
Abstract
Several isoforms of superoxide dismutase (SOD), including copper/zinc (cytosolic) and manganese (mitochondrial), exist. In the human placenta, SOD may prevent excessive superoxide accumulation and any potential deleterious oxidative effects. In pre-eclampsia, increased levels of lipid peroxide and decreased SOD activity have been described in the placenta. Oxidative stress such as occurs in pre-eclampsia can alter expression of SOD isoforms. The objective of this study was to localize the copper/zinc and manganese SOD isoforms in the placenta using immunohistochemistry and to compare localization and intensity of immunostaining in tissues from normotensive pregnancies with those from pregnancies complicated by pre-eclampsia and/or intrauterine growth restriction (IUGR). Western blotting with specific antibodies recognized a 17-kD copper/zinc and a 23-kD manganese SOD subunit in placental homogenates. Intense immunostaining for the manganese SOD isoform was seen in villous vascular endothelium, but only faint staining was found in the syncytiotrophoblast or villous stroma. In serial sections, intense immunostaining for copper/zinc SOD was seen in certain cells of the villous stroma but only faint immunostaining in syncytiotrophoblast and vascular endothelium. No apparent differences in localization or intensity of immunostaining for either isoform were seen between tissues of normotensive or pre-eclamptic pregnancies, with or without IUGR. The different cellular localizations of the SOD isoforms suggest that they fulfill different functional roles within the placenta.
I
The action of superoxide is normally limited by its low lipid solubility, its limited membrane transport, and also by its removal by SOD, the rate constant of which is 2 × 109 M-1s-1 (Huie and Padmaja 1993). However, macrophages, epithelial, endothelial, and interstitial cells can be induced to simultaneously produce both NO and superoxide at high concentrations in a localized manner by a variety of inflammatory stimuli, such as sepsis and ischemia/reperfusion injury. NO and superoxide then react to produce peroxynitrite, a potent long-lived oxidant, which is a major product of activated macrophages. The rate constant for this reaction is 6.7 × 109 M-1s-1 (Beckman and Tsia 1994), which suggests that as superoxide production increases, peroxynitrite formation will be favored over superoxide removal by SOD, because NO is the only known biological molecule that is produced in high enough concentrations and reacts rapidly enough to outcompete SOD for superoxide. In the perfused human placental cotyledon, generation of superoxide by co-infusion of purine and xanthine oxidase into the fetal placental vasculature surprisingly gave a vasodilator effect (Holcberg et al. 1995a). This we deduced to be caused by the formation of peroxynitrite through the interaction of NO and superoxide, because the effect was inhibited by treatment with competitive NO synthase inhibitors. Interestingly, peroxynitrite has also been shown to act acutely as a vasodilator of the dog aorta (Liu et al. 1994) and of the isolated perfused rat heart (Villa et al. 1994). Hence, we believe that peroxynitrite generated from NO and superoxide was responsible for the acute vasodilatation of the preconstricted placental vasculature (Holcberg et al. 1995a).
SOD exists in several isoforms, including the cellular copper/zinc (cytosolic, 32 kD) and the manganese (mitochondrial, 80 kD) isoforms (Fridovich 1975), and an extracellular isoform (Marklund et al. 1982). These isoforms show no sequence homology with each other. SOD activity has been described in the human placenta, and normally placental lipid peroxides decrease but SOD activity increases throughout gestation (Sekiba and Yoshioka 1979; Takehara et al. 1990) presumably to prevent excessive superoxide accumulation and consequential deleterious effects in the placenta. In placentas of preeclamptic pregnancies, increased levels of lipid peroxides are seen (Wang et al. 1992), and decreased SOD activity has been described in patients with pregnancy-induced hypertension (Wisdom et al. 1991; Chen et al. 1994). This decreased activity of SOD in pregnancy-induced hypertension may be associated with increased superoxide production and lipid peroxidation. As a prelude to measurement of placental SOD activities, we determined to identify and localize the isoforms of SOD expressed in the human placenta. This is important because regional variation of SOD activities has been shown at different sampling sites in the placenta (Jendryczko et al. 1991), and both copper/zinc (Cu/Zn) and manganese (Mn) isoforms of SOD may be present in the placenta. There is also evidence that oxidative stress, such as is postulated to occur in pre-eclampsia, can alter the expression of SOD isoforms (Shull et al. 1991). Therefore, we used immunohistochemistry with specific antibodies against the copper/zinc and manganese SOD to localize these isoforms in tissue from normotensive pregnancies and compared localization and intensity of immunostaining at the light microscopic level in normotensive tissues with that from pregnancies complicated by preeclampsia and/or intrauterine growth restriction.
Materials and Methods
All procedures were performed under protocols approved by the Institutional Review Board of the University of Cincinnati Medical Center and the Ethical Committee of Glasgow Royal Infirmary. Placental villous tissue (1 × 1 × 1 cm) was collected immediately after delivery from normotensive pregnancies (n = 5) and from pregnancies complicated by either preeclampsia (n = 5), intrauterine growth restriction (IUGR; n = 5), or preeclampsia plus IUGR (n = 4). Villous tissue was then immediately flash-frozen in liquid nitrogen and stored at −70C until processed. Preeclampsia is defined as a blood pressure of 140/90 mmHg on at least two occasions at least 6 hr apart occurring after Week 20 of gestation and accompanied by proteinuria (>300 mg/liter in a 24-hr urine collection) and/or edema. IUGR is defined as a fetal weight less than the fifth percentile using standardized Scottish birthweight tables. The gestational ages of the patients were 30.9 ±1.7 weeks (normotensive), 35.4 ± 2.6 weeks (pre-eclampsia), 35.5 ± 1.3 weeks (IUGR), and 30.5 ±1.9 weeks (preeclampsia plus IUGR). Because all pathological tissues were pre term, the normotensive group was age-matched to the group with earliest gestation.
Serial sections of villous tissue were cut at 7 μ and immunostained using a monoclonal antibody to Cu/Zn SOD (Sigma; St Louis, MO) or a polyclonal antibody to Mn SOD (Fitzgerald; Concord, MA) with the Vectastain ABC Elite Kit (Vector Laboratories; Burlingame, CA) using DAB reagent. These tissues were incubated in primary antibody for 1.5 hr at 37C, followed by incubation in the secondary antibody. Sections were then blocked for endogenous peroxidase for 3 min using 3% H2O2 in water. Saponin (0.1%) was included in all washes and antibody solutions up to and including the secondary antibody incubation, after which it was excluded from all reagent mixtures. Sections were finally counterstained with hematoxylin and mounted in PBS/glycerol (1:9). The control incubations contained no primary antibody. All sections were then examined by one observer who was blinded to tissue identity, and the localization and intensity of immunostaining for either the Cu/Zn or the Mn SOD were noted.
For Western blot analysis, fresh placental villous tissue was homogenized in 0.32 M sucrose, 20 mM HEPES, 1 mM EDTA, 1 mM DTT, pH 7.8, with a polytron, three times for 10 sec on ice. The homogenates were centrifuged at 1000 × g for 15 min to remove debris, and the protein concentration of the supernatant was determined. Five μg total protein was loaded on 15% polyacrylamide gels (1 mm × 10 cm × 10 cm). Proteins were separated at 20 mAmps/plate at 4C and transferred to nitrocellulose membranes using a BioRad transblot apparatus (BioRad; Melville, NY). Membrane blots were blocked with 5% nonfat dry milk in 0.1% Tween-20 in Tris/saline buffer [50 mM Tris-HCl, pH 7.5, 500 mM NaCl (TTBS)] and incubated in either monoclonal anti-human Cu/Zn SOD antibody 1:250 (IgG1, ascites fluid; Sigma Immunochemicals, St Louis, MO) or polyclonal sheep anti-human Mn SOD, 1:100 (IgG fraction, preabsorbed with human tissue and serum polymers; Calbiochem-Novabiochem International, San Diego, CA) in TTBS for 1 hr at room temperature. Subsequently, membranes were treated with biotin-conjugated goat anti-mouse IgG, 1:1000 (Sigma ImmunoChemicals) or biotin-conjugated rabbit anti-sheep IgG 1:200 (Vector Laboratories) containing 1% goat serum or 1% rabbit serum, respectively. Protein bands were detected with the Vectastain ABC and DAB Peroxidase Substrate kits (Vector Laboratories). Washes in between treatment steps consisted of two quick rinses followed by three washes of 10 min each in TTBS.
Results
Examination of placental villous tissue taken from a normotensive term pregnancy (Figure 1) showed intense immunostaining for the Mn SOD isoform in the endothelium of the villous vasculature (Figure 1A) but only faint staining for Mn SOD in the syncytiotrophoblast and in the stromal cells of the villous core. This was in contrast to immunostaining for Cu/Zn SOD in the same tissue, where on a consecutive section (Figure 1B), intense immunostaining was observed in certain cells of the villous stroma. Faint positive immunostaining for Cu/Zn SOD was also seen in syncytiotrophoblast and vascular endothelium. However in vascular endothelium the intensity of Cu/Zn SOD immunostaining was much less than that of the Mn SOD isoform. In the absence of either primary antibody (Figure 1C), no immunostaining was apparent on the consecutive section of this or any other sample.
Examination of placental villous tissue taken from pregnancies complicated by either pre-eclampsia, IUGR, or pre-eclampsia plus IUGR showed that there were no apparent differences in localization and intensity of immunostaining for either Mn SOD or Cu/Zn SOD compared to tissues from normotensive pregnancies. Therefore, immunostaining for Mn SOD in the pre-eclamptic (Figure 2A) IUGR (Figure 2C), or pre-eclampsia plus IUGR (Figure 2E) tissue was still seen most intensely in the villous vascular endothelium and faintly in the syncytiotrophoblast and stroma. Moreover, the Cu/Zn isoform of SOD in the pre-eclampsia (Figure 2B) and IUGR (Figure 2D) samples, or in the pre-eclampsia plus IUGR (Figure 2F) samples, was seen again primarily most intensely in certain villous stromal cells, with weaker positive immunostaining in syncytiotrophoblast and vascular endothelium. In these pathological tissues, weak diffuse immunostaining for Cu/Zn SOD was also seen throughout the stroma. Again, in the absence of primary antibody (data not shown) no immunostaining was seen in the villous tissue from any of these pathological pregnancies. On Western blots (Figure 3), single immunostained bands of 17 kD (Cu/Zn SOD) and 23 kD (Mn SOD) were recognized by the SOD antibodies.
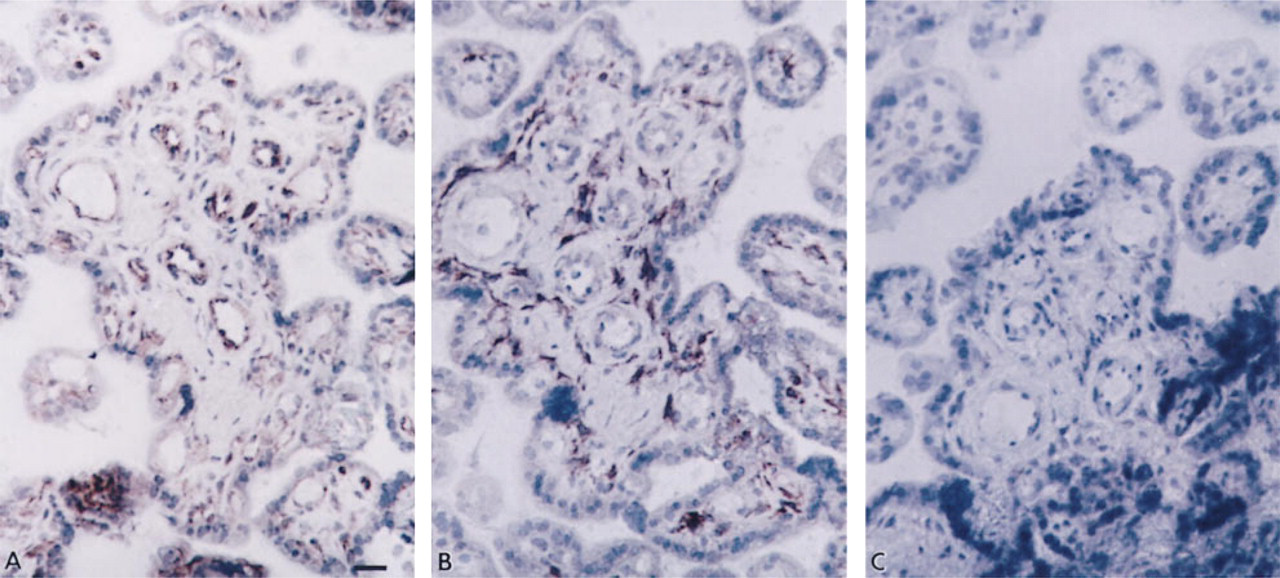
SOD isoforms in placental villous tissue of term normotensive pregnancy. Consecutive sections immunostained with (
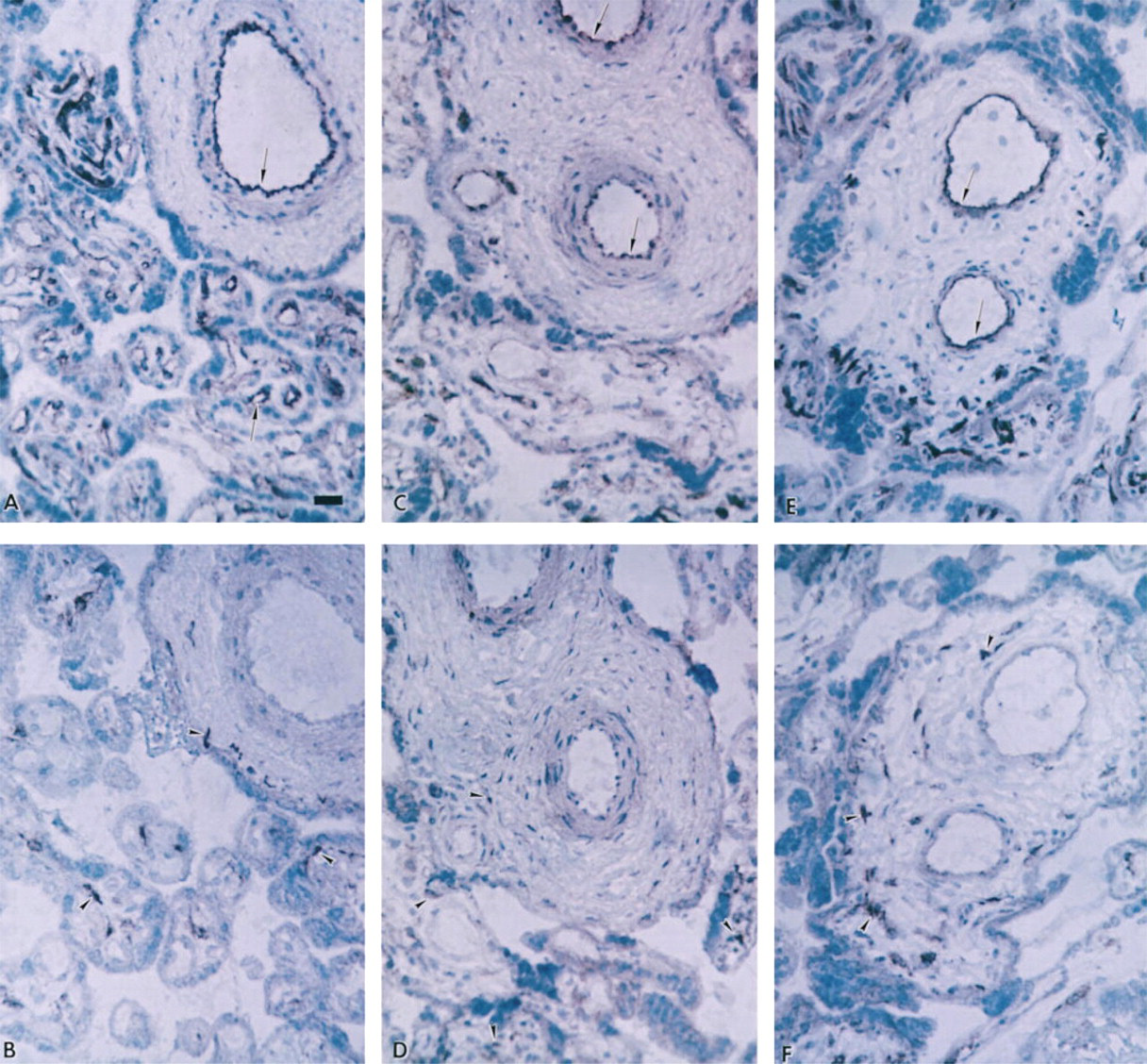
SOD isoforms in placental villous tissue of pregnancies complicated by pre-eclampsia, IUGR, or pre-eclampsia plus IUGR. Consecutive sections from villous tissue of pre-eclampsia (
Discussion
Using specific antibodies to both the Cu/Zn and Mn SOD isoforms, we have demonstrated the presence of both isoforms in human villous tissue by immunohistochemistry and Western blotting. It is apparent that the two isoforms demonstrate different cellular localizations within the placenta, which suggests that they may fulfill different physiological roles. The Mn SOD isoform appears to be mainly localized to fetal vascular endothelium, suggesting that it may play a role in scavenging superoxide at the level of the endothelium or elsewhere in the vasculature, thus preventing deleterious effects of superoxide on the vasculature. However, it may also play a crucial role in controlling the half-life of endothelium-derived NO (via regulation of superoxide concentrations) and thus in regulating vascular reactivity. Indeed, our previous studies, in which infusion of SOD into the fetal placental vasculature of the perfused placental cotyledon resulted in vasodilatation (Holcberg et al. 1995b), support this viewpoint. The relative activity of Mn SOD in the fetal vascular endothelium and hence its ability to scavenge superoxide may also play a role in the formation of peroxynitrite anion, which is formed from the interaction of NO and superoxide. The formation and presence of nitrotyrosine residues have been used as an index of peroxynitrite anion formation (Beckman et al. 1994). Nitrotyrosine residues have been demonstrated in atherosclerotic plaques (Beckman et al. 1994) and in lung sections of patients and animals with acute lung injury (Haddad et al. 1994), indicating areas of cell damage. In the villous tissue of normotensive pregnancies, we find few or no nitrotyrosine residues in the villous vasculature, which suggests that very little peroxynitrite is formed and which indicates that superoxide is effectively scavenged by SOD. However, in villous tissue of pregnancies complicated by pre-eclampsia with or without intrauterine growth restriction (Myatt et al. 1996), we find evidence of nitrotyrosine residues in both the fetal vascular endothelium and in the surrounding smooth muscle and villous stroma. This suggests that, in these pregnancies, there is increased production of peroxynitrite in the fetal placental vasculature, which leads to the formation of nitrotyrosine residues. We have also shown increased expression of the endothelial isoform of NO synthase in the fetal placental vasculature of pre-eclamptic pregnancies (Myatt et al. 1997), which we postulate may be an adaptive response to the increased fetal placental resistance characteristic of such pregnancies (Trudinger et al. 1985). In concordance with this apparent upregulation of endothelial NO synthase isoform, we also find increased concentrations of nitrate, the breakdown product of NO, in the umbilical circulation of pregnancies complicated by pre-eclampsia (Lyall et al. 1995) or intrauterine growth restriction (Lyall et al. 1996). Therefore, increased endothelial NO synthase expression may lead to increased NO formation, which may facilitate peroxynitrite production. However, we do not find any apparent difference at the light microscopic level in the intensity of staining for Mn SOD in the vascular endothelium when villous tissue of normotensive pregnancies are compared with those complicated by pre-eclampsia, with or without intrauterine growth restriction. Therefore, we cannot postulate that increased superoxide concentrations may arise in the vasculature due to decreased scavenging by SOD, although it remains to be established whether or not immunoreactivity correlates with enzyme activity.
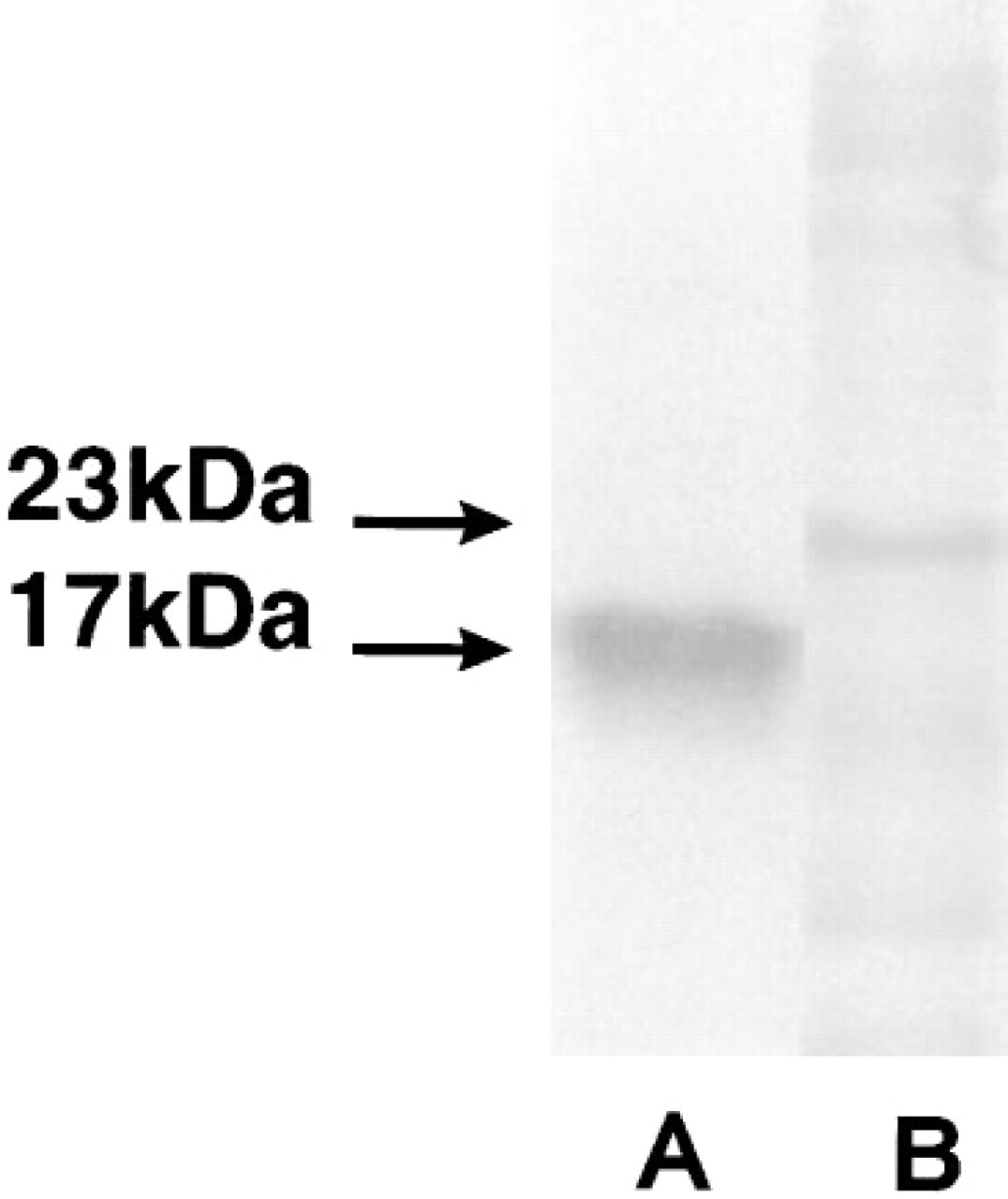
Western blot analysis of human placental villous tissue homogenate. Distinct bands were observed after labeling with monoclonal anti-human Cu/Zn SOD (Lane A) and monospecific polyclonal anti-human Mn SOD (Lane B) antibodies. The molecular weight of the labeled products is based on BioRad Kaleidoscope Prestained Standards.
In contrast to the Mn SOD isoform, the Cu/Zn SOD isoform is localized mainly in the villous stromal tissue. At this location, Cu/Zn SOD may protect the villous tissue against the deleterious effects of super-oxide generated by fetal macrophages (Hofbauer cells) or from other sources within villous tissue. Interestingly, neither the Mn SOD nor the Cu/Zn SOD isoform appeared to be present in syncytiotrophoblasts at the intensity found in the vascular endothelium and the stroma. Although this may be a relative difference in expression, this finding is surprising, given that Church et al. (1992) found that in vitro differentiation of cytotrophoblasts to syncytiotrophoblasts was associated with transient expression of Mn SOD. Increased Mn SOD gene expression or enzyme activity appeared to precede or to be coordinated with morphological and biochemical trophoblast differentiation, and hence may be a marker for cell differentiation. There are other examples in which cell transformation is associated with a loss in SOD activity (Allen 1991) and in which administration of SOD induces cellular differentiation. Because reactive oxygen species, including superoxide, may function as cellular second messengers involved in signal transduction processes, the SOD enzymes may therefore have an important regulatory role in cell signaling and cell proliferation.
The recognition that the different SOD isoforms are present at different cellular locations within villous tissue cautions against gross measurements of SOD activity in whole placental homogenates in relation to different pathological states of pregnancy. Although placental SOD activity increases throughout gestation and is accompanied by a decrease in placental lipid peroxide concentrations (Sekiba and Yoshioka 1979; Takehara et al. 1990), changes in villous volume, trophoblast surface area, and the degree of vascularization that may occur in pathological pregnancies, such as pre-eclampsia or intrauterine growth restriction, make it difficult to be able to draw meaningful comparisons between SOD activities in whole placental homogenate from normotensive and pathological pregnancies. Furthermore, oxidant stress, which may occur regionally in the placenta, can alter the expression of the SOD isoforms (Shull et al. 1991).
Footnotes
Acknowledgements
Supported by grants from NIH (HL47860; LM) and Action Research and by the Scottish Hospital Endowments Research Trust (FL, IAG).