
Abstract
We describe an immunohistochemical method that allows the detection of apoptotic cells in human epidermis by use of confocal laser reflectance and antibody-immunogold-silver complexes. For this purpose, the site of free 3′-OH DNA ends was detected by the reflectance from heavy metal products (anti-digoxigenin antibody-immunogold-silver complexes) instead of 3, 3′ -diaminobenzidine (DAB) reaction products in the conventional in situ nick end-labeling of DNA strand breaks (ISEL) technique. Localization of double-stranded DNA was demonstrated by the autofluorescence of methyl green. The ISEL technique using confocal reflectant laser microscopy (CRLM) clearly showed the most intense reflectance in the nuclei of granular cells, in contrast to only a weaker reflectance in those of basal cells. On the other hand, the extent of autofluorescence of methyl green was significantly more intense in the nuclei of basal cells and showed a reciprocal relation to that of the reflectance. Therefore, granular cells were most prone to apoptosis and did not contain double-stranded DNA, as indicated by the lack of stainability with methyl green. In addition, this method demonstrating the simultaneous localization of both free 3′-OH DNA ends and double-stranded DNA proved to have a wide range of applications, including the study of other DNA autolytic processes.
Keywords
T
In this study we modified the conventional ISEL technique to detect apoptotic cells in the human epidermis by using confocal reflectant laser microscopy (CRLM). Free 3′-OH DNA ends detected by the reflectance from the antibody-immunogold-silver complexes were present only in the nuclei of both the spinous and granulosa cells and not in the cytoplasm of the basal cells.
Materials and Methods
Human scalp skin samples were embedded in Tissue Mount (Ciba Medical; Saitama, Japan) and rapidly frozen in acetone cooled on dry ice to prepare 6-7-μm-thick cryosections. Glass slides coated with aminopropyltriethoxysilane were used to eliminate background staining on slides. The sections were fixed in freshly prepared 10% (w/v) paraformaldehyde in 0.01 M PBS for 10 min and postfixed in a 2:1 mixture of ethanol and acetic acid for 5 min at -20C. After washing with PBS three times for 15 min, the sections were incubated with PBS containing 0.5% H2O2 for 20 min at room temperature (RT) to inhibit endogenous peroxidase activity. A commercially available kit (the ApopTag peroxidase kit; Oncor, Gaithersburg, MD) was used for the detection of 3′-OH DNA ends in the sections. After washing with PBS for 15 min, the sections were soaked in the equilibration buffer of the kit for 10-15 sec at RT and then incubated at 37C for 60 min in a moist chamber with 54 μl of the working buffer containing terminal deoxynucleotidyl transferase (TdT), digoxigenin-11-dUTP, and dATP. The reaction was stopped by incubating the sections in a blocking buffer containing Na-citrate and NaCl at 37C for 30 min. After rinsing with PBS three times for 15 min, the sections were used for the conventional ISEL or ISEL using CRLM study.
For the conventional ISEL study, some sections were incubated with an anti-digoxigenin antibody conjugated to horseradish peroxidase at RT for 30 min. After incubation with the antibody, the peroxidase activity was examined by exposing the sections to a solution containing 0.05% DAB and 0.01% H2O2 in Tris buffer, pH 7.6, for 3-6 min at RT. The sections were counterstained with 1% methyl green.
For the ISEL using CRLM study, the remaining sections were incubated with sheep anti-digoxigenin antibody conjugated to 10-nm colloidal gold (British BioCell International; Golden Gate, UK) at RT for 1 hr. After washing in distilled water, the sections were immersed in a physical developer containing silver lactate (Zymed; San Francisco, CA) at RT for 5 min. The reaction of the silver enhancement to the antibody labeled with immunogold particles was stopped by washing in distilled water. The sections were counterstained with 1% methyl green (pH 4.0) for 1 hr at RT. After dehydration with butanol, they were mounted in Entellan Neu (Merck; Darmstadt, Germany). The confocal laser microscope used in this study was an LSM-10 type (Carl Zeiss; Jena, Germany) equipped with two modes for fluorescence and reflectance. Both an argon laser excitation filter at 488 nm and an emission filter were used for detection of the autofluorescence of methyl green, which stains double-stranded DNA. The reflectance from the antibody-immunogold-silver complexes was detected using an argon laser excitation filter at 514 nm.
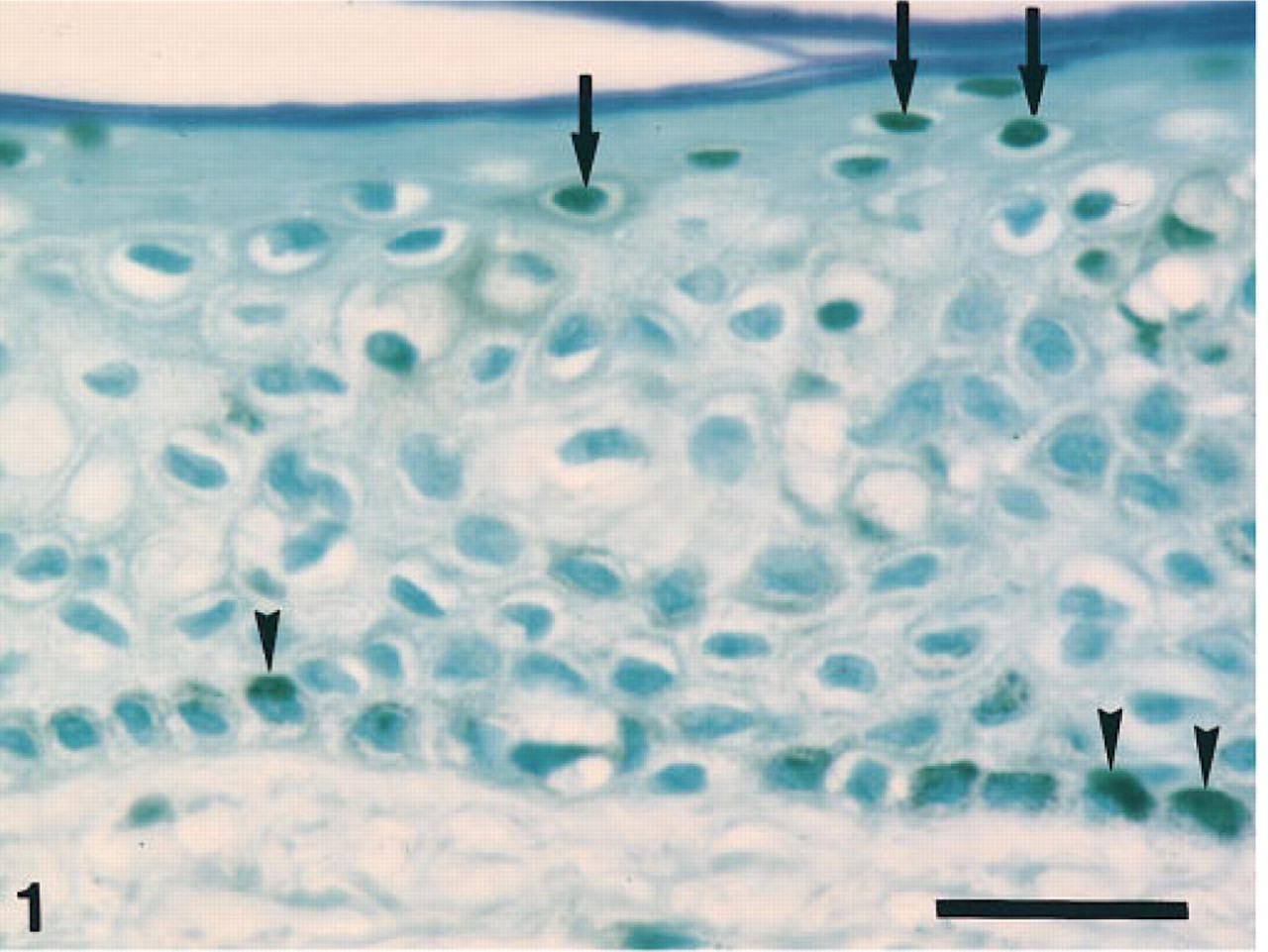
Conventional ISEL technique demonstrates the intense DAB reaction in both the nuclei of the granular cells (arrows) and the cytoplasm of the basal cells (arrowheads). Bar 5 27 μm.
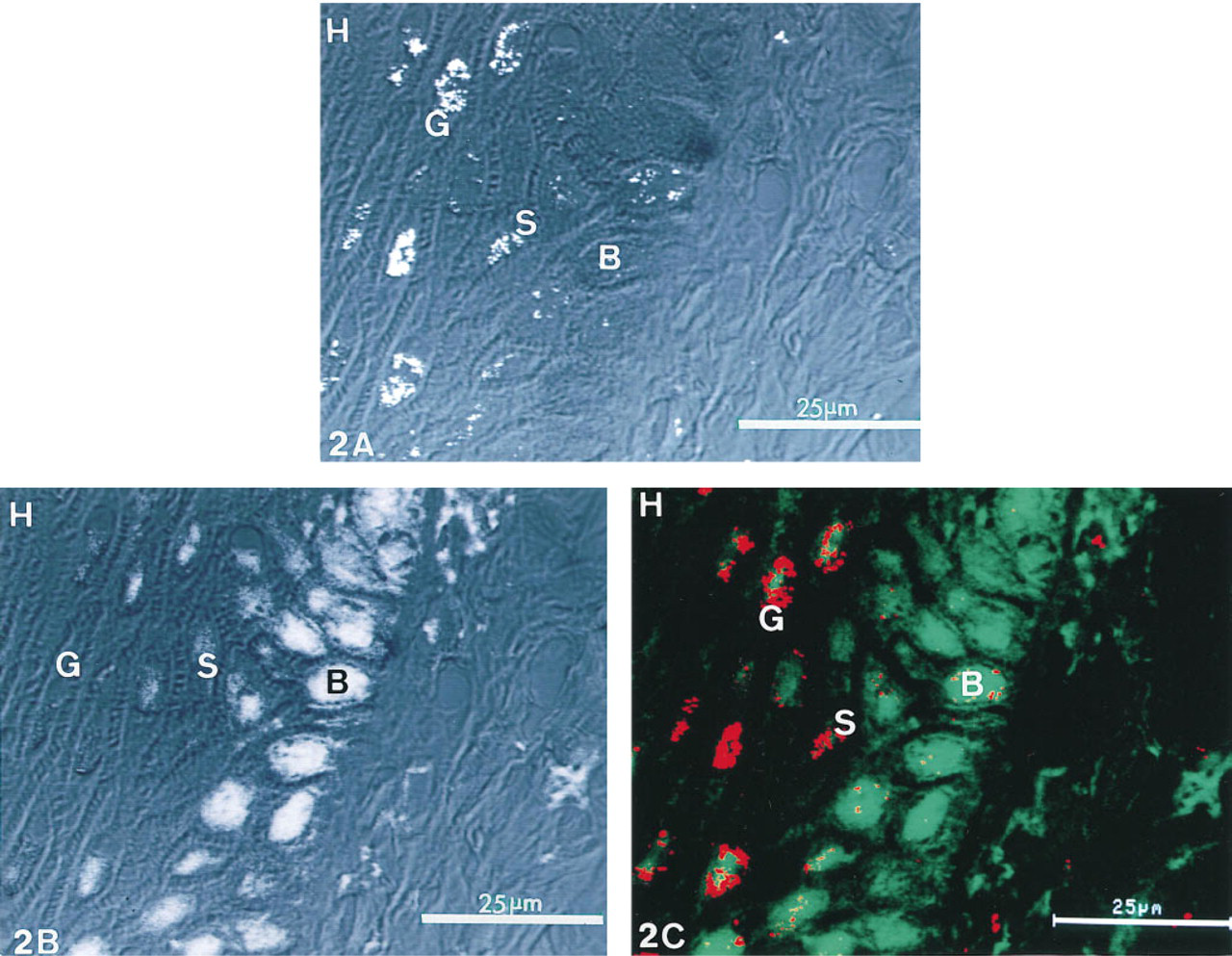
Images of free 3′-OH DNA ends detected by confocal laser reflectance from antibody-immunogold-silver complexes and double-stranded DNA stained with methyl green.
Negative controls were obtained by omitting either the antibody conjugates or the mixture of nucleotides and TdT.
Results
This conventional ISEL study on the human epidermis demonstrated that DAB reaction was observed in both the nucleus and cytoplasm of keratinocytes. The nuclei of most granular cells exhibited intense DAB reaction, although those of spinous cells lacked the reaction. An intense DAB reaction in the cytoplasm was observed in the basal cells and, therefore, often made it difficult to differentiate between the reaction in the nuclei and that in the cytoplasm. Only a few spinous and granular cells exhibited a weak DAB reaction in the cytoplasm (Figure 1).
The ISEL using CRLM study clearly showed that free 3′-OH DNA ends detected via the reflectance from the antibody-immunogold-silver complexes were present only in the nuclei of keratinocytes (Figure 2A). The intensity of the reflectance was different among all epidermal cell layers. The most intense reflectance was observed in the nuclei of granular cells. Moderately intense reflectance was detected in the nuclei of the spinous cells exhibiting a clustered staining pattern. The nuclei of most basal cells demonstrated only weak reflectance at the nuclear periphery. In contrast to the reflectance from antibody-immunogold-silver complexes, autofluorescence of methyl green, which stains double-stranded DNA, was detected in the nuceli of basal cells, but the granular cells lacked the autofluorescence or, if present, contained only weak autofluorescence (Figure 2B). The superimposed image of the reflectance and autofluorescence in the nuclei of spinous cells clearly demonstrated the different localization of the reflectance and autofluorescence. The reflectance was present at the nuclear periphery but the autofluorescence was demonstrated in the center of the nuclei (Figure 2C). The horny cells demonstrated neither reflectance nor autofluorescence (Figures 2A-C).
No reflectance was observed in the controls.
Discussion
Several studies have reported the localization of DNA fragmentation in normal human epidermis via the conventional ISEL technique (Gavrieli et al. 1992; Polakowska et al. 1994; Tamada et al. 1994), but there has been much discussion concerning which cell in the epidermis is most susceptible to treatment with the conventional ISEL technique, only granular cells (Gavrieli et al. 1992; Tamada et al. 1994) or both the granular and spinous cells (Polakowska et al. 1994). In the present study using the conventional ISEL technique, we demonstrated an intense DAB reaction in the nuclei of granular cells and in the cytoplasm of basal cells in the human epidermis. Our findings indicate that all keratinocytes possessing melanosomes with peroxidase activity in the cytoplasm react positively when the conventional ISEL technique with immunoperoxidase is used, regardless of the degree of DNA strand breaks.
Therefore, we applied the ISEL technique using antibody-immunogold-silver complexes instead of an antibody-immunoperoxidase complex for detection of apoptotic cells containing free 3′-OH DNA ends in the human epidermis. The immunogold-silver staining (IGSS) method is extensively employed for light, scanning electron, and transmission electron microscopy (Otsuki and Maxwell 1993) and exhibits a higher labeling sensitivity (more than fourfold) than does the indirect immunoperoxidase staining method (Scopsi et al. 1986; Hayat 1993). The use of CRLM combined with the IGSS method revealed weak reflectance in some nuclei of basal cells. This may be due to DNA strand breaks induced by DNA repair systems or to the site of active gene transcription located in the euchromatin (Thiry 1991; Migheli et al. 1995), rather than to oligonucleosomal degradation and background.
Methyl green has been known to stain doublestranded DNA (Franklin and Filion 1981; Melnick and Pickering 1988; Burres et al. 1993) and is weakly autofluorescent (Rothbarth et al. 1976). The ISEL using CRLM technique combined with methyl green staining enables the simultaneous localization of both 3′-OH DNA ends and double-stranded DNA in apoptotic cells and also the comparison of their concentrations. In this study, the superimposed image of both the reflectance from the antibody-immunogold-silver complexes and the autofluorescence of methyl green clearly demonstrated that apoptotic change involving the newly formed 3′-OH DNA ends began in the spinous cells and was typical in granular cells that lacked the double-stranded DNA. This topographical relationship between 3′-OH DNA ends and doublestranded DNA of keratinocytes is supported by the results of a biochemical study carried out by Suzuki et al. (1977) in which the distribution pattern of DNA in keratinocytes changed during keratinization, resulting in a lack of double-stranded DNA in all electronopaque areas of the nuclei in the granular cells.
Based on the above-mentioned findings, the ISEL using CRLM combined with methyl green staining has a wide range of applications, including the study of other DNA autolytic processes.