
Abstract
Pax2 has been identified as a key regulatory protein associated with renal developmental malformations. The purpose of this study was to determine whether Pax2 protein expression, and that of other proteins important for normal renal development, is abnormally distributed in the prenatal kidney of the Brachyrrhine (Br) mouse that displays heritable renal hypoplasia. Embryonic 3H1 +/+ and Br/Br mice were collected between E11.0 and E18.0. Routine light microscopy and immunohistochemical analysis using antibodies to Pax2, E-cadherin, fibronectin, laminin, and Type IV collagen were applied to sequential tissue sections. E-cadherin stained consistently in the renal tubules of both normal and mutant animals. Whereas the initial expression of Pax2 corresponded between normal and mutant kidneys, it became progressively limited to the nephrogenic zone in +/+ animals, while distributing erratically in the Br/Br kidney. Fibronectin was not expressed in the normal nephrogenic zone but remained abundantly distributed throughout the Br/Br kidney. Luminin and Type IV collagen staining revealed a deficiency in renal vasculature formation in Br/Br kidneys. Results suggest that initial morphological differentiation occurs normally in the Br kidney but that subsequent nephric formation is associated with abnormal distribution of Pax2 and ECM proteins. (
T
Many regulatory genes have been identified that appear to play a major role in kidney morphogenesis. Pax2, in particular, regulates nephric differentiation (Dressler et al. 1990; Dressler and Douglass 1992; Phelps and Dressler 1993). Pax2 protein is expressed in the developing metanephric blastema and epithelial cells at the time of ureteric bud invasion and becomes confined to the cortex, where it is seen within the nephrogenic zone and terminal ampullae (Dressler and Douglass 1992). Dressler et al. (1993) showed that Pax2 is required for normal kidney development, and persistent expression of this regulatory gene prevents subsequent differentiation of renal epithelial cells. Several kidney abnormalities result, including multifocal microcystic tubule dilations with protein-filled lumina, atrophic glomeruli, hypertrophic Bowman's capsules, and capillary abnormalities. Pax2 also appears to play a direct role in maintaining cystic renal epithelia (Ostrom et al. 2000).
Kidney abnormalities typically occur in offspring of irradiated mice and many defects have been reported (Little and Bagg 1924; Bagg 1925; Carter 1959; Center 1977). These include renal hypoplasia with reduced number of glomeruli or complete renal agenesis (Brown 1931; Carter 1959). The Br mouse is one such mutant, resulting from overexposure to γ-irradiation (Searle 1966). In addition to renal hypoplasia, the craniofacial region of the adult Br mouse is characterized by malformations that arise early in development (Lozanoff et al. 1994; Ma and Lozanoff 1993, 1996). The abnormal phenotype probably results from a dominant point mutation that is characteristic of radiation-induced malformations (Selby and Selby 1978). However, the mechanism underlying these malformations remains unclear. In light of the important role of Pax2 in renal development, it is possible that the mutation seen in the Br mutant involves expression of this protein. The purpose of this study was to determine whether abnormal morphological expression of Pax2 protein occurs in the developing Br kidney. In addition, the expression of E-cadherin, laminin, fibronectin, and Type IV collagen was delineated to determine whether abnormal distribution of these important renal proteins is associated with the Br mutation.
Materials and Methods
All procedures were carried out in accordance with IACUC specifications and performed according to procedures approved by the Laboratory of Animal Services, University of Hawai'i. Adult C3H/He × 101/H Br/+ (3H1 Br/+) mice were housed under standard conditions with a 12-h light cycle. They were supplied with tapwater and Rodent Mouse Chow ad libitum. Br/Br and +/+ embryos were obtained through reciprocal crosses between 3H1 Br/+ adults. Females were examined for a vaginal plug starting at 0800 hr and checked hourly over a 12-hr period. If none was present, the females were removed and re-mated the next day. The day on which a vaginal plug was observed was designated as Day 0 of gestation (E0). Pregnant females were sacrificed by cervical dislocation at 12-hr intervals between E11.0-E14.0 or at 24-hr intervals after E14.0 until 18 days of gestation. Embryos were removed from the uterus and rinsed in PBS (pH 7.4). Heads were removed and the body was retained for transverse sectioning. Chucks were suspended in isopentene and frozen in liquid nitrogen, and tissues were mounted using OCT. Tissue blocks were positioned in a cryostat (IEC-CTF) and serial sections were cut at 8 μm. Sequential sections were placed on five different gelatinized slides with four sections on each slide and air-dried for 1 hr. One of the sequential sections was stained with either toluidine blue or hematoxylin and eosin for anatomic reference.
Light Microscopy
For anatomic reference, kidneys from both 3H1 +/+ and Br/Br mice were processed for routine light microscopy. Frozen sections were collected (as above), fixed with 10% neutral buffered formalin (NBF), and stained with either toluidine blue or hematoxylin and eosin. Additional specimens were collected for plastic embedding and sectioning. Specimens were collected (as above) and fixed in NBF for at least 1 week. Specimens were dehydrated in a graded series of ethanol (50%, 70%, 95%, 100%) and embedded in plastic (JB-4 embedding kit). Blocks containing the tissues were cut in a transverse plane at 6 μm on a Sorvall (JB-4) microtome. Sections were stained with hematoxylin and eosin.
E-cadherin and Pax2
Tissue sections were fixed with 100% acetone at −20C for 10 min. Sections were washed twice (5 min each) with PBST (PBS and 0.05% Tween-20) at room temperature (RT). The antibodies used included 10 μg/ml of polyclonal rabbit anti-Pax-2 IgG (gift from Dr. G. Dressler) and rat anti-E-cadherin (Sigma; St Louis, MO) in 2% goat serum in PBST. After incubation of 1 hr in a humid chamber at RT, slides containing the sections were washed twice in PBST, 5 min for each wash. Secondary antibodies were diluted at 1:32 for FITC-conjugated goat anti-rabbit antibody (Sigma) and 1:20 for FITC-conjugated goat anti-rat antibody (Sigma) in 2% goat serum in PBST. Slides with tissue sections were incubated in a moist chamber for 30 min at RT, washed twice in PBST (5 min each) and coverslipped using Citifluor mounting medium (Marivac; Halifax, Canada). Tissues were viewed and photographed using a Zeiss fluorescence microscope equipped with a digital camera. Two sets of negative controls were processed in a similar fashion. In one set, PBS was substituted for the primary antibody. In the second set, same-species antiserum was substituted for the primary antibody. No qualitatively significant staining appeared on negative controls.
Fibronectin, Laminin, and Type IV Collagen
After sectioning, tissues were fixed in 100% methanol at −20C for 10 min. Tissue sections were washed for 5 min in PBS with 0.2% Triton X-100 (Sigma). Nonspecific binding of antibodies was blocked by incubation for 10 min with 2% skim milk at RT. Sections were flooded with PBS and primary antibodies were applied. Primary antibodies used in this study included anti-fibronectin (Calbiochem; La Jolla, CA), anti-laminin (Sigma), and polyclonal anti-collagen Type IV (ICN; Covina, CA). All primary antibodies were applied at a concentration of 1:100 in PBS and incubated in a moist chamber overnight at 4C. The following day, sections were warmed to RT and washed twice for 5 min in PBS. The secondary antibody was FITC-conjugated swine anti-rabbit IgG (Dako; Carpinteria, CA) at a concentration of 1:100 in PBS for all primary antibody preparations. Slides were incubated for 30 min at RT and washed in PBS. Tissue sections were coverslipped using Citifluor mounting medium and were then viewed and photographed. Two sets of negative controls were processed (as above for E-cadherin and Pax2), and qualitatively significant staining did not appear on negative controls.
In Situ Detection of Apoptosis
Kidney tissue sections were derived from normal and Br/Br mice at gestational days 12–14, dewaxed in xylene, and rehydrated through a graded series of ethanol and distilled water according to standard methods. A deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick endlabeling (TUNEL) assay was performed using the in situ cell death detection POD kit (Roche; Indianapolis, IN). The tissue sections were incubated with proteinase K–Tris-HCl 10 nM, pH 7.6 (20 μg/ml), for 30 min at 21C. The slides were incubated with a blocking solution (3% H2O2 in methanol) for 10 min at RT, followed by 2-min incubation at 4C with a 0.1% Triton X-100/0.1% sodium citrate solution. The tissue sections were labeled with the TUNEL reaction mixture as described by the manufacturer. The signal conversion was obtained using the converter POD system provided in the in situ cell death detection POD kit. The DAB substrate was added to the tissue sections for 5 min at RT to reveal the presence of the horseradish peroxidase. The stained slides were then viewed under a light microscope. A positive control included adult murine gut tissue. Negative controls consisted of sequential renal tissue sections that were processed on the same slide as treated experimental tissues. These negative controls were processed equivalently but they received the labeling solution without the terminal transferase.
Results
Gross Morphology and Light Microscopy
The external appearance of the gross kidney from newborn 3H1 +/+ mice showed a smooth surface with no irregularities (Figure 1A). The kidney from a typical 3H1 Br/+ mouse was smaller, with a rough and irregular surface indicative of multifocal cyst formation (Figure 1B). The Br/Br kidney was extremely hypoplastic, approximately the size of the adrenal gland (Figure 1C). The external surface also was rough in appearance, consistent with the formation of multifocal cyst formation in underlying renal tubules. Only +/+ and Br/Br kidneys were used in this study.
Histological examination of the normal developing ureteric bud as it initially branched in to the metanephric blastema at E11.5 showed aggregating cells surrounding the developing tubule (Figure 2A). Similarly, the E11.5 Br/Br kidney showed initial condensation of the surrounding mesenchyme (Figure 2B). Whereas the E12.0 +/+ kidney showed a continuing trend of mesenchymal condensation around the branching metanephric tubule system (Figure 2C), the developing Br/Br kidney lacked mesenchymal condensation (Figure 2D). Mesenchymal condensation continued at E13.0, with the +/+ kidney showing marked aggregation of the peritubular mesenchyme as well as an emerging medullary area of less condensed tissue (Figure 2E). The Br/Br kidney did show a condensing capsular area but peritubular mesenchyme remained loose, with minimal condensation (Figure 2F).
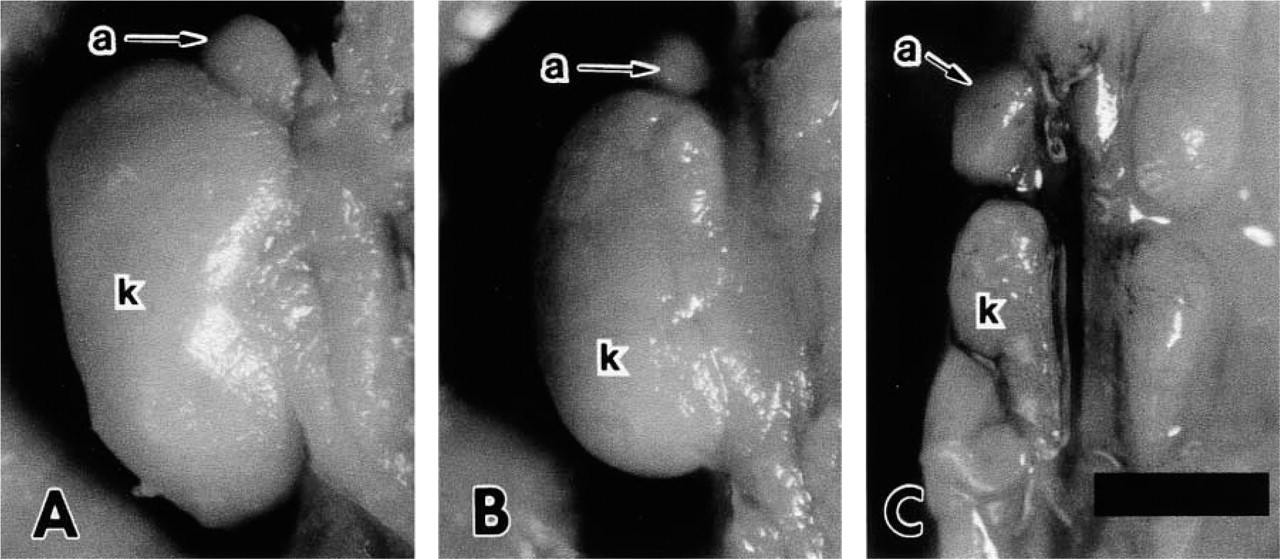
Kidneys (k) of +/+ (
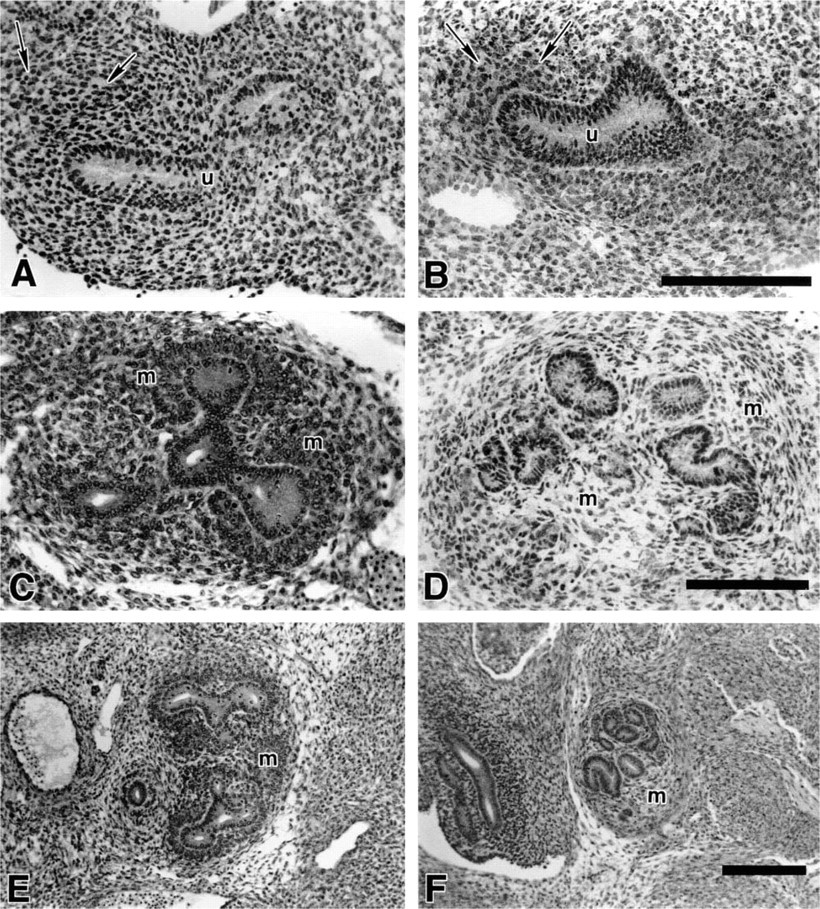
Light micrographs of the differentiating ureteric bud and metanephric blastema in +/+ and Br/Br mice at E11.5 (
Subsequent development of the normal kidney (E13.0) was marked by a well-distinguished nephrogenic region of the developing cortex, with an emerging medullary region lacking cellular condensation (Figure 3A). The Br/Br kidney continued to lack peritubular condensations and appeared greatly reduced in overall size as well as tubule number (Figure 3B). At E15, the normal kidney demonstrated a condensing nephrogenic zone at its periphery, with an abundance of branching tubules as well as a medullary region with qualitatively reduced cell number and density (Figure 3C). At this same developmental stage, the Br/Br kidney lacked a nephrogenic zone peripherally (Figure 3D). Although tubules were present, they appeared to lack an extensive branching pattern. The neonatal normal mouse displayed a well-defined nephrogenic zone peripherally (Figure 3E). At this point, the Br/Br kidney lacked a nephrogenic zone and displayed severe polycystic kidney disease (Figure 3F). Proximal tubules were severely distended and appeared to contain amyloid. Although glomeruli were present, they appeared less abundant than in the normal animal. Vascular distension was present, and the epithelium of some cysts appeared disrupted.
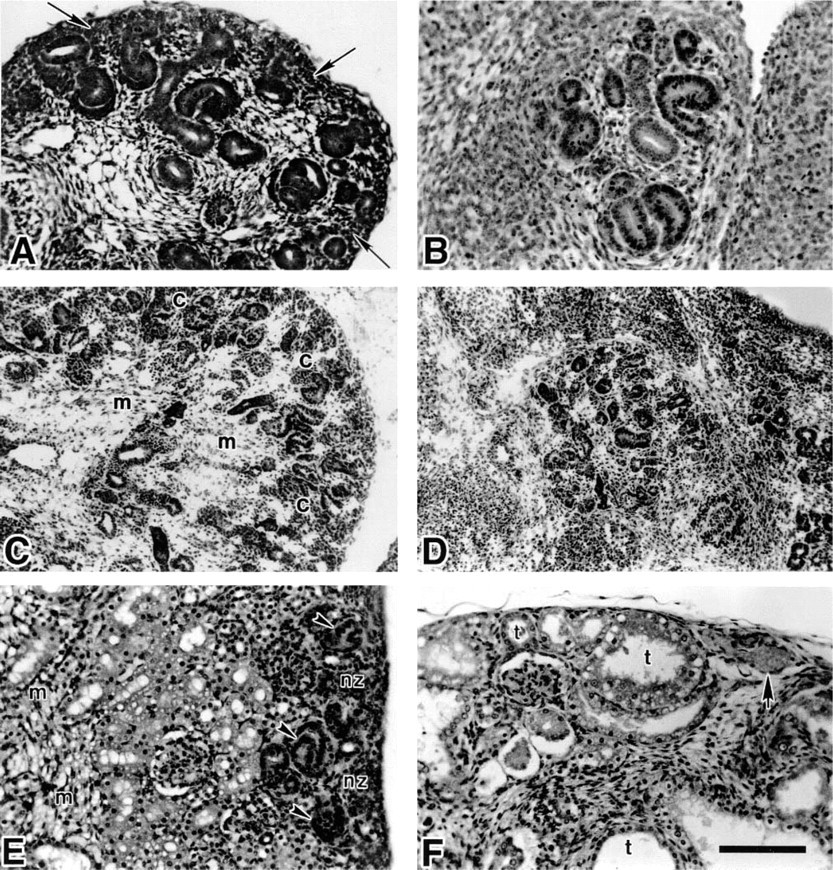
Light micrographs of renal development in +/+ (
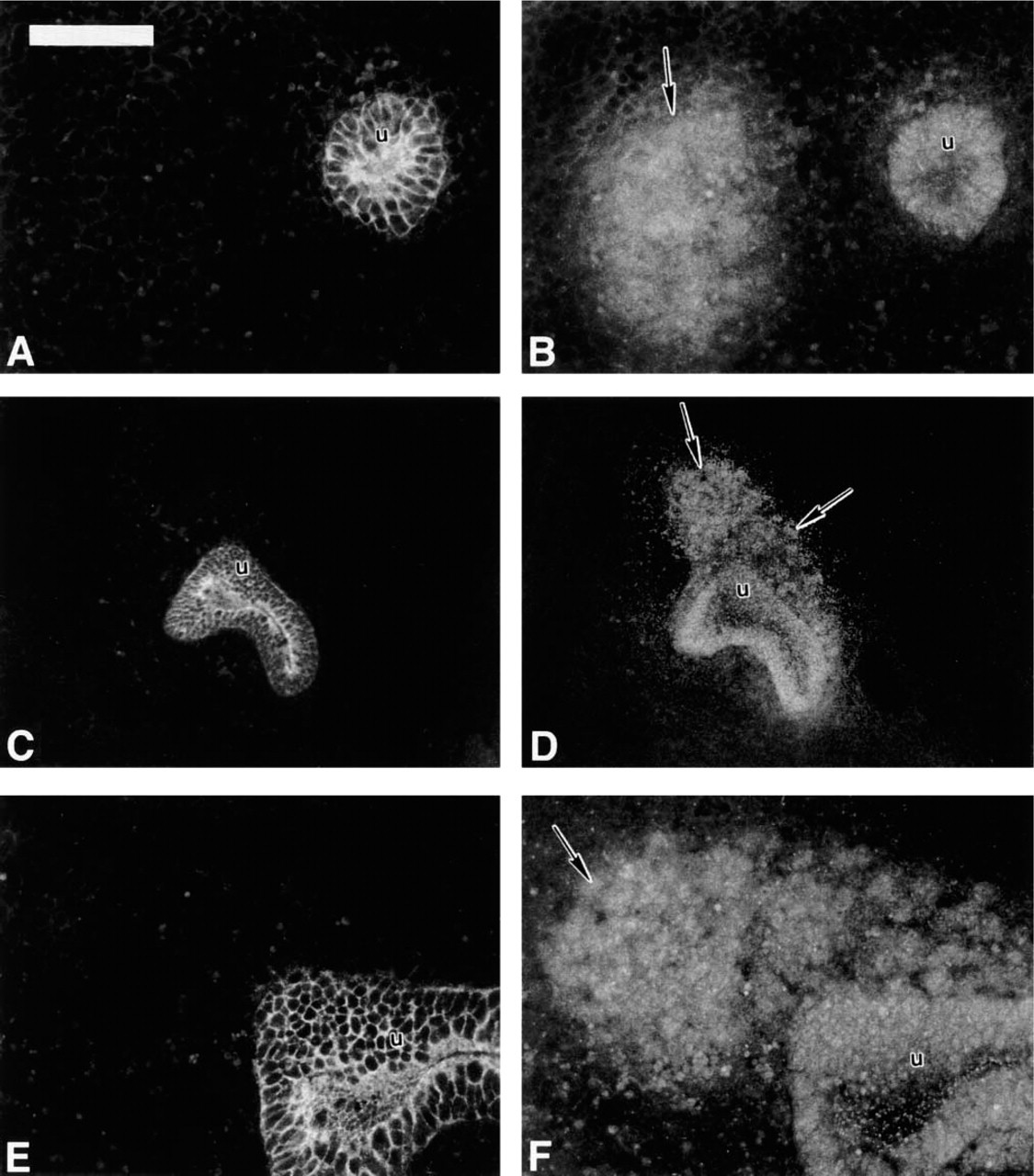
E-cadherin (
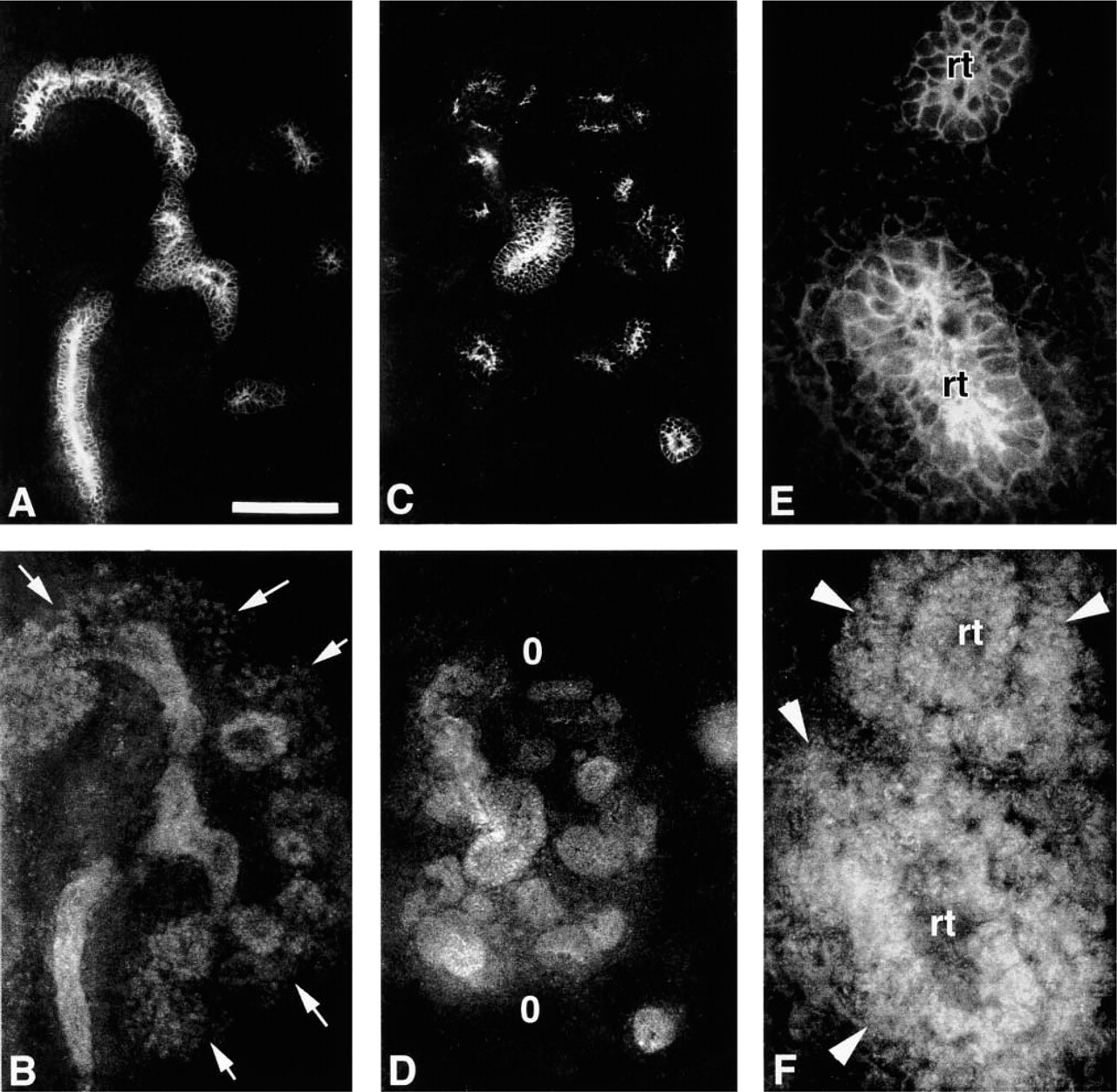
Distribution of E-cadherin and Pax2 in the normal (
E-cadherin and Pax2
Differentiation of the renal epithelium was characterized by a thick layer of E-cadherin along the borders of the renal tubule cells that became less pronounced abluminally in the +/+ mouse during the early stages of nephric development (Figure 4A). The Br/Br kidney showed a similar pattern of E-cadherin expression (Figures 4C and 4E). Just before nephric differentiation at E11.5 in the +/+ mouse, the expression of Pax2 was present in both the tubule epithelium and the peritubular condensate adjacent to the metanephric tubule (Figure 4B). Similarly, Pax2 was present in the renal tubules and peritubular metanephric blastema of the E11.5 Br/Br kidney during this initial induction period (Figures 4D and 4F).
E-cadherin and Pax2 distribution in E15 normal (
Distribution of E-cadherin continued to be concentrated along the luminal surface of the developing renal tubule system for both +/+ and Br/Br animals at E13.0 (Figures 5A, 5C, and 5E). The pattern of Pax2 expression appeared to differ, with +/+ animals showing a well-defined nephrogenic region peripherally in addition to Pax2 localization within the developing tubule network (Figure 5B). Pax2 localized in the renal epithelium of the E13.0 Br/Br kidney, but was conspicuously absent within the peripheral area of the kidney, reflecting a poorly developed nephrogenic zone (Figure 5D). Pax2 was still expressed in the mesenchyme of the Br/Br kidney, but staining was limited to peritubular areas (Figure 5F).
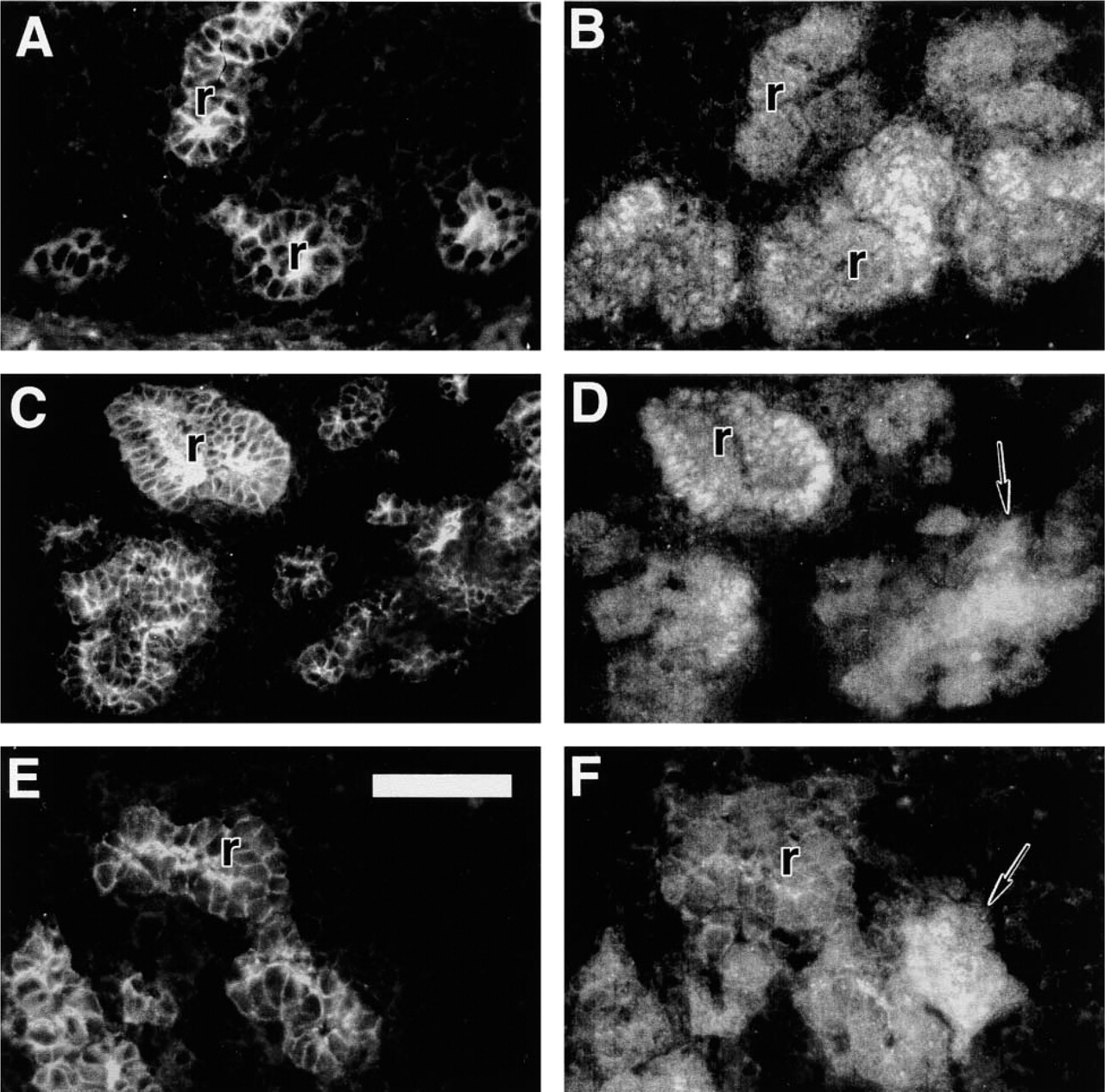
E-cadherin (
At E15, E-cadherin was prominently expressed in the developing renal tubules of both normal and mutant mice (Figures 6A, 6C, and 6E). As with the E13.0 stage, E15 +/+ mice displayed Pax2 prominently in the nephrogenic zone (Figure 6B). However, the E15 mutant kidney lacked a nephrogenic zone and Pax2 staining showed a discontinuous pattern, appearing only as infrequent clumps in some pertibular regions (Figures 6D and 6F).
E-cadherin remained prominently expressed in the developing renal tubules of both +/+ and Br mutant neonatal mice (Figures 7A and 7C). Pax2 distribution in the +/+ mouse was restricted to the cortical nephrogenic zone and newly emergent renal tubules (Figure 7B). This protein was distributed within the developing tubules and abundantly in the peritubular areas but was largely absent from more mature subcortical renal tubules, indicated by intense E-cadherin staining (Figures 7A and 7B). Pax2 distribution in the Br/Br mutants was expressed inconsistently in clumping arrangements around some renal tubules but not others (Figure 7D).
Fibronectin, Laminin, and Type IV Collagen
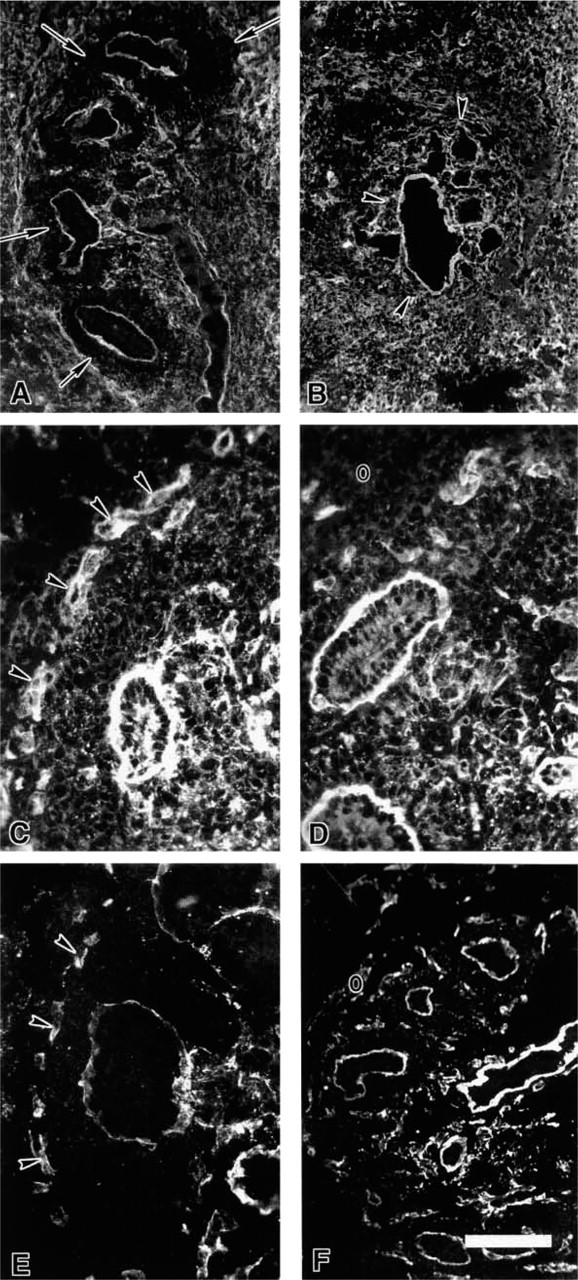
Fibronectin, laminin, and Type IV collagen were all clearly visible in the developing basement membranes of normal and mutant kidneys at E11.5 (Figure 8). Fibronectin appeared less concentrated in the condensing peritubular mesenchyme of Br/Br animals, corresponding to the normal condition (Figures 8A and 8B). Laminin formed a thick layer surrounding the developing ureteric bud while Type IV collagen was well expressed in the basement membranes of normal and mutant kidneys (Figures 8C–8F).
The distribution of fibronectin appeared to decrease in the developing cortical nephrogenic zone and particularly around the renal tubules, but increased toward the medullary interstitium in the normal animal at E13.0 (Figure 9A). However, fibronectin staining remained intense throughout the Br/Br kidney, showing no decrease in distribution cortically (Figure 9B). Although laminin and Type IV collagen were consistently expressed in the basement membranes of developing normal and mutant tubules, there was little evidence of emergent vasculature in the Br/Br nephrogenic area compared to the normal condition (Figures 9C–9F).
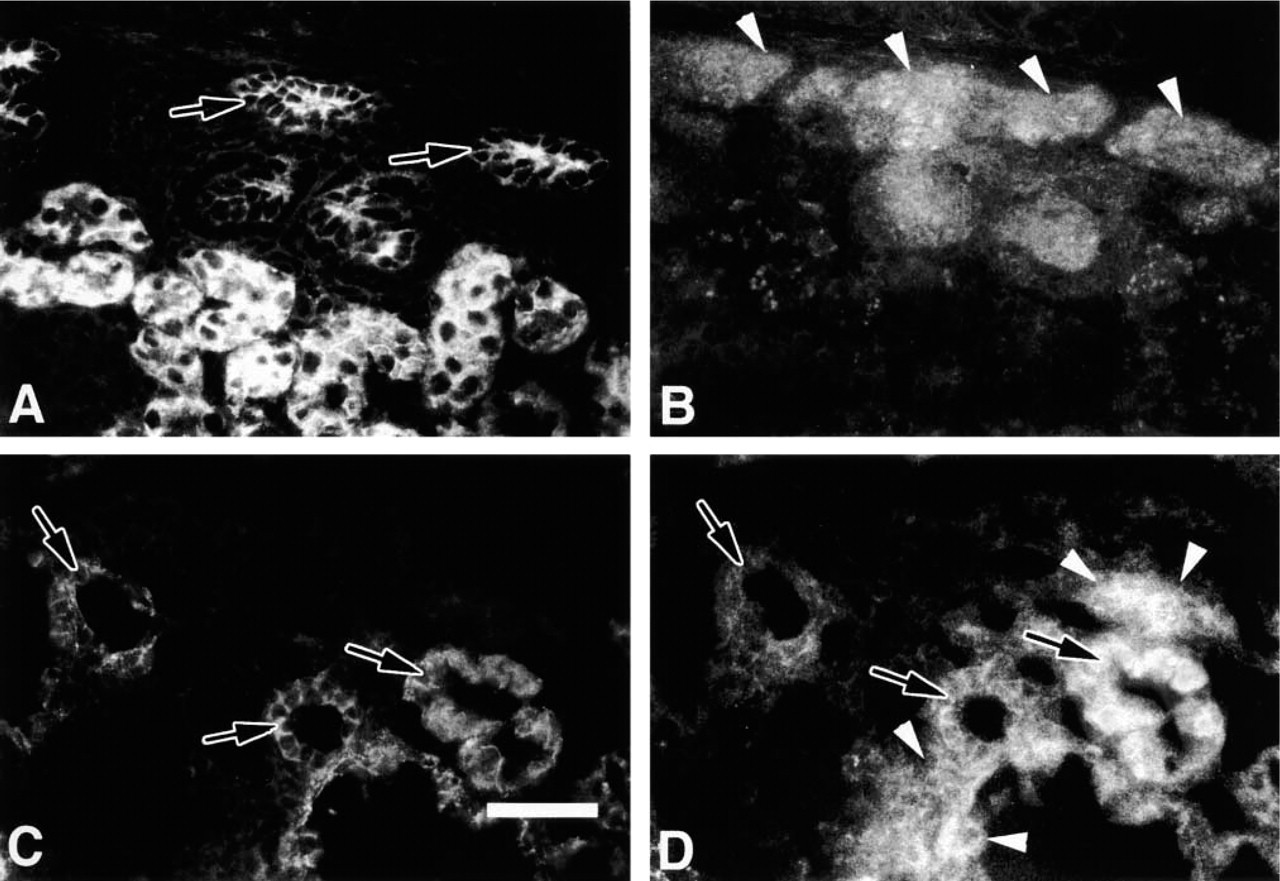
Extracellular matrix components are normally distributed in the normal (
Extracellular matrix molecule distribution in normal (
Distribution of extracellular matrix molecules at E15 in +/+ (
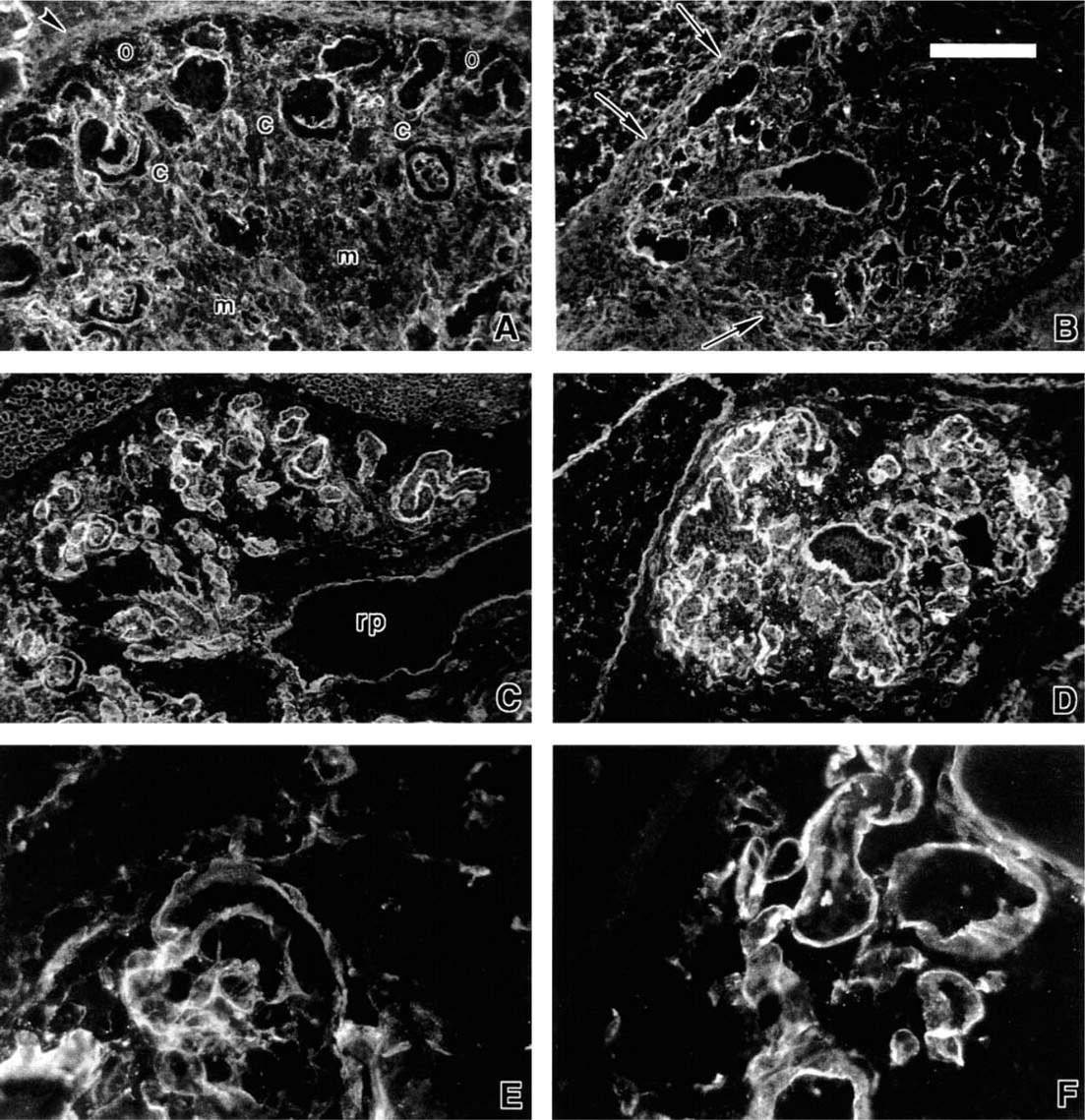
By E15, fibronectin was present in the medullary region but was absent in the nephrogenic zone located peripherally in +/+ animals (Figure 10A). It remained consistently expressed in the Br/Br kidney, with little evidence of a nephrogenic zone (Figure 10B). Both laminin and Type IV collagen remained prominently stained in the basement membrane of both +/+ and Br/Br mutants (Figures 10C–10F). In the +/+ kidney, strong laminin staining was present in the basement membrane of developing nephrons but appeared less intense in the developing renal pelvis, whereas laminin staining was consistently strong in the Br/Br kidney (Figures 10C and 10D).
Extracellular matrix molecule distribution in +/+ (
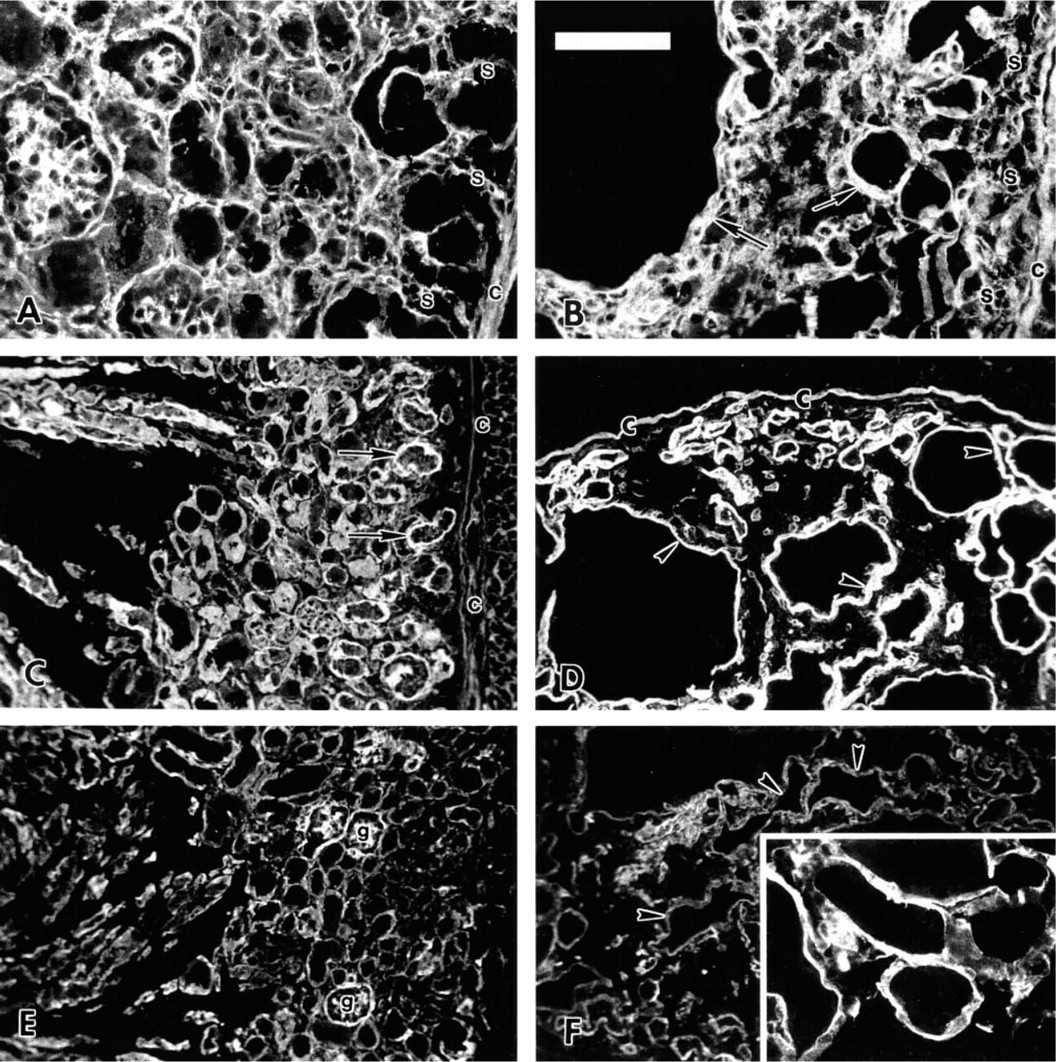
Fibronectin was sparsely distributed in the nephrogenic zone of the +/+ neonate but increased toward the medulla (Figure 11A). In the Br/Br newborn, however, fibronectin was consistently and densely dispersed throughout the entire kidney without any indication of a nephrogenic zone (Figure 11B). Laminin stained densely in the tubule basement membrane of +/+ mice, particularly in the developing nephrons located peripherally (Figure 11C). In Br/Br kidneys, laminin was particularly dense in cystic tubules (Figure 11D). Type IV collagen was also seen in the tubule basement membrane of +/+ and Br/Br neonates (Figures 11E and 11F), demonstrating a very irregular outline for mutant renal tubules, and it appeared thicker in enlarging cysts compared to normal tubules (Figures 11E and 11F). These two elements of the basement membrane were well expressed in the glomeruli of normal mice (Figures 11C and 11E), underscoring a reduced number of glomeruli in the Br/Br kidneys (Figures 11D and 11F).
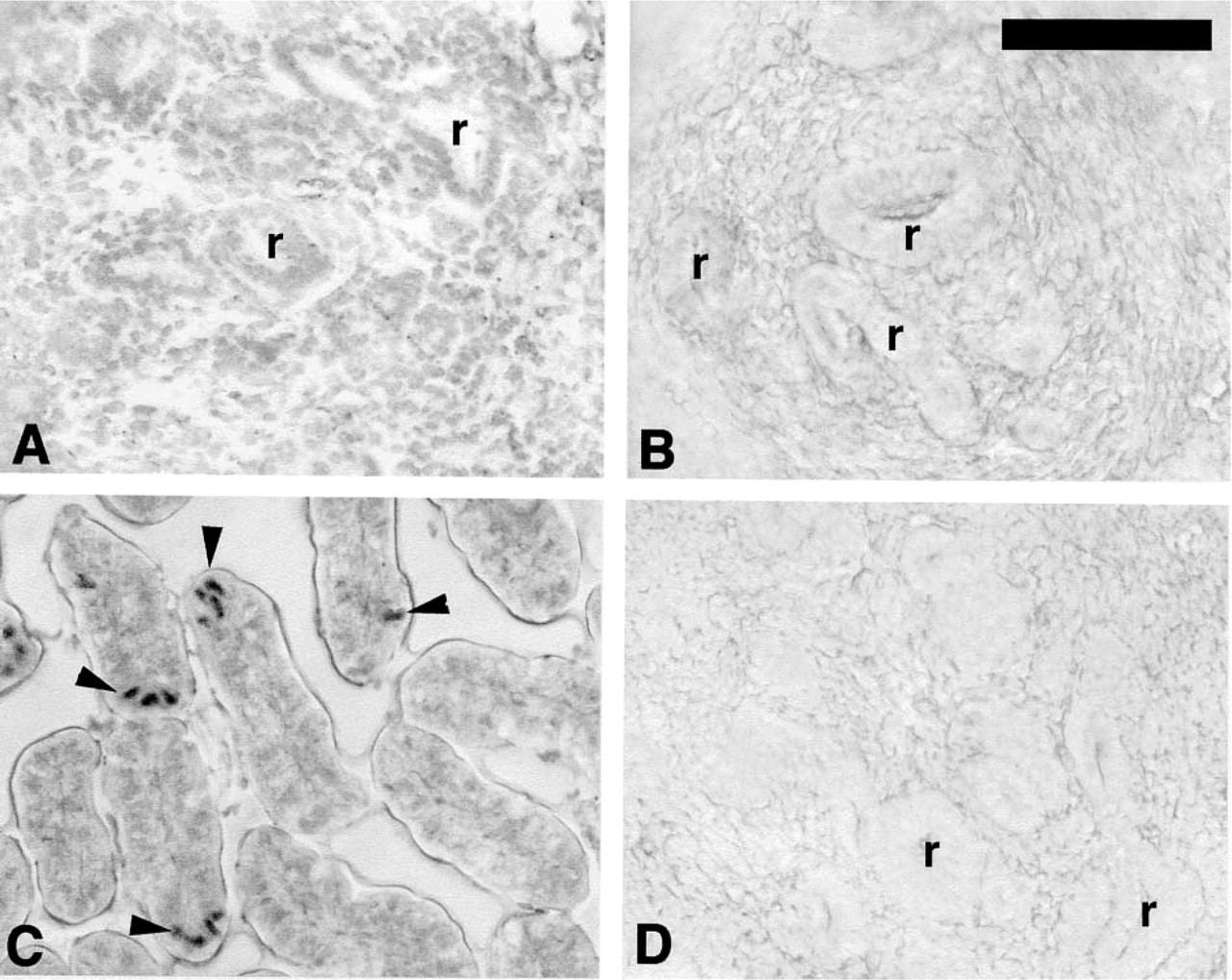
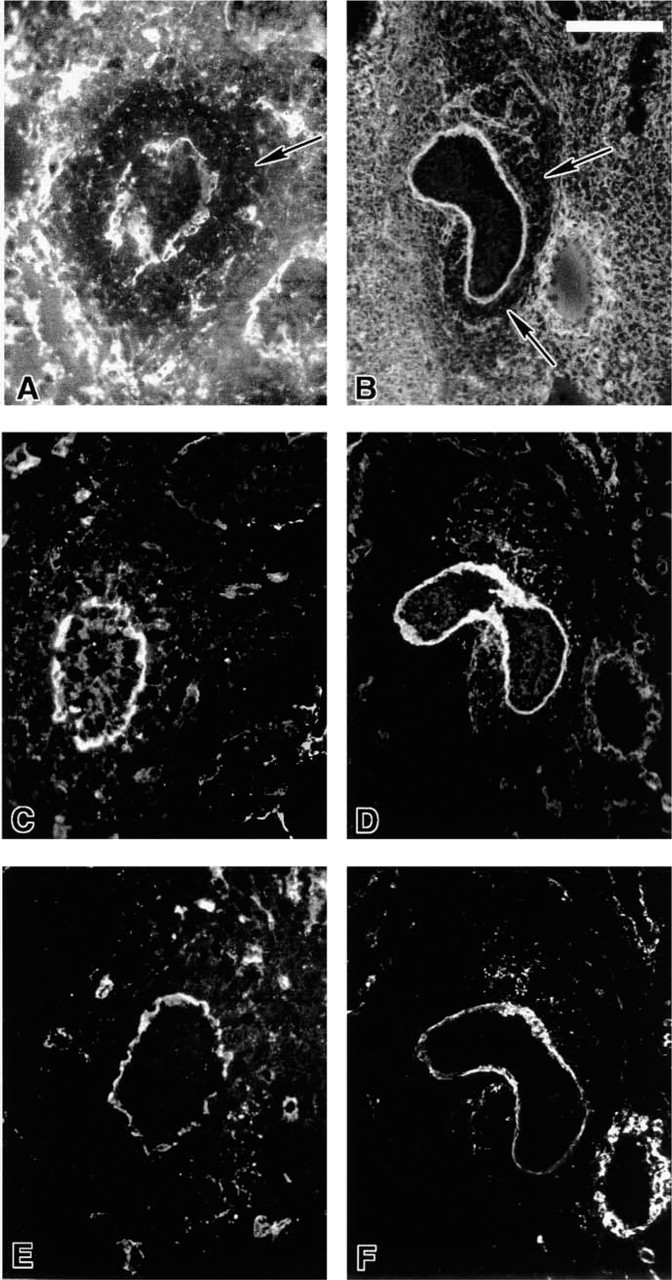
TUNEL staining of 3H1 +/+ (
Staining for apoptotic bodies revealed an absence of stained cells in both normal and Br/Br renal tissues between Days 12 and 14 (Figures 12A and 12B), even though positive controls showed clear staining in the murine gut while negative controls lacked all staining (Figures 12C and 12D).
Discussion
Epithelial cell proteins play a key role in the normal development of the kidney. E-cadherin is a transmembrane protein important for epithelial structure by maintaining and regulating cell–cell adhesion. It may play a role in epithelial morphogenesis by complexing with cytosolic catenin proteins, thereby linking it to the actin cytoskeleton and maintaining epithelial integrity (Balkovetz et al. 1997). E-cadherin staining occurred prominently along the borders of the cell membrane in both normal and mutant animals, consistent with its role as an epithelial cell adhesion molecule. It remained consistent throughout the prenatal period in the Br/Br mutants and its distribution corresponded to that seen in +/+ animals, suggesting that metanephric tubule derivatives were developing correctly throughout the prenatal period. However, ECM and Pax2 protein distributions in Br/Br animals revealed developmental disparities compared to +/+ animals.
Fibronectin, laminin, and Type IV collagen are important ECM proteins found in both the basement membrane and interstitium of the developing kidney. Ekblom (1981) showed that undifferentiated renal mesenchyme expresses fibronectin, thus promoting mesenchymal condensation. However, it is subsequently lost from the mesenchyme during the inductive process, being degraded by proteases. In this study, fibronectin was lost from the Br/Br peritubular mesenchyme at E11.5, coincident with initial differentiation of the mutant kidney. However, once this initial induction event occurred, fibronectin reappeared by E13.0 in the Br/Br kidney, where it remained intensely staining peripherally throughout the remainder of the prenatal period. Therefore, peripheral expression of fibronectin in the area where the nephrogenic zone should develop was associated with the deficient nephrogenesis seen in the mutant. Laminin and Type IV collagen are also important ECM proteins in the basement membrane of developing renal tubules. Similar to fibronectin, laminin staining decreased in the interstitium of the nephrogenic zone of the normal kidney, but basement membranes for newly emerging renal tubules maintained intense staining. Laminin in Br/Br tubules remained intensely stained throughout the kidney and was very dense in the basement membranes of cystic tubules of mutant neonates. Laminin is known to increase in the basement membrane of polycystic kidneys (Vleming et al. 1995). Therefore, the intense laminin staining may reflect the development of polycystic kidney disease in the Br mutants. Type IV collagen remained clearly visible in the mutant tubule basement membranes throughout the developmental period and appeared thicker in the enlarging cysts of Br/Br mutants, particularly later in development.
In the normal kidney, the appearance of Pax2 is a marker for renal mesenchymal cell induction, with the highest protein expression in proliferating epithelial cell precursors. Pax2 probably regulates nephron precursor cell proliferation while also modulating WT1 expression, which is required for induction and survival of the renal mesenchymal cells (McConnell et al. 1997). In this study, Pax2 expression diminished in the area of the medulla and eventually became confined to the cortical nephrogenic zone in normal animals. The Br mutants showed a normal Pax2 expression pattern at E11.5, the time of ureteric bud migration into the presumptive metanephric blastema. However, Br/Br tubule formation appeared to decrease, while peritubular mesenchyme showed limited Pax2 expression. By E13.0, cortical tissue was largely absent in Br/Br animals and Pax2 expression was limited to the medullary region, where it appeared in clumps around some of the tubules. The spatiotemporal Pax2 expression pattern as seen in this study was similar to that reported by Dressler et al. (1993), who showed that overexpression of Pax2 results in abnormal kidneys displaying multifocal, microcystic tubule dilatations, with lumina filled with protein and capillary abnormalities. The Br/Br neonatal mouse showed a very similar renal morphology, characteristic of nephrosclerosis and chronic polycystic renal disease. Although initial Pax2 expression occurs normally at E11.5 in the Br mouse, it becomes abnormally expressed during subsequent development, possibly preventing further proliferation and differentiation of renal nephric components.
Pax2 expression appeared inversely distributed compared to fibronectin. In the normal condition, fibronectin was well represented in the basement membrane and interstitium of the medulla of the kidney but diminished in the nephrogenic zone. Although fibronectin was diminished in the peritubular area at the time of differentiation in the Br/Br mutant, staining became consistently expressed in the periphery of the kidney, contrary to the diminished staining in this same area in the normal mouse. A mechanism explaining an inverse interaction between Pax2 and fibronectin has not been reported. However, it is possible that Pax2 is not available to interact with fibronectin in the Br/Br mutant kidney, because a nephrogenic zone is largely missing in the mutant. Further co-localization studies will be required to test the inverse relationship between Pax2 and fibronectin expression.
A notable absence of cortical blood vessels was evidence in the developing Br/Br mouse between E13.0 and E15 when tissues were stained for laminin and Type IV collagen. In the normal condition, a regular network of endothelial cells in the developing nephrogenic zone indicates a close relationship between nephrogenesis and vessel development (Kitamoto et al. 1997). It is possible that ureteric bud and/or metanephric blastemal cells guide migrating endothelial cells to the correct locations or stimulate differentiation of endothelial cells locally (Kitamoto et al. 1997). Disruption of nephrogenic zone formation in the Br mutant could likewise prevent necessary guidance interactions between vascular endothelial and renal epithelial cells. Because Pax2 showed widespread expression in the developing nephrogenic zone from E13.0 onwards in normal animals, its failed expression in the mutant could contribute to decreased vascularity. Further work will be required to clarify the relationship between Pax2 expression and the disruptive vascular pattern in the Br kidney.
The Br mouse showed renal tubule enlargement beginning by E15, with subsequent development of polycystic kidney disease. Neonatal animals displayed kidneys with constricted arterioles and fibrinoid tissue around glomeruli, as well as increased interstitial tissue with extremely large, dilated tubules, many containing casts. Ostrom et al. (2000) showed that Pax2 expression and gene dosage modifies the expression of renal cystic disease in young postnatal mice. These authors showed that murine Pax2 haploinsufficiency is associated with increased apoptosis and reduced cyst growth. However, our results suggest that apoptosis is not a critical factor in the development cysts at the earliest stage of development in the Br/Br mouse.
Radiation-induced mutations in mice have long been known to result in a variety of phenotypic defects involving the skeletal, nervous, urogenital, and reproductive systems (Carter 1959; Ehling 1965). The kidney is particularly prone to malformations but the specific effects are morphologically variable. Renal hypoplasia was shown to occur in the Br mouse in a heritable fashion (Ma and Lozanoff 1993), and the Br locus has been identified near the telomere on chromosome 17 (Beechey et al. 1997; Lozanoff et al. 1999). Although no known murine kidney genes exist in this region, this area also shows homology with human 2p21, an area occupied by genes associated with frontonasal dysplasia and median midfacial clefting (Schell et al. 1996). Interestingly, the Br/Br mouse also shows severe median midfacial clefting (Singh et al. 1998). It is possible that an unknown gene on murine chr17 may effect multiple transcriptional events affecting seemingly unrelated systems, as is the case for the sonic hedgehog-patched-GLI pathway resulting in facial and urogenital developmental defects (Walterhouse et al. 1999). Further work is required to map and clone the gene(s) responsible for the Br malformation to clarify this relationship.
Footnotes
Acknowledgment
Supported by AHA (Hawai'i Affiliate) 9951133Z.