
Abstract
Peroxisomes are cytoplasmic organelles involved in a variety of metabolic pathways. Thus far, the morphological and biochemical features of peroxisomes have been extensively characterized in adult tissues. However, the existence of congenital peroxisomal disorders, primarily affecting tissue differentiation, emphasizes the importance of these organelles in the early stages of organogenesis. We investigated the occurrence and tissue distribution of three peroxisomal enzymes in rat embryos at various developmental stages. By means of a highly sensitive biotinyl-tyramide protocol, catalase, acyl-CoA oxidase, and ketoacyl-CoA thiolase were detected in embryonic tissues where peroxisomes had not thus far been recognized, i.e., adrenal and pancreatic parenchyma, choroid plexus, neuroblasts of cranial and spinal ganglia and myenteric plexus, and chondroblasts of certain skeletal structures. In other tissues, i.e., gut epithelium and neuroblasts of some CNS areas, they were identified earlier than previously. In select CNS areas, ultrastructural catalase cytochemistry allowed identification of actively proliferating organelles at early developmental stages in several cell types. Our data show that in most organs maturation of peroxisomes parallels the acquirement of specific functions, mainly related to lipid metabolism, thus supporting an involvement of the organelles in tissue differentiation.
P
Although sharing some common features, peroxisomes show species and tissue specificity. In rat and human liver, for example, the organelles exhibit a roughly spherical shape and a finely granular matrix (Wedel and Berger 1975; Sternlieb and Quintana 1977), while more elongated peroxisomes are present in human kidney and gut (Roels and Goldfischer 1979; Roels et al. 1991). Differences in matrix composition and in the relative abundance of membrane proteins among various organs have been also reported (Usuda et al. 1991). Moreover, ultrastructural and enzymatic heterogeneity of peroxisomes inside a tissue and even within a single cell has been demonstrated (Stefanini et al. 1985; Angermüller and Fahimi 1988; Roels and Cornelis 1989; Roels et al. 1991; van den Munckhof 1996; Cimini et al. 1998). Finally, dishomogeneous localization of specific enzymes, such as urate oxidase, α-hydroxy acid oxidase B, and
The pivotal role of peroxisomes in development is emphasized by the existence of congenital diseases caused by the absence or impaired function of peroxisomes and leading in most cases to premature death (Wanders 2000; Moser 2000; Baumgartner and Saudubray 2002). In these disorders, both the function and the histology of several organs, such as liver, kidney, brain, adrenal gland, and testis, are profoundly affected (Powers et al. 1982; Goldfischer et al. 1983; Nissenkorn et al. 2001).
Concerning peroxisome biogenesis, the fission of pre-existing organelles as well as the sequential import of membrane and matrix proteins are widely accepted (Subramani 1993; Purdue and Lazarow 2001).
During prenatal development, peroxisomes have been detected in several mammalian organs, and an overview of the distribution of peroxisomal enzymes in the mouse embryo focused on selected prenatal stages has been recently presented (Grabenbauer et al. 2001).
The aim of this investigation was to study the presence and maturation of peroxisomes in rat embryos at various developmental stages, using polyclonal antibodies against the peroxisomal marker catalase (CAT) and two other enzymes, i.e, acyl-CoA oxidase (ACOX), and ketoacyl-CoA thiolase (THL), involved in the peroxisomal fatty acid β-oxidation system. To this purpose, we applied a highly sensitive IHC procedure based on the deposition of biotinylated tyramine on the tissue (Adams 1992), which was successfully used by our group in different organs (Moreno et al. 1995,1999; Farioli-Vecchioli et al. 2001a,b). In select regions of the nervous system, peroxisomes were also investigated at the ultrastructural level by CAT cytochemistry.
Materials and Methods
Animals
Female albino Wistar rats (Charles River; Milan, Italy) were placed with males overnight and examined the next morning for the presence of sperm in the vaginal smear. The day of sperm observation was considered as embryonic day 0.5 (E0.5). Animals were housed and handled according to the European Communities Council Directive of 24 November 1986 (86/609/EEC).
Fixation and Tissue Processing
Pregnant rats at days between E9.5 and E17.5 were sacrificed under deep anesthesia with Farmotal (Amersham Pharmacia Biotech Italia; Milan, Italy) 100 mg/kg bw injected IP. The uterus was quickly removed, immersed in Ringer's solution and sectioned in fragments corresponding to individual embryos.
At the earliest stages, uterine segments were fixed by immersion in Bouin. Starting from E12.5, embryos were extracted from the deciduas, weighed, and measured to confirm their gestational age, and fixed by immersion in Bouin for 1-20 hr at room temperature (RT). Embryos older than 15.5 days were cut along the sagittal plane 10 min after immersion in the fixative solution. After fixation, specimens were dehydrated in graded alcohols and embedded in paraffin.
Immunohistochemistry
Serial sagittal microtome sections (8 μm) were deparaffinized, transferred to methanol containing 0.3% H2O2 for 30 min at RT, and rehydrated. Slides were immersed in PBS containing 5% non-fat dry milk and 0.2% Triton X-100 for 1 hr at RT and then incubated for 24 hr at 4C with rabbit polyclonal antibodies in PBS containing 2.5% non-fat dry milk and 0.1% Triton X-100. Dilutions were as follows: anti-CAT (Rockland; Gilbertsville, PA) 1:500; anti-ACOX (generous gift of Prof. A. Völkl, University of Heidelberg, Germany) 1:200; anti-THL (generously supplied by Prof. T. Hashimoto, Shinshu University, Nagano, Japan) 1:500. In control sections, the primary antibody was omitted or substituted with normal rabbit serum. Immunocomplexes were revealed by an amplified ABC procedure (Adams 1992) since preliminary experiments performed by conventional ABC method gave a very weak signal.
Biotinylated goat anti-rabbit IgG and avidin-biotin-HRP (standard ABC kit) were purchased from Vector (Burlin-game, CA); normal goat serum, tyramine-HCl, and DAB were from Sigma Chemical (St Louis, MO); NHS-biotin was from Pierce Chemical (Rockford, IL).
For each developmental stage, several sections from at least three embryos were submitted to the localization of peroxisomal enzymes and examined in a Zeiss Axioplan 2 equipped with a video camera. In each section, organs and tissues were identified based on Hebel and Stromberg's (1986) description of rat embryology. The Kaufman's (1992) atlas of mouse development was also used, taking into account the reported differences in the timing of appearance of principal structures.
A semi-quantitative evaluation of the labeling intensity was independently performed by three researchers, who assigned scores ranging between - and + + +. Representative images were electronically captured; contrast and brightness were adjusted by Adobe Photoshop 5.5.
Cytochemistry
E10.5 to E13.5 embryos were excised from the decidua and immersed in 4% formaldehyde in 0.1 M cacodylate buffer, pH 7.4, containing 1% calcium chloride overnight at RT. E12.5 and 13.5 embryos were cut in fragments. Samples were then rinsed in the same buffer and incubated with The-orell buffer, pH 10.5, containing 0.2% DAB and 0.01% H2O2 for 3 hr at RT. For negative control, the incubation medium was supplemented with 0.1 M 3-amino-1,2,4-triazole (Sigma). After washings with 13% sucrose in H2O, specimens were postfixed in 1% OsO4 in 0.1 M cacodylate buffer containing 1.6% K3Fe(CN)6 for 1 hr at RT, dehydrated, and embedded in Epon. Ultrathin sections from select areas of neural tissue were contrasted with uranyl acetate and observed in a Philips CM120 electron microscope.
Results
Immunohistochemistry
In all the examined periods, peroxisomal enzymes were localized in the cytoplasm of a number of cell types. In several tissues the labeling had a granular appearance, with granules either distributed in the whole cytoplasmic compartment or concentrated in specific areas, whereas in other tissues the cytoplasmic compartments appeared more homogeneously stained. The nonspecific staining due to spontaneous biotinyl-tyramide binding was generally irrelevant. In all the sections incubated without the primary antibody, specific labeling was absent, while the nonspecific staining was similar to that found in immunoreacted specimens.
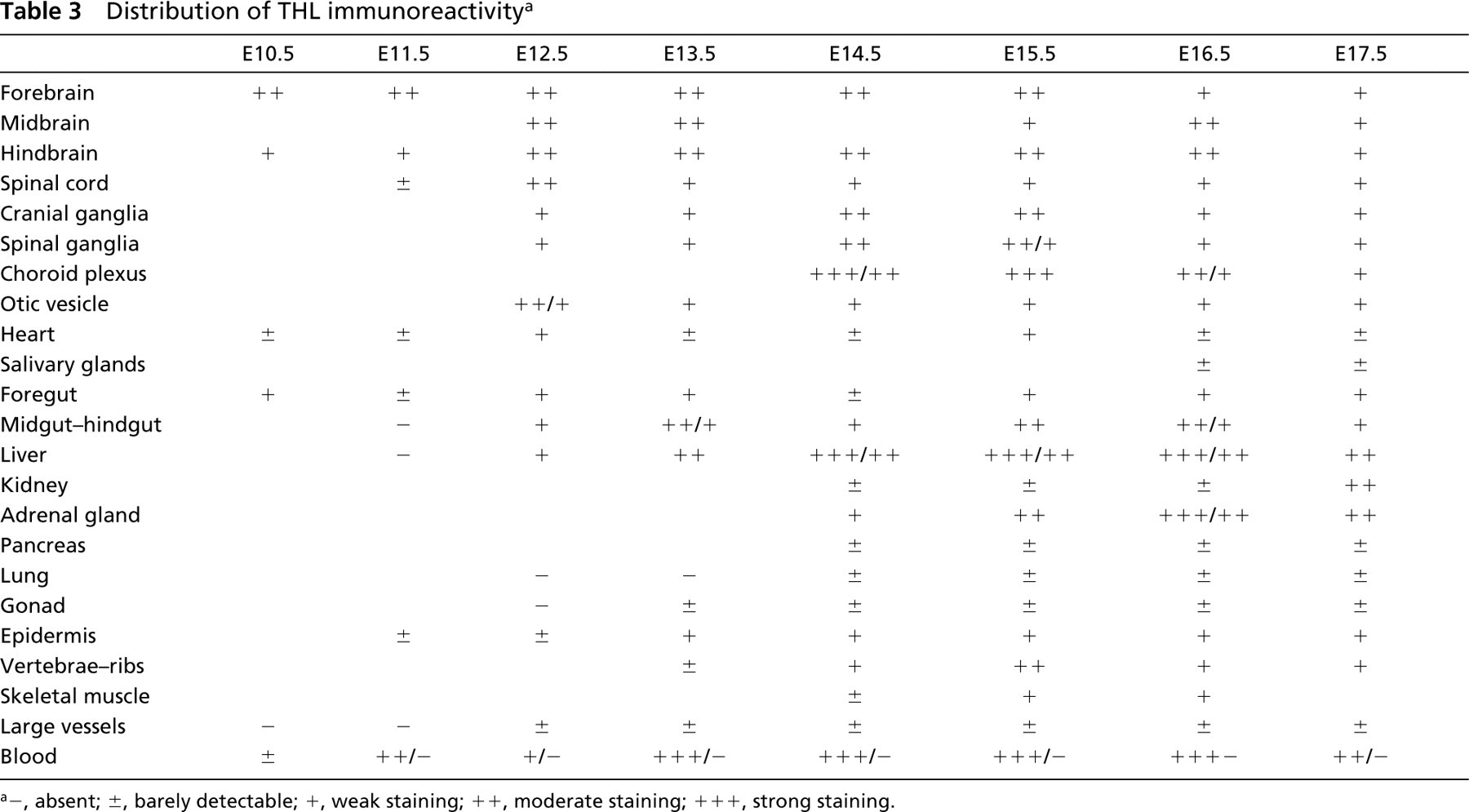
In a semi-quantitative attempt to summarize our data, we listed the major organs and systems at developmental stages between E10.5 and E17.5 on a scale of immunoreactivity (Tables 1–3). In organs composed of parenchyma and stroma, the parenchyma was usually more heavily labeled. For this reason, the results given in the tables refer to the parenchyma.
Distribution of CAT immunoreactivity a
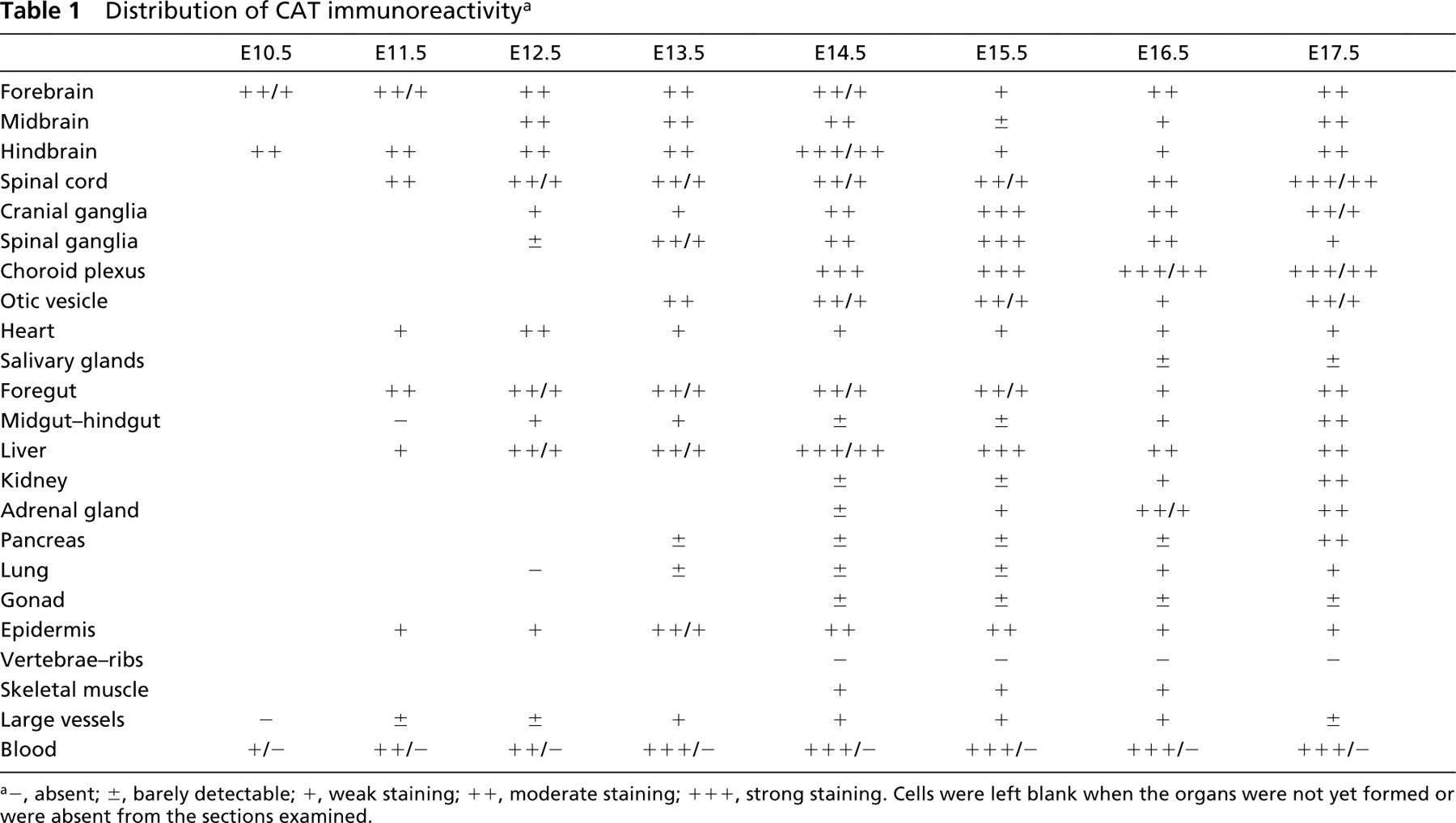
−, absent; ±, barely detectable; +, weak staining; + +, moderate staining; + + +, strong staining. Cells were left blank when the organs were not yet formed or were absent from the sections examined.
Distribution of ACOX immunoreactivity a
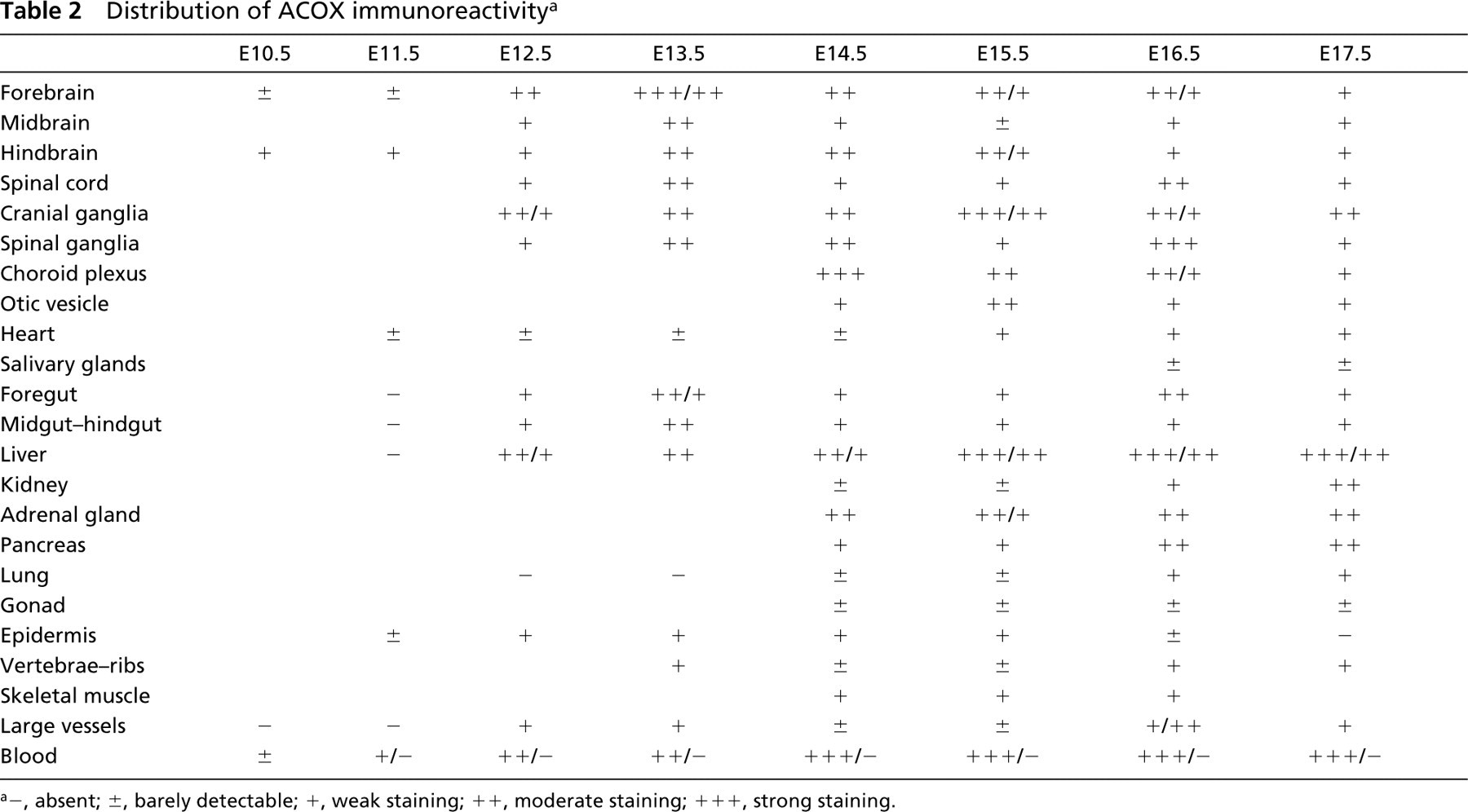
−, absent; ±, barely detectable; +, weak staining; + +, moderate staining; + + +, strong staining.
Distribution of THL immunoreactivity a
−, absent; ±, barely detectable; +, weak staining; + +, moderate staining; + + +, strong staining.
A detailed description of the distribution of the enzymes in various organs is given below.
Brain
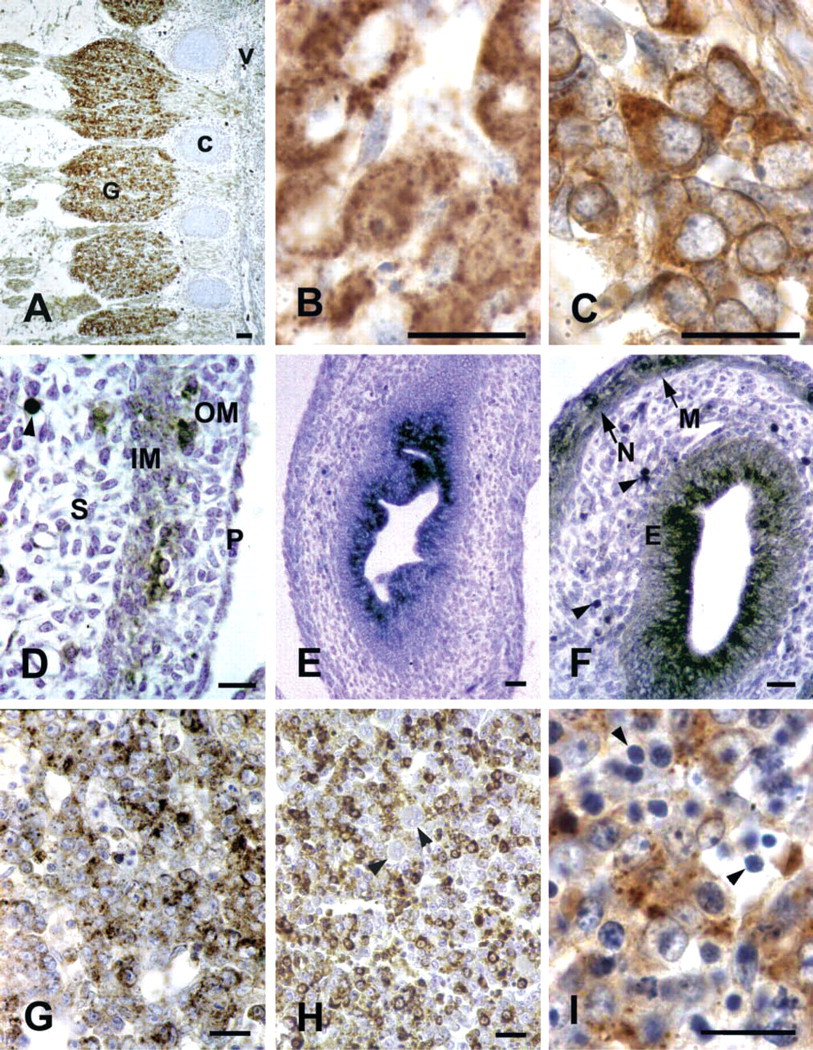
In the walls of cerebral ventricles, immunore-activity to the peroxisomal enzymes was detected starting from E9.5 and showed little variation during the periods examined. In the telencephalon, positive neuronal cells were more numerous at the the pial surface (Figures 1A and 1B) with respect to the ventricular zone, the roof of which showed relatively higher munoreactivity than the floor. In the midbrain, labeled neuroblasts were more randomly distributed, while in the hindbrain immunoreactive cells were more common in the ventricular area (Figures 1C and 1D) than in the pial zone. Moreover, during development, neuroblasts localized in the floor of the IV ventricle became progressively more immunoreactive than those in the roof.
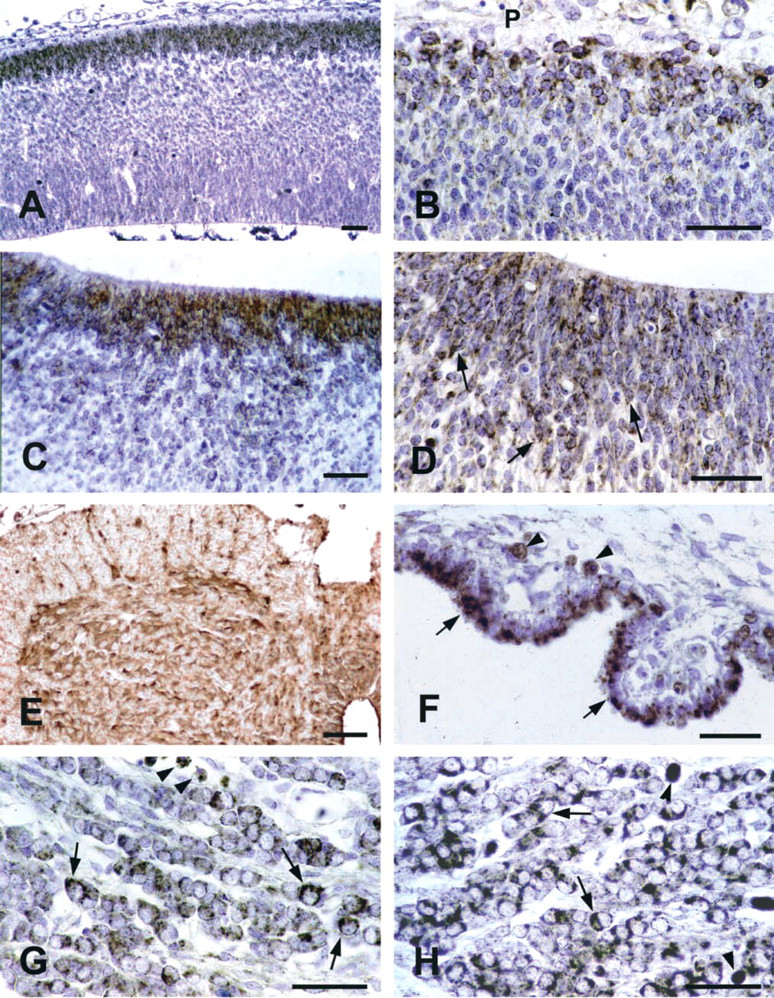
IHC of peroxisomal enzymes in nervous system. (
Spinal Cord
At all the examined developmental stages, all the neuronal cells showed high degrees of CAT staining, while ACOX and THL were less abundant (Figure 1E).
Cranial and Spinal Ganglia
Starting from E12.5 and especially around E15.5-16.5, ganglia were intensely immunolabeled by peroxisomal antibodies. For each enzyme, the positivity was restricted to neuronal cells. Individual neuroblasts showed different degrees of staining (Figures 1G, 1H, and 2A-2C).
IHC of peroxisomal enzymes in different tissues. (
Choroid Plexus
Starting from their first identification (E14.5), the choroid plexus showed strong immunoreactivity to the three enzymes. This pattern remained unchanged during the whole period examined, the labeling being concentrated in the apical portion of the epithelial cells (Figure 1F). The junctional epithelium between the choroid plexus and the adjacent ependyma was also stained.
Heart
In the myocardium, moderate to low staining for the three proteins was observed at all the stages examined. Labeling degrees were similar across different cardiac regions and among adjacent cardiomyocytes (not shown).
Salivary Glands
At E16.5, a mild positivity to anti-ACOX and anti-THL was found in the parenchyma of the submandibular gland.
Gut
In the oropharyngeal epithelium, CAT was detected as early as E11.5, while other enzymes first appeared at E12.5. High staining levels were present until the end of the period examined. In the stomach epithelium, all the enzymes were immunodetected starting from E13.5 and high staining degrees were found at E15.5 and later. In duodenal mucosa, especially high levels of ACOX and THL were reached at E16.5. In the midgut, ACOX, THL, and CAT were detected in cells randomly located in the epithelium starting at E12.5. High ACOX and THL positivity was achieved at E13.5 and maintained later. Differences in staining degrees occurred among midgut sections. No labeling was found in the hindgut whenever it was recognized.
In all the intestinal regions, submucosa was constantly negative, whereas weak CAT and ACOXimmunostaining was found in smooth muscle layers. In addition, starting at E15.5, ACOX immunoreactive cells showing neuronal morphology, probably corresponding to the prospective myenteric plexus, were recognized (Figures 2D-2F).
Liver
CAT was immunohistochemically detected in the hepatic primordium at E11.5, when ACOX and THL immunoreactivities were still negligible. In the following days the liver parenchyma showed increased staining for all the peroxisomal enzymes (Figures 2G-2I), remarkably high levels being found after E15.5. Differences in the labeling intensity were observed among individual embryos. At early developmental stages, virtually all the hepatocytes contained peroxisomal enzymes (Figure 2G). By contrast, starting at E14.5, the staining for each protein was restricted to groups of hepatocytes interspersed among others that were only faintly stained or even immunonegative (Figure 2H). At all the stages examined, finally, megakaryocytes were unstained, while a faint labeling was present in normoblasts.
Kidney
In the metanephros, CAT was significantly present from E15.5, while ACOX and THL were first found at E16.5 and E17.5, respectively. In the entire period examined, epithelial cells of the tubular portions of developing nephrons and parenchymal cells located in the superficial cortex were remarkably labeled. A weak staining was occasionally present in glomeruli (Figures 3A and 3B).
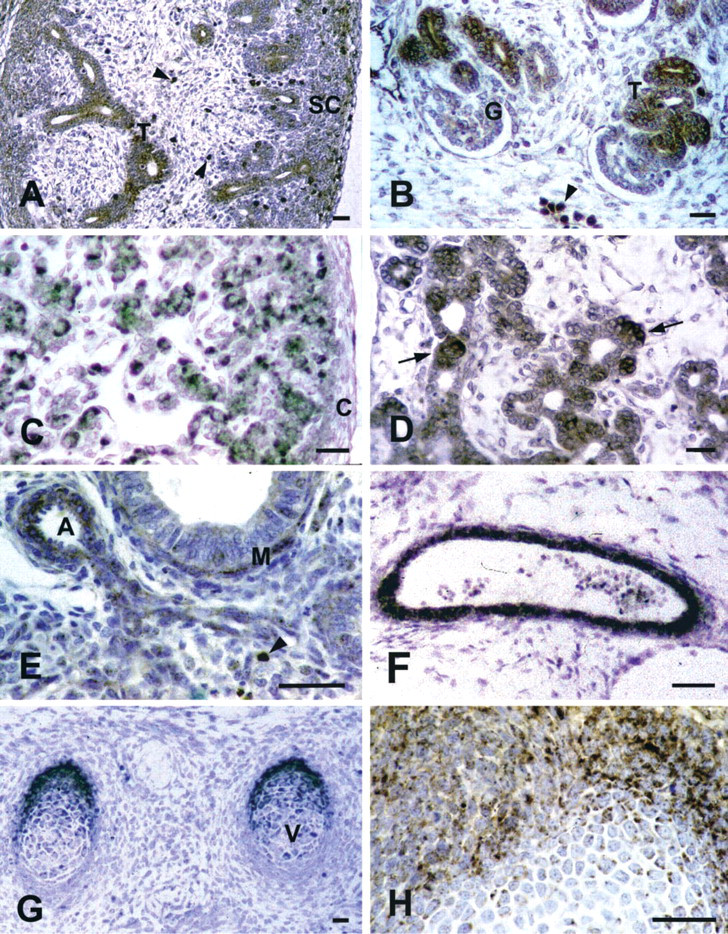
IHC of peroxisomal enzymes in different tissues. (
Adrenal Gland
Starting from E14.5, when the organ was first identified, the peroxisomal enzymes were immunodetected in randomly distributed parenchymal cells. Different staining intensities were also observed (Figure 3C).
Pancreas
All the peroxisomal markers were first detected in the pancreatic parenchyma at E14.5. Starting from E16.5, levels of CAT and ACOX were found. At E17.5, in particular, remarkable ACOX immunoreactivity was shown by individual cells of the tubular wall and by closely integrated epithelial cell groups, presumably corresponding to developing islets (Figure 3D).
Lung
Lung parenchyma was unlabeled after immunolocalization of the three enzymes during most of the period examined. Mild staining was detected in the bronchiolar epithelium at E17.5. At this stage, muscle layers of bronchioles and vessels showed strong, ACOX immunoreactivity (Figure 3E).
Large Vessels
Starting from E12.5-E13.5 until the end of the period examined, CAT and ACOX were detected in the walls of the main arteries (Figure 3F), such as the dorsal aorta and arteries of branchial archs. Large veins, including cardinal veins and primary head veins, showed weaker staining. At all stages, labeling for THL was low to absent.
Skeleton
Results mainly refer to vertebrae and ribs, the primordia of which were recognized starting from E13.5 and E14.5, respectively. Whereas CAT and ACOX remained almost undetectable throughout the period examined, THL immunoreactivity abruptly increased at E15.5, decreasing thereafter. In the majority of cartilage structures, the most intensely labeled chondroblasts were peripherally located (Figures 3G and 3H).
Blood
Many cells showing medium to high labeling levels for each peroxisomal marker were present inside vessels and heart lumina. Most of them were recognized as erythrocytes.
Cytochemistry
In specimens submitted to ultrastructural catalase cytochemistry, DAB reactivity was restricted to organelles recognizable as peroxisomes. The presence of the CAT inhibitor 3-amino-1,2,4-triazole in the incubation medium completely abolished the positivity.
DAB-reactive organelles were detected, starting from E13.5, in neuroblasts of several CNS areas, i.e., medulla oblongata, olfactory bulb, cerebellum, trigeminal ganglion, and retina. In all the tissues examined, peroxisomes frequently showed elongated shapes and tail-like expansions. Non-homogeneous distribution of CAT inside individual organelles was also consistently found (Figure 4).
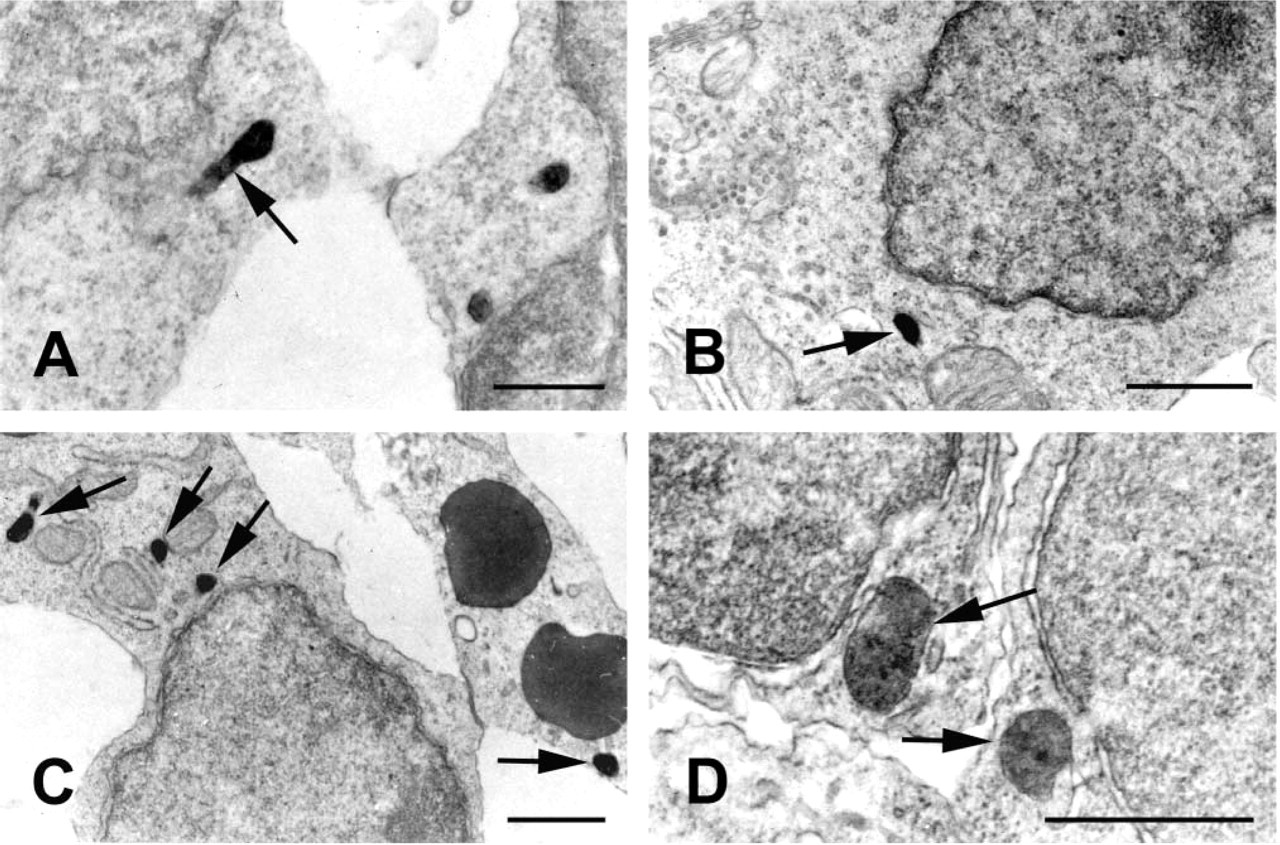
CAT cytochemistry in central nervous system. (
Discussion
The highly sensitive immunolocalization procedure employed here enabled us to detect peroxisomal proteins in the cytoplasmic compartment of cells in early developing rat tissues.
The granular labeling observed in the majority of tissues is consistent with the intraperoxisomal localization of these enzymes. The rather homogeneous staining of the cytoplasmic compartment obtained in some tissues may depend on massive deposition of the immunoreaction product.
The early appearance of peroxisomal enzymes during organogenesis is in agreement with the available literature (see references below). However, our study extends previous knowledge because in some organs peroxisomal markers were identified earlier than previously and in others their presence was detected here for the first time. In the following paragraphs, results obtained on different organs or tissues are discussed in view of the available literature.
Hepatic Parenchyma
The detection of CAT at E11.5 and its progressive increase confirm previous ultrastructural and IHC results (Tsukada et al. 1968; Grabenbauer et al. 2001). Moreover, the IHC patterns shown by the three enzymes appear in agreement with the widely accepted theory concerning sequential import of proteins during peroxisomal maturation (Stefanini et al. 1985; Espeel et al. 1997; Depreter et al. 1998).
At later stages, the differential immunoreactivities shown by individual hepatocytes confirm previous cytochemical data describing intra- and intercellular peroxisomal heterogeneity (Stefanini et al. 1985; Anger-müller and Fahimi 1988). It is worth recalling that in pathological conditions, e.g., in livers of patients affected by severe peroxisomal disorders, a mosaic distribution of parenchymal cells with or without peroxisomes has been described (Espeel et al. 1995).
Kidney Proximal Tubules
The relatively late appearance of peroxisomal markers in rat embryonic kidney when compared with mouse (Pipan and Psenicnik 1975; Goeckermann and Vigil 1975) and the regional selectivity shown by these organelles within the developing nephrons are consistent with previous ultrastructural and IHC data (Stefanini et al. 1994; Johkura et al. 1998,2000). Moreover, the earlier detection of CAT with respect to β-oxidation enzymes confirms that peroxisomal maturation occurs through the sequential import of matrix proteins.
In proximal tubules of adult mammalian kidney, characterized by high rates of lipid metabolism, peroxisomes are engaged in several pathways, such as amino acid reabsorption and interconversion, gluconeogenesis, or drug biotransformation (Zaar 1992). During prenatal life in tubular epithelial cells, peroxisomal biogenesis parallels the achievement of specific differentiation hallmarks, such as brush borders, apical vacuolar apparatus, and basolateral membrane infolding (Stefanini et al. 1994; Johkura et al. 1998). Formation of these structures requires high rates of lipid synthesis and turnover, functions in which peroxisomes are known to be involved.
Gut Epithelium
The appearance of CAT in the rat embryonic oropharyngeal epithelium at E11.5 represents the earliest detection of this enzyme in rodent digestive tract. In the other gut regions, the early appearance and the growing concentrations of the enzymes observed are in overall agreement with results obtained in mice and humans (Pipan and Psenicnik 1975; Calvert and Menard 1978; Dauça et al. 1996; Grabenbauer et al. 2001). In the mucosa of the stomach and the duodenum, in particular, the appearance and subsequent increase of peroxisomal markers parallel the beginning and the progression of glandular and villous differentiation.
In mammalian gut epithelium, peroxisomal β-oxidation is centrally involved in its intense lipid catabolism (Small et al. 1980; Cablé et al. 1993). Therefore, the appearance and maturation of peroxisomes can be considered as enterocyte differentiation markers. Moreover, during embryonic development, acquisition of features typical of the mature enterocyte, involving novel membrane production, presumably imply intense lipid anabolism by peroxisomes.
Pancreas, Adrenal Gland, and Salivary Glands
Although the occurrence and features of peroxisomes in the parenchymal compartment of rodent adult pancreatic, adrenal, and salivary glands have been previously described (Magalhães and Magalhães 1971; Beard 1972; Hand 1973; Arnold and Holtzman 1975; Hanker et al. 1977), to our knowledge we are the first to report the presence of peroxisomal enzymes in these organs during rat embryogenesis.
Peroxisomes of the adrenal cortex, owing to their content in sterol carrier protein-2 (van Amerongen et al. 1989), have been implicated in steroidogenesis (see Magalhães and Magalhães 1997). In some human peroxisomal disorders, the defective removal of very long chain fatty acids causes the formation of lamellar aggregates in adrenocortical cells even at prenatal stages (Powers et al. 1982; Goldfischer et al. 1983). This may reflect the early engagement of peroxisomal fatty acid β-oxidation enzymes in lipid metabolism of the organ.
Concerning the developing pancreas, peroxisomal fatty acid β-oxidation may play a key role in differentiating β-cells before their metabolic switch from intense fatty acid oxidation towards a high rate of glucose oxidation (Bliss and Sharp 1994).
Lung Parenchyma
The weak immunostaining shown by rat bronchiolar epithelium at E17.5 is consistent with previous reports on mouse embryonic lung, in which at comparable developmental stages peroxisomes could be detected only by ultrastructural cytochemistry (Schneeberger 1972). In the perinatal period and in adult life, by contrast, peroxisomes are abundant in non-ciliated bronchiolar cells and in type II pneumocytes (Petrik 1971), where they are believed to participate in the synthesis and turnover of surfactant (Ossendorp et al. 1994).
CNS
In the rat brain, peroxisomes have thus far been described in adult (Novikoff and Novikoff 1973; McKenna et al. 1976; Holtzman 1982; Moreno et al. 1995,1999; Farioli-Vecchioli et al. 2001a; Zaar et al. 2002) and neonatal (Arnold and Holtzman 1978) animals, even though in other species, markedly in humans, peroxisomal proteins have been immunodetected even during prenatal development (Houdou et al. 1991,1993; Imamura et al. 1994; Miyawaki et al. 1995; Itoh et al. 1999)
Here we report the presence of peroxisomal markers in the various rat CNS regions starting from E9.5. The early appearance and sustained expression of peroxisomal enzymes in neuroblasts of several brain regions, as well as in spinal cord, could be related to specific roles played by the organelles in the achievement and maintenance of neuron-specific functions. For example, the high membrane turnover rate necessary for neuron processes to form, extend, and reach their targets, might involve activation of peroxisomal fatty acid metabolism.
Concerning the regional distribution of immunolabeled neuroblasts, it is interesting to note that they are particularly abundant in areas, such as the cortical plate and the cerebellar neuroepithelium, that are selectively affected by generalized peroxisomal disorders (Powers and Moser 1998).
Interestingly, the various brain regions show differential staining of neuronal cells with respect to their maturation degree. In fact, while in the telencephalon the labeling is more concentrated in differentiated neurons than in neuroblasts of the ventricular zone, in the hindbrain immunoreactivity is more abundant in neuroblasts of the periventricular area.
At the electron microscopic level, our most relevant finding concerns the relatively frequent occurrence of elongated organelles with a rather inhomogeneous CAT content. This peculiar peroxisome staining, suggesting active proliferation and ongoing maturation, is consistent with previous reports (Fahimi et al. 1993; Cimini et al. 2000) and with the widely accepted theory of peroxisomal biogenesis (Subramani 1993).
Ganglia
The intense and early peroxisomal immunostaining found in virtually all the neuroblasts belonging to cranial and spinal ganglia is a novel finding. As far as we know, available data have been limited to satellite and Schwann cells inside dorsal root or sympathetic ganglia (Citkowitz and Holtzman 1973; Arnold and Holtzman 1975).
Myenteric Plexus
The presence of ACOX in neurons of prospective myenteric plexus, detected especially at E16.5 both in stomach and in several intestinal tracts, is here reported for the first time and deserves a deeper investigation.
Choroid Plexus
The high concentration of peroxisomal enzymes detected in epithelial cells of this organ after E14.5 is a novel finding. We suggest that peroxisomes play a pivotal role in several functions of choroid plexus epithelium, such as secretion and reabsorption of molecules of the cerebrospinal fluid (CSF). Peroxisomes are actively involved in the metabolism of prostaglandins (Lazarow and de Duve 1976), major components of the CSF (Chodobski and Szmydynger-Chodobska 2001). Furthermore, the organelles may be specifically implicated in epithelial differentiation, particularly in membrane turnover necessary for brush border and basolateral infolding formation.
Skeletal, Cardiac, and Smooth Muscle
To our knowledge, our results on peroxisomal enzymes in the skeletal muscle and myocardium of prenatal rat are the first on this topic because available data are limited to adult animals (Hand 1974; Tarach 1978). Further studies are needed to clarify peroxisomal functions in these tissues, especially in view of the central role played by the organelles in the intense lipid catabolism occurring in the adult heart (Hinton and Price 1993).
Our data concerning the presence of CAT and ACOX in myocytes belonging to gut and large vessels are difficult to interpret because previous reports dealing with peroxisomes in adult or embryonic smooth muscle are few and conflicting. For example, CAT, ACOX, and their mRNAs were detected in mouse gut myocytes starting from E14.5 (Grabenbauer et al. 2001), whereas cytochemical methods failed to reveal peroxisomes in human embryonic intestine (Dauça et al. 1996).
Cartilage
To our knowledge, the presence of peroxisomes in chondrocytes has thus far been suggested only in adult mammal epiphyseal growth plate, on the basis of histochemical data (Davis et al. 1989). Therefore, our IHC findings in rat embryonic skeleton are novel, although preliminary. In particular, the sharp increase of THL IHC found in vertebral primordia at E15.5 could indicate a specific role of this enzyme in the maturation and/or function of cartilaginous tissue. Such a hypothesis is indirectly supported by the defect in THL import found in some peroxisomal diseases, such as the rhizomelic form of chondrodysplasia punctata, characterized by abnormal skeletal development (Heikoop et al. 1990).
Blood
In erythroblasts, CAT immunostaining might simply reflect the presence of soluble enzyme, in analogy with what has been reported in mature erythrocytes. On the other hand, the occurrence of β-oxidation enzymes might be related to the presence of peroxisomes, thus far unidentified in immature blood cells.
Concluding Remarks
Here we show that peroxisomal enzymes are widely expressed throughout prenatal development and that their immunoreactivity patterns are temporally correlated with the acquisition of specific catabolic or anabolic pathways, and more generally with the appearance of differentiation hallmarks.
For example liver, gut and kidney peroxisomes are likely to prepare the respective organs for the high rate of fatty acid oxidation characteristic of maturity. Interestingly, in these organs the high levels of peroxisomal enzymes parallel the expression of the peroxisome proliferator activated receptor α (PPARα) (Braissant and Wahli 1998), known to induce fatty acid β-oxidation enzymes (Lemberger et al. 1996). Moreover, in gut, kidney, and choroid plexus epithelia, peroxisomal enzymes, which increase with the formation of brush border and basal infoldings, presumably participate in the intense lipid anabolism needed for membrane production and turnover. Similarly, in the CNS, where all the PPAR isotypes were immunodetected (Braissant and Wahli 1998), the role exerted by peroxisomes in lipid metabolism might be crucial in the process of neuronal elongation. PPARs (Braissant and Wahli 1998; Keller et al. 2000) and peroxisomes might also be important in regulating cartilage maturation. In the adrenal gland, the early detection of peroxisomal enzymes might correspond to their engagement in the early beginnings of steroidogenesis, while in lung their relatively late appearance would be consistent with the late onset of surfactant synthesis.
Footnotes
Acknowledgements
We wish to thank Prof A. Völkl (University of Heidelberg, Germany) and Prof T. Hashimoto (Shinshu University, Nagano, Japan) for generously providing primary antibodies.
We also gratefully acknowledge Prof Maria Paola Cerù (University of L'Aquila, Italy) for critical review of the manuscript.