
Abstract
Duffy antigen is a chemokine binding protein expressed on the surface of erythrocytes and postcapillary venular endothelial cells. It binds selective CXC and CC chemokines with high affinity. Although Duffy antigen is present in the normal pulmonary vascular bed, it is not known whether its expression is altered by innate inflammatory responses in the lungs. We studied Duffy antigen expression by immunohistochemistry in autopsy lung specimens from 16 cases of suppurative pneumonia, 11 cases of acute lung injury, and seven normal lungs. In lungs with suppurative pneumonia, Duffy antigen was expressed in higher numbers of pre- and postcapillary parenchymal vessels compared to normal specimens or specimens with acute lung injury (p<0.03 and p<0.02, respectively). Lungs with suppurative pneumonia also showed Duffy antigen expression on the alveolar septa, whereas this was a rare finding in normal specimens or in acute lung injury (p<0.02). Furthermore, Duffy antigen labeling of the alveolar septa localized to regions with airspace accumulation of neutrophil-rich exudates. In summary, Duffy antigen expression is increased in the vascular beds and alveolar septa of the lung parenchyma during suppurative pneumonia, suggesting that Duffy antigen may have a functional role in the lung parenchyma during inflammation.
T
Duffy mRNA and protein are upregulated during inflammatory conditions in the kidney, suggesting that the selective chemokine binding properties of Duffy antigen may be biologically relevant during inflammation (Liu et al. 1999; Segerer et al. 2000). A study in transgenic mice showed that targeted overexpression of the Duffy antigen in endothelial cells attenuated the angiogenic effect of the mouse CXC chemokine macrophage inflammatory peptide-2 (MIP-2) (Du et al. 2002). Dawson and colleagues (2000) showed that mice with a targeted deletion of the Duffy antigen receiving IP injections of lipopolysaccharide (LPS) have significantly increased inflammatory changes in the lungs and liver. By contrast, Luo et al. (2000) showed that Duffy-null mice have impaired neutrophil recruitment into organs such as the lungs at a later time point after a lower dose of IP LPS. Although the three findings differ, they are all consistent with the hypothesis that the Duffy antigen participates in regulating effective chemokine concentrations in tissue (Dawson et al. 2000). Although human lungs express Duffy mRNA (Neote et al. 1994) and the Duffy antigen is detectable on normal venular endothelium and type I epithelial cells of the lungs by immunoelectron microscopy (Chaudhuri et al. 1997), the relationship between Duffy antigen expression and neutrophil accumulation in the lung parenchyma during inflammation in humans has not been clearly defined.
Our goal was to determine how the tissue distribution of Duffy antigen in human lungs is modified during suppurative pneumonia, a process associated with recruitment of neutrophils into airspace units. We used a monoclonal antibody (MAb) that specifically recognizes the Duffy antigen (Segerer et al. 2000,2001; Wasniowska et al. 2002). During suppurative inflammation there was an increase in the number of Duffy positive pre- and postcapillary microvessels compared with normal lungs or specimens with acute lung injury. In the majority of lungs with suppurative pneumonia, Duffy antigen expression was also detectable on the alveolar septa. This is in contrast to normal lungs and specimens with acute lung injury, in which alveolar septal immunostaining was a rare finding. Moreover, the alveolar septal immunostaining observed during suppurative pneumonia co-localized to regions of neutrophilic airspace accumulation. The enhanced expression of Duffy antigen in the pulmonary parenchymal microvessels and the alveolar septa suggests that the Duffy antigen may have a biological role in the lungs during suppurative inflammation.
Materials and Methods
Lung Tissue Specimens
This study was approved by the Institutional Review Board of the Human Subjects Division at the University of Washington. Formalin-fixed, paraffin-embedded lung tissues were identified from autopsies performed during the years 2000–2001 at the VA Puget Sound Medical Center (Seattle, WA). Lung tissue was collected at autopsy and routinely fixed by intratracheal instillation of formalin at 20 cm H2O pressure. After inflation, the lungs were immersed in formalin, then embedded in paraffin and sectioned by standard methods. Hematoxylin- and eosin-stained slides from each lung specimen were initially reviewed by a pathologist and organized into three groups on the basis of light microscopic findings: (a) lungs with suppurative airspace pneumonitis; (b) acute lung injury; and (c) normal lungs. Specimens with the histological diagnosis of suppurative airspace pneumonitis showed extensive neutrophil infiltration of the airspaces. Specimens with acute lung injury were defined as tissue sections with evidence of parenchymal injury such as protein-rich exudates in the airspaces, intra-alveolar fibrin deposition, alveolar microhemorrhage, or hyaline membrane formation, in the absence of a prominent neutrophilic airspace process. When the patterns overlapped, as in cases where there was evidence of airspace accumulation of neutrophils in a few locations on the slide and therefore only a minor component of the pathology, the specimens were classified according to the predominant histological finding. Normal lungs showed no evidence of an inflammatory cell infiltrate or parenchymal injury. A total of 16 cases of suppurative pneumonia, 11 cases of acute lung injury, and seven cases of normal lungs were studied.
Immunohistochemistry
Formalin-fixed, paraffin-embedded lung tissue samples were cut into 4-μm sections, deparaffinized in xylene, and rehydrated in a series of graded ethanol. Microwave treatment in citrate-based buffer was used for antigen retrieval (Vector Labs; Burlingame, CA). Endogeous peroxidases and biotin were blocked using 3% H2O2 in methanol and an avidinbiotin blocking kit (Vector Labs). The specificity of MAb αFy6 (clone 2C3) used in this study was determined by immunoblotting, and epitope mapping showed that it binds to the epitope 24FEDVW28 located on the amino-terminal extracellular domain of the Duffy glycoprotein (Segerer et al. 2000; Wasniowska et al. 2002). An irrelevant monoclonal IgG1 isotype antibody (Dako Diagnostics; Glostrup, Denmark) was used as the control primary antibody. A biotinylated horse anti-murine IgG served as the secondary antibody (Vector Labs). A commercially available peroxidase-conjugated streptavidin complex was used for signal amplification (ABC Elite kit; Vector Labs). The color substrate was formed using diaminobenzidine and nickel chloride. Slides were counterstained with methyl green.
Morphometric Analysis
The number of immunoreactive pre- and postcapillary vessels in the lung parenchyma in each tissue section was scored semiquantitatively by counting the total number of immunostained vessels in five high-powered fields (X200). The number of immunostained vessels was normalized to the number of alveoli in each high-powered field to yield the number of immunostained pre- and postcapillary parenchymal vessels in each alveolar unit. High-powered fields were selected sequentially from a predetermined list of 31 random x,y coordinates. High-powered fields that did not contain more than 75% lung parenchyma were excluded from the scoring. Mean scores were derived from the data of two observers. Each specimen was also scored for the presence or absence of Duffy immunostaining on alveolar septal capillaries (absent = 0, present = 1). Review of slides was performed by three independent observers, including the clinical pathologist who performed the autopsy. In cases where there was disagreement about the presence of alveolar septal immunostaining, specimens were scored as “1” if two observers agreed on its presence. Nonparametric analysis was performed using the Mann-Whitney U-test. No patient identifiers were linked to the tissue samples, consistent with the guidelines of the Institutional Review Board. Therefore, demographic or clinical information could not be obtained, and only autopsy-related diagnoses were available.
Results
The analysis showed two regions in which Duffy immunostaining was readily detectable: the bronchiolar region and the lung parenchyma. The immunostaining in the lung parenchyma was subdivided into (a) the pre- and postcapillary microvessels and (b) the alveolar septa. The terminology of arteriole vs venule was not applied in describing the vessels of the lung parenchyma because vessels less than approximately 50 μm in diameter lack well-defined smooth muscle layers, preventing a clear distinction between arterioles and venules. Precapillary vessels were defined as all vessels located within parenchymal units with diameters greater than those of capillaries. These were often found at the junction of two or more converging alveolar septa (Figures 1A and 1B, arrowheads). Postcapillary vessels were identified by location outside parenchymal units within interlobular septa (Figure 1C, black arrowheads).
Alveolar Septal Immunostaining
Whereas Duffy antigen was detectable in the majority of specimens with suppurative pneumonia (Figure 2B), it was rarely detectable on the alveolar septa of normal lungs or specimens with acute lung injury (Figures 1D–1F, arrows). Semiquantitative analysis showed a significant relationship between the presence of Duffy antigen labeling on the alveolar septa and suppurative pneumonia (Figure 3). The immunostaining on the alveolar septa formed many circumscribed channels that often contained erythrocytes or leukocytes in their lumens, suggesting that the immunostaining outlined capillaries rather than type I epithelial cells of the alveolar septa (Figure 2E, arrows). Within the same lung specimen, the distribution of alveolar septal immunostaining tended to occur in alveoli with neutrophil accumulation (Figure 2D), whereas, in alveoli with relative neutrophil sparing, alveolar septal immunostaining was not present (Figure 2C).
Specimens with acute lung injury, characterized by protein-rich exudates in the airspaces that reflect the loss of integrity to the alveolar capillary epithelial membrane, showed almost complete absence of Duffy antigen labeling in all vessels throughout the parenchyma (Figure 1E). Although other specimens in the lung injury group showed Duffy antigen expression on pre- and postcapillary vessels, alveolar septal staining was absent in the majority of cases (Figure 1F). In normal lungs, Duffy antigen labeling of the alveolar septa was not detectable in 6/7 specimens. The normal lung specimen showing Duffy antigen-positive alveolar septal staining was obtained from a patient with a postmortem diagnosis of clostridial infection with gas-filled cystic lesions of the liver. Unlike the patchy alveolar staining detected in lungs with suppurative pneumonia, the alveolar septal immunostaining was uniformly distributed throughout the lung tissue (not shown).
Pre- and Postcapillary Microvessel Immunostaining
Semiquantitative data showed that the number of Duffy-positive pre- and postcapillary microvessels was significantly higher in lungs with extensive airspace accumulation of neutrophils (Figure 4). In general, specimens in the normal or acute lung injury group showed only low-level reactivity for Duffy antigen that had a patchy distribution in pre- and postcapillary vessels (Figures 1D and 1F).
Bronchiolar Vessel Immunostaining
Normal lung tissue showed constitutive expression of Duffy antigen in the small and medium-sized bronchiolar vessels. The presence of parenchymal inflammation did not appear to influence the expression of Duffy antigen in the vessels surrounding the small airways (data not shown).
Discussion
The purpose of this study was to define the expression of the Duffy antigen in vivo during lung inflammation. The main focus was to determine whether enhanced expression of the Duffy antigen occurs in the lung parenchyma during suppurative pneumonia, a process associated with neutrophil accumulation in the airspaces. In normal lungs there was low-level Duffy antigen expression in the pre- and postcapillary vessels of the lung parenchyma. In both normal lungs and lungs with acute lung injury, Duffy antigen expression on the alveolar septa was infrequent. In contrast, lung specimens with suppurative pneumonia showed increased Duffy antigen expression in precapillary and postcapillary vessels, as well as the alveolar septa.
This interpretation is based on the following semiquantitative analysis. First, the number of pre- and postcapillary vessels showing Duffy antigen labeling was higher in specimens with suppurative pneumonia compared with the acute lung injury or normal groups (p<0.02 comparing suppurative vs acute lung injury; p<0.03 comparing suppurative vs normal). Second, the number of lung specimens in the suppurative pneumonia group containing alveolar septal immunostaining was significantly higher than either the acute lung injury group or the normal group (p<0.02). In addition, in each specimen alveolar septal staining was localized to areas of neutrophil accumulation. Finally, the expression of Duffy antigen during suppurative pneumonia was specific to the parenchymal vessels, and no increase in labeling was identified on bronchiolar vessels. Taken together, these findings show an association between suppurative pneumonia and an increase in Duffy antigen expression in the parenchymal vascular beds and the alveolar septa.
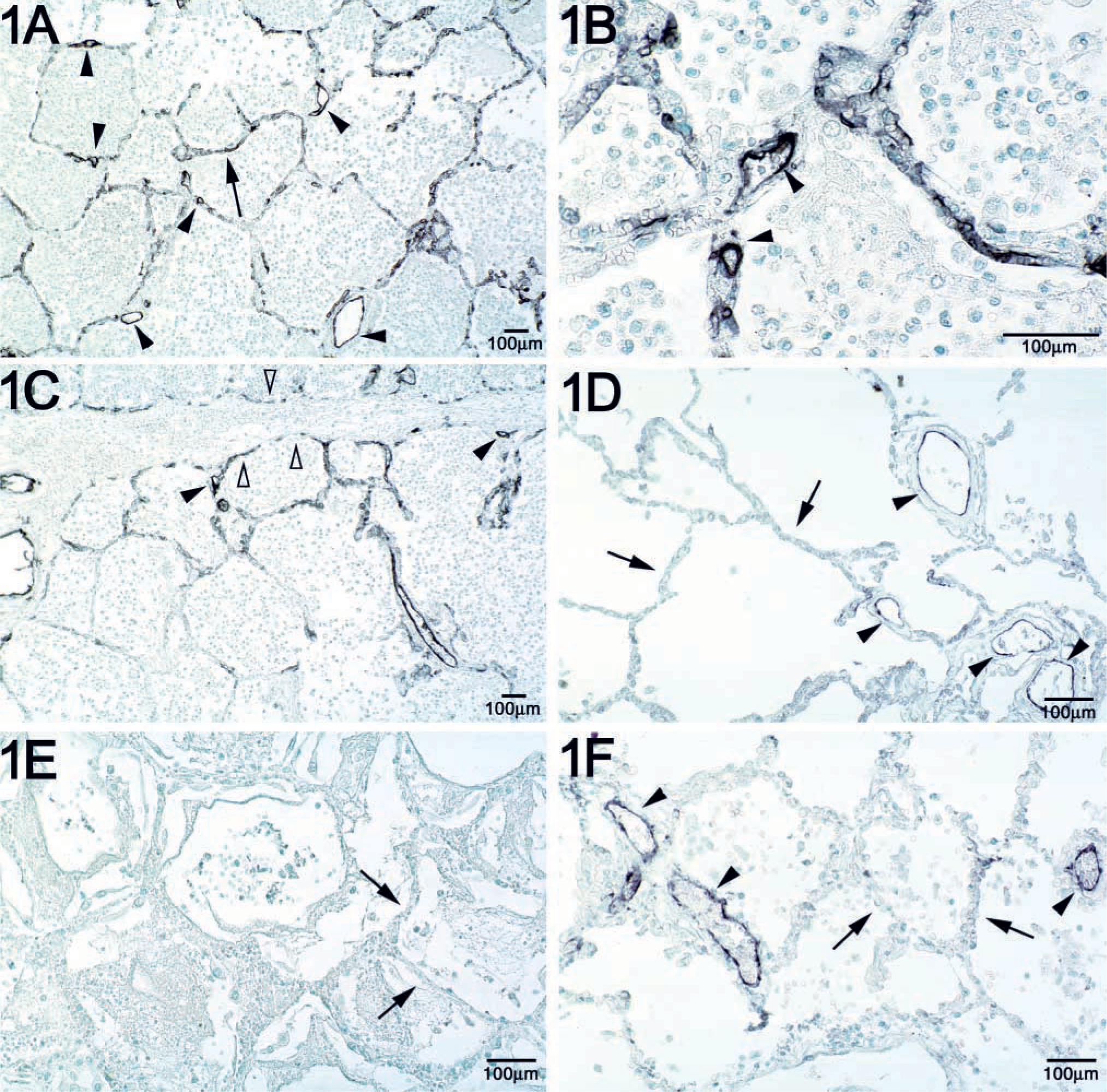
Duffy antigen expression on lung parenchymal pre- and postcapillary vessels. Immunohistochemistry was carried out using MAb αFy6. Duffy antigen staining is reflected by the black color product. Arrows indicate the alveolar septa, and black arrowheads indicate pre-or postcapillary vessels. (
Although an increase in Duffy expression has been detected during inflammatory states of the kidney (Liu et al. 1999; Segerer et al. 2000,2001), no information is available about the effect of inflammation on Duffy antigen expression in the lungs. We chose to focus on suppurative pneumonia as an example of acute inflammation not only because of its clinical importance but because of the neutrophil-predominant infiltration of the airspaces. We hypothesized that the expression of Duffy antigen, which binds CXC chemokines, is enhanced at sites of neutrophil migration in the lungs. The data show detectable constitutive expression of Duffy antigen on pre- and postcapillary parenchymal vessels and bronchiolar vessels but not on the alveolar septa. The number of pre- and postcapillary vessels positive for Duffy antigen increases during suppurative pneumonia. There is also a significant relationship between the presence of Duffy antigen labeling on the alveolar septa and suppurative pneumonia.
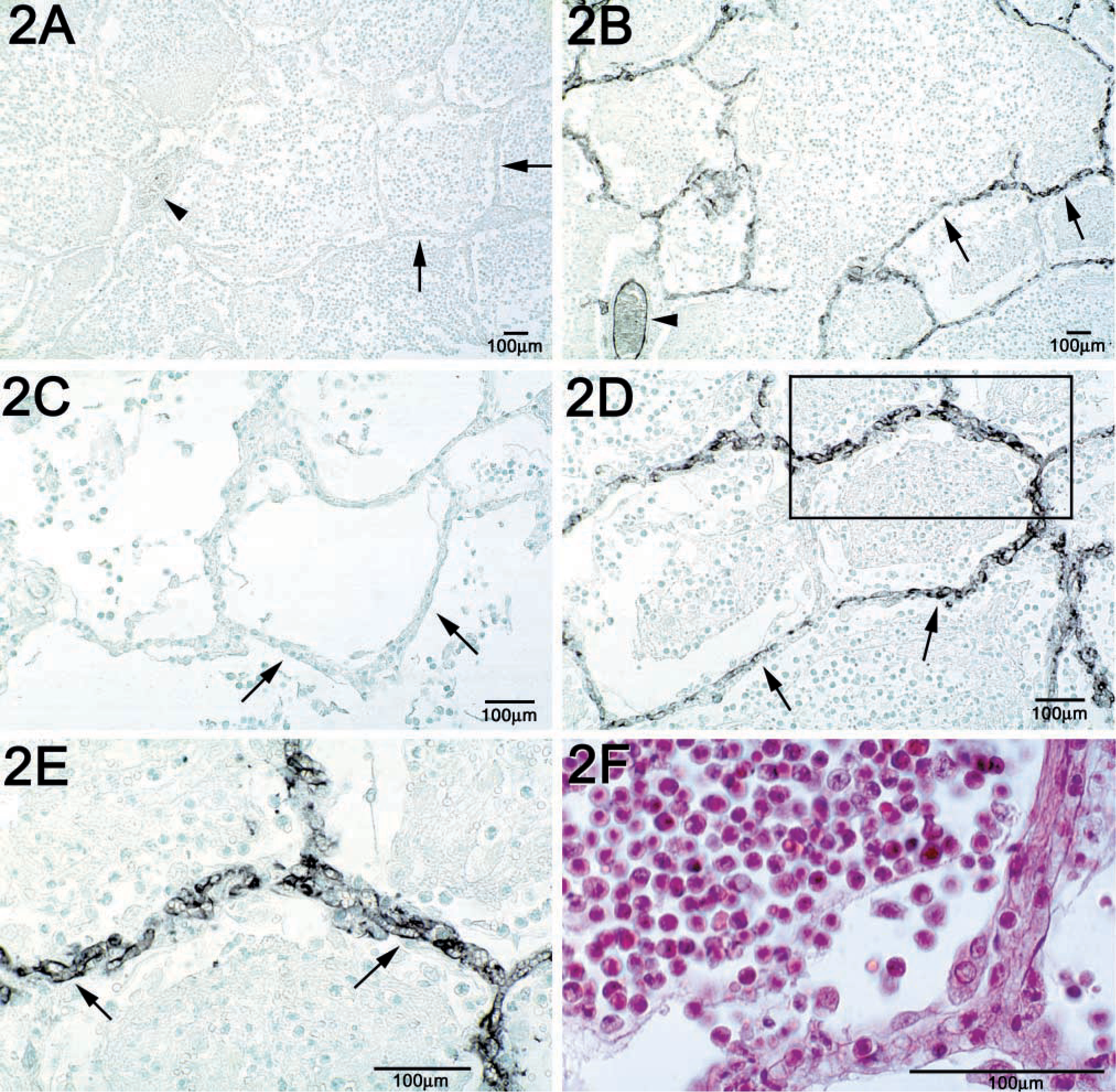
Duffy antigen is expressed on alveolar septa during suppurative pneumonia and co-localizes to areas of prominent neutrophilic airspace accumulation. (
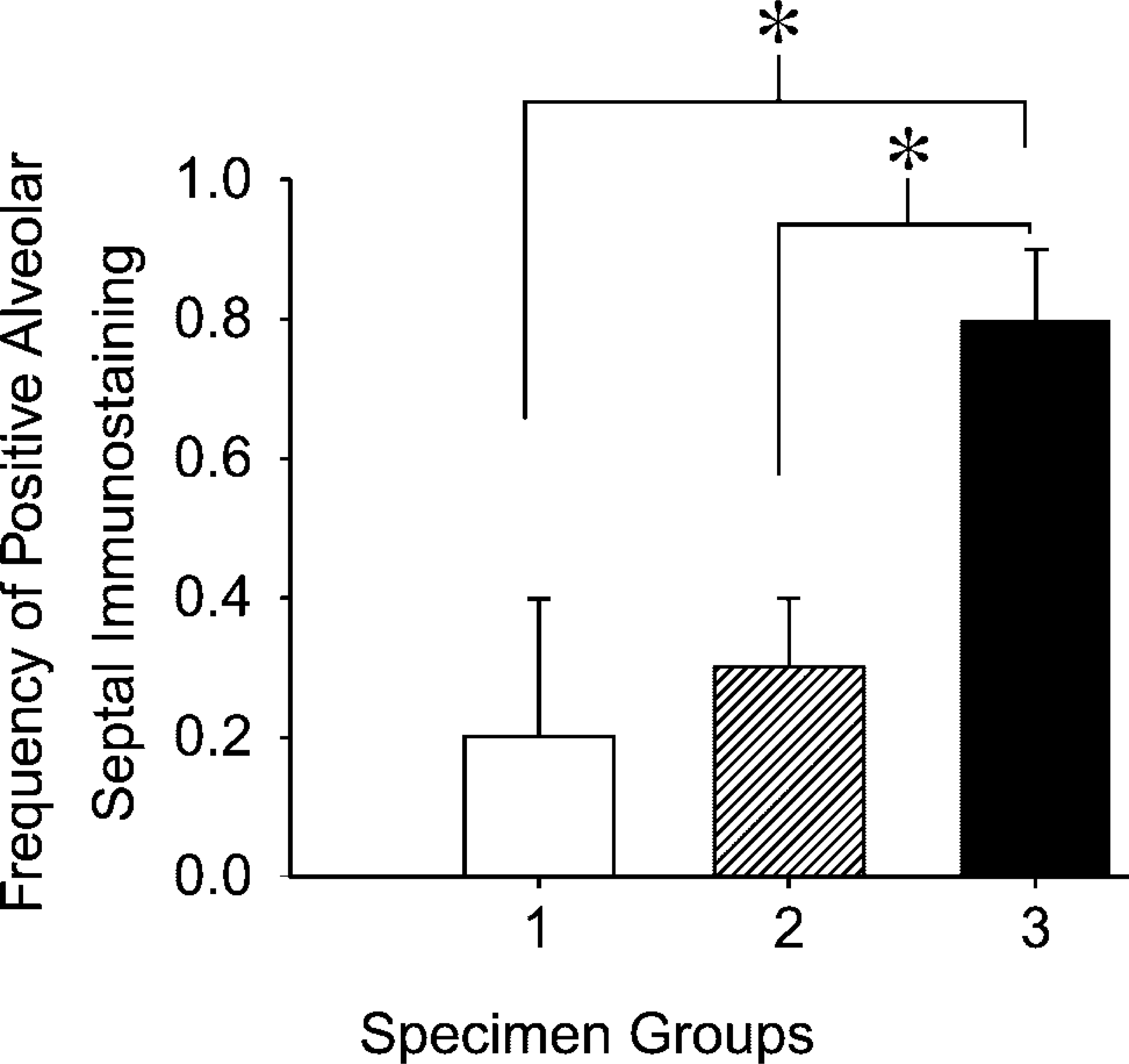
Alveolar septal immunostaining occurs more frequently in lungs with suppurative pneumonia. Specimen groups include group 1 (normal lungs), group 2 (acute lung injury), and group 3 (suppurative pneumonia). The data are the percentage of specimens in each group with immunostaining along the alveolar septae. ∗ p<0.02 using the Mann-Whitney U-test comparing suppurative with normal and suppurative with acute lung injury.
Our finding is the first report of enhanced expression of the Duffy antigen in the pulmonary parechymal microvessels and the alveolar septa during lung inflammation. Others have shown that Duffy antigen is expressed on bronchiolar capillaries, venules, and type I epithelial cells in the normal lung using immunoelectron microscopy and a polyclonal antibody directed against the carbohydrate portion of the Duffy antigen (Chaudhuri et al. 1997). Our light microscopic analysis suggested that positively stained alveolar septal cells have the appearance of capillaries rather than type I alveolar epithelial cells, based on their tubular shape in some cuts and the presence of erythrocytes and leukocytes within their lumen. Because the limitation of the study is that an endothelium-specific marker was not used to confirm alveolar septal immunostaining as endothelial in origin, we cannot conclusively determine that the alveolar septal immunostaining observed represents alveolar capillaries as opposed to type I epithelial cells. Nevertheless, these studies demonstrate increased Duffy antigen expression in suppurative pneumonia and suggest that the Duffy protein can be regulated during inflammation.
There was variability in Duffy antigen expression within individual specimens in each group, and disease variability may account for some of the differences. Clinical information such as severity of disease, duration of illness, and co-morbid conditions was not available, and only diagnoses made from postmortem examination of tissue were available for this study. Interestingly, one specimen obtained from a patient with widespread clostridial infection diagnosed at autopsy showed intense Duffy antigen expression throughout the parenchymal vascular bed and the alveolar septa despite histologically “normal” lungs. This stands in contrast to all other “normal” lung specimens that showed low-level Duffy antigen staining on vessels and absence of staining on the alveolar septa. Although the lungs from the patient were histologically normal, it is tempting to speculate that a soluble mediator(s) in the circulation may have stimulated enhanced Duffy expression.
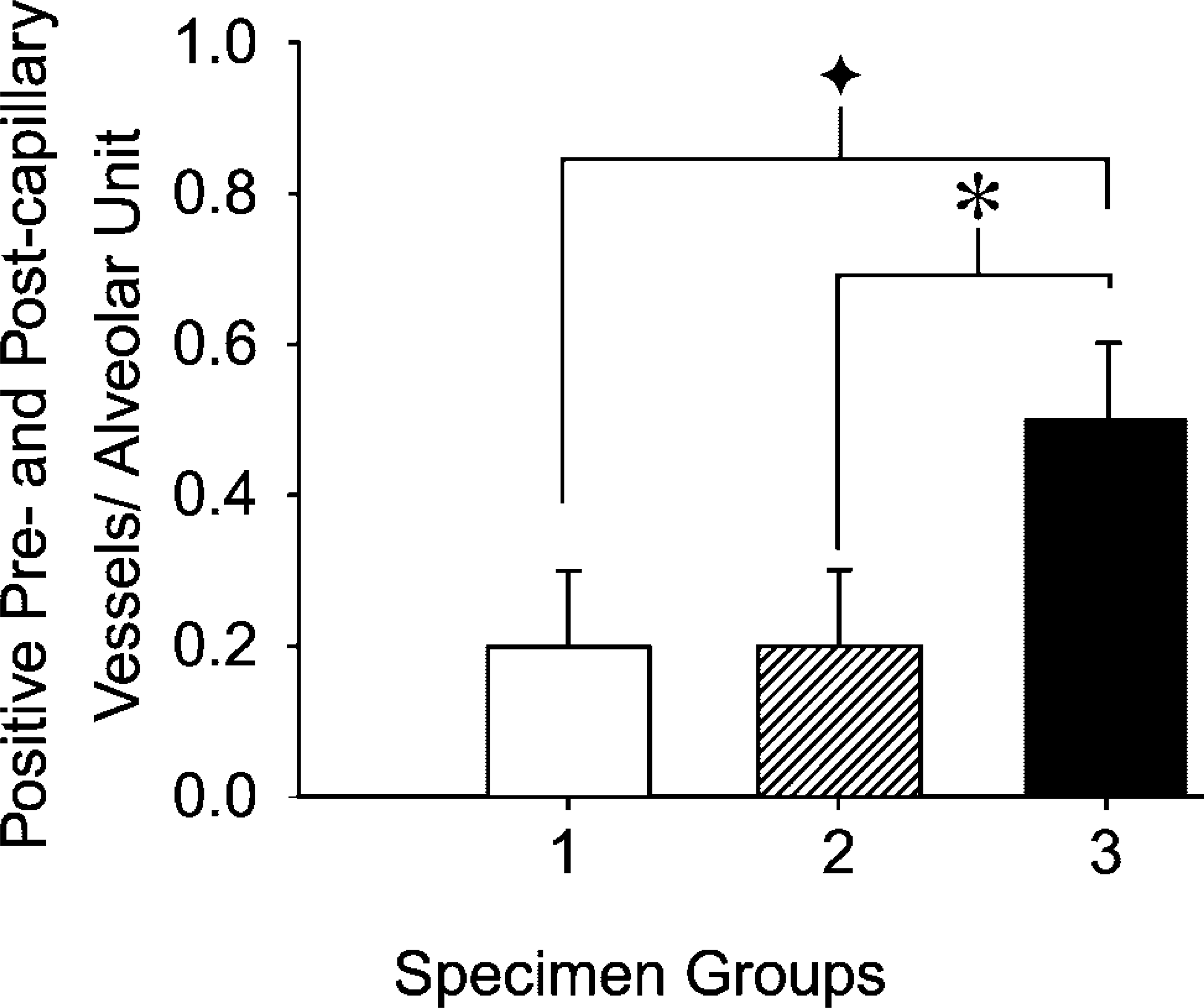
The number of positively staining pre- and postcapillary microvessels is higher in lungs with suppurative pneumonia. Specimen groups include group 1 (normal lungs), group 2 (acute lung injury), and group 3 (suppurative pneumonia). ∗ p<0.03 comparing suppurative with normal, and ∗ p<0.02 comparing suppurative with acute lung injury using the Mann-Whitney U-test.
Polymorphisms in the Duffy antigen locus could also account for some of the variability observed. The red blood cells (RBCs) of individuals homozygous for the Fyx polymorphism (Arg89Cys) have only 10% of Duffy expression and 25% binding of [125I]-IL-8 compared with RBCs of individuals with the more prevalent Fya or Fyb polymorphisms (Tournamille et al. 1998; Yazdanbakhsh et al. 2000). Because the Fyx polymorphism results from a point mutation within the coding region of the Duffy gene, the nonerythroid cells should also show reduced expression and chemokine binding (Olsson et al. 1998; Parasol et al. 1998; Tournamille et al. 1998; Wasniowska et al. 2002).
In contrast, the GATA1 promoter mutation that renders erythrocytes Duffy-negative in the majority of African-Americans is a mutation occurring in the erythroid-specific promoter region and would not alter Duffy expression on nonerythroid cells (Olsson et al. 1998; Peiper et al. 1995; Tournamille et al. 1995). Of the 34 specimens examined, three were Duffy-negative on erythrocytes but positive on vessels, and one was Duffy-negative on erythrocytes with extremely low-level to no staining on vessels. It is possible that the three specimens with negative staining on erythrocytes but positive staining on venules were obtained from individuals with the GATA1 promoter mutation that effectively silences Duffy expression on erythrocytes but not on nonerythroid cells (Tournamille et al. 1995). It is also possible that the single specimen lacking erythrocyte staining and showing near-absent levels of staining on vessels was derived from an individual with the Fyx mutation. This lung section showed diffuse alveolar damage consistent with acute lung injury (Figure 1E), and the near-absent immunoreactivity of this lung section could be explained by the specimen originating from an individual with the Fyx mutation. Therefore, the Fyx but not the GATA1 mutation can account for some variability in the nonerythroid Duffy antigen expression among individual specimens of a particular group. However, this accounted for a relatively minor number of samples in our study (1/34 samples). This frequency agrees with the 2.5–3.5% frequency of the Fyx mutation found in the White population reported in the literature (Olsson et al. 1998; Yazdanbakhsh et al. 2000).
Two in vivo animal studies have shown that Duffy-knockout mice can exhibit differential responses to an inflammatory stimulus such as lipopolysaccharide (LPS), depending on the dose, timing of the stimulus, and the focus of the experimental design (Dawson et al. 2000; Luo et al. 2000). This attests to the complex role that Duffy antigen may play during inflammation. Dawson and colleagues (2000) observed an exaggerated neutrophil influx into the liver and lungs 2 hr after an IP injection of LPS (30 mg/kg) in Duffy-null mice. Their work suggests that the absence of the chemokine sink effect by circulating Duffy-null RBCs may promote distant organ inflammation when such systemic inflammatory doses of LPS are administered. However, Luo and colleagues (2000) observed impaired neutrophil recruitment into the peritoneum, intestine, and lungs 24 hr after an IP injection of LPS (10 mg/kg) in Duffy-null mice. They found similar results when thioglycolate was used as the inflammatory stimulus. The findings of Luo and colleagues suggest that Duffy antigen may participate in the neutrophil migratory process. This is in keeping with the concept that, on endothelial cells, Duffy antigen may serve as a mediator of chemokine transcytosis, thus facilitating neutrophil recruitment (Middleton et al. 1997). The biological role of enhanced Duffy antigen expression in the lung microvasculature during suppurative pneumonia remains undefined and could play an anti-inflammatory role in the lungs by binding excess chemokines and might reduce chemokine availability to neutrophils. Alternatively, enhanced Duffy antigen expression could serve a proinflammatory role in the lungs by participating in chemokine transport across the endothelium, as others have suggested (Peiper et al. 1995; Chaudhuri et al. 1997; Middleton et al. 1997), and thereby facilitate neutrophil recruitment into organ-specific sites.
In the lungs, Duffy antigen is constitutively expressed at low levels on pre- and postcapillary vessels of the parenchyma. During suppurative pneumonia there is increased Duffy antigen expression in the parenchymal microvessels and the alveolar septa. These observations suggest that expression of Duffy antigen is regulated in the lung microvasculature by the inflammatory process and that enhanced expression of Duffy antigen may play an important role in chemokine mediation of neutrophil recruitment in the lungs during suppurative pneumonia.
Footnotes
Acknowledgements
Supported by the VA Medical Research Service (HL10470, HL70178, HL69955, HL030542, HL63652, and DK47659).
We gratefully acknowledge Jeanna Strout and Eden Palmer for their assistance with the image preparation.