
Abstract

S
Structure and Organization of the Growth Cone
The growth cone has two major domains characterized by the cytoskeletal components found mainly therein. The peripheral domain (Figures 1a–1c, p) is rich in actin filaments, which are organized into different types of structures. The filaments in parallel bundles are called ribs, if they do not protrude from the body of the growth cone, or filopodia, if they extend into finger-like projections. In between these rib structures at the membrane are often found a meshwork of filaments in regions that are extending forward. These membrane expansions are called lamellipodia, if they are closely associated with the substratum, or membrane ruffles, if they appear on the apical surface of the growth cone. Ruffles are usually formed from lamellipodia that fail to adhere to the substratum and collapse back into the growth cone. A veil is a region of expanding membrane between two filopodia. Veils can adhere to the substratum to generate a lamellipodium, or they can lift back and generate a ruffle. Actin within the p domain is undergoing dynamic assembly that lead to membrane protrusion. Recruitment and activation of myosin lead to contraction and the retrograde flow of actin back towards the central (c) domain (Forscher and Smith 1988).
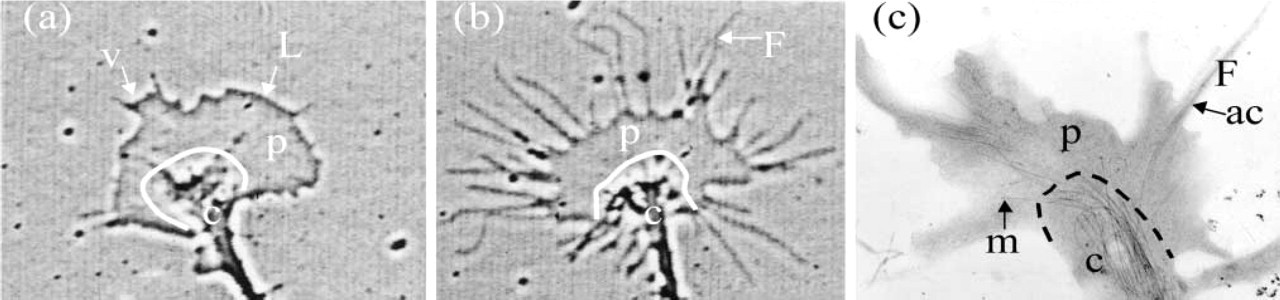
Phase-contrast (
The central domain (Figures 1a–1c) is rich in microtubules and membranous organelles. Microtubules are oriented with their plus ends towards or within the p domain (Gordon-Weeks 1993) and serve as the major cytoskeletal structure on which rapid vesicular transport occurs (Schnapp et al. 1987). Vesicle delivery to the growth cone is necessary to maintain the supply of membrane used for neurite elongation. The majority of membrane addition for neurite extension occurs at the growth cone (Lockerbie et al. 1991). Fluorescent speckle microscopy has been used to examine the dynamics of microtubules that extend into the actin-rich domain of lamellipodia in non-neuronal cells (Salmon et al. 2002). Microtubules extending into this domain are often caught up in the retrograde flow of actin and are observed to buckle and even break. Similar results have been observed for microtubules extending into the p domain of growth cones (Schaefer et al. 2002).
Microtubules extending into this p domain appear to be important for neurite pathfinding (Gordon-Weeks 1991; Tanaka and Sabry 1995; Tanaka et al. 1995; Williamson et al. 1996). Earlier work on neurite growth using drugs that targeted either actin filaments or microtubules suggested that neurite extension can occur in the absence of actin dynamics through microtubule-dependent processes (Marsh and Letourneau 1984; Bentley and Toroian-Raymond 1986). Path-finding, however, did not occur in the absence of actin dynamics, suggesting that actin is required for guidance in response to environmental cues but that microtubule-based growth alone could extend neurite processes. Recent studies suggest that important interactions occur between microtubules and actin filaments in the pathfinding process, and that it is the capture by actin filaments of microtubules randomly penetrating into the p domain that is an important step in guidance (Schaefer et al. 2002).
The formation of focal adhesions is coupled with the actin assembly-based protrusion of membrane at the leading edge of growth cones (Steketee and Tosney 2002). These adhesions provide substrate attachment of the expanding filopodia and lamellipodia. Maintenance of these adhesions is necessary for growth cone spreading and advance (Mikule et al. 2002). Once adhesion has occurred, the growth cones can generate contraction through the activity of myosin (see Brown and Bridgman, this issue). In one model of growth cone advance, the clutch hypothesis, the contractile events in the growth cone are coupled to either retrograde flow of the actin or to rapid forward advance, depending on whether or not the adhesion of the growth cone to the substrate is coupled by clutch molecules that can translate the retrograde forces on the actin to forward advance (Lin and Forscher 1995).
Mechanisms of Regulating Growth Cone Guidance
The mechanisms by which extracellular guidance cues regulate growth cone turning and pathfinding have not been completely elucidated for any single guidance molecule, although significant progress in this area has been made during the past few years. The pioneering studies of Alan Hall and co-workers, using fibroblasts as a model system, demonstrated that Rho family GTPases are master regulators of actin organization (Jaffe and Hall 2002). These GTPases also control actin-based processes in growth cones (reviewed in Kuhn et al. 2000). Results from recent studies by many groups investigating receptors for both attractive and repulsive guidance cues have demonstrated that these receptors are often linked, either directly or indirectly, to one of these GTPases (Song and Poo 1999; Forcet et al. 2002; Ming et al. 2002). Downstream of these GTPases are a number of kinases that can alter the activity of actin and tubulin binding proteins and regulate cytoskeletal dynamics (Banerjee et al. 2002; Geneste et al. 2002).
Recent studies also suggest that local translation of mRNA in axons and dendrites can also play a key role in regulating growth cone navigation. In response to repulsive guidance cues, local protein synthesis appears to be needed to bring about the morphological changes in growth cone structure in response to sema 3A or netrin-1 (Campbell and Holt 2001) or adaptation to gradients of netrin-1 (Ming et al. 2002). Other studies suggest that the synthesis of new receptors can alter growth cone responsiveness to local cues and thus aid in altering a growth response (Brittis et al. 2002). Such studies imply that the targeting and delivery of specific mRNAs to the growth cones (Bassell et al. 1998) can regulate growth cone behavior.
The known molecular mechanisms involved in growth cone motility and neurite extension provide several possible modes for regulation of growth cone navigation. Extracellular guidance cues could be presented in patterns whereby only a single response (attraction or repulsion) of the growth cone is elicited. In regions where mixed cues are present, growth cones must integrate these signals into a single growth response. Such integration could occur at several levels, including the following: (a) localization, activity, or expression of guidance cue receptors; (b) signal transduction from the receptor to the Rho family GTPases; (c) regulation of the kinases and phosphatases downstream of the GTPases; (d) regulation of proteins that control cytoskeletal dynamics; (e) regulation of vesicle formation and delivery; (f) targeting specific mRNAs to growth cones; (g) regulation of growth cone adhesion; and (h) activation and recruitment of the contractile motors. There is some compelling evidence that involves each of these steps.
The papers that follow are review articles based on a symposium at the Sixth Joint Meeting of The Japan Society of Histochemistry and Cytochemistry and The Histochemical Society held in Seattle, Washington in July, 2002. These reviews focus on some of the areas described above. They start with a focus inside the growth cone looking at the molecules and pathways regulating the actin cytoskeleton, the role of myosin II isoforms in growth cone activity, and the role of SNARE proteins in neuronal outgrowth. They then address how neurotrophic factors and repulsive cues signal growth cones and conclude with a look at how an extracellular gradient of repulsive cues affects growth cone behavior.
Footnotes
Acknowledgements
Supported by the Alzheimer's Association (grant IIRG-01-2730) and the National Institutes of Health (grants GM35126 and NS40371).
I thank Barbara Bernstein, O'Neil Wiggan, and Patrick Sarmiere for valuable discussions and comments on this article.