
Abstract
Many studies have examined the localization of gangliosides using anti-ganglioside antibodies, although widely differing conclusions have been reached. We now demonstrate that the apparent localization of gangliosides can be greatly influenced by the fixation method. Using monoclonal antibody (MAb) A2B5 (which reacts with a variety of gangliosides), hippocampal neurons were labeled at the cell surface when incubated with the antibody before fixation, but when incubated after fixation the cells displayed a variety of labeling patterns, depending on the fixation method. Biochemical analysis demonstrated that some of the fixatives (particularly acetone and methanol) significantly reduced or completely depleted cellular gangliosides, implying that the immunoreactivity observed with A2B5, and with other antibodies, was not due to gangliosides. When neurons were incubated with an anti-GD1b antibody prefixation, uniform labeling of the plasma membrane was observed, but after ganglioside depletion using biochemical inhibitors of ganglioside synthesis no cell surface labeling was detected. However, even in cells depleted of gangliosides, labeling of both the cell surface and intracellular compartments was observed when the anti-GD1b antibody was applied after fixation. Moreover, after fixation, antibodies to GM4 and GD2 reacted with hippocampal neurons, although these ganglio-sides are absent from these neurons. In contrast, the JONES antibody (which reacts with 9-O-acetylated GD3) labeled neurons with a similar pattern, essentially irrespective of the fixation method. These observations demonstrate that great care must be taken in assigning gangliosides to specific cell populations or to intracellular locations solely on the basis of use of anti-ganglioside antibodies, and suggest that optimal fixation conditions must be established for each anti-ganglioside antibody.
The sialic acid-containing glycosphingolipids, the gangliosides, may play important roles in both neuronal (Ledeen and Wu 1992) and non-neuronal (Hakomori and Igarashi 1993; Zeller and Marchase 1992) tissues. Most of the proposed functions of ganglio-sides are related to their localization at the plasma membrane, where they have been shown by biochemical analyses to be enriched (Yu and Saito 1989), although low levels of gangliosides have also been detected in intracellular membranes (Matyas and Morre 1987).
Many studies have attempted to determine the cellular localization of gangliosides using anti-ganglio-side antibodies (reviewed in Schwarz and Futerman 1996). In contrast to most glycerolipids and sphingolipids, gangliosides contain antigenic carbohydrate moieties, which has facilitated the production of a large number of polyclonal and monoclonal antibodies against all of the major and most of the minor gangliosides (Schwarz and Futerman 1996). However, immunolocalization of gangliosides has often yielded contradictory data. For example, using two different antibodies to ganglioside GD3, opposite conclusions were reached about the presence of GD3 in rat cerebellar Purkinje cells. With monoclonal antibody (MAb) R24 no labeling of Purkinje cells was observed (Goldman et al. 1984), but labeling was observed with the LB1 antibody (Reynolds and Wilkin 1988). Although these differences could be caused by each antibody recognizing different epitopes on ganglioside GD3, a more likely explanation is that the different fixation methods used in each study (Reynolds and Wilkin 1988) are responsible for the different results.
We now report that the immunolocalization of gangliosides can be greatly influenced by the fixation techniques used, at least in cultured hippocampal neurons, which contain high levels of gangliosides expressed at the cell surface (Hirschberg et al. 1996; Sofer and Futerman 1995; Harel and Futerman 1993). Incubation with anti-ganglioside antibodies before fixation (prefixation) yields reproducible patterns of immunofluorescence using all antibodies tested, whereas incubation with the antibodies after various fixation techniques (postfixation) often leads to ambiguities about the localization of a particular ganglioside. Moreover, in some cases ganglioside depletion from the tissue did not affect antibody binding. Together, these data demonstrate that optimal fixation techniques must be established for each anti-ganglioside antibody and suggest that biochemical analysis (or other non-immunohistochemical analysis) should be used, where possible, to confirm that a particular ganglioside is present in a particular tissue.
Materials and Methods
Cell Culture and Reagents
Hippocampal neurons (taken from embryonic Day 18 rats) were cultured as described for morphological (Schwarz et al. 1995; Harel and Futerman 1993; Goslin and Banker 1991) and biochemical (Hirschberg et al. 1996) analyses. FB1, an inhibitor of dihydroceramide synthase (Merrill et al. 1996) was obtained from the Division of Food Science and Technology (CSIR, Pretoria, South Africa). d-Threo-1-phenyl-2-decanoylamino-3-morpholino-1-propanol (PDMP), an inhibitor of glucosylceramide synthase (Radin and Vunnam 1981) was from Matreya (Pleasant Gap, PA). FB1 and PDMP were dissolved in Hepes buffer (20 mM, pH 7.4) and added to cultures to give a final concentration of 10 μM and 50 μM, respectively (Schwarz et al. 1995). All other solvents and chemicals were of analytical grade.
Antibodies
The following MAbs were used in this study: A2B5 (Boehringer-Mannheim; Mannheim, Germany), which reacts with a variety of polysialo-gangliosides (see Schwarz and Futeran 1996); B17 (IgM subclass) (provided by Dr. Israel Pecht, Department of Immunology, Weizmann Institute of Science), which reacts with ganglioside GD1b (Harel and Futerman 1993; Ortega et al. 1990); JONES (IgM subclass) (provided by Dr. C.J. Barnstable, Yale University Medical School), which reacts with 9-O-acetylated-GD3 (Blum and Barnstable 1987; Constantine-Paton et al. 1986); BMA190 (Behring Werke; Marburg, Germany), which reacts with GD2; R24 (IgG subclass) (Matreya), which reacts with GD3 (Pukel et al. 1982; Dippold et al. 1980). An anti-GM4 poly-clonal antibody (IgG subclass) (Matreya) was also used. Biotin-conjugated anti-rabbit or anti-mouse IgG (H + L), and indocarbocyanine (Cy3)-conjugated streptavidin, were obtained from Jackson Immunoresearch Labs (West Grove, PA).
Indirect Immunofluorescence Microscopy
Coverslips containing neurons from 0-, 3-, and 7-day-old cultures were washed in Hank's balanced salt solution (HBSS) containing 3% (w/v) bovine serum albumin for 5 min: Then the neurons were either incubated directly with primary antibodies (37C, 30 min) and then fixed with 4% formaldehyde (either freshly depolymerized from paraformaldehyde, or frozen aliquots) for 30 min at 37C, or fixed before incubation (37C, 30 min) with the primary antibodies. The following fixation methods were used: (a) 4% formaldehyde, 30 min, 37C; (b) methanol, 10 min, −20C; (c) acetone, 1 min, −20C; (d) 4% formaldehyde, 20 min, 37C, followed by acetone, 1 min, −20C. MAbs B17, JONES, and R24, and the polyclonal anti-GM4 antibody were diluted 1:50, BMA190 was diluted 1:1, and A2B5 was diluted 1:100, all in HBSS containing 3% bovine serum albumin. Biotin-conjugated anti-rabbit or anti-mouse IgG (H + L) antibodies and indocarbocyanine (Cy3)-conjugated streptavidin were used for detection. Cells were observed with a Plan Apochromat X63/1.4 NA oil objective of a Zeiss Axiovert 35 microscope with an appropriate filter for Cy3 fluorescence.
Biochemical Analysis of Ganglioside Synthesis and Content
Gangliosides were metabolically labeled by incubation with 5 × 106 cpm of [4,5-3H]-dihydrosphinganine (10 Ci/mmol) (Hirschberg et al. 1993) on Day 0 in culture. After 6 hr, unincorporated [4,5-3H]-dihydrosphinganine was removed by washing, and [3H]-sphingolipids and [3H]-gangliosides were extracted on Day 3 in culture as described (Hirschberg et al. 1996; Schwarz et al. 1995). Briefly, neurons were removed from the coverslips by scraping with a rubber policeman into water, lyophilized, and resuspended in 6 ml CHCl3/CH3OH/H2O/pyridine (60:30:6:1 v/v/v/v) at 48C for 48 hr with stirring. Phospholipids were degraded by 100 mM methanolic NaOH (2 hr, 37C). Samples were desalted by reverse-phase chromatography on an RP-18 column (Williams and McCluer 1980), and [3H]-sphingolipids and [3H]-gangliosides separated by thin layer chromatography (TLC) using CHCl3/CH3OH/aqueous 9.8 mM CaCl2 (60:35:8 v/v/v) as the developing solvent. [3H]-Sphingolipids and [3H]-gangliosides were visualized using a BAS-TR2040S imaging plate, a FUJI BAS 1000 phosphorimager, and Mac BAS v2.0 software (Fuji; Tokyo, Japan).
Results
MAb A2B5 reacts with a number of gangliosides, including GT3, 9-Oacetylated-GT3, GQ1b, GD3, GD2, GQ1c, sulfatide, and several minor gangliosides (Fredman et al. 1984; and reviewed in Schwarz and Futerman 1996), and has been widely used in immunofluorescence studies. When hippocampal neurons were incubated with A2B5 before fixation, the plasma membrane of the cell body and of the neuronal processes (both axons and dendrites) was uniformly labeled (Figures 1A and 1B). However, not all of the neurons on the coverslip were A2B5-positive (Figures 1A and 1B). When cells were first fixed with formaldehyde and then incubated with A2B5, a similar number of A2B5-positive neurons were observed, but rather than the uniform labeling obtained when neurons were incubated with the antibody before fixation (Figures 1A and 1B), a non-uniform “striped” labeling pattern was observed (Figures 1C and 1D). In contrast, glial cells, which are occasionally found on neuronal coverslips (Hirschberg et al. 1996) were labeled with a “spotty” pattern (inset to Figure 1D). The striped labeling pattern of neurons was abolished by treatment with formaldehyde and acetone (Figures 1E and 1F), which resulted in bright labeling of all neurons in the cell body and labeling of some processes. After acetone fixation alone, the cell body was brightly labeled (Figures 1G and 1H), and after methanol fixation the nuclear envelope, the nucleolus, and other organelles in the cell body were labeled (Figures 1I and 1J). Therefore, the localization of the A2B5 antigen(s) appeared to vary widely depending on the fixation method. No labeling was observed with biotin-conjugated anti-rabbit or anti-mouse IgG (H + L) antibodies or with indocarbocyanine (Cy3)-conjugated streptavidin in the absence of primary antibodies.
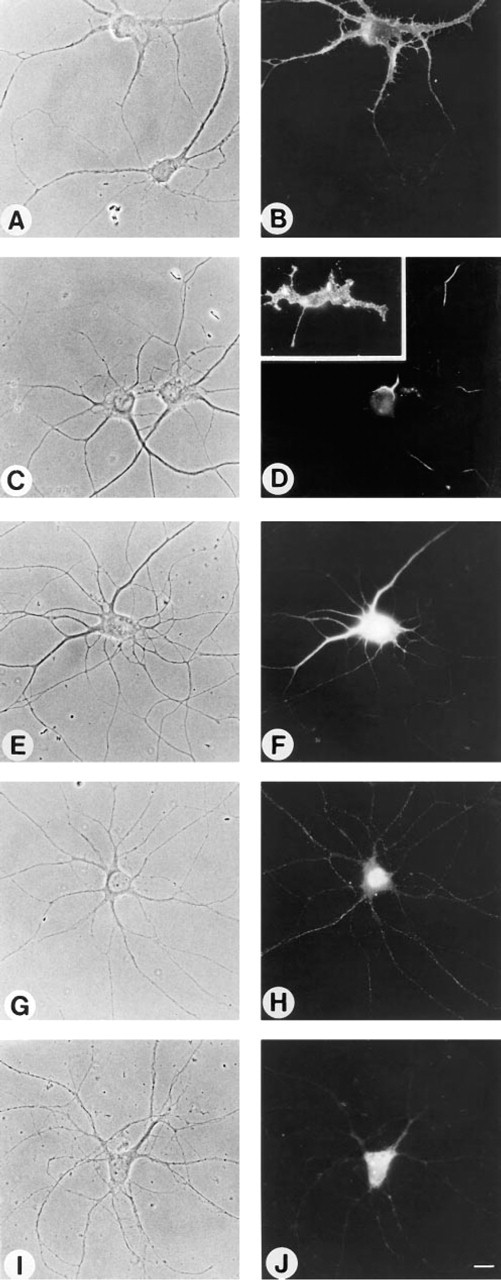
Effect of fixation methods on binding of MAb A2B5 to 7-day-old cultures of hippocampal neurons. Neurons were either fixed with formaldehyde after incubation with antibody A2B5 (
To analyze whether the labeling observed after each fixation method was due to endogenous gangliosides, neurons were metabolically labeled with [4,5-3H]-dihydrosphinganine to label sphingolipids and gangliosides, and then fixed by the four methods used above (Figure 1). To ensure no loss of cellular sphingolipids and gangliosides, neurons were scraped directly into distilled water and then lyophilized (Hirschberg et al. 1996). After methanol treatment, no [3H]-gangliosides remained associated with the neurons (Figure 2), indicating that the labeling pattern observed with A2B5 (Figures 1I and 1J) was not due to binding of A2B5 to cellular gangliosides. Likewise, after acetone treatment, the level of cell-associated [3H]-gangliosides was reduced by 56% compared to control cells (Figure 2), and after treatment with formaldehyde and acetone, [3H]-ganglioside levels were reduced by 46%, implying that the labeling observed with A2B5 after these fixation methods may also not be due to binding of A2B5 to cellular gangliosides. In contrast, formaldehyde fixation alone reduced levels of [3H]-gangliosides only by 10% (Figure 2).
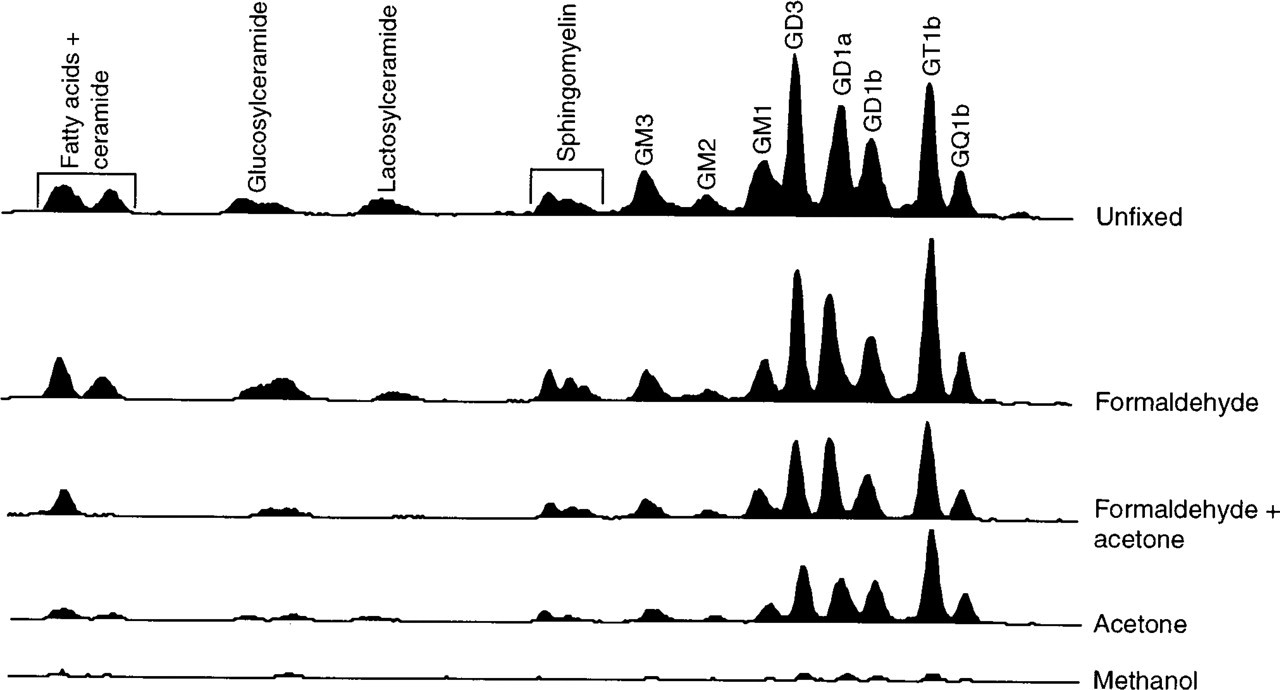
TLC analysis of cell-associated [3H]-sphingolipids and [3H]-gangliosides after various fixation methods. The origin of the TLC plate is on the right; the identity of [3H]-sphingolipids and [3H]-gangliosides was determined using authentic standards.
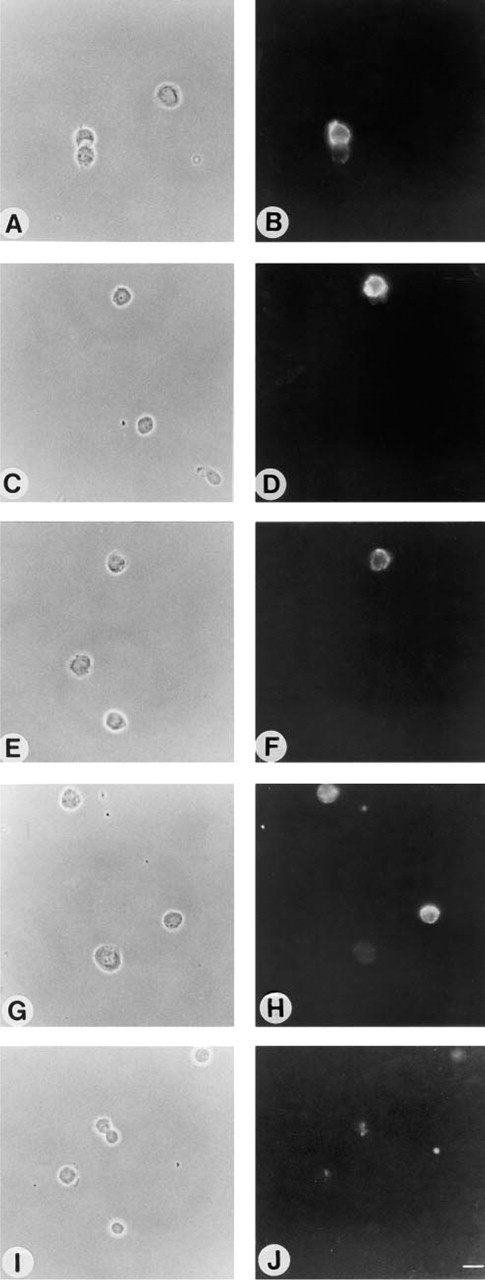
We next compared the binding of a highly specific MAb, B17 (which binds to ganglioside GD1b), to hippocampal neurons after various fixation methods. When 3-day-old neurons were incubated with B17 before fixation, uniform labeling of the cell surface was observed (Figures 3A and 3B). However, after incubation with either PDMP (Figures 3C and 3D) or FB1 (Figures 4A and 4B), both of which inhibit ganglioside synthesis (Futerman 1994), no labeling of the cell surface was observed even when neurons were incubated with B17 before fixation. This is consistent with previous studies demonstrating that ganglioside GD1b could not be detected by biochemical or immunohistochemical analysis after inhibition of ganglioside synthesis (Harel and Futerman 1993). Similarly, levels of cell surface cholera toxin, which binds to ganglioside GM1, were significantly reduced after incubation with FB1 (Sofer and Futerman 1995).
Whereas no labeling with B17 was seen when neurons were incubated with the antibody before formaldehyde fixation in PDMP- or FB1-treated cells (Figures 3C, 3D, 4A, and 4B), a variety of labeling patterns were seen after incubation with the antibody after fixation, including striped labeling in the neuronal processes (formaldehyde fixation; Figures 4C and 4D), bright labeling of the whole cell (formaldehyde and acetone fixation; Figures 4E and 4F), labeling of the cell body and nuclear envelope (acetone fixation; Figures 4G and 4H), and labeling of the cell body (methanol; Figures 4I and 4J). Because GD1b levels were almost completely abolished by PDMP and FB1 treatment, these labeling patterns cannot reflect the actual intra-cellular location of GD1b. Likewise, a striped labeling pattern was also observed when formaldehyde-fixed neurons were incubated with a polyclonal antibody to GM4 (Figure 5B) and with MAb BMA190, which reacts with GD2 (Figure 5D), although when incubated with the antibody before formaldehyde fixation no labeling was observed (Figures 5A and 5C). Because GD2 is expressed mainly in undifferentiated tissues and in tumor cells (Hakomori 1986), although GD2 is an intermediate that does occur at low levels in neurons (Hirschberg et al. 1996), and because GM4 is expressed mainly in oligodendrocytes (Yu et al. 1989; Yu and Iqbal 1979) and is not expressed in hippocampal neurons (Hirschberg et al. 1996), these data indicate that incubation with anti-ganglioside antibodies after formaldehyde fixation can lead to erroneous localization of gangliosides in neurons.
In contrast to the variety of labeling patterns observed with different fixation methods for the four antibodies discussed above, the labeling patterns observed with the JONES antibody (which reacts with 9-O-acetylated GD3) (Figure 6), or with antibody R24 (which reacts with ganglioside GD3) (not shown), were not affected by fixation. JONES labeled ~30% of neurons 2 hr after neurons were plated on glass cover-slips (Figure 7A), and a similar number of JONES-positive cells and a similar pattern of labeling were observed after all fixation techniques (Figures 6C-6H), with the exception of methanol, which completely abolished labeling (Figures 6I and 6J). The number of JONES-positive cells decreased significantly as neurons matured, and labeling was most prominent in cells that had not yet extended neuronal processes (Figure 7A). This is consitent with previous studies that used this antibody to examine the developmental profile and cellular localization of 9-O-acetylated GD3 (Blum and Barnstable 1987; Constantine-Paton et al. 1986). In contrast, when analyzed before fixation (Figures 3A and 3B), antibody B17 reacted mainly with cells that had already extended neuronal processes (Figure 7B), consistent with previous immuno-histochemical analysis using this antibody (Harel and Futerman 1993).
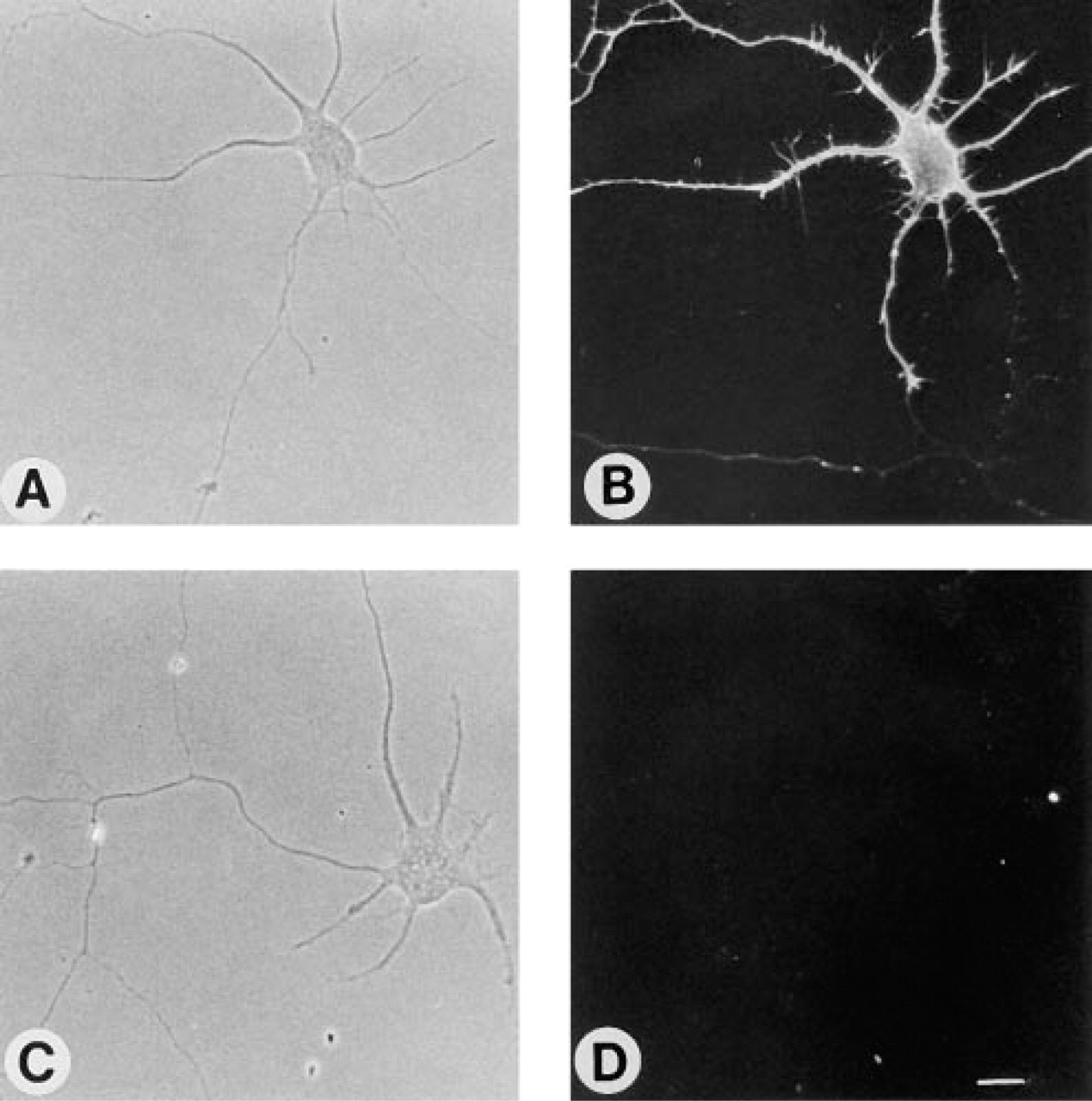
Effect of PDMP on binding of MAb B17 to 3-day-old cultures of hippocampal neurons. Control neurons (
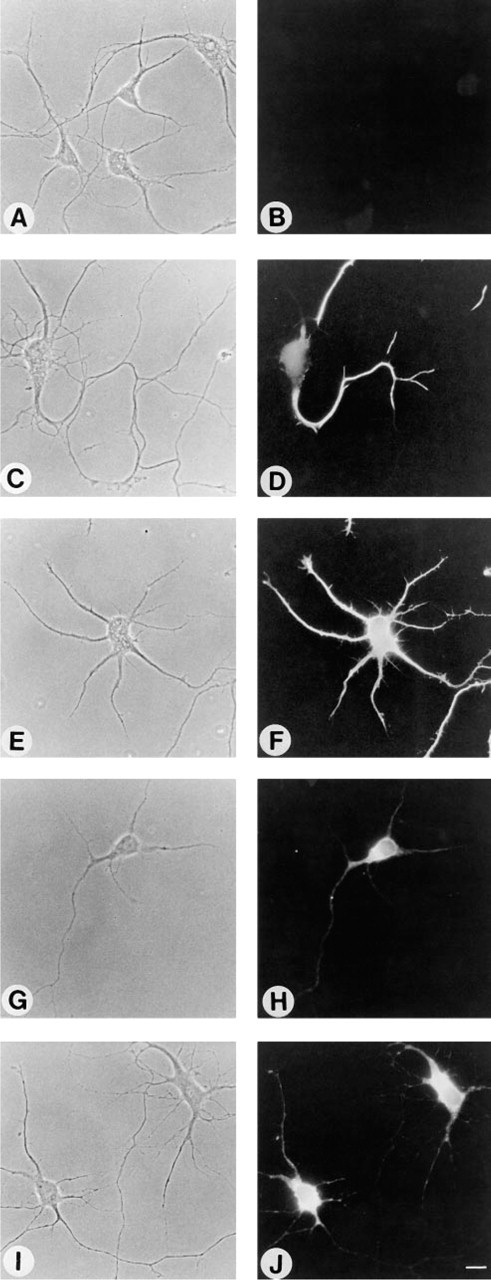
Effect of fixation methods on binding of MAb B17 to 3-day-old cultures of hippocampal neurons after reduction of sphingolipid and ganglioside levels by FB1. Neurons were either fixed with formaldehyde after application of antibody B17 (
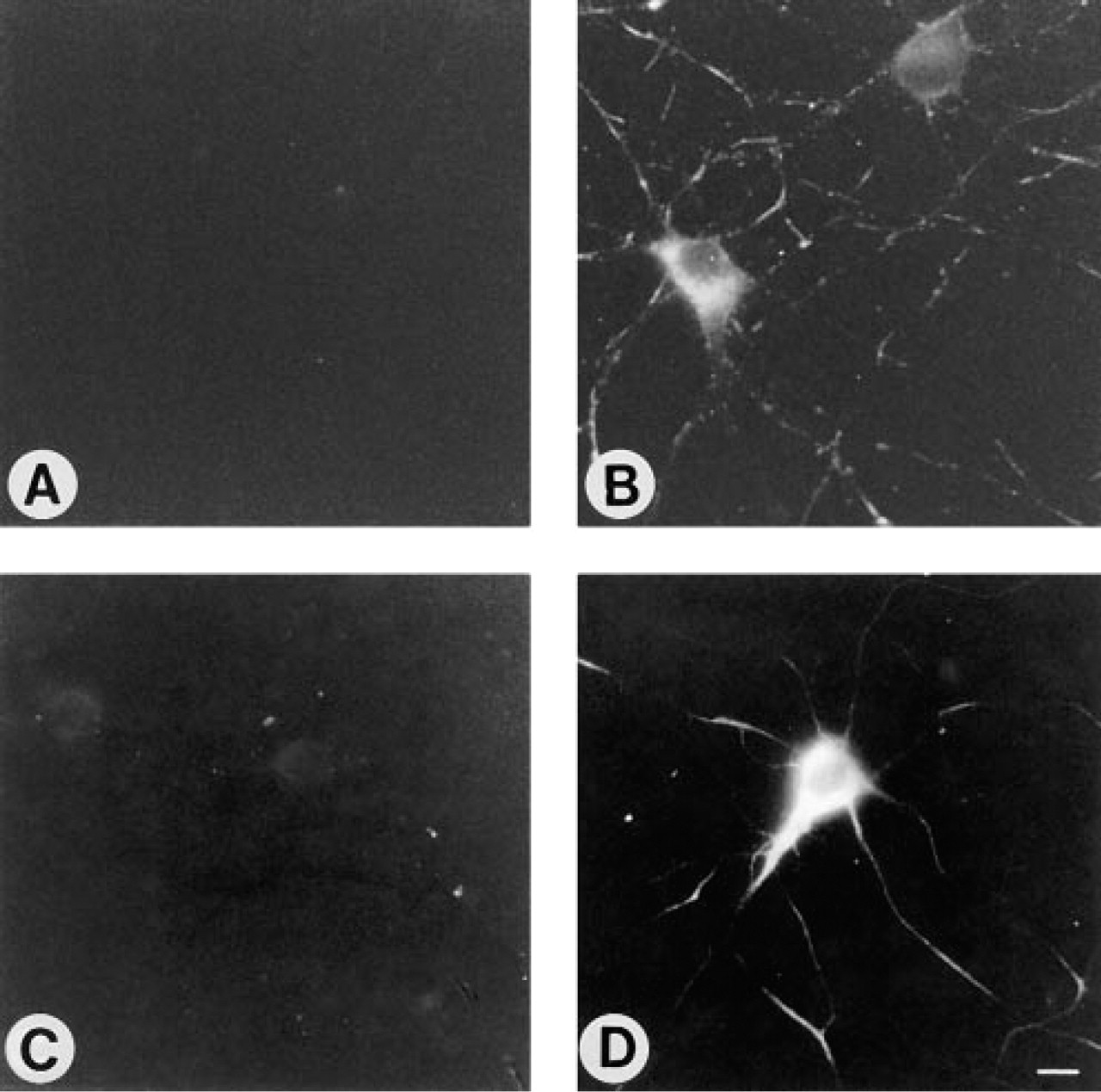
Effect of formaldehyde fixation on binding of an anti-GM4 polyclonal antibody (
Discussion
Anti-ganglioside antibodies have become a popular tool for localizing gangliosides in neuronal and non-neuronal tissues (Schwarz and Futerman 1996; Yu and Saito 1989). However, because of the unique solubility properties of lipids (Stein and Stein 1971), and because gangliosides are expressed mainly at the cell surface (van Echten and Sandhoff 1993), mild fixation, if fixation is needed at all, must be used to localize gangliosides, at least in dissociated cultures, although stronger fixation and permeabilization are presumably needed in whole tissues. If fixation is necessary, then optimal fixation conditions must be established for each anti-ganglioside antibody. For example, whereas the JONES MAb is relatively insensitive to the type of fixative, A2B5 and an anti-GD1b antibody (B17) gave distinct labeling patterns depending on the type of fixative. Moreover, biochemical analysis confirmed that the labeling could not have been due to binding to gangliosides. This is the first time that direct analysis of the effect of fixation methods on ganglioside levels has been reported, although an earlier study compared the binding of HNK-1-reactive antigens with levels of sulfoglucuronyl glycolipids (Prasadarao et al. 1990).
Effect of fixation methods on binding of JONES MAb to hippocampal neurons immediately after plating. Neurons were either fixed with formaldehyde after incubation with JONES (
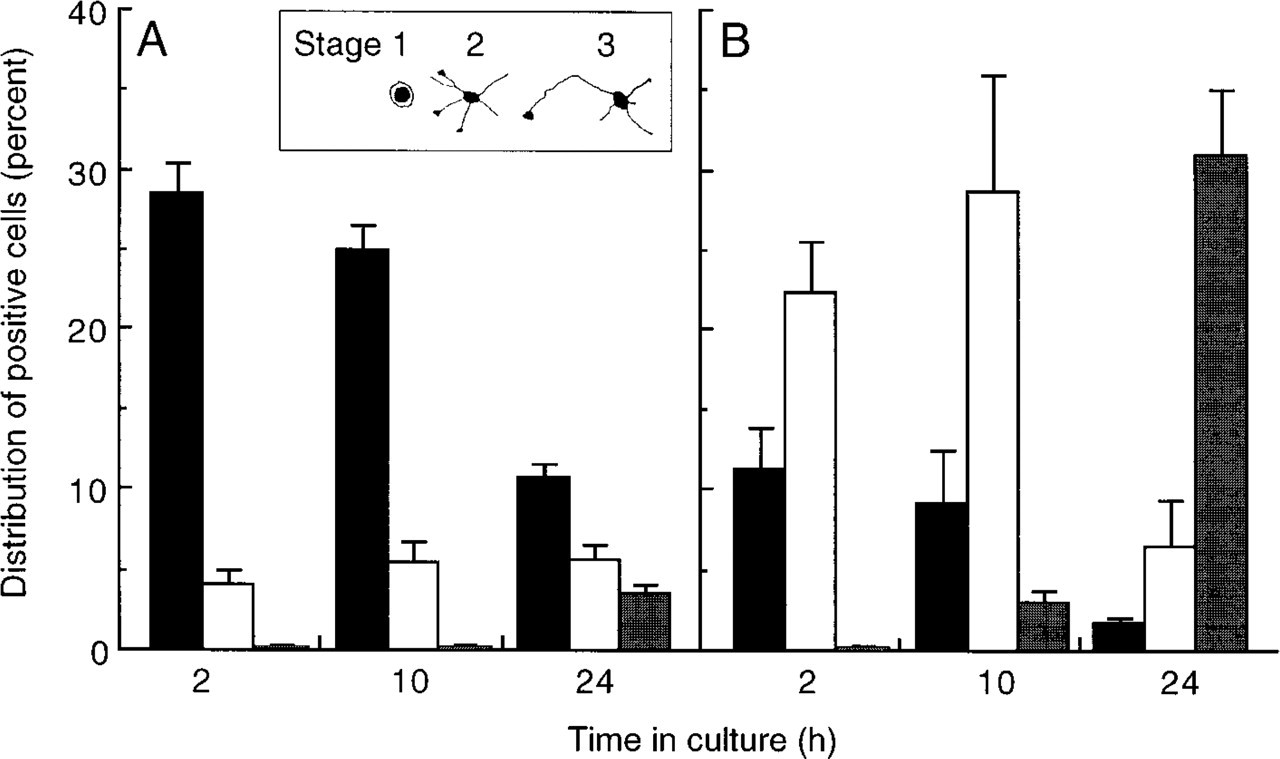
The number of JONES- and B17-positive cells at each developmental stage. Neurons were labeled before fixation at various times in culture with JONES (
Organic solvents, such as acetone and methanol, are particularly problematic as fixatives because both solubilize gangliosides to various extents. Our biochemical analysis demonstrated that methanol completely solubilizes gangliosides from hippocampal neurons, with acetone also solubilizing gangliosides by >50%. This implies that binding of anti-ganglioside antibodies to methanol-treated tissues may be due to crossreactivity of the antibodies with unrelated epitopes, and also suggests that published conclusions using acetone as a fixative may need to be re-evaluated. It has been suggested that gangliosides become antigenic only after acetone treatment, because acetone may act to promote the accessibility of the ganglioside (Graus et al. 1994) or may unmask membrane-bound carbohydrates (Thorpe and Kerr 1994). However, no GM3 was detected with an anti-GM3 antibody in prefixed cryostat sections after acetone treatment (Daniotti et al. 1994) or after tratment of formaldehyde-fixed tissue sections with methanol, acetone, or ethanol/acetic acid (Reynolds and Wilkin 1988).
Mild fixatives such as formaldehyde can also cause artifactual labeling. Therefore, an anti-GM4 antibody bound to formaldehyde-fixed hippocampal neurons, even though GM4 is absent from these neurons and is found only in glia (Yu et al. 1989; Yu and Iqbal 1979). In many cases, a non-uniform “striped” labeling pattern was observed after formaldehyde fixation, which does not indicate that the ganglioside is present in a non-uniform distribution over the neuronal cell surface but rather that formaldehyde interferes with the antigenicity of the gangliosides, or with the binding of the antibodies, resulting in unusual labeling patterns.
The reason for the differences in the sensitivity of various antibodies to different fixatives is not clear, but may be due to differences in the solubility of the glycolipid antigens, to differences in accessibility, or to nonspecific binding of certain antibodies to related carbohydrate antigens that may become accessible after treatment with various fixatives. However, it is clear that the differences are not due to the immunoglobulin class, because both B17 and JONES antibodies are of the IgM class but each antibody behaves differently under various fixation conditions.
In summary, whereas previous workers have recognized that the antigenicity and immunogenicity of gangliosides and other glycosphingolipids depends on the density of the antigen on the cell surface, the chain length of the fatty acids and their α-hydroxylation, and the cryptic behavior of glycosphingolipids (Hakomori 1986), the present study demonstrates that the method of fixation is vitally important for unambiguous conclusions to be drawn about the localization of a ganglioside in biological tissues using anti-ganglioside antibodies. This is of importance not only for localizing gangliosides in biological tissues (Schwarz and Futerman 1996) but also for using anti-ganglioside antibodies as diagnostic tools (Bernhard et al. 1992; Lloyd et al. 1992).
Footnotes
Acknowledgements
Supported by the German-Israel Foundation for Scientific Research and Development.
We thank Dr Israel Pecht for providing the B17 antibody, Dr Colin J. Barnstable for providing the JONES antibody, and Behring Werke AG for providing BMA190. We also thank Rivi Zisling for expert help in preparing and maintaining hippocampal cultures. AHF is the incumbent of the Recanati Career Development Chair in Cancer Research.