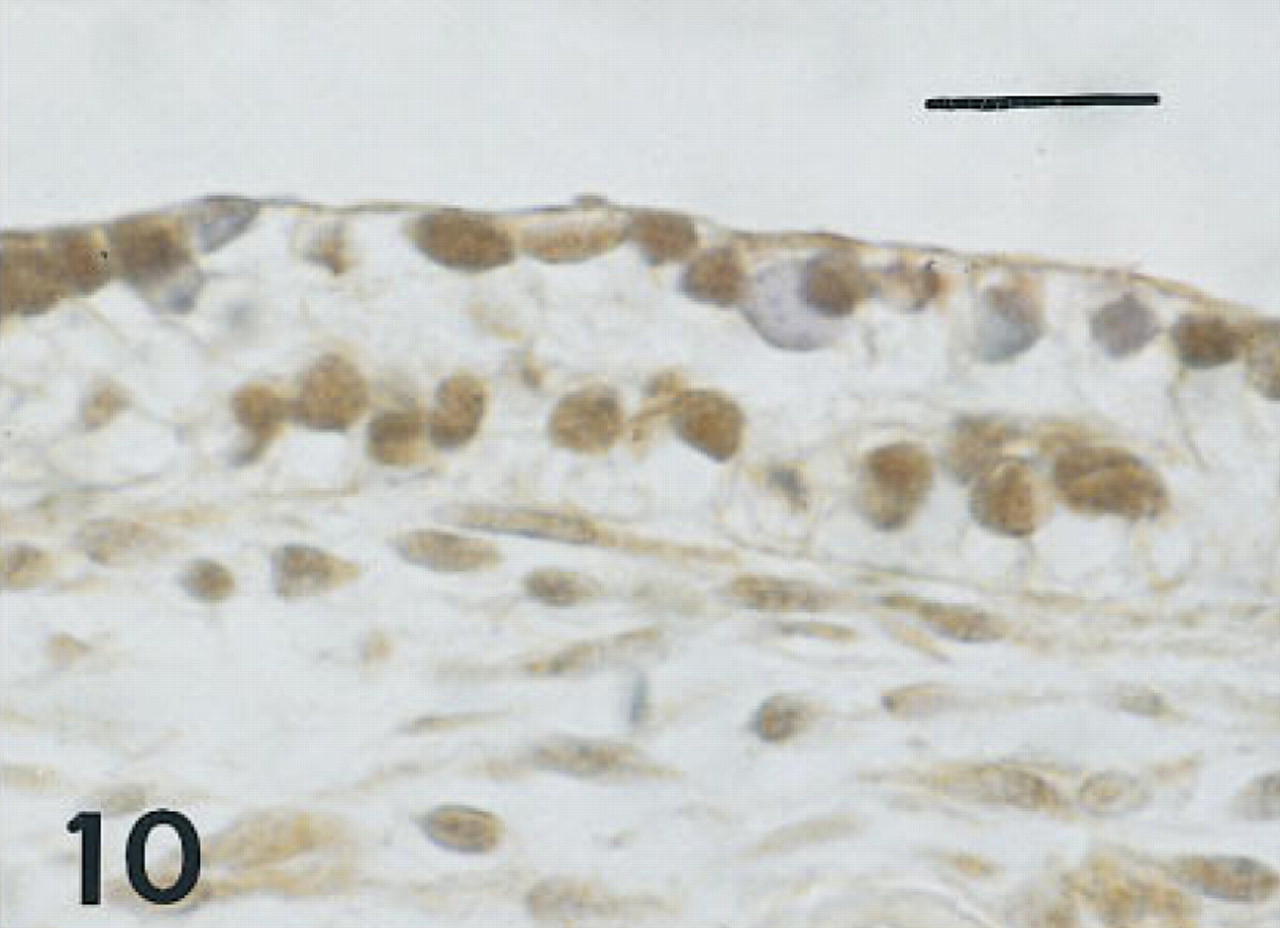
Trachea of the same fetus of 10 weeks' gestation as shown in Figure 9, immunostained for TTF-1. Basal nuclei and some surface nuclei are immunolabeled (brown). Peroxidase-conjugated streptavidin and hematoxylin. Bar = 16 μm.
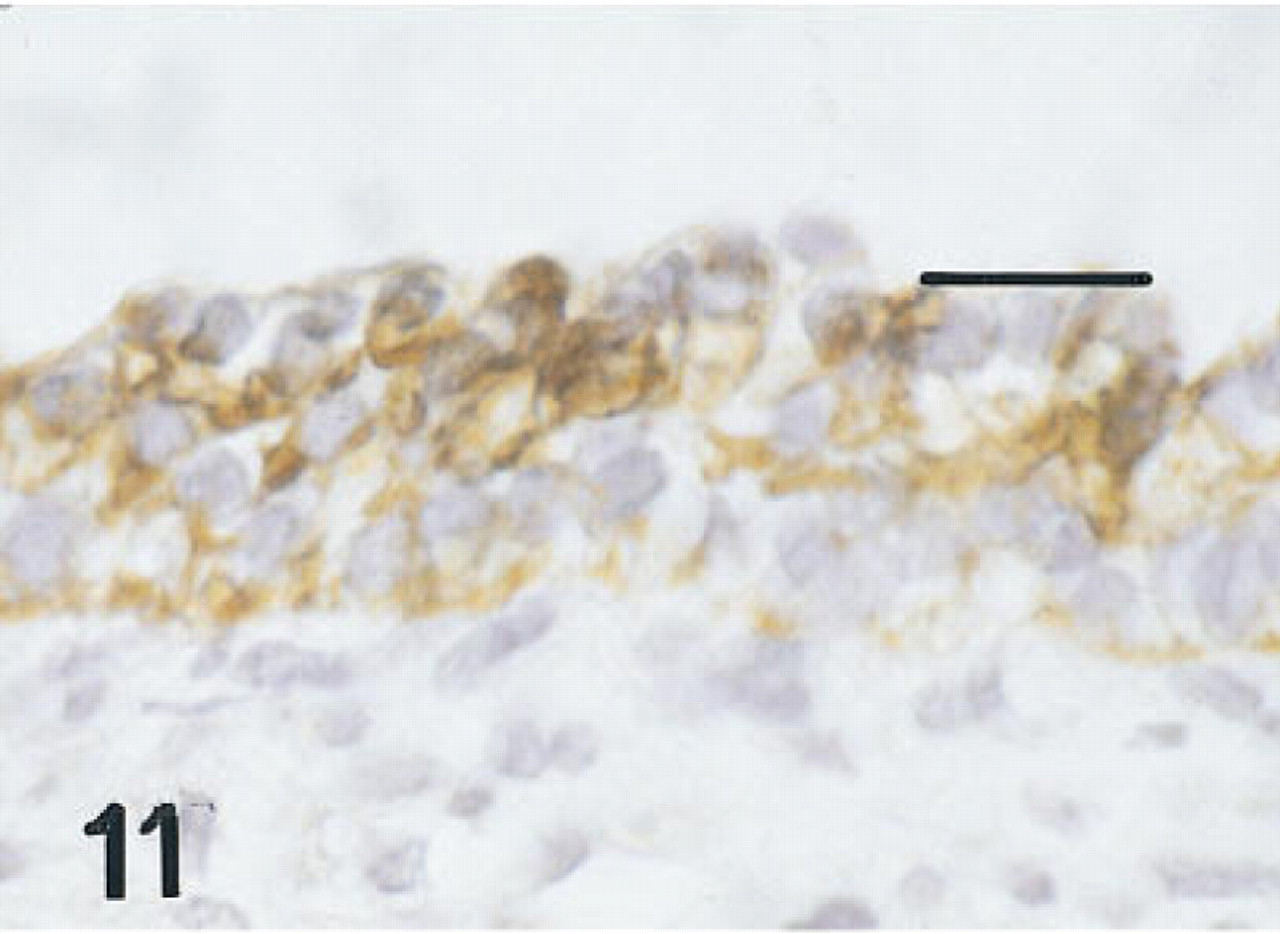
Trachea of a fetus of 12 weeks' gestation immunostained for proSP-B. All membranes at all levels in the epithelium are immunolabeled. Peroxidase-anti-peroxidase and hematoxylin. Bar = 16 μm.
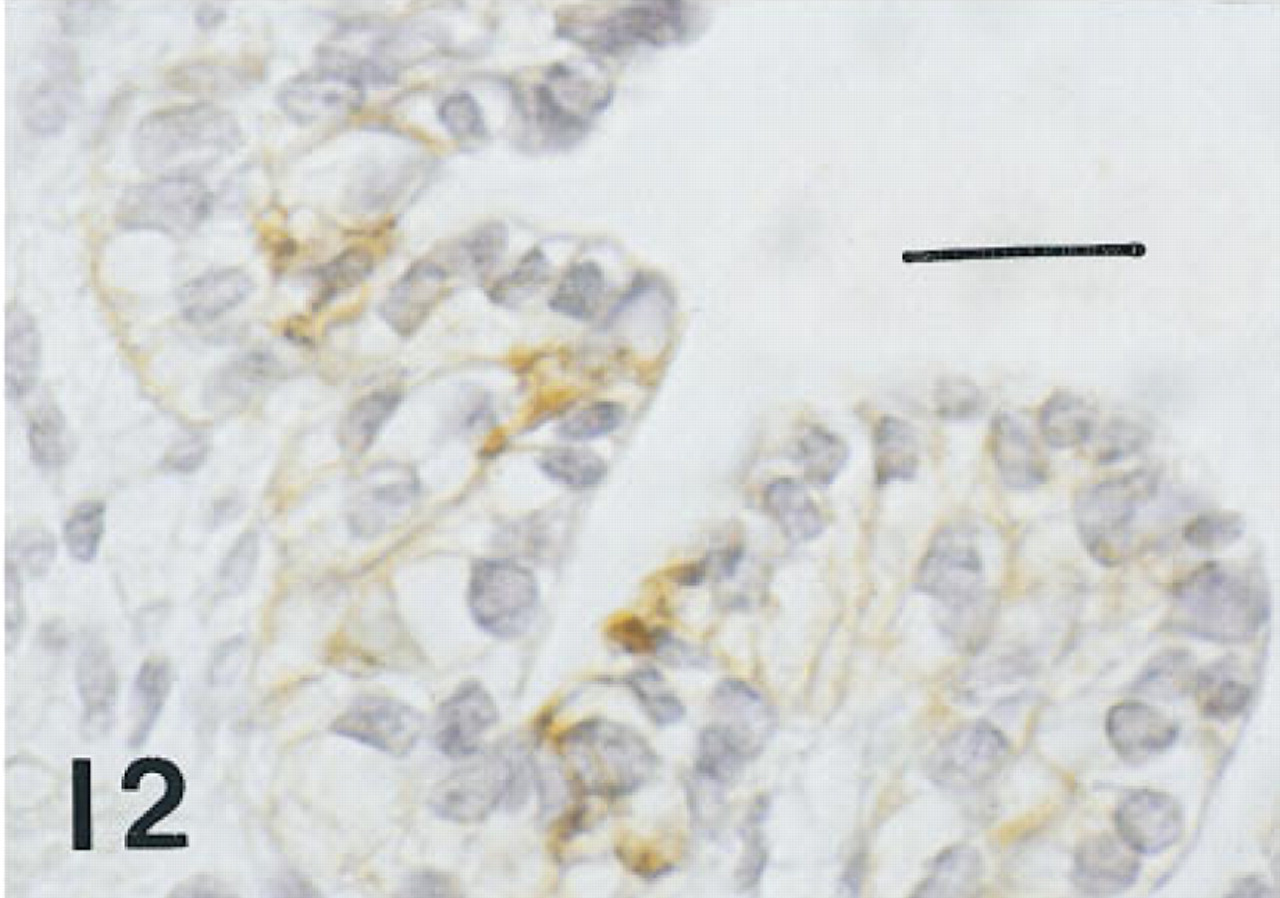
Trachea of the same fetus of 10 weeks' gestation shown in Figure 1, immunostained for CCSP. Membranes of nonciliated surface cells are immunolabeled. Peroxidase-anti-peroxidase and hematoxylin. Bar = 16 μm.
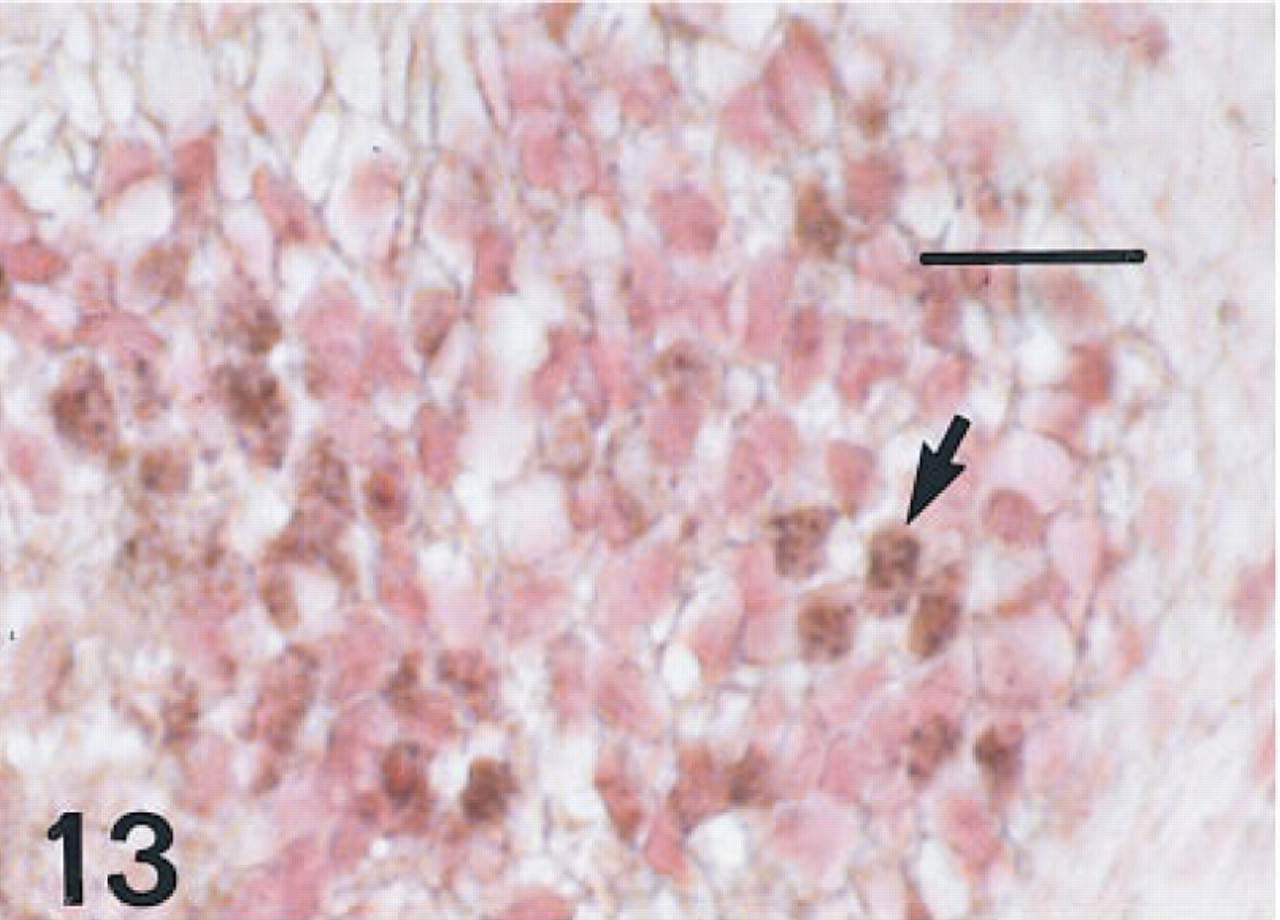
Esophagus of a fetus of 12 weeks' gestation immunostained for HNF-3β, showing multiple nuclei (arrows) in the basal layers of a cross-sectional area. Peroxidase-conjugated streptavidin and nuclear fast red. Bar = 16 μm.
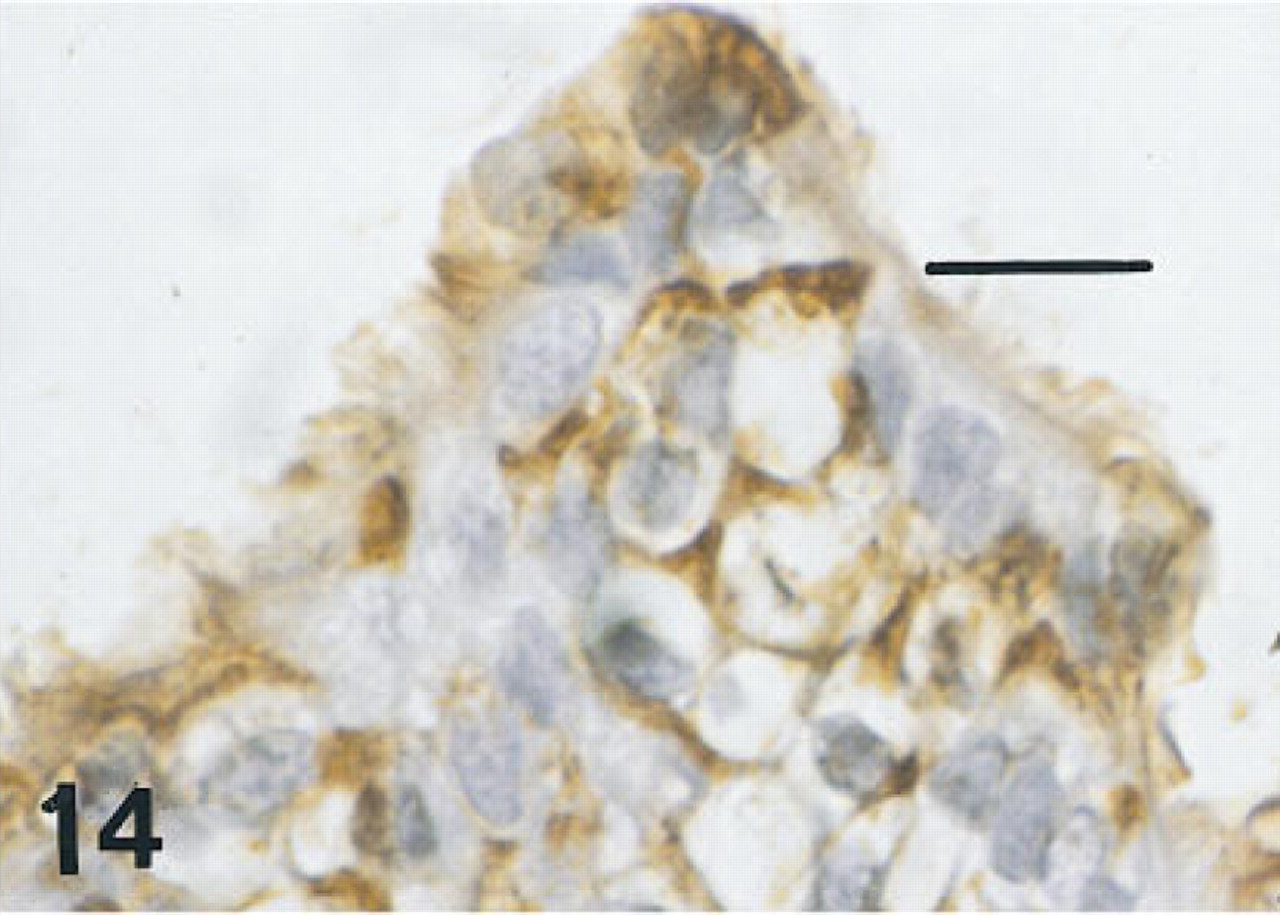
Esophagus of the same fetus of 12 weeks' gestation as shown in Figure 13, immunostained for SP-A. There is intense membrane labeling of nonciliated cells near the luminal surface. Basal cells (not shown) were unlabeled. Peroxidase-anti-peroxidase and hematoxylin. Bar = 16 μm.
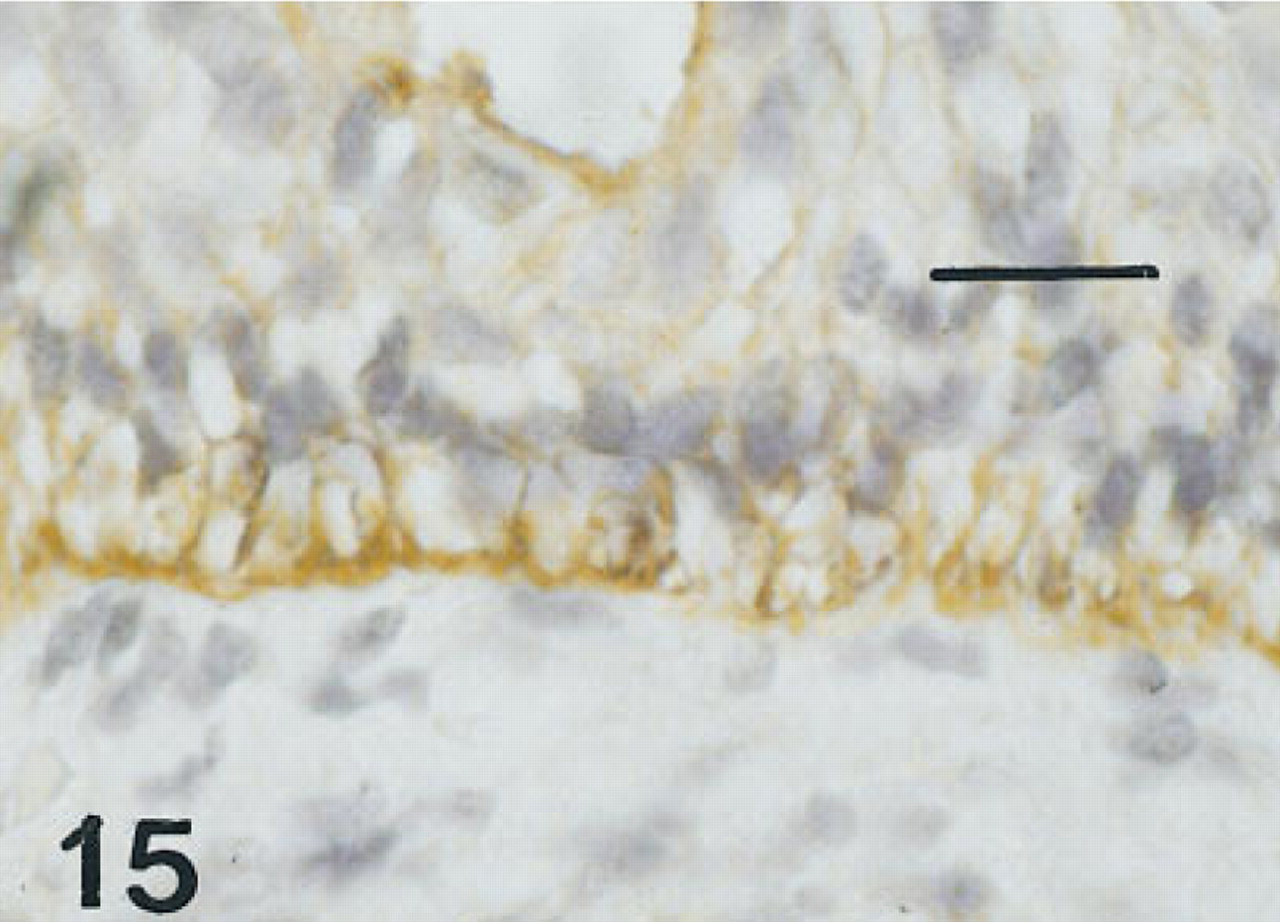
Esophagus of the same fetus of 12 weeks' gestation as shown in Figure 13, immunostained for proSP-B. There is pronounced immunolabeling of the membranes of basal cells. Peroxidase-anti-peroxidase and hematoxylin. Bar = 16 μm.
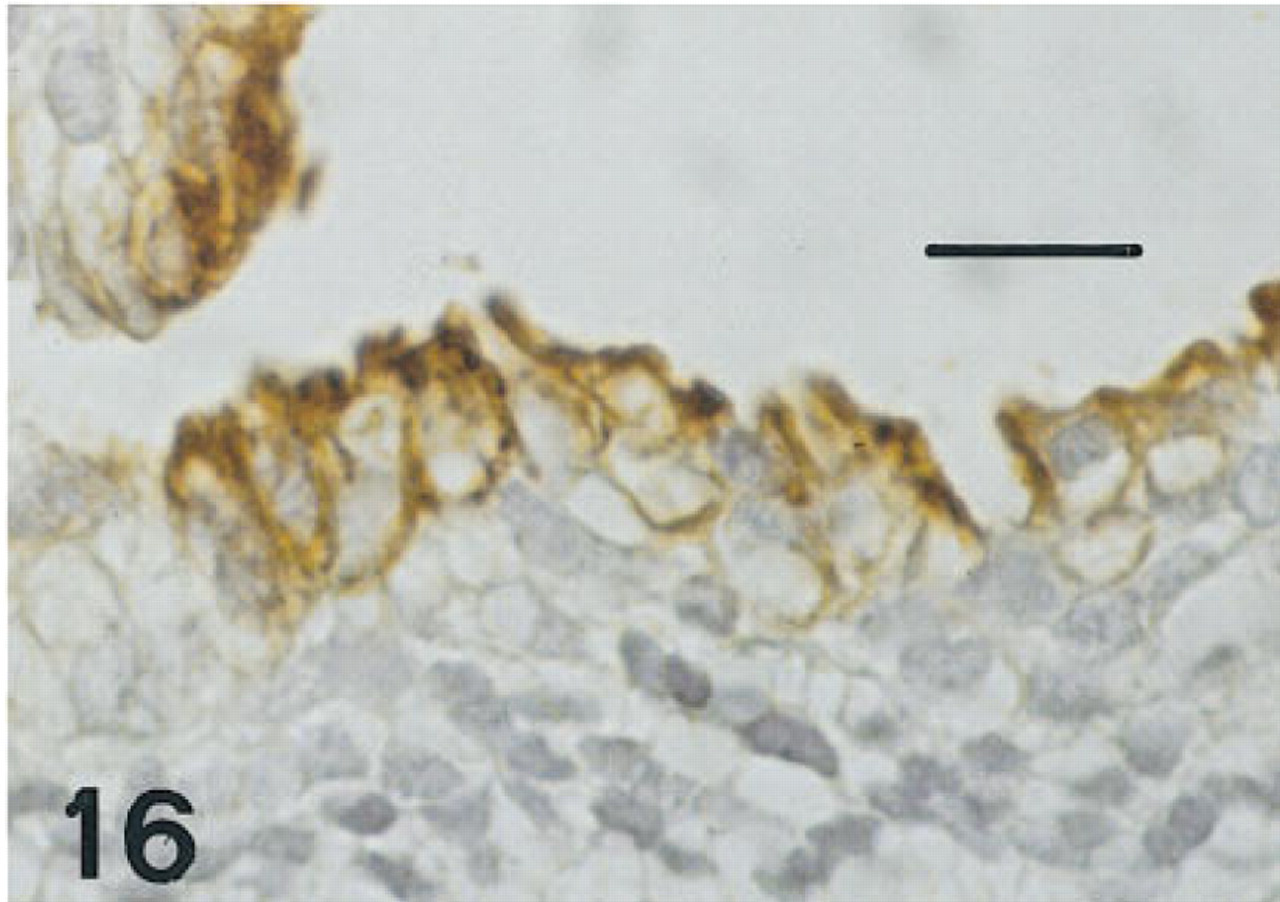
Esophagus of the same fetus of 12 weeks' gestation as shown in Figure 13, immunostained for CCSP. There is intense immunolabeling of both membranes and cytoplasm of nonciliated cells at the luminal surface. Basal cells and ciliated cells are unlabeled. Peroxidase-anti-peroxidase and hematoxylin. Bar = 16 μm.
Very immature mice have shown immunolabeling of all epithelial cells of distal airways with CGRP, CC-10, and SP-A, and later the emergence of staining of differentiated cell types (Wuenschell et al. 1996). Human fetal tissues were not available before 10 weeks of gestation, so the possibility exists that younger fetuses might have had different patterns of immunolabeling, including that of NEBs.
The expression of HNF-3β and TTF-1 was distributed widely in regions of the lung undergoing regeneration after injury. Whereas HNF-3β and TTF-1 were lost consistently in areas of squamous metaplasia and severe lung injury in infants with HMD and BPD, labeling was observed for both proteins in regions undergoing regeneration, often most apparent abutting pleural surfaces and lobar septa. Regions lacking HNF-3β stained with cytokeratin CAM 5.2, demonstrating that antibody accessibility was preserved in these areas. The presence of TTF-1 and HNF-3β in the relining cell population may therefore represent the recapitulation of transcriptional signaling that was also observed during development. HNF-3β was detected in various nonpulmonary tissues, all derivatives of the foregut endoderm. HNF-3β overlapped with TTF-1 in the lung but only transiently in the esophagus and not in other nonpulmonary tissues. Expression of HNF-3β in basal cells of the esophagus and in the stomach and duodenum is also consistent with a potential role in commitment of progenitor cells to certain cell lineages in the developing gastrointestinal tract. A developmental role for HNF-3β is also suggested in that immunolabeling was absent from nonpulmonary tissues of older subjects.
The pattern of increasing restriction of HNF-3β in endodermal tissues during advancing human development is similar to that in the developing mouse. HNF-3β was detected in the mouse CNS and in tissues derived from foregut endoderm, including the lung, esophagus, small intestine, and pancreas (Zhou et al. 1996).
However, this pattern of expression in the developing mouse lung was distinct from that of the human being expressed in the former to a greater degree in conducting airways than in terminal airways. In contrast, the expression of both HNF-3β and TTF-1 in human lung consistently demonstrated an increase in intensity and numbers of cells immunolabeled from the conducting to peripheral airways, most likely reflecting the known proximal to distal gradient of lung epithelial cell differentiation. The biological importance of the differences in the human and mouse distribution may reflect differences in cell type or subtle differences in the function of HNF-3β in these two species.
HNF-3β binds to consensus elements present in a number of genes expressed in foregut endodermal tissues, including those of CCSP and surfactant proteins in the lung (Bohinski et al. 1994). The restriction of extrapulmonary sites of expression of HNF-3β to fetuses below 20 weeks of gestation is consistent with its potential role in gastrointestinal, pancreatic, and tracheal development. Serial sections of tracheas showed the expression of HNF-3β and TTF-1 in subsets of cells, including basal cells, consistent with the known role of both transcription factors in the activation of genes for SP-A, proSP-B, proSP-C, and CCSP. These findings support the concept of shared precursors for the cells expressing these markers. Subsets of cells in the esophagi of early fetuses were also labeled with these antisera. Expression of these proteins was lost in trachea and esophagus with increasing divergence and commitment to differentiated cell phenotypes.
In summary, the temporal-spatial distribution of HNF-3β was assessed in developing human lung and other foregut-derived tissues. HNF-3β was found in the first trimester in subsets of cells that also expressed TTF-1, SP-A, proSP-B, proSP-C, and CCSP in the lung, the trachea, and the esophagus. Expression of both HNF-3β and TTF-1 was increasingly restricted to the distal airways during differentiation. HNF-3β was restricted ultimately to Type II epithelial cells in the postnatal lung and was absent in extrapulmonary tissues. The present studies support the concept that cells expressing HNF-3β and TTF-1 share a close lineage relationship early in development, with disappearance of expression correlating with divergence of cell phenotypes. The persistence of immunolabeling of cuboidal relining cells and Type II cells suggests that HNF-3β and TTF-1 also play a role in influencing cell differentiation and gene expression in the process of repair of the postnatal lung.
Footnotes
Acknowledgements
Supported by HL56387 SCOR and HL14214 SCOR.
We thank Dr Robert Costa of the University of Chicago (Chicago, Illinois) for the use of the antibody generated against recombinant HNF-3β and Dr Gurmukh Singh of Veterans Affairs Medical Center (Pittsburgh, Pennsylvania) for the use of the antibody generated against CCSP. We also thank Ms Sandra Olson and Mr Fred Morris for expert technical assistance, Mr Brent Weedman for photographic assistance, Mr Robert Vantrease for assistance with illustrations, and Ms Nancy L. Hanna for preparation of the manuscript.