
Abstract
We studied carbohydrate residues of glycoproteins and proteoglycans (PGs) in peritoneal Pacinian corpuscles of five adult cats. Terminal monosaccharides of glycoproteins and related polysaccharides were identified by lectin histochemistry and the PGs and glycosaminoglycans (GAGs) by specific antibodies. The most intensive lectin staining reactions indicated an abundance of glycoconjugates with terminal mannose (Man) or sialic acid residues, but no complex-type oligosaccharides were detected within the corpuscles. Terminal fucose (Fuc) and galactose (Gal) residues typical for O-linked mucin-type glycoproteins generally associated with high water binding capacity were also absent. Antibodies against unsulfated chondroitin (C-0-S), chondroitin-4-sulfate (C-4-S), and decorin showed positive reactions in the interfibrillar spaces between the lamellae, around collagen fibers, and around the lamellae of the perineural capsule, especially in the outer parts known to contain Type II collagen. Biglycan showed a preference for the innermost part of the perineural capsule (intermediate layer), known to contain Type V collagen. Collagen V and biglycan are both linked to growth processes. Hyaluronic acid (HA), chondroitin-6-sulfate (C-6-S) chains, and a chondroitin sulfate proteoglycan (CSPG) were colocalized in the terminal glia. The study of carbohydrates with high water binding capacity may contribute to our understanding of the high viscoelasticity of Pacinian corpuscles.
P
The single afferent axon is myelinated. However, its myelin sheath is lost before the axon enters the inner core. The nuclei of terminal glial cells are arranged in a beaded fashion, such that the lamellae of two glial cells form one half of the sheath of the axon. The terminal glial cells are covered by the basal lamina, around which collagen fibers form a network between the perineural lamellae extending into the central cleft. The perineural capsule is composed of flattened perineural cells forming, together with their basement membranes, up to 70 lamellae separated by 10-μm-wide interspaces. The inner core is surrounded by three parts of the perineural capsule: the intermediate layer (growth zone), the outer core, and the external capsule (Halata 1975, 1993; Ide et al. 1988; Munger et al. 1988; Zelena 1994; Pawson et al. 2000) (Figure 1).
Laminin and Type IV collagen are components of all structures of the corpuscle, with the exception of the axon. The S-100 protein is exclusively localized in the capsular cells and the inner core. Pacinian corpuscles have some immunohistochemical characteristics in common with Schwann cells and perineural fibroblasts of other neural structures (Vega et al. 1994, 1995b, 1996). The proteoglycan (PG) β-amyloid precursor protein has been found in the inner core, outer core, and capsule of Pacinian corpuscles (Vega et al. 1995a).
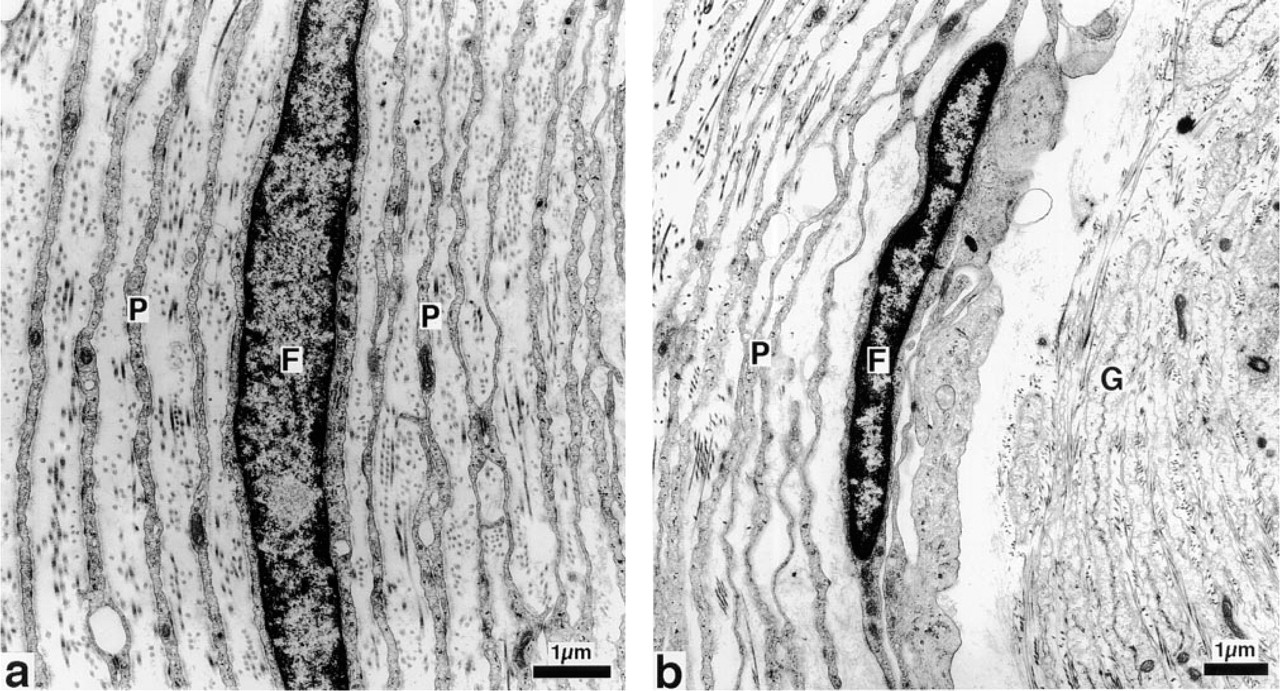
Mesenterial Pacinian corpuscle of the cat. Electron microscopic photograph of the perineural capsule (
The extracellular matrix is one morphological hallmark of these receptors and hence, as is also the case in other tissues, their mechanical features must be largely determined by the substances present in the interfibrillar matrix. Like most other secreted proteins, proteins of the extracellular matrix are glycosylated. In addition to glycoproteins, glycosaminoglycans (GAGs) form the major constituents.
To date, no analysis of the carbohydrate components of the extracellular matrix of Pacinian corpuscles exists. The aim of this investigation therefore was to analyze the composition of mainly terminal carbohydrate residues of glycoconjugates, using lectin histochemistry, and to characterize the proteoglycans by the use of specific monoclonal antibodies and hyaluronectin.
Materials and Methods
Subperitoneal Pacinian corpuscles were dissected out from the mesentery of five adult cats, fixed in 4% neutral buffered formalin (for 48 hr at 4C), and embedded in paraffin. Five-μm-thick sections were deparaffinized, rehydrated, and used for routine hematoxylin and eosin staining and for the following histochemical procedures.
Lectin Histochemistry
Rehydrated sections were pretreated for 15 min with 0.1% trypsin in distilled water, washed in Tris-buffered saline (TBS: 50 mM Tris, 150 mM NaCl, 1 mM MgCl2, 1 mM CACl2, pH 7.4) and incubated with 10 μg/ml biotinylated lectins (for source of the lectins, their abbreviations, and their nominal carbohydrate specificities, see Table 1). SBA, WGA, UEA-I, LCA, and PHA-L were supplied by Sigma (Deisenhofen, Germany). The other lectins were in-house preparations (EJM van Damme, WJ Peumans). The incubation was performed for 1 hr at room temperature (RT). After careful washing in TBS, the sections were incubated with biotin–avidin complexes with alkaline phosphatase as a marker enzyme (Vector ABC Kit; Burlingame, CA; dilution 1:250 in TBS) for 30 min. After a careful wash in TBS, naphthol-AS-biphosphate was used as the phosphatase substrate and hexazotized New Fuchsin was used for simultaneous coupling. The reaction was finally stopped after 30 min with 4% buffered formalin. For nuclear counterstaining, Mayer's hematoxylin 1:1 diluted in distilled water was used and the slides were mounted in a water-soluble mounting medium.
Because our material was limited, the following reactions were performed as control reactions in only a part of the samples: succinyl WGA (Vector), UDA (Urticaria dioica, inhouse supply), CMA (Chelidonium majus, in-house supply), and the reaction with WGA after neuraminidase digestion. The results of these reactions are therefore not contained in the tables.
Lectins
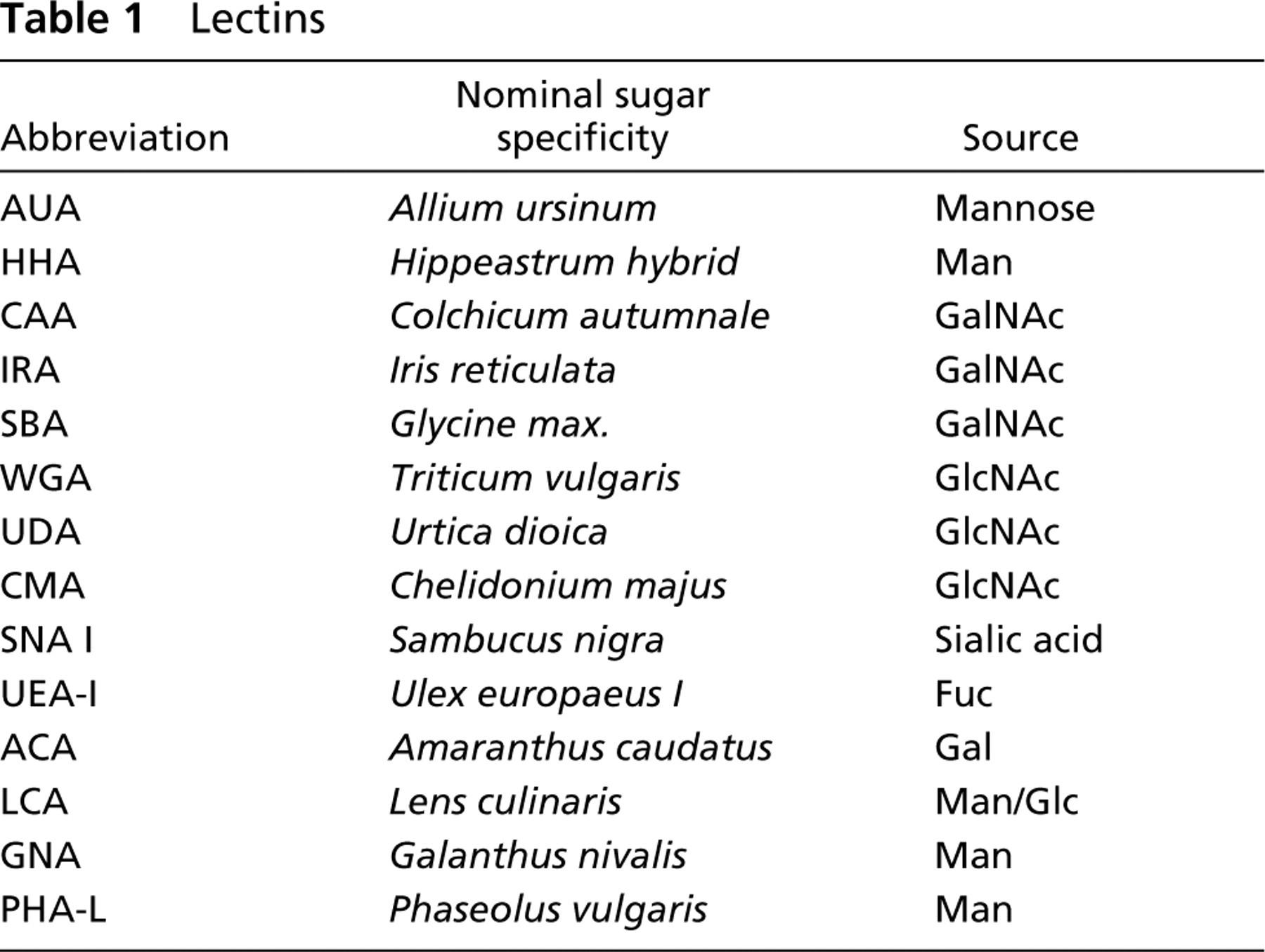
The specificity of the lectin binding was controlled by incubation of the lectins with their specific inhibitory monosaccharides (100 mM final concentration). For the neuraminic acid-reactive lectins, neuraminidase predigestion of the sections was used.
Proteoglycan Immunohistochemistry
For the immunostaining using side chain-specific chondroitin (Ch) antibodies and protein-specific antibodies against a chondroitin sulfate proteoglycan (CSPG), paraffin sections were rehydrated. Antibodies against unsulfated chondroitin (C-0-S), chondroitin-4-sulfate (C-4-S), and chondroitin-6-sulfate (C-6-S) were obtained from Seikagaku (Tokyo, Japan) and the CSPG antibody from Sigma. In the case of the antibodies against C-0-S, C-4-S, and C-6-S, the reaction of the first antibody was preceded by chondroitinase ABC (Sigma) digestion to unmask the specific antigen epitope. Sections were rehydrated and incubated with 0.5 U of chondroitinase ABC in 1 ml 0.25 M Tris buffer (pH 8.0) for 4 hr at 40C in a moist chamber. Slides with (C-0-S, C-4-S, and C-6-S) or without predigestion (CSPG, decorin, biglycan) were carefully washed with PBS and incubated with the first antibodies (dilution 1:200) overnight at 4C in a moist chamber. This was followed by a careful wash and an incubation with a secondary biotinylated antibody (rabbit IgG) and, after a further wash, a rabbit–PAP complex was applied. After a further wash, an incubation with an ABC complex followed (Davidoff and Schulze 1990). The peroxidase reaction was visualized by incubation with 0.05% diaminobenzidine (DAB) and 4 μg/ml glucose oxidase. Finally, the slides were dehydrated and mounted in Crystal Mount (Biomeda; Foster City, CA).
For staining of biglycan, slides were pretreated with 1.5% H2O2 to inhibit endogenous peroxidase, followed by incubation with a rabbit polyclonal antiserum against a synthetic peptide from the C-terminus of human biglycan core protein (generously provided by Dr. P.J. Roughley, Shriners Hospitals for Crippled Children, affiliated with McGill Univ., Montreal, Canada) as the first antibody (diluted 1:300). This was followed by a secondary biotinylated anti-rabbit antibody. The further procedure was the same as with the other antibodies.
Decorin was detected with an anti-human decorin antibody (Biomol, Hamburg, Germany; dilution 1:300), and the same procedure as with biglycan was applied.
Omission of the first antibodies was used as negative control. In the case of chondroitin-proteoglycans, omission of chondroitinase digestion as well as omission of the first antibody after chondroitinase digestion served as further controls. Rat articular cartilage, mast cells, and human arteries contain these GAGs and were used as positive controls.
Hyaluronectin Histochemistry
For detection of hyaluronic acid (HA), the slides were treated with 0.3% H2O2 in absolute methanol for 20 min to block endogeneous peroxidase and washed in 0.05 M Trisbuffered saline (TBS), pH 7.6. The slides were incubated with biotinylated sheep brain hyaluronectin (isolated by B. Delpech; diluted 1:20 in TBS) for 1 hr, followed by a wash in TBS. After incubating the sections with an avidin–peroxidase complex for 20 min (streptavidin ABC complex; DAKO, Hamburg, Germany), a careful wash in TBS followed. Finally, an incubation in 0.05% DAB in TBS containing 0.6% H2O2 for 10 min was conducted to visualize the peroxidase. The sections were counterstained with hematoxilin, dehydrated, and mounted in Eukitt (Dr. Kindler GmbH; Freiburg, Germany).
To test the specificity of the hyaluronectin binding, sections pretreated with testis HAse served as controls. Twenty units HAse from sheep testes (Type III, EC 3.2.1.35; Sigma) was dissolved per ml in 0.02 M NaH2P04 with 0.077 M NaCl and 0.01% bovine serum albumin, pH 5.35. A parallel section was incubated with the buffer lacking enzyme and slides were incubated for 4 hr at 37C. Incubation was repeated after addition of the same fresh enzyme solution for 20 hr, followed by a wash with distilled water.
Type IV Collagen
Type IV collagen was demonstrated by a polyclonal antibody reacting with human and other mammalian Type IV collagen (Quartett; Berlin, Germany). Incubation with the first antibody (dilution 1:40) was followed, after careful washes in buffer, by a biotinylated secondary antibody, followed by a wash and a biotin–streptavidin–alkaline phosphatase complex. Visualization of the phosphatase with New Fuchsin was carried out according to standard procedures. Nuclei were counterstained with Mayer's hematoxylin. After dehydration, slides were mounted in Crystal Mount.
Semiquantitative evaluation was performed by blind estimation of reaction intensities of structures in the corpuscles.
The terms used for different parts of the perineural capsule in this paper are compatible with the terms used by Pawson et al. (2000) as follows (terms of Pawson et al. in parentheses): innermost lamellae (intermediate layer or growth zone); inner lamellae (outer core); outer lamellae (external capsule); outer limiting lamella. We prefer general terms because these parts are not strictly distinguishable from each other.
Semiquantitative evaluation of staining intensities of lectins in different structures of Pacinian corpuscles a

aOutlL, outer limiting perineural lamella; OutL, outer perineural lamellae (external capsule); InnL, inner perineural lamellae (outer core); InnML, innermost perineural lamellae (intermediate layer or growth zone); Isp, interfibrillar spaces between perineural lamellae; Fib, interlamellar collagenous fibers; Term, terminal glia; CT, surrounding connective tissue; Cont, positive staining in negative controls. Reaction intensities (means of all specimens): + + + +, intense reaction; + + +, medium reaction; ++, positive reaction; +, weak reaction; -, negative reaction.
Results
Lectin Histochemistry
Results concerning the lectin binding pattern are summarized in Table 2. The mannose (Man)-specific AUA labeled all neuronal and neuroglial elements of Pacinian corpuscles intensively (Figure 2a), whereas the Man-specific HHA, which has a similar nominal specificity for mannose, preferentially labeled the outer limiting perineural lamella. In contrast to the neural and neuroglial elements, the surrounding normal connective tissue showed a high reaction intensity with both lectins.
Of the N-acetyl-galactosamine (GalNAc)-specific lectins, CAA stained all perineural lamellae (perineural cells and basement membranes), collagen fibers, and the interlamellar fluid intensely (Figure 2b). With IRA, the outer limiting lamella of the perineural capsule was intensely labeled. Staining of medium to high intensity was visible within the innermost lamellae and collagen fibers between lamellae (Figure 2c). SBA showed the same staining pattern. Common to all GalNac-specific lectins was the distinct reactivity of the connective tissue surrounding the corpuscles.
WGA, which is able to react with N-acetyl-glucosamine (GlcNAc) and sialic acid, labeled all structures of the Pacinian corpuscles, including the surrounding connective tissue, very intensely (Figure 2d). In the corpuscle, the innermost lamellae were stained most intensely. A staining pattern similar to that of WGA could be observed with the sialic acid-specific lectin SNA-I (Figure 2e). Controls in sample tests showed that there was no staining with succinyl WGA and that after neuraminidase digestion a distinct reduction of the staining was found, especially in the outer lamellae and outer limiting lamella. With UDA and CMA a faint reaction of the innermost lamellae was found in most of the Pacinian corpuscles, whereas single corpuscles remained completely unstained.
The fucose (Fuc)-specific lectin UEA-I and the galactose (Gal)-specific ACA (Figure 2f) labeled predominantly the surrounding connective tissue fibers.
There are two Man-specific lectins with monosaccharide specificities different from the types mentioned above. LCA reacts with mannose/glucose. Its reaction with the structures of the corpuscle were similar to those of HHA (Figure 2g). In contrast, GNA showed no reaction at all (Figure 2h).
PHA-L, specific for complex-type glycoproteins, did not label any structures of the Pacinian corpuscles, except for the endothelium of blood vessels in and around the corpuscles.
Control preincubations with monosaccharides or neuraminidase predigestion of the sections resulted in lack of reactivity with respect to the Pacinian corpuscles. An exception was AUA, where the reaction was not totally extinct within the corpuscle, and CAA, where it was only slightly reduced after the monosaccharide preincubation (Table 2).
Glycosaminoglycan and Proteoglycan Histochemistry
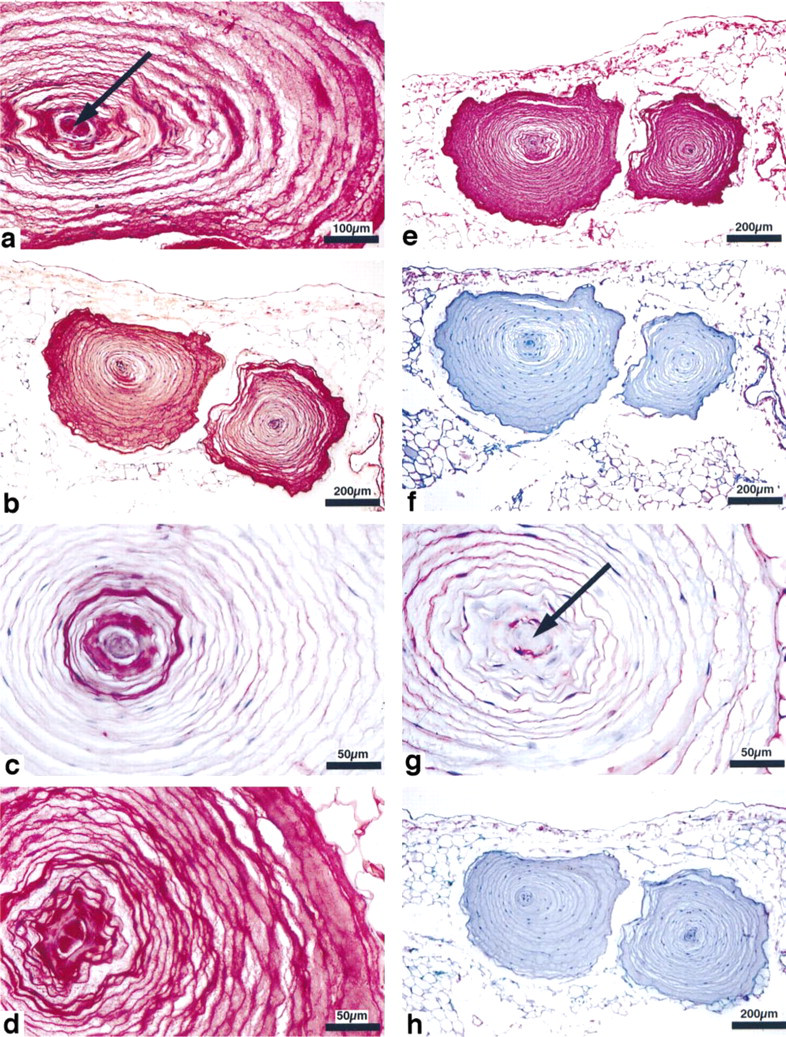
Lectin histochemistry of Pacinian corpuscles. (

Proteoglycan histochemistry of Pacinian corpuscles. (
The C-4-S antibody stained the lamellae of the perineural capsule. The most intense labeling was found in the outer limiting perineural lamella (Table 3, Figure 3c). In contrast, the interfibrillar space between the lamellae showed low staining intensity.
With the C-6-S antibody, the only structures showing highly intense staining were the terminal glial cells and the outer limiting lamella of the perineural capsule. The perineural lamellae showed some staining. The interfibrillar space between the perineural lamellae remained unstained (Table 3; Figure 3e).
The CSPG antibody produced a pattern very similar to that of C-6-S, whereby the staining intensity in most of the structures was slightly lower.
The decorin reaction was found in most structures of the Pacinian corpuscle and showed its highest labeling densities in the perineural lamellae and in connection with the interlamellar fibers (Figure 3f).
The biglycan antibody reacted most intensely with both the innermost and outer limiting lamellae of the perineural capsule. The interlamellar fibers and interspaces remained almost unstained, as did the axon (Figure 3g).
Controls lacking the first antibody, with or without previous chondroitinase digestion, were negative. Controls without chondroitinase digestion sometimes showed an antibody reaction located especially in perineural cells with the C-0-S and C-4-S antibodies. In the undigested controls of 15–20 evaluated corpuscles, 44% remained unstained with the C-0-S, 65% with the C-4-S, and 96% with the C-6-S antibody (Table 3, Figure 3b,d).
HA was present only in the terminal glia and the outermost perineural lamella, but not in the interfibrillar spaces between the perineural lamellae (Figure 3h). The controls were negative.
The staining pattern of Type IV collagen is already known (see above). It has been used to distinguish cellular from pericellular structures of the lamellae.
Discussion
This study was undertaken to identify the carbohydrate composition of the Pacinian corpuscles, which are morphologically characterized by their large volume of extracellular matrix. Because the extracellular matrix is rich in glycoproteins and glycosaminogly-cans, which are representative of the water binding capacity of tissues, these carbohydrate residues are of functional interest. Staining with lectins for different carbohydrate residues indicates a clear-cut carbohydrate distribution pattern within the corpuscle. Sialic acid (SNA-I), and Man (AUA) residues are present within the whole Pacinian body, whereas GalNAc (SBA, IRA, CAA) is present only in defined single structures. IRA and SBA, in particular, show a preferential localization in the innermost lamellae and thus a co-localization with LCA, Type V collagen, and biglycan (see below). The GlcNAc-specific lectins UDA and CMA produce only faint reactions of the innermost lamellae. The WGA reaction, which may be caused by GlcNAc or by sialic acid, is intense in all structures of the corpuscle, similar to the sialic acid-specific SNA-I. However, in contrast to SNA-I, WGA shows a preferential reaction with the innermost lamellae. Controls with succinyl WGA show negative reactions, whereas neuraminidase reduced staining intensity, especially in the outer lamellae of the perineural capsule. Therefore, WGA staining may be produced mainly by reaction with sialic acid, while in the innermost lamellae traces of GlcNAc can be shown, which are not present in all of the corpuscles. Gal and Fuc are present in the surrounding connective tissue and are absent within the Pacinian corpuscles themselves. The different Man-specific lectins show a considerable heterogeneity in their binding pattern, indicating that Man residues in different linkages are present within the corpuscle. A heterogeneity of Man-specific lectin binding similar to that found in Pacinian corpuscles has been observed in the human leukemic mast cell line HMC-I (Schumacher et al. 1998). LCA, which is specific for both Man and Glc, labels the different structures of the Pacinian corpuscle almost indiscriminately. GNA, which recognizes Man only and reacts with different di- and oligosaccharides, which may be branched, e.g., [(Man α1,6 (α1,3-Man) Man-α1,6-(α2,3-Man)Man], is much more limited in its binding (Hester and Wright 1996; Wright and Hester 1996). The other two Man-specific lectins AUA and HHA are similarly broad with respect to their binding. AUA resembles LCA and HHA resembles GNA. Because AUA (van Damme et al. 1993) binds to Man only and its binding pattern is so similar to LCA, this result might indicate that in Pacinian corpuscles LCA recognizes only Man residues.
Semiquantitative evaluation of staining intensity of proteoglycan and glycosaminoglycan immunoreactions a

aOutlL, outer limiting perineural lamella; OutL, outer perineural lamellae (external capsule); InnL, inner perineural lamellae (outer core); InnML, innermost perineural lamellae (intermediate layer or growth zone); Isp, Interfibrillar spaces between perineural lamellae; Fib, interlamellar collagenous fibers; Term, terminal glia; CT, surrounding connective tissue; Cont, chondroitinase digested controls. Reaction intensities (means of all specimens): ++ ++, intense reaction; ++ +, medium reaction; ++, positive reaction; +, weak reaction; −, negative reaction.
For the rapid transmission of pressure, it is important for the Pacinian corpuscle to possess high viscoelasticity. Accordingly, a high water content of the extracellular spaces is to be expected. In principle, two main types of water binding molecules can be found within the extracellular matrix: mucins and glycosaminoglycans/proteoglycans, including HA. Mucins are O-linked high molecular weight glycoproteins, with the sugar component usually exceeding that of the protein. Fuc, Gal, and GalNAc are the most likely typical terminal carbohydrate residues of O-linked mucins (Campbell 1999) and their absence, as indicated by the lack of UEA-I and ACA binding, indicates that O-linked glycoproteins are unlikely candidates for the water binding capabilities of the extracellular matrix of the Pacinian corpuscles.
In contrast to the absence of carbohydrate residues typical for O-linked glycoproteins in the extracellular spaces, Man residues are abundantly present. Man residues are typical for the core region of N-linked glycoproteins which are added to the protein backbone in the rough endoplasmic reticulum and are further modified in the Golgi apparatus. These types of glycoproteins are either membrane or secretory glycoproteins (Brooks et al. 1987; Schumacher 1995). As proved by the reaction with PHA-L, the N-linked glycoproteins contain no complex-type oligosaccharides and only the Man core structures.
The other group of glycoconjugates studied in these experiments are the PGs and GAGs. Antibodies directed against Ch substances react with the content of interlamellar spaces. HA is restricted to the lamellae of the terminal glia. It is therefore not present in the perineural capsule and cannot account for their water binding capacity. The absence of HA further implies that no chondroitin proteoglycan complexes with HA, like those of versican, can be present in the interlamellar spaces of the perineural capsule. Complexes of this type are normally responsible for the consistency of cartilage (connected with Type II collagen) and arteries (Sames 1994) and would hence have explained the cartilage-like elasticity of the Pacinian corpuscle.
This elastic consistence may, however, be functionally explained by PG monomers with side chains of high water binding capacity. Such PGs may be bound to the collagen fibers, which form a dense network in the interlamellar spaces of the perineural capsule. Recently, interlamellar collagen in the outer core and external capsule has been characterized as a Type II collagen, while the intermediate layer contains Type V collagen (Pawson et al. 2000). Decorin, a small PG with up to two side chains composed of GalNAc and glucuronic or iduronic acid, is known to interact with collagens Type I and Type II (Kresse et al. 1994.) We demonstrated the presence of decorin in the interlamellar spaces around collagen fibers in those parts of the corpuscle known to contain Type II collagen, where the specific decorin antibody stains with high intensity. In contrast, biglycan, with side chains of similar composition compared to those of decorin, is restricted to the perineural lamellae. This is in agreement with the fact that it has been localized in intracellular and pericellular compartments (Bianco et al. 1990; Schaefer et al. 1998). Biglycan has also been localized in connective tissue (Huang et al. 1999). It is known to interact with collagen Type I (Schönherr et al. 1995) and Type V (Hocking et al. 1996). In our study it is visible only in the lamellae of the intermediate zone. This zone has been postulated to be a growth zone known to contain collagen Type V, which is suggested to play a role in growth and development. In immature corpuscles, this zone is most prominent (see Pawson et al. 2000). In this context, it is of interest that biglycan was proposed to support cell proliferation (Kamo et al. 1993; Veness-Meehan et al. 1994; Oritani and Kincade 1996; Gruber et al. 1997; Tomoyasu et al. 1998).
Water binding capacity is especially influenced by the nature of the carbohydrate side chain and the density of side chains on the core protein. The side chain-specific antibodies with the most intensive reactions are the C-4-S and C-0-S antibodies. Part of the reacting GAGs are expected to be constituents of decorin and biglycan. We speculate that this does not exclude the presence of PGs with higher density of side chains, and therefore higher water binding capacity.
Chondroitin sulfates are reduced by chondroitinase to short stubs located at the protein core, resulting in an artificial antigen (Couchman et al. 1984). Controls without chondroitinase digestion should therefore remain unstained. However, such controls show a reaction with the C-0-S antibody and a very faint reaction with the C-4-S antibody, which may be the result of an unspecific antibody reaction. In contrast to usual negative controls, there may exist specific substrates that are able to react with the antibody even without chondroitinase digestion. Therefore, a possible explanation for positive reactions could be that the antigen resembles an early stage of polysaccharide synthesis, because carbohydrate synthesis starts at the protein core (review in Sames 1994). Such early substrates may therefore react with Ch and CS antibodies similar to the artificial antigen produced by chondroitinase. Most of these early substrates may be located in the cell and may be unsulfated, because sulfation of carbohydrates starts late during synthesis. However, only the difference between reactions in the digested sample and undigested sample has been taken as a result.
The antibodies against CSPG and C-6-S show only weak reactions in the interlamellar spaces of the perineural capsule. With respect to the similar reaction of both antibodies, it can be assumed that C6-sulfated CS represents the main pool of side chains of the CSPG.
Despite its mesenchymal origin, the lectin reaction of the perineural capsule of the Pacinian corpuscle differs from that of surrounding subperitoneal connective tissue. Although fibers of the outer limiting perineural lamellae run continously into the fibers of surrounding connective tissue, it reacts in a different way, e.g., it shows no or only weak reactions with ACA, GNA, UEA-I, and HHA, which is in contrast to the intense reactions of the surrounding connective tissue. PGs and GAGs show also a specific pattern in the corpuscle compared to surrounding connective tissue. Therefore, the Pacinian corpuscle has its own particular pattern of glycoconjugates, which differs from that of connective tissue. Inside the corpuscle, defined structures show individual reaction patterns with the same lectin or PG antibody.
The corpuscle therefore represents a differentiated structure in terms of the pattern of extracellular substances. We cannot definitely decide if the perineural capsule, containing Type II collagen, chondroitin-type glycosaminoglycans, decorin, and a special spectrum of terminal monosachcharides (or oligosaccharides, respectively) represents a water binding matrix structure similar to cartilage tissue, with which it shares mechanical qualities. An interlamellar solution, with less stringent binding to structure components and special colloidal characteristics, could be discussed alternatively.