
Abstract

P
Neurons from different subregions of the rat brain, including the cerebellum and the hippocampus, express PTHrP and PTH/PTHrP receptors (Weir et al. 1990; Ureña et al. 1993). In the rat, PTHrP has also been detected in the meninges but not in astrocytes that have PTH/PTHrP receptors (Struckhoff and Turzynski 1995). We evaluated the presence of PTHrP in human astrocytomas, using immunohistochemistry and Western immunoblot analysis.
Four low-grade astrocytomas and five high-grade astrocytomas (Daumas–Duport et al. 1988) were studied. Fresh tumor samples were fixed overnight in 10% buffered formalin and embedded in paraffin before sectioning. The histological features were evaluated on hematoxylin- and eosin-stained tumor sections. Cytologically, the constituent tumor cells varied in size and in histological appearance. They consisted of small cells with hyperchromatic nuclei and larger cells with glassy cytoplasm and the morphological appearance of astrocytes.
Immunostainings were performed with anti-PTHrP antisera C6 and C13, recognizing the C- and N-terminal region of PTHrP, respectively (Albar et al. 1996), and MIB 1 monoclonal specific antibody (Immunotech; Marseille, France), recognizing the Ki-67 proliferation marker, at 1:100 dilution, by the avidin–biotin–peroxidase complex method. Paraffin-embedded tissue sections (5 μm) were mounted on glass slides pretreated with 1 g/liter poly-
Positive immunostaining for PTHrP was found in all nine astrocytomas with antisera C6 and C13. With either antiserum, the staining was diffuse in the cytoplasm of neoplastic cells and was prominent in the larger cells with a more astrocytic appearance (Figures 1A and 1B). These cells were abundant in the five high-grade astrocytomas, whose proliferative activity, expressed as percentage of MIB 1-positive nuclear area was (mean ± SD) 17 ± 5%, greater than 5 ± 3% in the four low-grade cases. Adjacent nonneoplastic cortex in the tissue samples did not stain for PTHrP. No immunostaining was observed on incubation without these primary antisera (Figure 1C).
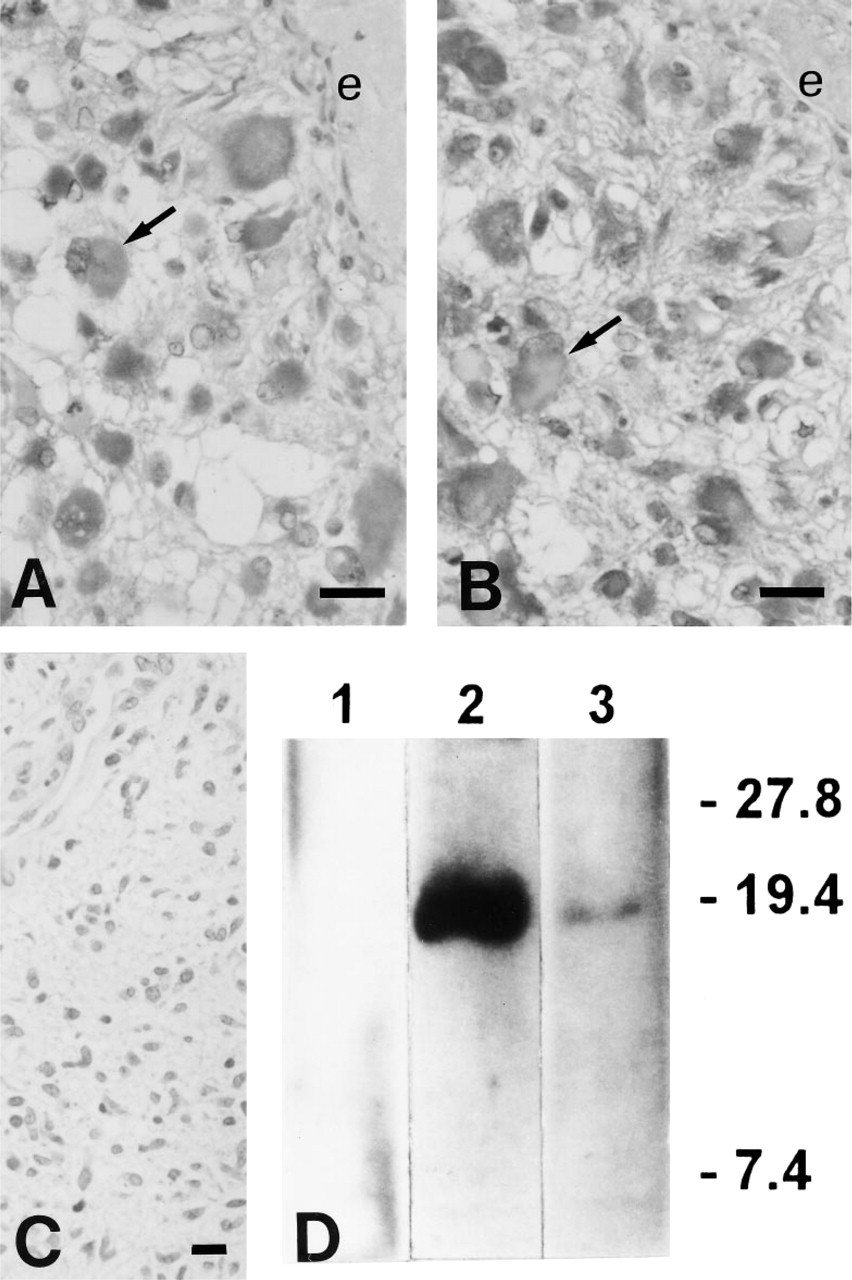
Intense PTHrP immunostaining in larger cells (arrows) in a high-grade astrocytoma with anti-sera C6 (
One of the tumors, an astrocytoma of high grade 4, was selected to further characterize PTHrP by Western immunoblotting. A fresh tumor sample (1.2 g) was extracted with 0.2 M HCl/8 M urea/0.1 M cysteine and then with 0.1 M NaCl/20% ethanol/10 mM acetic acid (de Miguel and Esbrit 1992). Aliquots of tumor extract were run on a 15% sodium dodecylsulfate-polyacrylamide gel and then electrophoretically transferred onto nitrocellulose membrane filters. The blotted membrane was blocked with 5% bovine serum albumin and then incubated for 1 hr with anti-sera C6 and C13, at 1:5000 and 1:2000 dilution, respectively, at RT. The membrane was then incubated with peroxidase-labeled goat anti-rabbit IgG and developed by an enhanced chemiluminescence method (Amersham; Poole, UK). A protein band with an apparent molecular weight of 18 kD, consistent with the mature PTHrP form, was detected with either anti-PTHrP antiserum (Figure 1D).
All our cases of astrocytoma were normocalcemic at presentation. In this regard, several studies indicate that PTHrP expression can occur in a variety of tumors in the absence of hypercalcemia (Ralston et al. 1991; Abdeen et al. 1995).
PTHrP is widely expressed during normal fetal development and is present in various adult tissues, in which it appears to modulate cell growth and/or differentiation (Philbrick et al. 1996). At least in some tumors, either PTHrP overproduction in a fashion dissociated from the control of cell proliferation/differentiation or PTHrP gene re-expression could affect their own growth (Henderson et al. 1991; Albar et al. 1996).
In the rat, the PTH/PTHrP receptor, but not PTHrP, is highly expressed in cortical astrocytes, in which PTHrP[1–34] induces a process-bearing cell morphology and increases cellular glial fibrillary acidic protein (Struckhoff and Turzynski 1995). In these cells, PTHrP inhibits transforming growth factor-β-stimulated release of dipeptidyl peptidase II, a glial enzyme involved in glial scarring (Struckhoff 1995). Therefore, in the rat brain PTHrP might control normal astrocyte differentiation by complex paracrine mechanisms.
Taken together, the latter findings and our present results suggest that production of PTHrP by human astrocytomas may be associated with dysregulated mechanisms of growth and/or differentiation in malignant astrocytoma cells. Elucidation of the factors that induce PTHrP expression and its role in these tumor cells await further studies.
Footnotes
Acknowledgements
Supported in part by a grant from Fondo de Investigación Sanitaria of Spain (FIS 94/269). F. de M. is the recipient of a fellowship from the Spanish Ministry of Health (BAE 95/5438).
We thank Ms Trinidad Carrizosa for her technical assistance.