
Abstract
Human prostate cancer cells (DU145) implanted into nude mice are deficient in DNase activity. After administration of a vitamin C/vitamin K3 combination, both alkaline DNase (DNase I) and acid DNase (DNase II) activities were detected in cryosections with a histochemical lead nitrate technique. Alkaline DNase activity appeared 1 hr after vitamin administration, decreased slightly until 2 hr, and disappeared by 8 hr after treatment. Acid DNase activity appeared 2 hr after vitamin administration, reached its highest levels between 4 and 8 hr, and maintained its activity 24 hr after treatment. Methyl green staining indicated that DNase expression was accompanied by a decrease in DNA content of the tumor cells. Microscopic examination of 1-μm sections of the tumors indicated that DNase reactivation and the subsequent degradation of DNA induced multiple forms of tumor cell death, including apoptosis and necrosis. The primary form of vitamin-induced tumor cell death was autoschizis, which is characterized by membrane damage and the progressive loss of cytoplasm through a series of self-excisions. These self-excisions typically continue until the perikaryon consists of an apparently intact nucleus surrounded by a thin rim of cytoplasm that contains damaged organelles.
P
The results of previous studies demonstrate that the activity of alkaline DNase (DNase I; EC 3.1.21.1) and acid DNases (DNase II; EC 3.1.22.1) is inhibited in non-necrotic cells of malignant tumors in men and in experimental animals, as well as during the early stages of experimental carcinogenesis (Taper 1967; Taper and Bannasch 1976; Fort et al. 1974; Taper et al. 1971a, b). Because alkaline and acid DNase reactivation occurred in spontaneous and induced tumor necroses and regression (Taper 1980; Taper et al. 1981), compounds capable of reactivating DNase in tumor cells should offer a great potential for therapeutic intervention in cancer. It has been found that vitamin K3 (2-methyl-1-4-naphthoquinone) selectively reactivated alkaline DNase in malignant tumor cells, whereas vitamin C (ascorbic acid, or sodium ascorbate) exclusively reactivated acid DNase (Taper 1980). A combined administration of vitamin C and vitamin K3 (at a single IP dose of vitamin C = 1 g/kg body weight and vitamin K3 = 0.01 g/kg) in ascites tumor-bearing mice synergistically increased the lifespan of the mice by 45%. Separate administration of vitamin C increased lifespan by 14.7% and vitamin K3 alone by 1.07% (Taper et al. 1987). Combined vitamin C and K3 treatment also potentiated the effects of chemotherapy induced by six different cytotoxic drugs (Taper et al. 1987), sensitized a mouse tumor that was resistant to vincristine (Taper and Roberfroid 1992), and potentiated the therapeutic effects of radiotherapy (Taper et al. 1996).
The tumor growth-inhibiting and -chemosensitizing effects of the vitamin C and K3 combination were confirmed by in vitro experiments with different lines of human tumor cells (Noto et al. 1989; De Loecker et al. 1993). These in vitro studies were extended to a battery of human urological tumor cell lines, including DU145, an androgen-independent prostate carcinoma cell line (Gilloteaux et al. 1995; Jamison et al. 1997; Venugopal et al. 1996a, b; Jamison et al. in press). In this study, we have investigated the therapeutic affect of the vitamin C and K3 combination on human prostate carcinoma cells (DU145) that had been transplanted into 5-week-old male NCr athymic nude mice.
Materials and Methods
Tumor Cells
The human androgen-independent prostate carcinoma DU145 cell line was obtained from the American Type Culture Collection (Rockville, MD) and was grown in McCoy's 5A medium (M5A; Gibco, Grand Island, NY). All media were supplemented with 10% fetal bovine serum (FBS; Gibco), 2.2 g/ liter NaHCO3 (Gibco), and 50 μg/ml gentamicin sulfate (Fisher Scientific; Pittsburgh, PA). Cells were grown as adherent cells in a humidified atmosphere at 37C in 5% CO2. When the tumor cells were used for experiments, they were harvested with trypsin-ethylenediamine tetraacetic acid (EDTA) (JRH Biosciences; Lenexa, KS), washed with PBS, pH 7.4, and resuspended in M5A. Cells were counted with a hemocytometer using trypan blue (Gibco) exclusion to determine the number of viable cells.
Reagents
Vitamin C (VC) and menadione bisulfite (VK3) were purchased from Sigma Chemical (St Louis, MO) and were dissolved in PBS to create oral vitamin doses of VC = 15 g/liter and VK3 = 0.15 g/liter. The IP doses of the vitamins were VC = 1 g/kg body mass and VK3 = 10 mg/kg body mass as optimized and described by Taper and co-workers (1987). All vitamins were prepared in a darkened laminar flow hood to prevent photo-inactivation.
Animals
Male athymic nude mice (NCr-nu/nu; 4 weeks old) were purchased from Taconic Farms (Germantown, NY) and maintained in microinsulator cages (within the AALAC-accredited NEOUCOM Comparative Medicine Unit) in a pathogen-free isolation facility. After a 1-week isolation period, 1 × 106 DU145 cells in a 100-μl volume of M5A medium were injected SC into the median and dorsal scapular region. After 4 weeks, the mice were weighed and divided into six groups of four animals each. Five of the six groups were given 100 μl of the VC/VK3 combination by gavage and then immediately given an additional 100-μl dose of the vitamin combination by IP injection. Control mice received two 100-μl doses of PBS. The mice were sacrificed by carbon dioxide inhalation at 1, 2, 4, 8, or 24 hr after vitamin administration. The tumors were then surgically removed and trisected. One third of the tumor was flash-frozen in liquid nitrogen and stored at –80C. The second third of the tumor was fixed in Carnoy's fixative for 3 hr, processed, and embedded in paraffin. The final third of the tumor was fixed in buffered glutaraldehyde and processed in epoxy resin for future electron microscopic observations as described below.
All animal procedures were performed under protocols approved by the Northeastern Ohio Universities College of Medicine Animal Care and Use Committee.
Plastic Sections
One third of each tumor from vitamin-treated and sham-treated mice was fixed in buffered glutaraldehyde solution (3.2% in 0.1 M phosphate buffer, pH 7.32) and processed for ultrastructural observations in epoxy resin (Polysciences, PolyBed 812; Warrington, PA) (Gilloteaux et al. 1995). One-μm-thick sections were cut and stained with Toluidine blue. Several of these specimens were examined, and selected micrographs were used in this presentation. Photomicrographs were taken with a Leitz and an Olympus AX 70 Provis photomicroscope. The numbers of mitotic, apoptotic, and necrotic cells were quantified by scoring 500 tumor cells per field in five random fields per tissue section. Statistical analysis was done by means of an unpaired Student's t-test to compare control and test sections.
Histochemical Detection of DNase Activity
The histochemical activity of alkaline and acid DNases was detected in cryosections of the frozen tumor specimens using a modified Gomori's lead nitrate method (Vorbrodt 1961; Taper 1968). Fresh cryostat tissue sections were fixed for 15 min in 10% formol-calcium, briefly washed in distilled water, and incubated for 3 hr at 37C in a medium that contained lead nitrate and DNA. For acid DNase activity, the reaction buffer contained 0.2 M acetate buffer, pH 5.9, and acid phosphatase. For alkaline DNase activity, the reaction buffer contained 0.2 M Tris-maleate, pH 8.0, magnesium chloride, and alkaline phosphatase. After incubation, the sections were washed for 5 min in distilled water, stained in dilute ammonium sulfide solution, washed again in distilled water, and mounted with aquamount. The sites of enzymatic activity were revealed by deposits of black lead sulfide. Control sections incubated in medium without substrate were included and exhibited no histochemical staining.
Histochemical Detection of Nucleic Acids
Nucleic acid staining was performed using the classic methyl green-pyronin technique of Brachet (1946) as modified by Godlewski and Vorbrodt (1954). The embedded tumors were cut into 5-μm sections, dewaxed in two xylene treatments, and brought to distilled water through ethanol changes (100%, 75%, 50%). The sections were stained for 2 hr with methyl green-pyronin Y in 0.1 M acetate buffer, pH 4.7, washed rapidly with water, and then dried with filter paper. The sections were dehydrated in 95% ethanol:100% isopropyl alcohol (1:7) and then in 100% isopropyl alcohol. The sections were then cleared in xylene and mounted with aquamount. Methyl green staining intensity of five fields of 100 control cells and five fields of 100 vitamin-treated cells was measured with an image analyzer and recorded in terms of optical density (Schulte et al. 1992). Statistical analysis was done by an unpaired Student's t-test to compare control and test sections.
Results
Table 1 and Figure 1 summarize the results of the histochemical determination of alkaline and acid DNase activity in the tumors at different time intervals after single-dose administration of IP and oral vitamins C and K3 or PBS administration. As shown in Table 1, the sham-treated tumors exhibited little if any alkaline and acid DNase activity. However, after combined vitamin C and K3 treatment, alkaline DNase activity appeared sooner than the acid DNase activity. This positive alkaline DNase activity was significant, appeared as early as 1 hr after vitamin administration, and decreased slightly until 2 hr after vitamin treatment. Alkaline DNase activity gradually diminished until 8 hr after vitamin administration and then disappeared. Acid DNase activity appeared 2 hr after vitamin administration, reached its highest level 4–8 hr after treatment, and maintained its activity 24 hr after treatment.
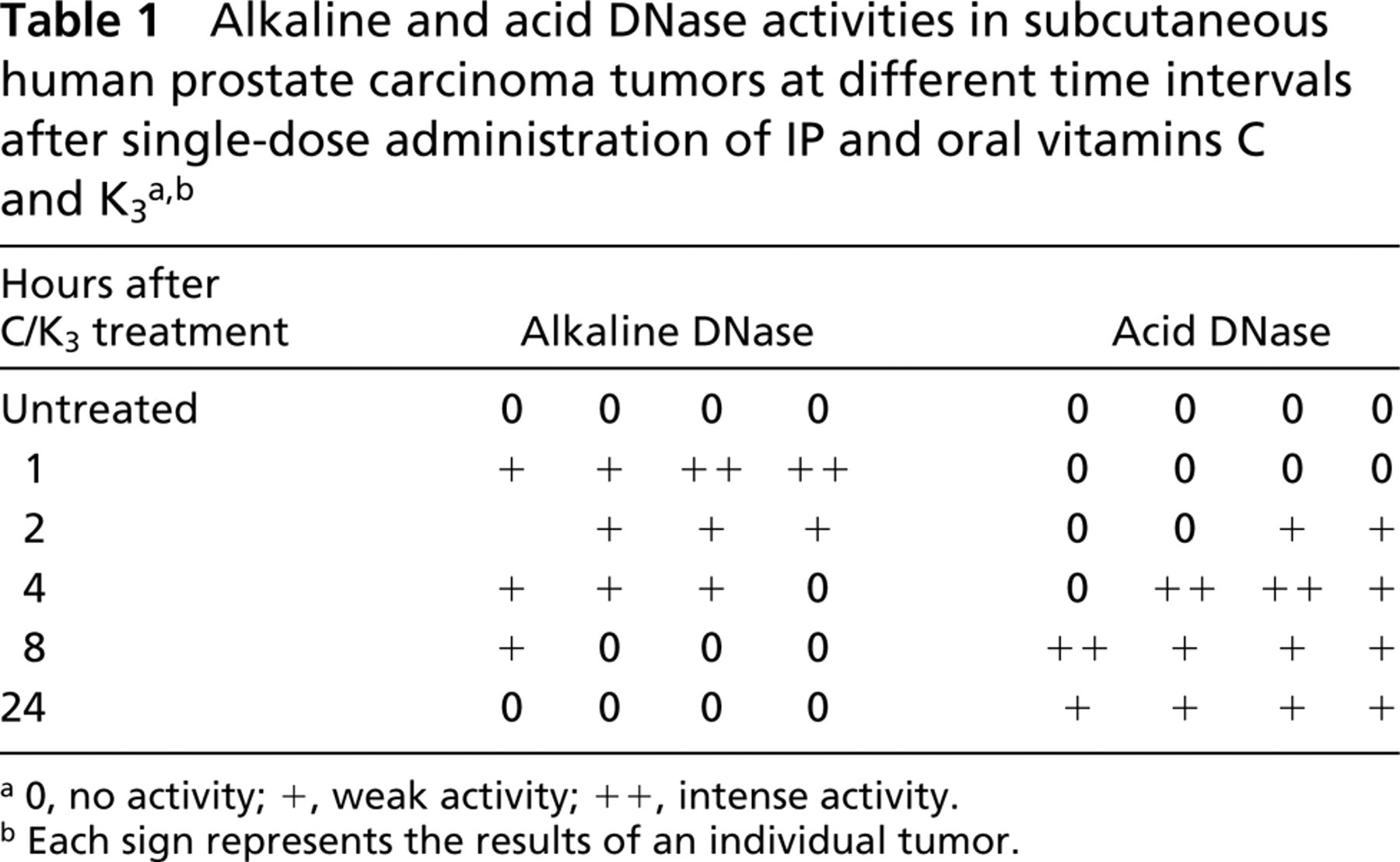
a0, no activity; +, weak activity; ++, intense activity.
bEach sign represents the results of an individual tumor.
Figure 1 is a montage of representative light micrographs of tumor sections that were stained for alkaline or acid DNase activities. One hour after combined vitamin C and K3 injection, alkaline DNase demonstrated distinct activity in the cytoplasm. However, the majority of alkaline DNase activity was found in the nuclei of tumor cells (Figure 1, 1h). As described in Table 1, alkaline DNase activity gradually decreased and was not evident after 4 hr. Very low to undetectable alkaline DNase activity was seen in the sham tumor cells (Figure 1, C). Conversely, little acid DNase activity was observed until 4 hr after vitamin treatment. At this time, intense acid DNase activity was detected primarily in the nuclei of the tumor cells (Figure 1, 4h). Acid DNase activity gradually decreased. Substantial activity was still evident 24 hr after vitamin treatment. Sham-treated tumor cells exhibited little acid DNase activity (Figure 1, C).
The consequence of the reactivation of alkaline and acid DNases could be seen in tumor cells 24 hr after vitamin C and K3 treatment. When the nucleic acids of sham-treated tumor cells were stained with methyl green-pyronin Y, intense methyl green staining was visible in the nucleus and intense pyronin Y staining was observed in the cytoplasm (Figure 2A). In vitamin-treated tumor cells (Figure 2B), the methyl green staining was greatly diminished, indicative of a decrease in DNA content. Densitometric analysis of the staining intensity of control and vitamin-treated tumor cells produced optical density values of 0.63 ± 0.11 and 0.37 ± 0.07, respectively. These values indicate a statistically significant (p < 0.01) decrease in DNA staining. Pyronin Y staining diminished at a more rapid rate and to an even greater extent than the methyl green staining, indicative of decreased RNA content. This suggests that this combined vitamin treatment may also reduce transcription or induce the activation of RNases.
Figure 3 is a set of micrographs of hematoxylin-and eosin-stained sections of sham-treated and vitamin-treated tumors 4 hr after treatment. The cells in the sham-treated tumor (Figure 3A) exhibited pleomorphic cellular and nuclear morphology. The presence of mitotic figures indicated that cells in this tumor were still actively dividing. Sections of the control tumor exhibited an average of 5 ± 1 mitotic figures/500 cells. Conversely, the cells in the vitamin-treated section (Figure 3B) averaged 1.23 ± 0.84 mitotic figures/500 cells. Although the incidence of mitosis was small, the difference between control and vitamin-treated cells was statistically significant (p < 0.01). The nuclei of some cells appeared pyknotic, whereas other cells contained nuclei with marginated chromatin and predominant condensed nucleoli. This section was also characterized by the presence of many vacuolated cells and the presence of necroses.
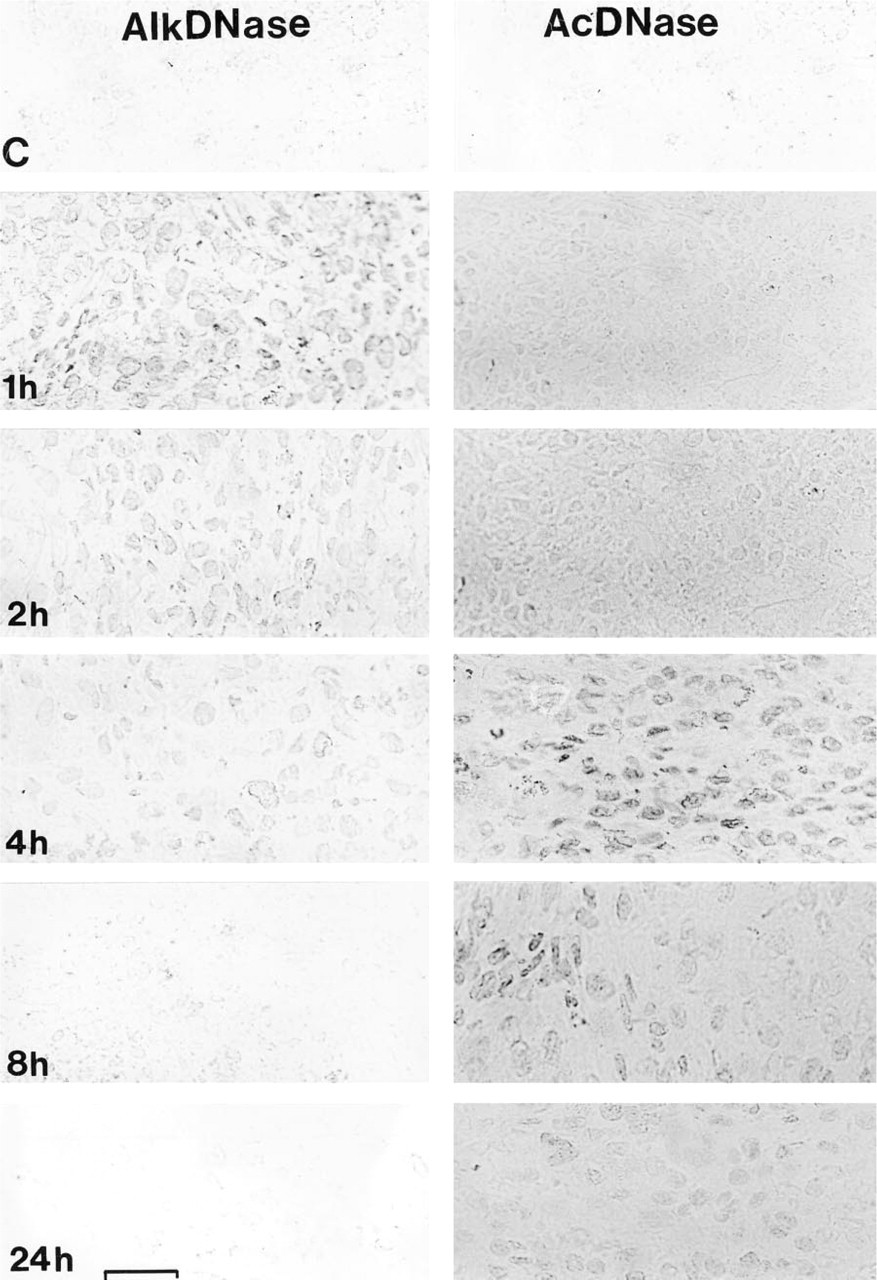
Representative montage illustrating reactivation of DNases in an implanted human prostate tumor after single-dose administration of IP and oral vitamins C and K3. (
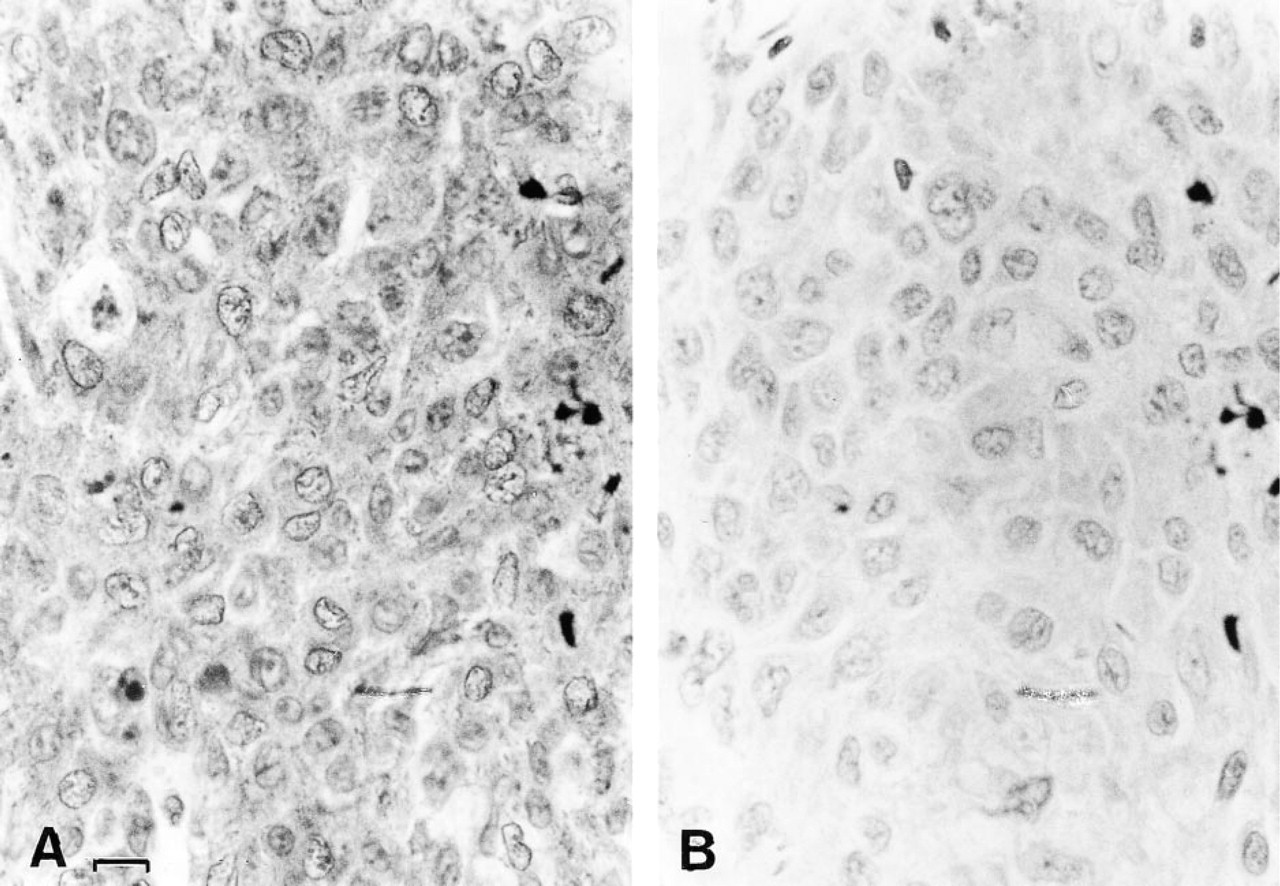
Methyl green-pyronin Y staining of sham-treated and vitamin-treated implanted tumors. The sham-treated tumor cells (
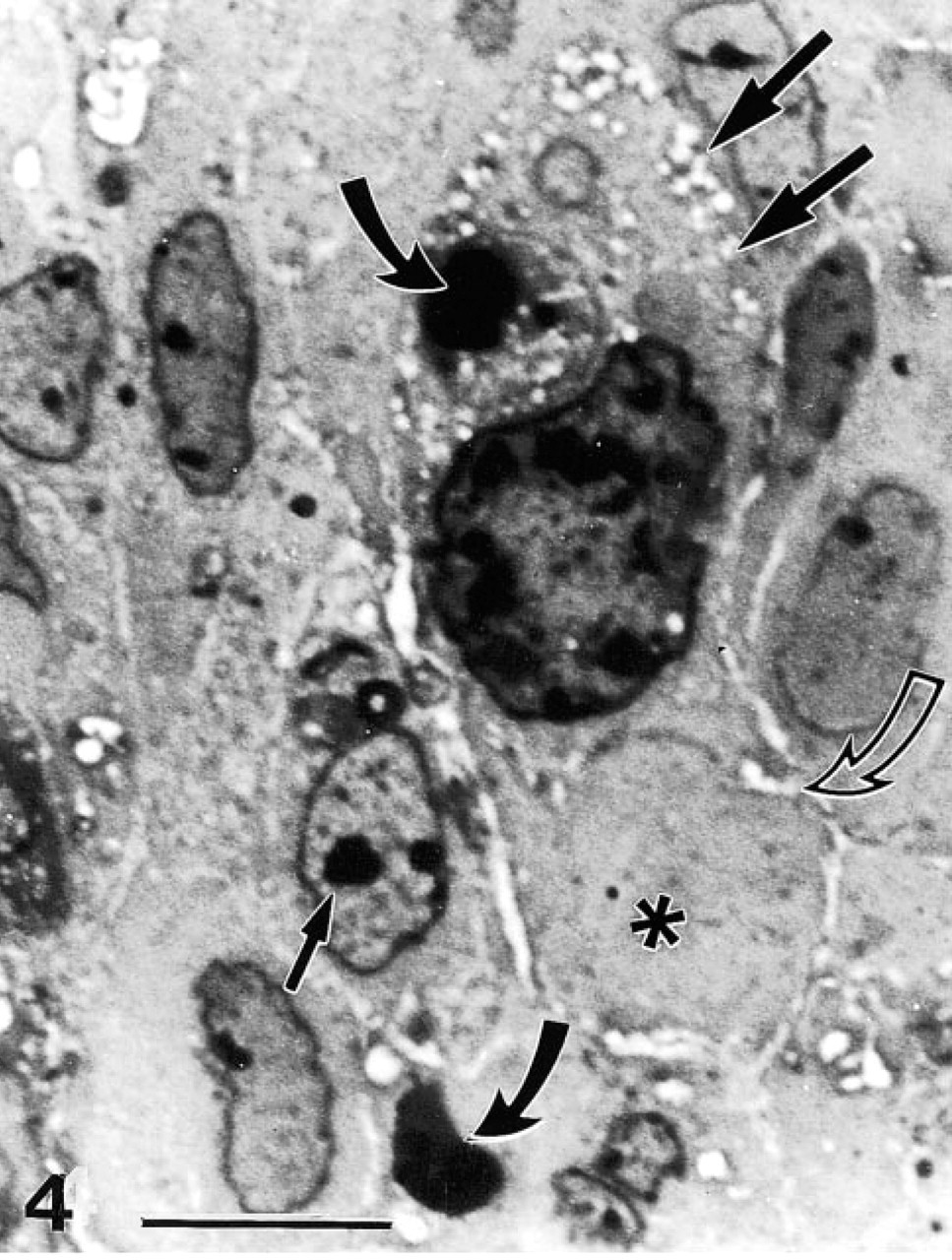
Figures 4–6 is a gallery of light micrographs of Toluidine blue-stained sections of vitamin-treated tumors 4 hr after treatment, depicting tumor cells in various stages of cell injury and cell death. These micrographs are characteristic of this tumor and reveal a plain, compact mass with few capillaries and little to no mitotic activity. The poor vascularization of these tumors (v = 250 mm3) is probably not a function of vitamin treatment because angiogenesis in tumors derived from DU145 cells is greatly diminished compared to that of other prostate cancer cell lines (Connolly and Rose 1998). However, the cells comprising the tumor exhibited cellular and nuclear pleiomorphism. Nuclear pleomorphism was especially evident in the broad range of nuclear size. In Figure 4, the peripheral cytoplasm of the large tumor cell, with the enormous nucleus, displayed a great deal of damage in the form of fine plasmalemmal vacuolizations and superficial exoplasmic vacuoles. The membrane defects associated with these vacuolizations suggest that this large cell was in the process of excising a large autoschizic body which is delineated by a peripheral line of vacuoles (curved open arrow). In addition, this large cell contained a large phagosome (curved dark arrow) digesting an apoptotic (nuclear) body that was the remnant of an adjacent apoptosis. The neighboring cells also possessed a small number of vacuoles and phagosomes. Many of these cells showed nuclei with condensed eccentric to centrally located nucleoli and marginated chromatin.
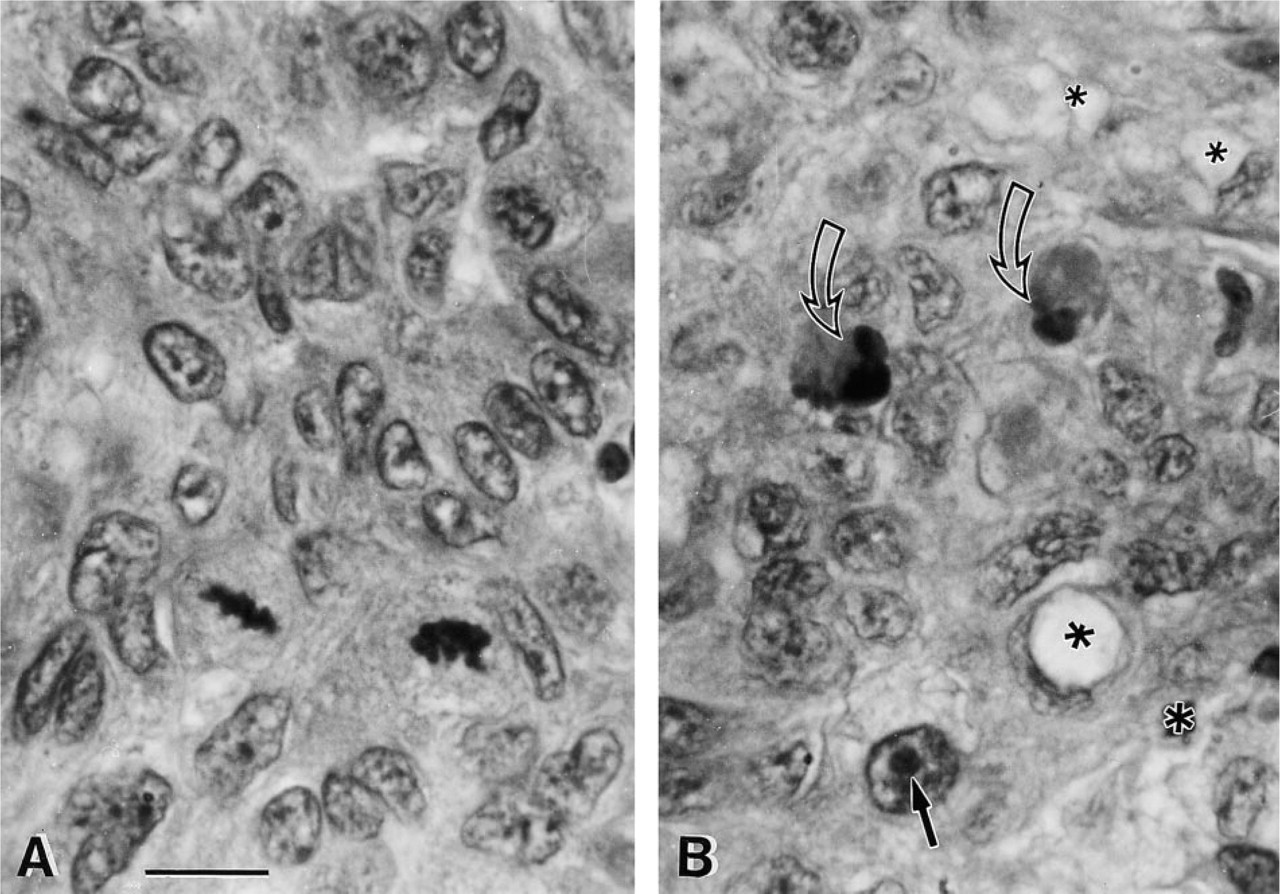
Hematoxylin- and eosin-stained implanted tumor. (
Figures 5 and 6 display similar but more exacerbated cell changes that result in multiple types of cell death, including necrosis, apoptosis, and autoschizis. Whereas necrosis, apoptosis, and autoschizis were not observed in control tumor cells, an average of 9.0 ± 2.1 autoschizic, 3.0 ± 1.1 necrotic, and 1.97 ± 0.45 apoptotic cells were observed per 500 cells after vitamin treatment. These results are consistent with the results obtained with annexin V-propidium iodide-labeled cells (Ervin et al. 1999). In the lower right-hand corner of Figure 5, the pale staining cells with the round to ovoid euchromatic nuclei are intact tumor cells that have not yet been affected by vitamin treatment. The cells shown in the inset are from a field of view adjacent to that shown in Figure 5. These cells are undergoing apoptosis and show crescentic condensation of chromatin, cell rounding and greatly reduced amounts of cytoplasm that are characteristic of this type of cell death. The majority of the remaining cells that display hyperchromaticity and intense vacuolization are in the process of cell necrosis through autoschizis.
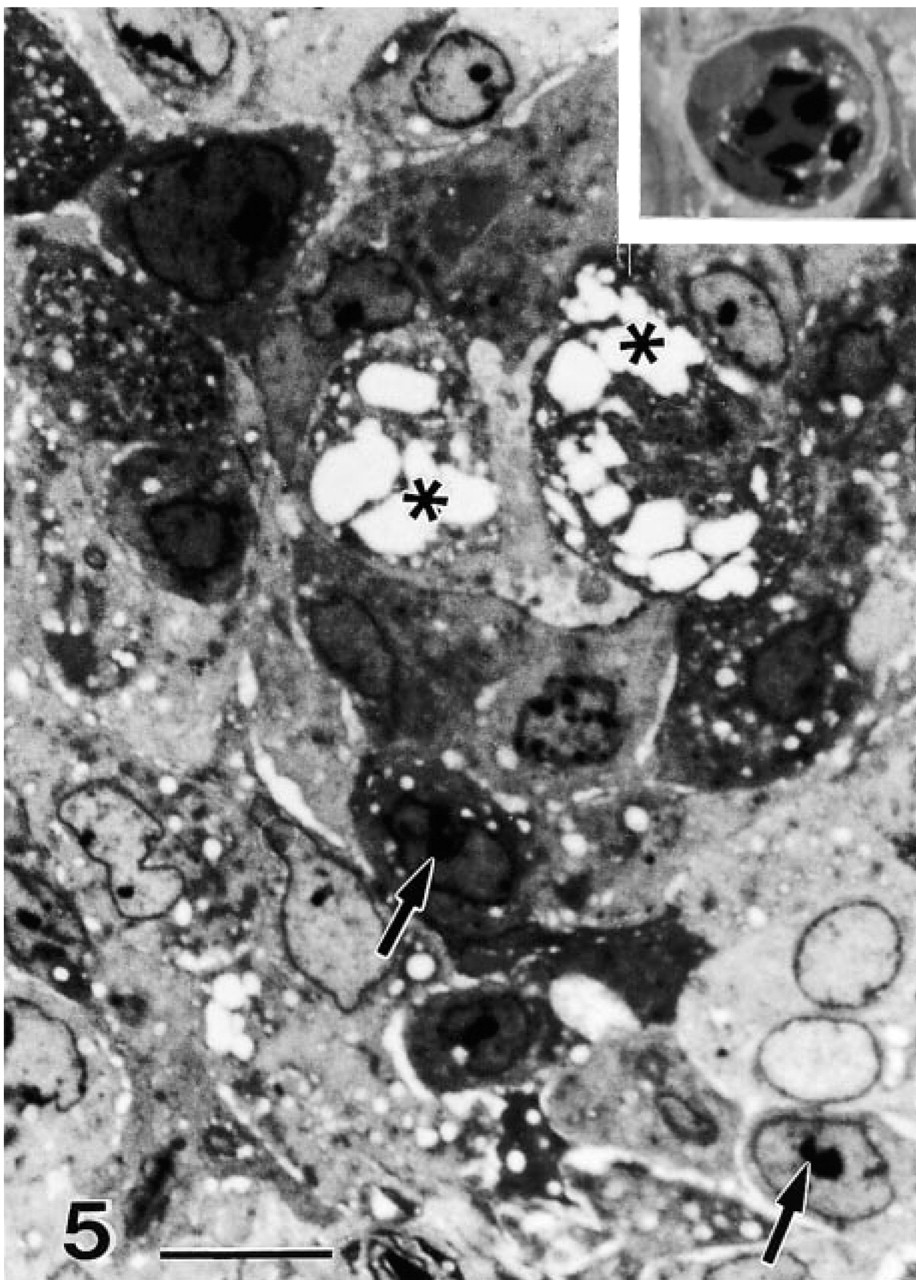
Small arrows indicate examples of nucleolar condensation and overall condensation of nucleoplasm and cytoplasm. The pale-stained cells in the lower right are intact DU145 cells. Stars indicate vacuolizations that lead to cytoplasmic excisions, while the nucleus remains intact. (
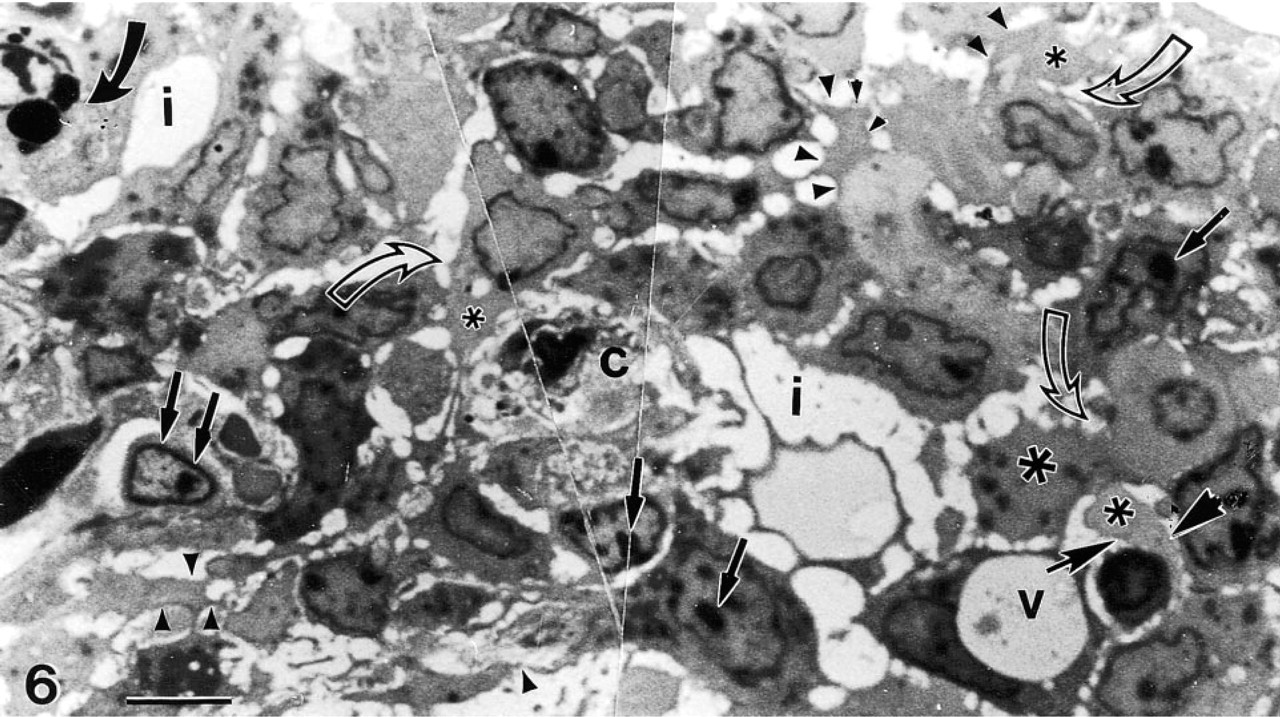
A photomontage of a peripheral area of the tumor depicting a capillary (c) surrounded by several vacuolated (v) tumor cells, undergoing nuclear changes preceding necrosis or autoschizis [condensation and segregation of nucleoli (small straight arrows)]. Some of these cells contain large cytoplasmic phagosomes (curved dark arrow), while the majority of the cells are undergoing autoschizis and exhibit cytoplasmic excisions (stars surrounded by small arrowheads), which result in exaggerated intercellular spaces (i).
The process of autoschizis is more evident in Figure 6, in which the vacuolizations are marked by stars and the cytoplasmic excisions are labeled with arrowheads, with intercellular vacuolizations and exaggerated intercellular spaces (i). These autoschizic tumor cells were found lining the necrotic foci of the tumor, which often form around capillaries such as the one in the center of Figure 6. These cells are characterized by exaggerated membrane damage and by a process of progressive loss of cytoplasm in which the perikaryal compartment separates from the main cytoplasmic cell body in a series of self-excisions. These self-excisions (dark arrowheads with short shafts) typically continue until the perikaryon consists of an apparently intact nucleus and a thin rim of cytoplasm containing damaged organelles. The excised cytoplasmic blebs are primarily filled with dense glycogen patches but contain no organelles or nuclear fragments. These cells are characterized by exaggerated membrane damage and by progressive loss of cytoplasm in which the perikaryal compartment separates from the main cytoplasmic cell body in a series of self-excisions. During autoschizis, nucleoplasm becomes more chromatic and nuclear size decreases. Therefore, the size of the resultant autoschizic cell or body is much smaller than the tumor cell from which it originated.
Discussion
Because endonuclease activation is one of the earliest changes denoting irreversible commitment to cell death, it is generally believed to be involved in the triggering of cell death rather than to be the result of it. Several candidate deoxyribonuclease molecules have been identified in various cell lines and tissues, including the caspase-activated DNA fragmentation factor caspase-3-activated DNase (DFF40/CAD) nuclease, DNase I, and DNase II. DFF endonuclease is primarily responsible for mediating DNA laddering during apoptosis after its activation by caspases (mainly caspase 3). DFF is composed of two subunits, a 40-kD caspase-3-activated nuclease and its 45-kD inhibitor. This protein complex resides in the nucleus and is activated by caspase-3 cleavage of the inhibitor and its subsequent dissociation from the endonuclease. The activated endonuclease exhibits a pH optimum of 7.5, requires Mg2+ (not Ca2+), and is inhibited by Zn2+ (Inohara et al. 1999; Widlak et al. 2000). Although the pH optimum, nuclear localization, and other factors suggest that the DFF40/CAD nuclease may be the reactivated alkaline DNase, a number of other factors argue that another DNase is primarily responsible for the DNase cleavage observed during vitamin-induced tumor cell death. First, autoschizis rather than apoptosis is the predominant form of vitamin-induced tumor cell death, and the DFF40/CAD nuclease is associated with apoptotic cell death. Second, preliminary studies conducted in our laboratories indicate that caspase-3 activation by the vitamin combination is marginal and only occurs 3 to 4 hr after vitamin treatment (data not shown). Third, electrophoretic gels of DNA isolated from cell lines after vitamin treatment reveal a smearing pattern with a subdued laddering pattern embedded in it, which occurs as early as 1 hr after vitamin treatment (Ervin et al. 1999). These results reveal a DNA degradation pattern that is inconsistent with DFF40/CAD nuclease activation and which precedes caspase-3 activation. Although these observations do not eliminate DFF40/CAD participation in later stages of the process, they do suggest that other DNases are involved.
Two of these DNases (DNase I and DNase II) are of particular interest because of their broad tissue distribution and because they have been implicated as possible effectors of apoptosis (Peitsch et al. 1994; Krieser and Eastman 1998). DNase I has an alkaline pH optimum around 7.5, with enzymatic activity occurring in a pH range of 5.5–9.0. Although DNase I requires micromolar concentrations of both Ca2+ and Mg2+ for optimal enzymatic activity, the enzyme may be activated by 1 mM Mg2+ alone. Subcellular fractionation studies have shown that DNase I is localized primarily in the mitochondria, with activity also being detected in the nucleus (Beaufay et al. 1959). A more recent immunolocalization study indicates that substantial amounts of DNase I are also found in the endoplasmic reticulum/nuclear envelope (Peitsch et al. 1993).
In contrast to DNase I, DNase II (acid DNase) does not require divalent cations for its activity (Allfrey and Mirsky 1952; Peitsch et al. 1994). It has an acidic pH optimum at about 5.0, with enzymatic activity occurring in a pH range of 3.0–7.0. Subcellular fractionation studies demonstrate that although the specific activity of DNase II is highest in the lysosome, over 50% of the total cellular DNase II is located in the nuclear fraction. The presence of nuclear DNase II has also been confirmed in fixed tissue sections by histochemical detection with the lead nitrate method (Vorbrodt 1961). Nuclear DNase II is very stable and is only slightly depleted after 24 hr, even in the presence of cycloheximide (Barry and Eastman 1993).
The activity of alkaline and acid DNases is inhibited in non-necrotic cells of malignant tumors in humans and experimental animals (Taper 1967; Taper et al. 1971a, b; Fort et al. 1974; Taper and Bannasch 1976). Reversal of this inhibition of DNase activity in malignant tumors has been linked to necrotic events and to efficient tumor therapy (Williams et al. 1974; Taper et al. 1981). In a study employing transplantable murine hepatoma cells, DNases have been shown to be reactivated in necrotic tumor cells after in vivo irradiation or in vitro treatment with a variety of different compounds.
The compounds that reactivate alkaline and acid DNase activities demonstrate a distinct enzymatic specificity (Festy and Paoletti 1963; Taper et al. 1981). Sulfhydryl agents, including vitamin K3, exclusively reactivate alkaline DNase. Reducing agents, including vitamin C, reactivate acid DNase without any effect on alkaline DNase. The reactivation of alkaline DNase activity occurs rapidly (within 1 hr) and is of shorter duration than the reactivation of acid DNase. The reversal of acid DNase deficiency is delayed compared to that of alkaline DNase. However, once acid DNase activity is restored, its half-life is much longer than that of alkaline DNase (Taper et al. 1981).
Although the relatively rapid reversiblity of the deficiency of alkaline DNase argues against a real decrease in enzyme content and the subsequent de novo synthesis of the DNase, the later time course of acid DNase activity could involve activation of both preexisting and de novo acid DNase synthesis (Taper et al. 1981). At least in the case of the alkaline DNase, these observations point to a masking of enzymatic activity, either by structural latency (i.e., the release of sequestered DNases from other intracellular compartments) or by the presence of enzyme inhibitors (Sierakowska and Shugar 1977). Furthermore, the lack of reactivation of DNases in tumor cells after treatment with phospholipases, as well as the inhibition of usually active DNases in normal rat liver by tumor homogenate, suggests that DNase deficiency in malignant cells is due to the presence of inhibitors and is not linked to structural latency (Taper et al. 1981).
Protein nuclease inhibitors that bind to DNase molecules in a 1:1 ratio have been found in normal tissues (Lindbergh and Skoog 1970; Lesca 1976) and may even be increased in tumor cells (Loiselle and Carrier 1963). In the case of DNase I, monomeric actin has been implicated as the inhibitory molecule (Lacks 1981; Peitsch et al. 1994). A monomeric natural protein inhibitor of DNase II has been purified to homogeneity from bovine liver (Lesca 1976). Small shifts in pH have been shown to affect the interaction between the inhibitor and DNase II and lead to recovery of nuclease activity (Lesca and Paoletti 1969), whereas DNase I is reactivated by sulfhydryl reagents (Festy and Paoletti 1963). However, the effects (if any) of vitamin C and vitamin K3 on the DNase inhibitor complexes has not been eluicidated.
The results of the current study demonstrate that sham-treated tumor cells are essentially devoid of DNase activity. After administration of a vitamin C/Vitamin K3 combination, both alkaline DNase (DNase I) and acid DNase (DNase II) activity was detected in cryosections of the frozen tumor specimens by a modified Gomori's lead nitrate method. Alkaline DNase activity appears as early as 1 hr after vitamin administration, decreases until 2 hr, and disappears by 8 hr after vitamin administration. Acid DNase activity appears 2 hr after vitamin administration, reaches its highest level 4–8 hr after treatment, and maintains its activity 24 hr after treatment. Methyl green staining indicates that DNase activity is accompanied by a decrease in DNA content of the tumor cells. Examination of 1-μm sections of the vitamin-treated tumors indicates that vitamin-induced DNase reactivation and the subsequent degradation of DNA are sufficient to induce tumor cell death. Although both necrotic and apoptotic cell death are observed in the tumor, the primary form of vitamin-induced tumor cell death is autoschizis, a novel type of necrosis characterized by exaggerated membrane damage and the progressive loss of cytoplasm through a series of self-excisions. These self-excisions typically continue until the perikaryon consists of an apparently intact nucleus surrounded by a thin rim of cytoplasm containing damaged organelles (Gilloteaux et al. 1998, 1999; Ervin et al. 1999; Gilloteaux et al. in press).
In vivo studies designed to determine the effect of vitamin administration on the lifespan of nude mice demonstrated that mice receiving both oral and IP vitamin lived significantly longer (p < 0.01) than control mice. The results of additional in vivo studies, designed to determine the effect of vitamin administration on the growth of solid tumors in nude mice, demonstrated that administration of clinically attainable doses of oral vitamins ad libitum in drinking water could significantly reduce the growth rate of solid tumors in nude mice (p < 0.05). Finally, nude mice receiving the vitamin combination did not exhibit any significant bone marrow toxicity, changes in organ weight, or pathological changes in these organs (Jamison et al. in press). Additional experiments are in progress using transgenic mice that more closely mimic the development and metastases of prostate cancer in humans. Among the different mechanisms that may be involved in this therapeutic effect of combined vitamins against cancer, the most plausible is a stimulation of a redox cycling system that produces H2O2 and other active oxygen species that are involved in cell membrane lipid peroxidation, DNase activation, and DNA breaks leading to cell death (Gilloteaux et al. 1995; Jamison et al. 1996, 1997; Venugopal et al. 1996a, b). Moreover, the most important fact is that this action of combined vitamins C and K3 appeared to be selective for cancer cells (Taper and Roberfroid 1992). As opposed to normal organs and tissues, cancer cells are usually deficient in catalase, superoxide dismutase, and/or glutathione peroxidase, which constitute the cellular armamentarium system/defense system against free radicals (Sinha and Mimnaugh 1990). This hypothesis is supported by the fact that simultaneous administration of vitamin C and K3 with catalase suppressed the potentiating and sensitizing effect of these vitamins (Noto et al. 1989). Because the vitamin combination is a chemosensitizer (Taper et al. 1987) and a radiosensitizer (Taper et al. 1996), combined vitamin C and K3 administration can be considered as a new, non-toxic adjuvant cancer therapy, which can be easily introduced into the classical protocols of clinical cancer therapy without any supplementary risk for patients.
Footnotes
Acknowledgments
Supported by grants from the American Institute for Cancer Research, 97B048, the Summa Health System Foundation of Akron, Ohio, and the Hess-Roth Kaminski & Maxon Foundation of Erie, Pennsylvania.