
Abstract
To test the hypothesis that the c-mpl ligand is not a primary factor in thromb-ocytopoiesis, we investigated the biological effects of recombinant human (rh) c-mpl ligand on differentiation of murine progenitor cells and on maturation of the cultured murine megakaryocytes under serum-free conditions on the basis of ploidy distribution, megakaryocyte/platelet-specific surface antigen CD 61 [glycoprotein (GP) IIIa], and cytoplasmic acetylcholinesterase (AchE) expression in vitro. In addition, we studied the effect of c-mpl ligand on proplatelet formation (PPF) by murine mature megakaryocytes. AchE was less strongly expressed in cultured megakaryocytic cells stimulated by c-mpl ligand than in those stimulated by recombinant murine (rm) IL-3 + rh IL-6 during the differentiation of progenitor cells. Less CD 61 was expressed by c-mpl ligand during both the differentiation of progenitor cells and the maturation of megakaryocytes compared with that by rm IL-3 + rh IL-6. Endomitosis, however, was more stimulated by c-mpl ligand than by rm IL-3 + rh IL-6 under both conditions. Furthermore, PPF of mature megakaryocytes was not stimulated by c-mpl ligand. These results indicate that c-mpl ligand stimulates the nuclear development of megakaryocytic cells but that it does not stimulate cytoplasmic maturation and PPF as much as IL-6. These data strongly suggest that c-mpl ligand is not a primary factor in platelet production.
Keywords
C-mpl ligand is a novel cytokine that stimulates mega-karyocytopoiesis and thrombocytopoiesis (de Sauvage et al., 1994; Kaushansky et al., 1994; Lok et al. 1994; Wendling et al., 1994). It is reported to stimulate the expansion of megakaryocytic progenitor cells in vivo, resulting in increased numbers of megakaryocytes in mouse bone marrow and spleen, followed by increased numbers of platelets in the peripheral blood (Kaush-ansky et al. 1994). This cytokine also supports mega-karyocytic colony formation and induces an increase in size and ploidy of megakaryocytes in vitro (Kaushansky et al. 1994). C-mpl is expressed only in mega-karyocytic lineage cells, including CD34-positive cells, megakaryocytes, and platelets (Debili et al. 1995a). Therefore, c-mpl ligand is functional only in mega-karyocyte/platelet lineage cells (Methia et al. 1993) and has been called thrombopoietin (TPO) (Metcalf 1994). Gurney et al. (1994) generated c-mpl-deficient mice (c mpl -/-) by gene targeting. The c-mpl-/- mice had an 85% decrease in the number of platelets and mega-karyocytes but normal amounts of other hematopoietic cells. This result indicates that c-mpl ligand plays significant roles in megakaryocytopoiesis and thrombocytopoiesis. In addition, 15% of platelets are considered to be produced through a salvage pathway other than that of c-mpl ligand/c-mpl. Recently, Shivdasani et al. (1995) reported megakaryocytopoiesis and throm-bocytopoiesis in mice lacking the hematopoietic sub-unit of the heterodimeric erythroid transcription factor NF-E2. They lack circulating platelets despite the presence of the megakaryocytes in bone marrow. These findings provide insight into the regulation mechanism of megakaryocyte maturation and thrombocytopoiesis. Therefore, in this study we investigated the biological effects of c-mpl ligand on megakaryocyte differentiation and maturation and on platelet production in vitro.
Materials and Methods
Reagents and Animals
Recombinant human (rh) c-mpl ligand and rh interleukin 6 (IL-6) were generously provided by Kirin Brewery (Tokyo, Japan) and Ajinomoto (Tokyo, Japan), respectively. Recombinant murine (rm) IL-3 and FITC-labeled anti-murine CD 61 antibody were obtained from Pharmingen (San Diego, CA). Propidium iodide (PI) was purchased from Sigma (St Louis, MO). Male ddY mice were obtained from Shizuoka Laboratory Center (Shizuoka, Japan).
Cell Culture
Preparation of Nonadherent and Nonphagocytic Cells
Male ddY mice 6–8 weeks old were used in all studies. Nonadherent and nonphagocytic cells were obtained as described (Rinehart et al., 1978; Kumagai et al. 1979) with minor modification. After murine bone marrow cells were suspended in CATCH medium (Levine et al. 1980) ([Ca,-Mg-free Hanks balanced salt solution with 0.38% sodium citrate, 1 mM adenosine, 2 mM theophylline, prostaglandin E1 (1 mg/ml), (Sigma) and 3.5% bovine serum albumin (BSA, Fraction V; Sigma)], platelets were removed by mild centrifugation (100 × g, 20 min). The pellet was suspended in IMDM (Life Technologies; Grand Island, NY) supplemented with 10% fetal bovine serum (FBS) (ICN Biomedicals; Costa Mesa, CA) and cultured in 5% CO2 and 95% O2 for 90 min. Silica gel (KAC-2; JIMRO, Takasaki, Japan), which was used for phagocytosis by phagocytic cells, was added to the nonadherent cell suspension and cultured in 5% CO2 and 95% O2 for 60 min with shaking every 15 min. The cell suspension was layered onto Lympholyte M (Cedar Lane Laboratories; Hornby, Ontario, Canada) and centrifuged at 400 × g for 20 min. The cells between Lympholyte M and the medium were collected and washed twice with IMDM.
Culture of Nonadherent and Nonphagocytic Cells
Nonadherent and nonphagocytic cells were cultured in IMDM supplemented with 1% Nutridoma (Behringer Mannheim Biochemica; Mannheim, Germany) with rh c-mpl ligand (20 ng/ml) or rm IL-3 (10 U/ml) + rh IL-6 (100 ng/ml) at a density of 1 × 106 cells/ml for 3 days. The cultured megakaryocytes were partially purified using an albumin gradient as described (Ito et al. 1996). They were cytospun on the slides and stained for AchE.
Nonadherent and nonphagocytic cells were cultured for 4 days under the same conditions as described above. Two-color analysis (CD 61 expression and ploidy distribution) of the cultured cells was performed as described below.
Nonadherent and nonphagocytic cells were cultured in IMDM supplemented with 1% Nutridoma and c-mpl ligand (20 ng/ml) for 3 days. The cells were washed twice with IMDM and re-cultured with various concentrations of rh c-mpl ligand (1, 20, and 100 ng/ml) or rh IL-6 (10 and 100 ng/ml) for 2 additional days at a density of 1 × 106 cells/ml. These cells were analyzed flow cytometrically as described below.
To estimate the stimulating activity of mpl ligand on DNA development of megakaryocytes, culture of nonadherent and nonphagocytic cells was performed for 4 days with various concentrations of rh c-mpl ligand (0.2, 2, and 20 ng/ml), and nonadherent and nonphagocytic cells were cultured with rh c-mpl ligand (20 ng/ml) for 3, 4, and 5 days. The ploidy distribution of the cultured megakaryocytes was analyzed flow cytometrically as described below.
AchE Staining of the Cultured Megakaryocytes
The cells cytospun on the slides with 2% BSA were fixed with 1% glutaraldehyde for 10 min. (Nakeff et al. 1976), washed twice with phosphate buffered saline (PBS) and stained for AchE as described (Karnovsky and Roots 1964).
Two-color Flow Cytometry of Cultured Megakaryocytes: Analysis of Ploidy Distribution of Cultured Megakary-ocytes and CD 61 Expression at each Ploidy Class
Cultured megakaryocytes were stained with FITC-labeled antimurine CD 61 antibody (Pharmingen) and propidium iodide (Sigma) for CD 61 and nucleus in megakaryocytes, respectively, according to the method described (Jackson et al. 1984). Murine lymphocytes were used as the definition of 2N ploidy. The dots of cells with each ploidy (8, 16, 32N) that were positive for CD 61 expression in the dimension with the ploidy distribution (PI fluorescence) and forward scatter were gated, as shown in Figure 2. A histogram of the FITC fluorescence of gated cells with each ploidy class was constructed and the mean FITC fluorescence intensity was calculated by the Cell Quest program (Becton Dickinson; Mountain View, CA) (Figure 2). The level of CD 61 expression was expressed as mean FITC fluorescence. FITC-labeled beads (Becton Dickinson) were used as the internal control for FITC fluorescence. The fluorescence of the beads was adjusted at 70 in the FITC fluorescence histogram.
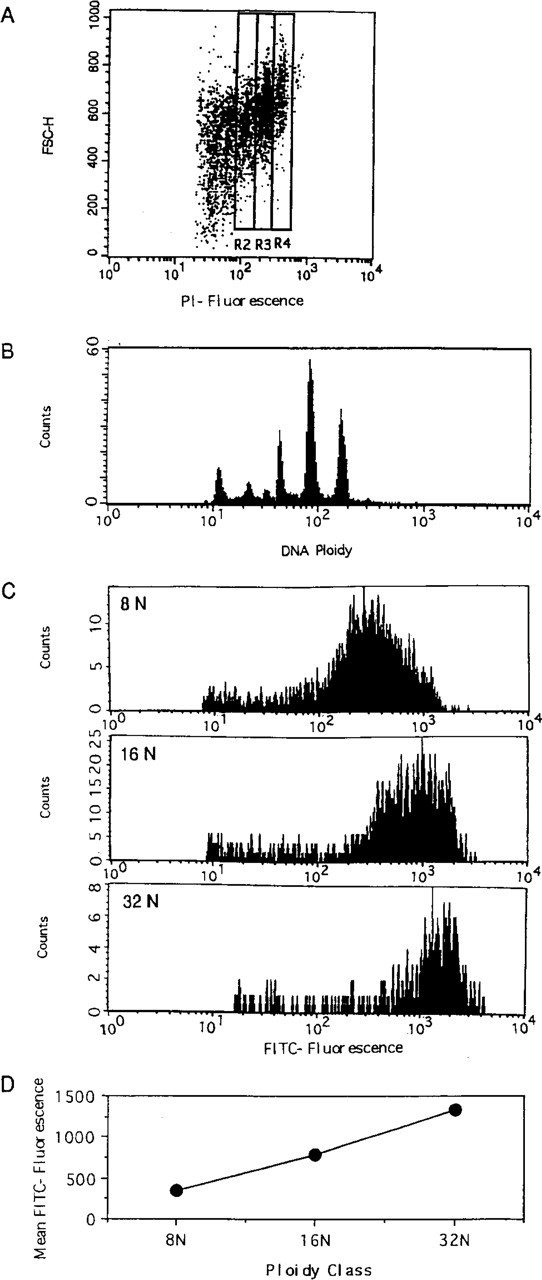
CD 61 fluorescence of murine bone marrow megakaryocytes with each ploidy class. Murine bone marrow megakaryocytes were stained with FITC-labeled anti-murine CD 61 antibody and propidium iodide (PI). (
Proplatelet Formation by the Murine Megakaryocytes
Murine megakaryocytes were partially purified from bone marrow cells using an albumin gradient as described (Ito et al. 1996). The megakaryocytes were cultured in IMDM containing 5% albumin (BSA Fraction V) and rh c-mpl ligand or rh IL-6 (100 ng/ml). After a 24-hr incubation, the cells were fixed with 1% glutaraldehyde and stained for AchE (Karnovsky and Roots 1964). Under inverted microscopy at 400-fold magnification, the numbers of AchE-positive cells and megakaryocytes with proplatelet formation were counted. The proportion (%) of megakaryocytes with proplatelet formation (% megs w/ PPF) is expressed using the following formula:
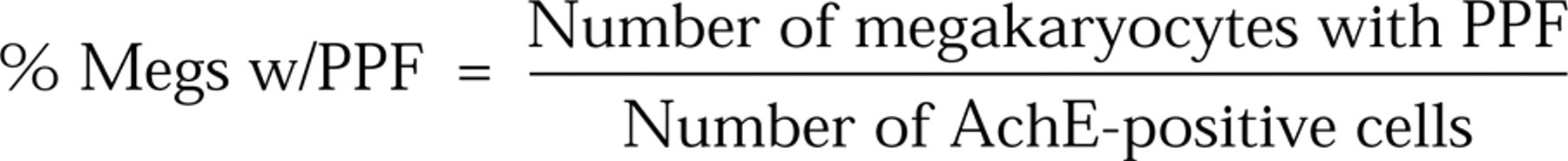
Statistical Analysis
Student's t-test was used to determine statistically significant differences.
Results
Culture of Nonadherent, Nonphagocytic Cells
To evaluate the stimulatory activity of rh c-mpl ligand on the cytoplasmic maturation of megakaryocytic cells during differentiation, we cultured murine bone marrow cells for 3 days. AchE expression by the cultured megakaryocytes stimulated with rh c-mpl ligand (20 ng/ml) was compared with that induced by rm IL-3 (10 U/ml) + rh IL-6 (100 ng/ml). Figure 1 shows the intensity of AchE staining and the size of each megakaryocyte. Most of the cultured megakaryocytes, including large and small cells, stimulated by rh c-mpl ligand were much less stained for AchE, whereas those cultured with rm IL-3 + rh IL-6 were stained more intensely. The megakaryocytes stimulated by rh c-mpl ligand, however, were larger than those stimulated by rm IL-3 + rh IL-6. To confirm these results, we evaluated the expression of the platelet-specific surface marker CD 61 by the cultured megakaryocytes for 4 days. In preliminary experiments, we performed two-color analysis of the cultured megakaryocytes with various concentrations of rh c-mpl ligand (0.2, 2, or 20 ng/ml) for 4 days (Figure 3). Because 20 ng/ml of rh c-mpl ligand stimulated the highest shift of ploidy distribution in megakaryocytes, the concentration of rh c-mpl ligand (20 ng/ml) was chosen in the following experiments (Tables 1 and 2). The mean FITC fluorescence (CD 61 expression) at each ploidy (8 N, 16 N, 32 N) was approximately 1.4–1.7-fold increased (1.4-, 1.62-, and 1.74-fold increase in megakaryocytes with 8 N, 16 N, and 32 N ploidy class, respectively) in the cultured megakaryocytes stimulated by rm IL-3 + rh IL-6 over those stimulated by rh c-mpl ligand (Table 1). In addition, the maturation activity of rh c-mpl ligand or rh IL-6 on the mature megakaryocytes was evaluated. We cultured murine bone marrow cells with rh c-mpl ligand (20 ng/ml) to become mature megakaryocytes, then incubated them for 2 more days with rh c-mpl ligand (1, 20, or 100 ng/ml) or rh IL-6 (10 or 100 ng/ml). Rh IL-6 (100 ng/ml) stimulated a 1.2-, 1.25-, 1.44-, and 1.47-fold increase of CD 61 expression in the megakaryocytes with 8 N, 16 N 32 N, and 64 N ploidy class, respectively, over those stimulated by rh c-mpl ligand (20 ng/ml) (Table 3). However, c-mpl ligand stimulated a shift towards a higher ploidy class of megakaryocytes than rm IL-3 + rh IL-6 in nonadherent and nonphagocytic cells cultured for 4 days (Table 2). In addition, the ploidy distribution of megakaryocytes after 3 days of culture with rh c-mpl ligand (20 ng/ml), followed by 2 additional days of culture with rh c-mpl ligand (1, 20, or 100 ng/ml) or rh IL-6 (10 or 100 ng/ml) is shown in Table 4. The megakaryocytes with higher ploidy class were observed more frequently in the cultures with c-mpl ligand than in those with rh IL-6. To estimate the stimulatory effect of c-mpl ligand on DNA development of megakaryocytes, the culture period was extended to 5 days with rh c-mpl ligand (20 ng/ml). The ploidy distribution of the megakaryocytes is shown in Figure 4. After culture for 5 days, megakaryocytes with high ploidy class (32 N, 64 N, and 128 N) were frequently observed.
AchE expression by cultured megakaryocytes. Nonadherent and nonphagocytic cells were cultured with rh c-mpl ligand (20 ng/ml) (
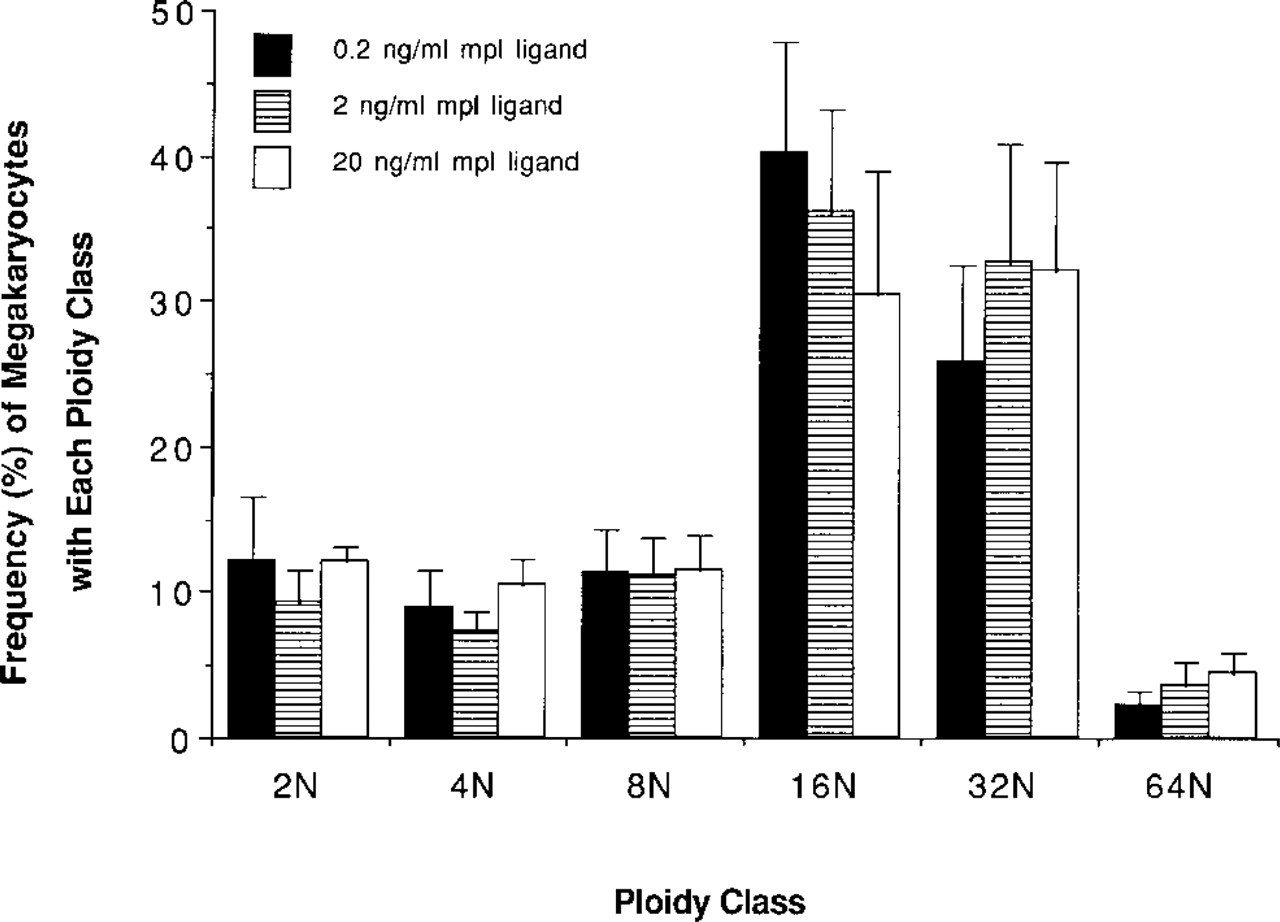
Dose-response change of ploidy class in megakaryocytes. Nonadherent and nonphagocytic cells were cultured for 4 days.
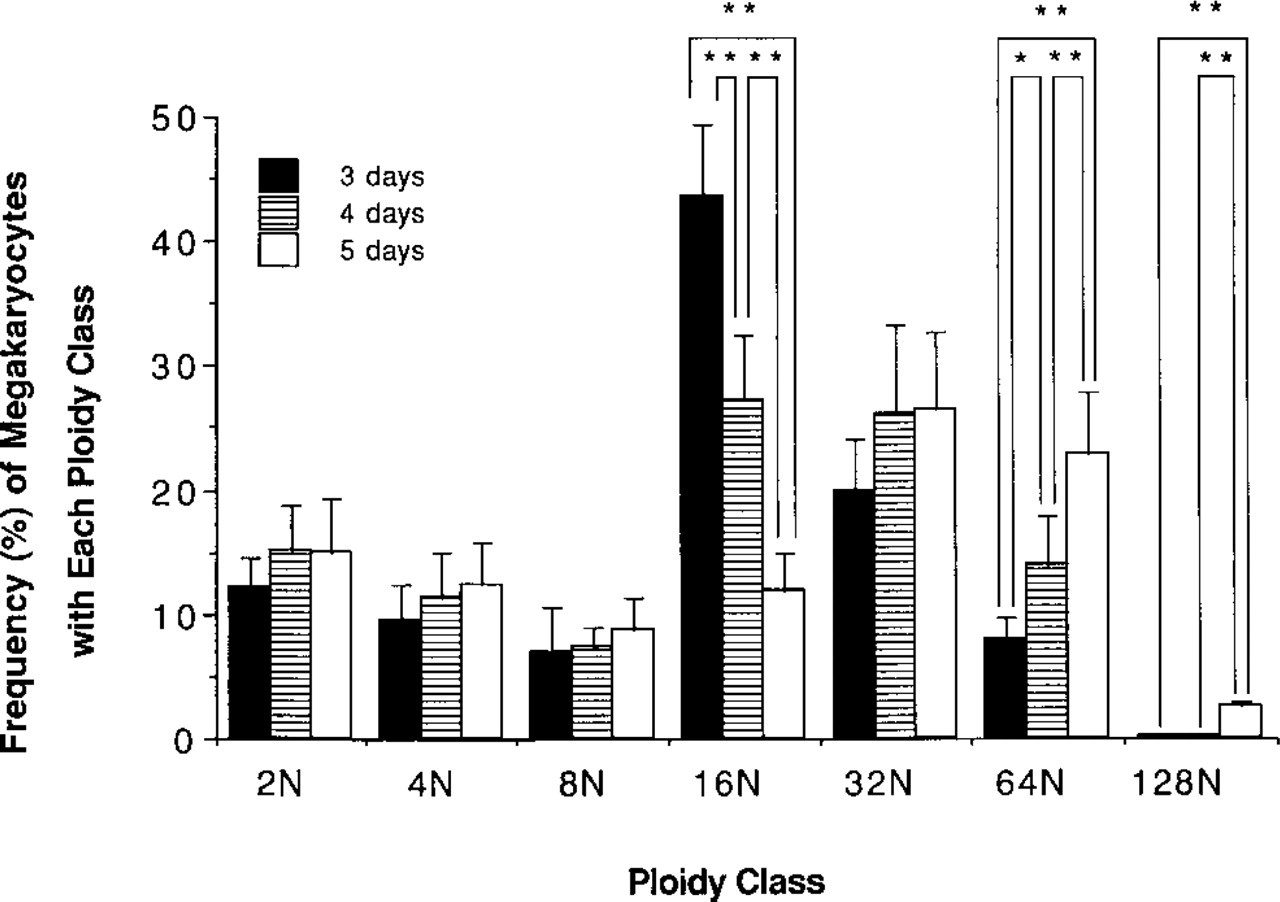
Time course of ploidy class in megakaryocytes. Nonadherent and nonphagocytic cells were cultured with 20 ng/ml of rh c-mpl ligand. ∗ Significantly increased or decreased (p<0.05); ∗∗ (p<0.01).
GP IIIa (CD 61) expression of murine megakaryocytes cultured for 4 days. a

Nonadherent and nonphagocytic cells were cultured for 4 days. The mean FITC fluorescence of the cultured megakaryocytes with each ploidy class, stimulated by mpl ligand (20 ng/ml), is expressed as 100%. Data are expressed as % (±SD) of four independent experiments. Figures in parentheses are the mean of the number of megakaryocytes examined for CD 61 expression analysis at each ploidy class.
Significantly increased (p<0.01).
Ploidy distribution of murine megakaryoctyes cultured for 4 days a
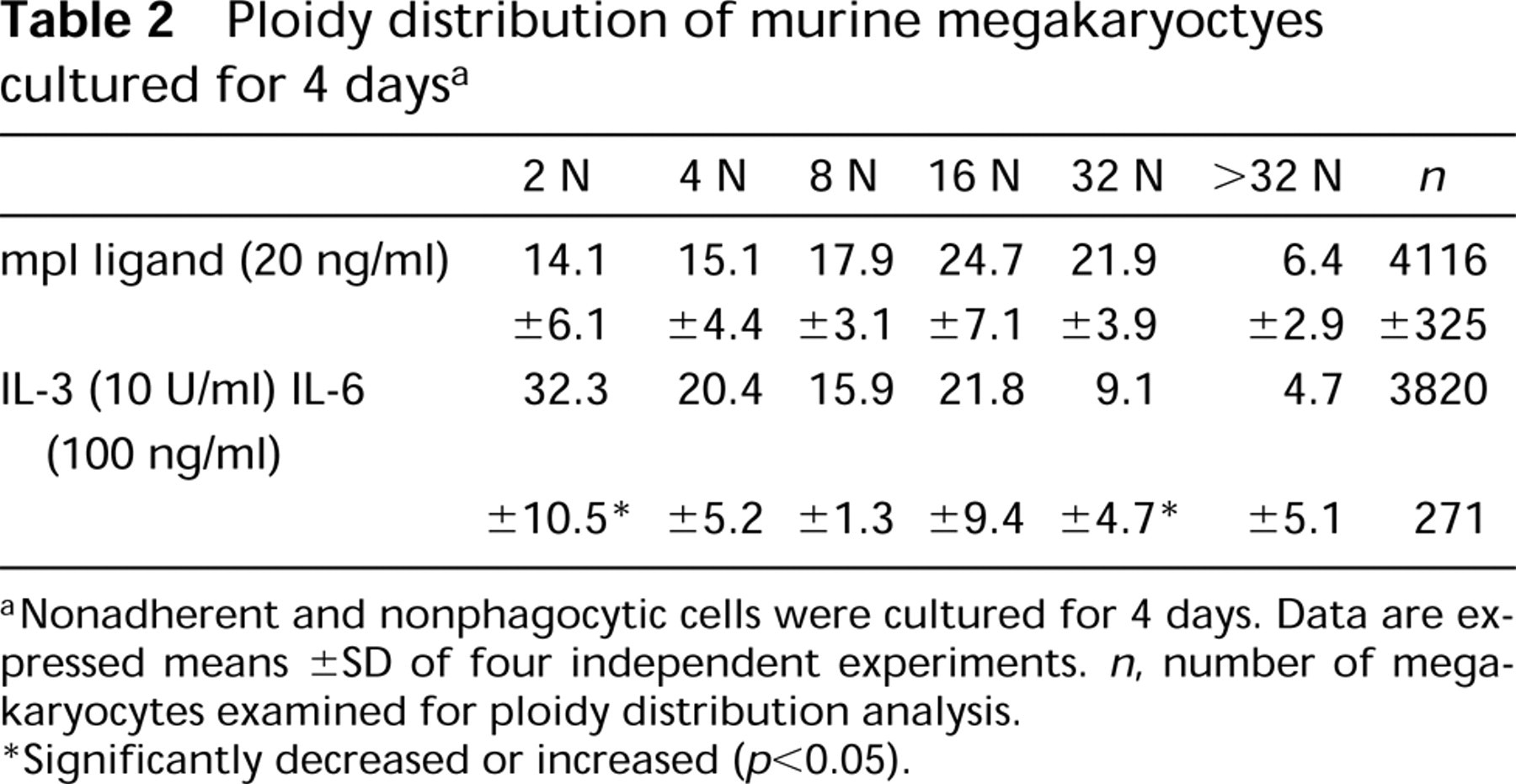
Nonadherent and nonphagocytic cells were cultured for 4 days. Data are expressed means ± SD of four independent experiments. n, number of megakaryocytes examined for ploidy distribution analysis.
Significantly decreased or increased (p<0.05).
GP IIIa (CD 61) expression of murine megakaryocytes cultured for 3 days + 2 days a

Nonadherent and nonphagocytic cells were cultured with c-mpl ligand for 3 days. The cells were recultured for an additional 2 days. The mean FITC fluorescence intensity of the cultured megakaryocytes with each ploidy, stimulated by mpl ligand (20 ng/ml), is expressed as 100%. Data are expressed as % (means ± SD) of four independent experiments. Figures in parentheses are the mean of the number of megakaryocytes examined for CD 61 expression analysis at each ploidy class.
Significantly increased (p<0.05)
significantly decreased or increased (p<0.01).
Ploidy distribution of cultured murine megakaryocytes a
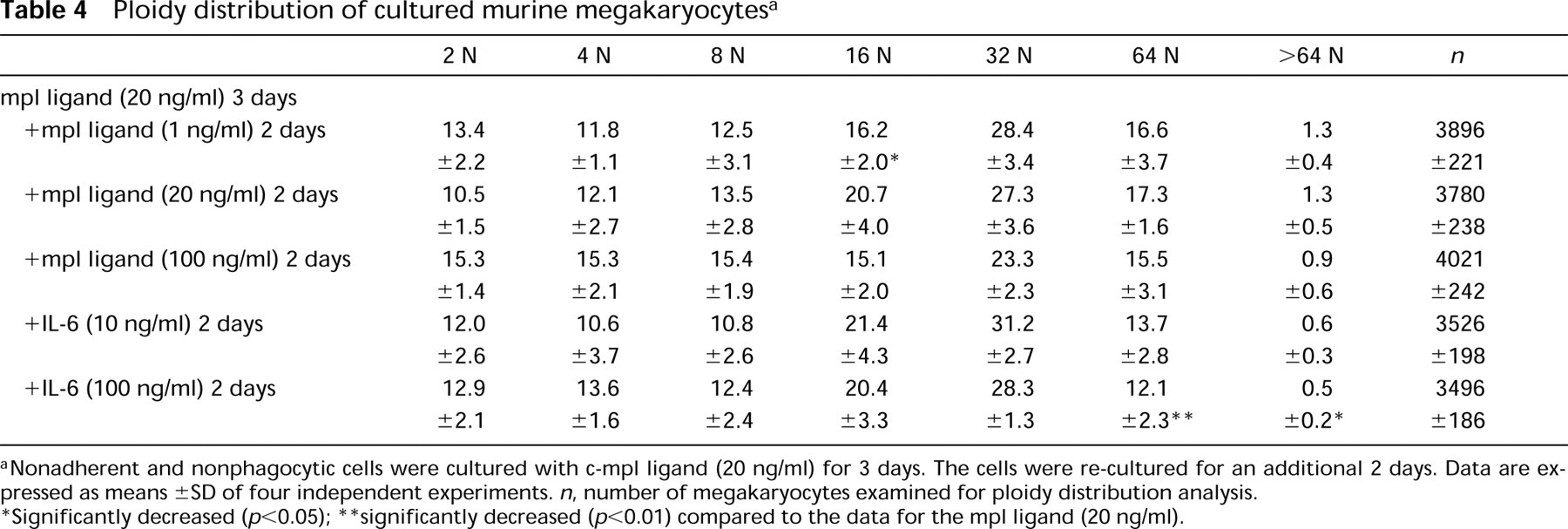
Nonadherent and nonphagocytic cells were cultured with c-mpl ligand (20 ng/ml) for 3 days. The cells were re-cultured for an additional 2 days. Data are expressed as means ± SD of four independent experiments. n, number of megakaryocytes examined for ploidy distribution analysis.
Significantly decreased (p<0.05)
significantly decreased (p<0.01) compared to the data for the mpl ligand (20 ng/ml).
These results indicate that c-mpl ligand plays a significant role in stimulating the endomitosis of megakaryocytes but a lesser role in the cytoplasmic maturation of megakaryocytic cells during differentiation and maturation, based on ploidy distribution and on AchE and CD 61 expressions.
Proplatelet Formation by Megakaryocytes
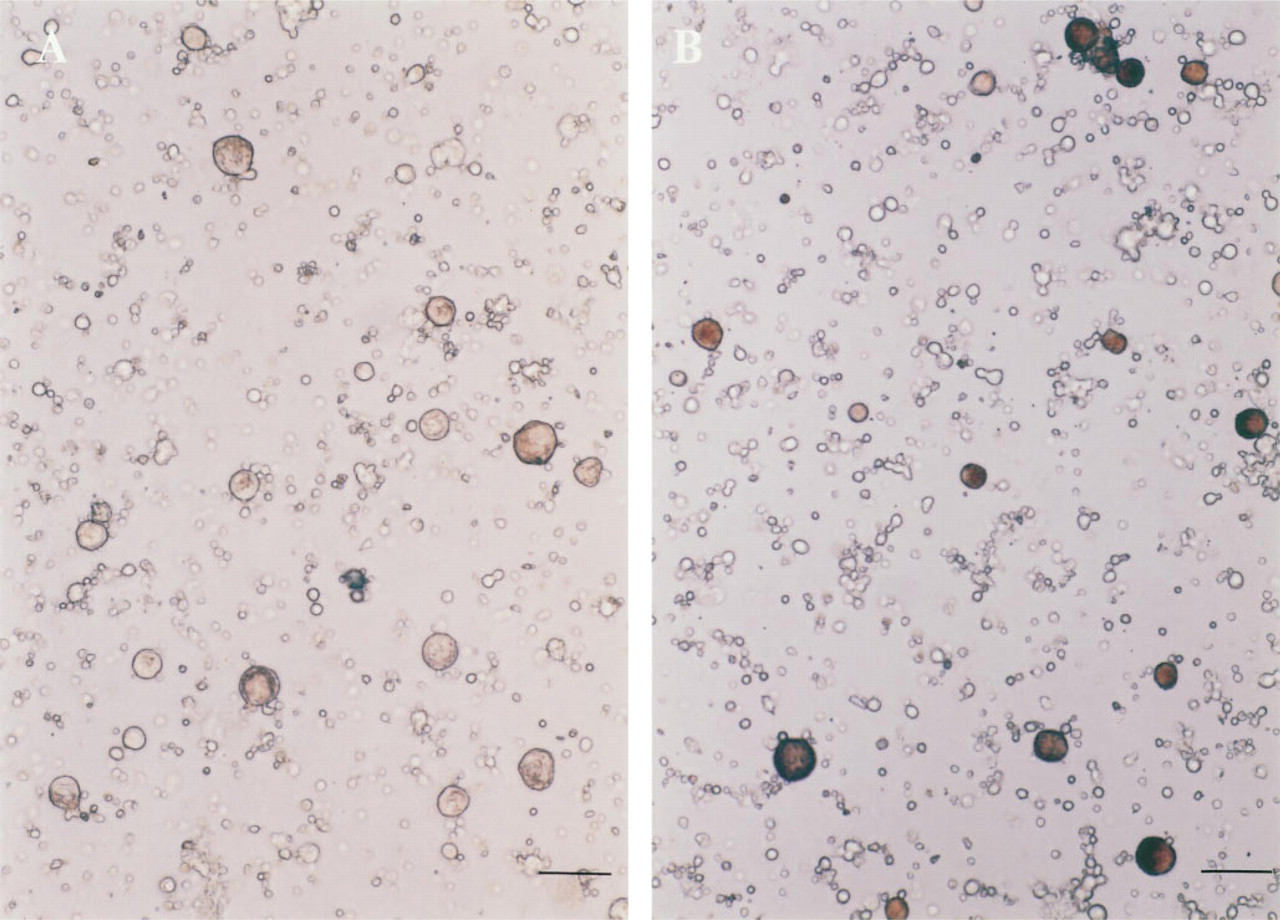
A total of 2 ± 0.3 × 104 (mean ± SD; n = 4) megakaryocytes were obtained from two mice. The purity of the megakaryocytes was 12.1 ± 2.0% n = 4), determined by AchE staining. The maturation stages of the purified megakaryocytes were estimated by the measurement of the ploidy class. More than 95% of the megakaryocytes were those with 8 N, 16 N, 32 N, and greater ploidy classes. Typical ploidy distributions of the purified megakaryocytes are shown in Figure 5. After a 24-hr incubation, the number of megakaryocytes with PPF was 17.6 ± 1.4 %, 16.0 ± 2.1%, 17.3 ± 2.3%, 7.2 ± 1.1% (p<0.01), and 22.3 ± 2.5% (p< 0.05, n = 4), stimulated by medium alone (control), rh c-mpl ligand (0.2 ng/ml), rh c-mpl ligand (2 ng/ml), rh c-mpl ligand (20 ng/ml), and rh IL-6 (50 ng/ml), respectively (Figure 6). A typical megakaryocyte with PPF stimulated by rh IL-6 (50 ng/ml) is shown in Figure 7.
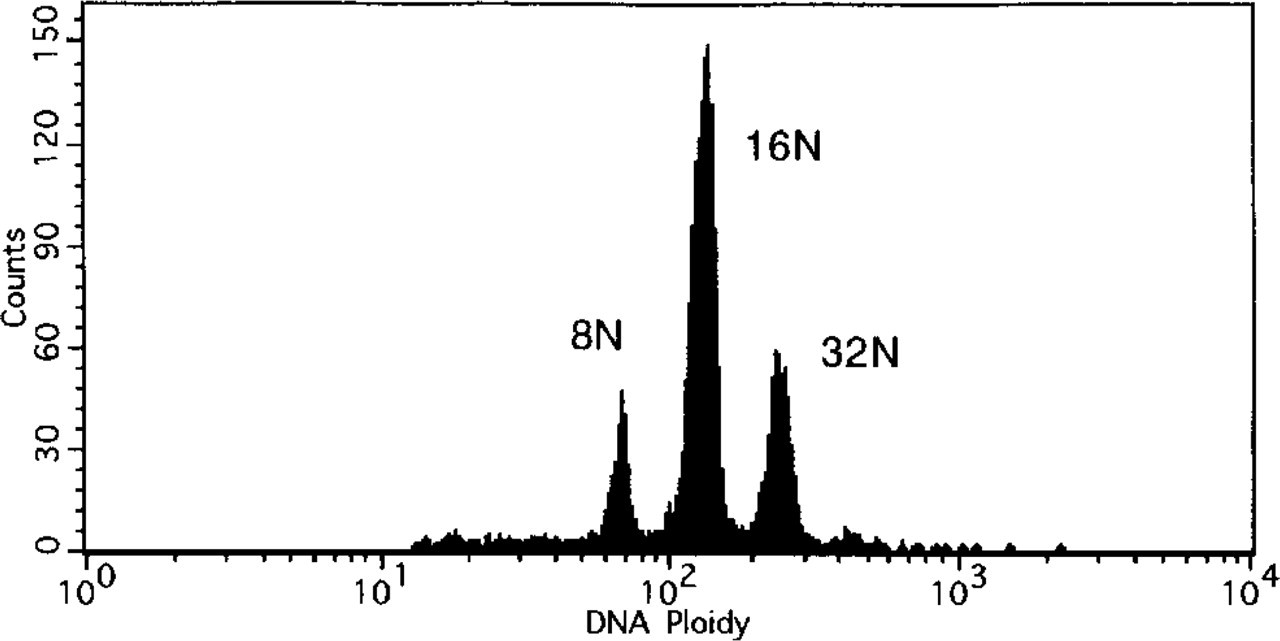
Histogram of DNA ploidy in partially purified megakaryocytes that were used for proplatelet formation experiments.
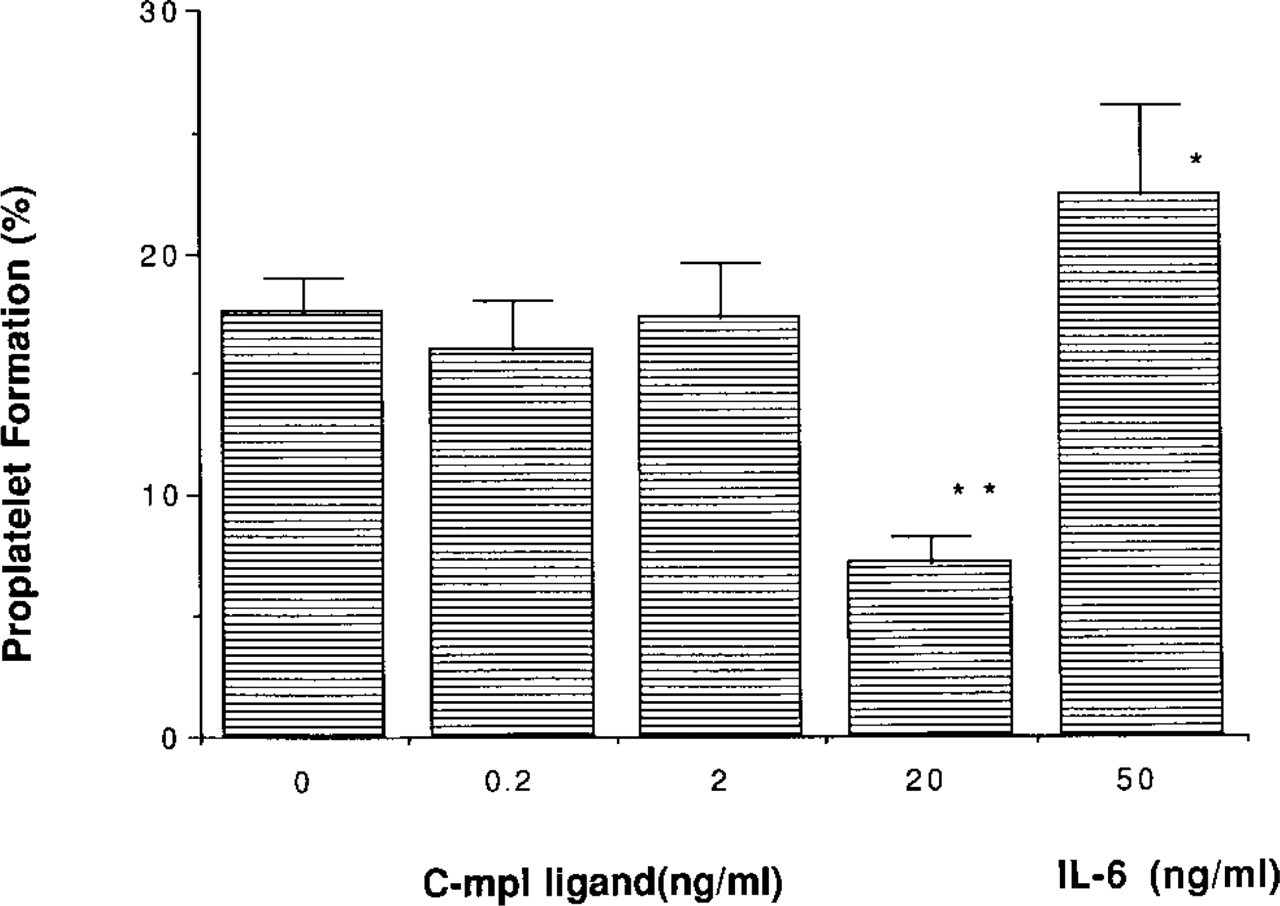
The ratio (%) of megakaryocytes with proplatelet formation, stimulated by rh c-mpl ligand or rh IL-6. ∗ Significantly increased or decreased (p<0.05); ∗∗ (p<0.01).
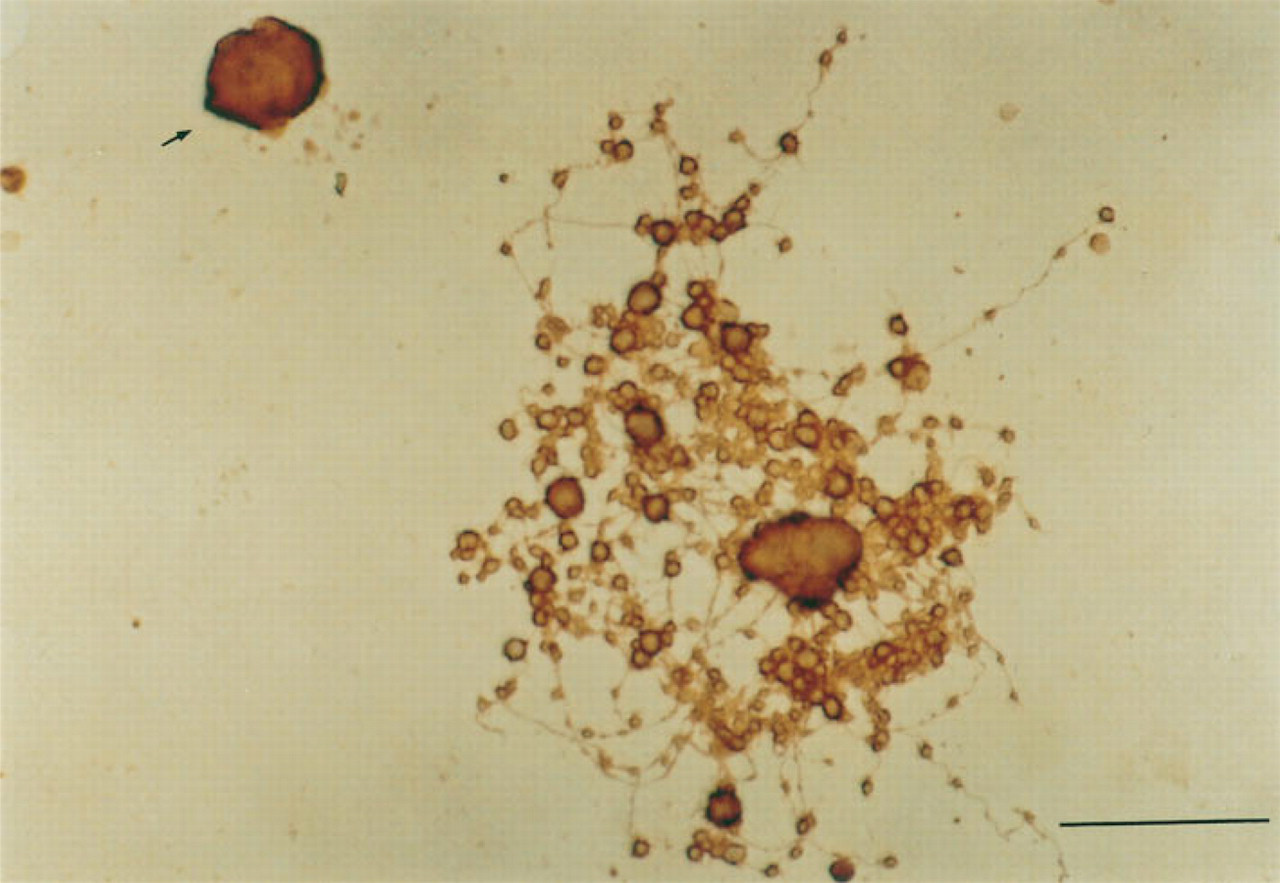
A megakaryocyte with proplatelet formation. Arrow shows a megakaryocyte without proplatelet formation. Bar = 50 μm.
Discussion
Many reports have described the biological effects of mpl ligand in vivo and in vitro (Bartley et al. 1994; de Sauvage et al., 1994; Kaushansky et al., 1994; Kuter and Rosenberg 1994; Wendling et al. 1994). Mpl ligand can act to stimulate CD 34 progenitor cells to proliferate and differentiate into megakaryocytes (Zeigler et al. 1994; Guerriero et al. 1995). In vivo administration of mpl ligand to mice induced a remarkable increase in megakaryocytic progenitor cells in bone marrow and spleen as well as thrombocytosis in peripheral blood (Kaushansky et al. 1994). Gurney et al. (1994) described mpl knockout mice with an approximately 80% decrease in the platelet count in the peripheral blood and an 85% decrease in megakaryocytes in the bone marrow. On the basis of these observation, c-mpl ligand is considered to be the physiological regulator of megakaryocytopoiesis. These results also suggested that a salvage pathway other than the mpl/mpl ligand system produces platelets. Recently, Shivdasani et al. (1995) revealed that NF-E2 knockout mice lacked circulating platelets and died of hemorrhage. Although the serum level of mpl ligand was slightly elevated above that of the normal control (Shivdasani, personal communication), electron microscopic observations of megakaryocytes in these mice revealed a striking deficiency in the number of granules and an absence of platelet regions, resulting in failure of platelet release (Shivdasani et al. 1995). This suggests that mpl ligand is not a primary regulator of cytoplasmic maturation of megakaryocytes and platelet release from megakaryocytes. Therefore, we investigated the biological effects of mpl ligand on these processes.
At first, we focused on AchE expression of the cultured megakaryocytic cells during differentiation, stimulated by mpl ligand or IL-3 + IL-6 for 3 days. AchE has been used as a differentiation/maturation marker for megakaryocytic cells during differentiation (Burstein et al. 1985; Nagasawa et al. 1988; Hill et al., 1992). Figure 1 shows that morphologically recognized megakaryocytes cultured with mpl ligand expressed less AchE than those with IL-3 + IL-6, but that larger megakaryocytes were more frequently observed in the cultures with mpl ligand than with IL-3 + IL-6. Furthermore, smaller cells than morphologically recognized megakaryocytes were more intensely stained for AchE, stimulated by IL-3 + IL-6, than those by mpl ligand. This result indicated that mpl ligand stimulated less AchE expression during differentiation from immature cells to morphologically recognizable megakaryocytes than IL-3 + IL-6 did. Kaushansky et al (1995), however, indicated that mpl ligand could induce the full development program required to produce mature platelets in electron microscopic observations of megakaryocytes. Zeigler et al. (1994) reported that murine stem cells undergo differentiation to produce megakaryocytes with proplatelet formation. However, the potential importance of serum and matrigel components in this maturation sequence in Zeigler's study (1994) was not discounted. Although we have not performed an electron microscopic study, Debili et al. (1995b) reported that a majority of the megakaryocytes showed maturation defects related to an imbalance between the synthesis of α-granules and the demarcation membrane at the ultrastructural level under serum-free conditions.
Several groups of investigators have determined the effects of mpl ligand on the expression of platelet-specific membrane proteins. Bartley et al. (1994) and Papayannopoulou et al. (1994) have detected GP Ib and GP IIb/IIIa on the surface of selected human CD34+ cells grown in the presence of rh mpl ligand and human plasma. de Sauvage et al. (1994) have shown that full-length rh mpl-ligand stimulates the formation of GP IIb/IIIa-positive cells from peripheral blood progenitors. According to their observations, mpl ligand can act on progenitor cells to differentiate into mega-karyocytic lineage cells, which express megakaryo-cyte/platelet-specific proteins GP Ib or GP IIb/IIIa. In their experiments, however, GP Ib or GP IIb/IIIa expression, measured by flow cytometry, was evaluated qualitatively but not quantitatively. We established a means of quantifying the intensity of CD 61 expression of megakaryocytes with various ploidy classes quantitatively (Figure 2). Similar analysis was performed to estimate granulocyte colony-stimulating factor (G-CSF) receptor expression in human or murine cells (Shinjo et al. 1995). To confirm the appropriateness of this procedure, murine bone marrow megakaryocytes were analyzed by this method. A linear relationship was obtained between mean fluorescence [CD 61 (GP IIIa) expression] and ploidy class in normal murine bone marrow megakaryocytes (Figure 2). Using this method, CD 61 (GP IIIa) expression was evaluated in cultured megakaryocytes. CD 61 expression was stimulated to a lesser extent during differentiation and maturation of the immature cells to mature megakaryocytes by c-mpl ligand than that stimulated by IL-3 + IL-6 under our conditions (Table 1). IL-3 has been recognized as a megakaryocyte colony-stimulating factor (Meg-CSF) (Ihle et al. 1983; Williams et al. 1985), and IL-6 as a megakaryocyte differentiation factor (Ishibashi et al. 1989; Kimura et al. 1990). Although c-mpl ligand has been reported to have both activities (de Sauvage et al., 1994; Kaushansky et al. 1994; Lok et al., 1994; Wendling et al., 1994; Zeigler et al., 1994; Kaushansky 1995), it had less differentiation activity in regard to GP IIIa expression than IL-6, based on the results obtained from murine bone marrow cells cultured for 4 days (Table 1). We also investigated the maturation effect of c-mpl ligand on cultured mature megakaryocytes. Murine nonadherent and nonphagocytic cells in bone marrow were cultured with c-mpl ligand for 3 days to form morphologically recognizable mature megakaryocytes. CD 61 expression after 2 additional days in culture with c-mpl ligand or IL-6 was evaluated using the same flow cytometric procedure. Mpl ligand stimulated the CD 61 expression of mature megakaryocytes less than IL-6 (Table 3). On the basis of expression of membrane protein, CD 61, and the cytoplasmic protein AchE, our results support the above hypothesis that c-mpl ligand does not play important roles in the megakaryocyte cytoplasmic maturation process during differentiation and maturation (Zeigler et al. 1994; Debili et al. 1995b).
The mpl ligand stimulated DNA development in the megakaryocytes cultured with mpl ligand for 3, 4, and 5 days (Tables 2 and 4). Megakaryocytes of the 64 N ploidy class tended to be increased in numbers among the cultured megakaryocytes stimulated by mpl ligand (20 ng/ml) for 3 days and 2 additional days with mpl ligand (Table 4). A significant difference were not observed between the mpl ligand and the IL-6 group except for the IL-6 (100 ng/ml) group (Table 4), probably due to the biological effect of IL-6, which stimulates an increase in ploidy class of megakaryocytes in vitro (Ishibashi et al. 1989; Kimura et al. 1990). However, all studies, including ours, support the notion that c-mpl ligand stimulates endomitosis in vitro and in vivo (Kaushansky et al. 1994; Broudy et al. 1995).
In addition, we investigated the stimulatory activity of c-mpl ligand on proplatelet formation (PPF) by megakaryocytes. PPF is considered to be the process of functional platelet production in vitro (Figure 7). We found that c-mpl ligand did not stimulate the PPF process. However, other investigators have reported results to the contrary (Miyazaki et al. 1994; Zeigler et al. 1994). They did not use mature megakaryocytes obtained directly from bone marrow cells but used cultured megakaryocytes derived from progenitor cells in bone marrow cells. Because megakaryocyte lineage cells produce cytokines such as IL-6 during development (Navarro et al. 1991), the finding that c-mpl ligand stimulated the PPF process might be due to a synergistic effect with these cytokines. Furthermore, they included plasma or serum in their culture media, so a synergistic effect of some factors in the plasma or serum with c-mpl ligand might have produced PPF activity. Choi et al. (1995) reported no effects of c-mpl ligand on megakaryocyte PPF using soluble mpl. Our results are consistent with their finding in megakaryocyte PPF.
Recently, de Sauvage et al. (1996) concluded that c-mpl ligand was not critical to the final step of platelet production, based on the findings from c-mpl ligand knock-out mice. In addition, treating wild-type mice with pharmacological doses of mpl ligand caused a 20-fold increase in megakaryocytes but only a fourfold increase in platelets (Kaushansky et al., 1994; Lok et al. 1994), suggesting that an other mechanism(s) or factor(s) may be required for platelet formation. More recently, Emmons et al. (1996) reported that the serum mpl ligand level appeared to be regulated by megakaryocyte mass rather than by the circulating platelet level, based on the human mpl ligand levels in patients with thrombocytopenia. Their results support the hypothesis that c-mpl ligand does play a physiological role in megakaryocytopoiesis but not in thrombocytopoiesis, which is consistent with the results of this study.
In summary, mpl ligand stimulates the endomitosis of megakaryocytes but not cytoplasmic maturation as IL-6 does. In addition, it does not stimulate the PPF process in vitro. On the basis of these data, c-mpl ligand is not likely to be involved in the process of platelet production.