
Abstract
Epithelial cells attach to the basement membrane through adhesive contacts between the basal cells of the epithelium and the proteins of the extracellular matrix (ECM). The hemidesmosome (HD) is a specialized cell-ECM contact, that mediates the attachment of the epithelial cell basal surface to the ECM. In bronchial epithelial cells, the protein components that constitute the HD have not been demonstrated. Using immunohistochemical techniques, we determined that normal human bronchial epithelial (NHBE) cells express the HD cell surface integrin α6β4 and produce laminin 5, the ECM protein associated with HDs. Furthermore, expression of the HD-associated structural proteins, bullous pemphigoid antigens 1 (BPAG 1) and 2 (BPAG 2), was demonstrated in NHBE cells by immunofluorescence microscopy and immunoblot analyses. In addition, we confirmed the presence of laminin 5 in the basement membrane (BM) of bronchial epithelial biopsy specimens and of BP230, BP180, and the α6β4 integrin heterodimer at the site of bronchial epithelial cell-ECM interaction in vivo. Finally, using electron microscopy, we were able to demonstrate intact HDs in a glutaraldehyde-fixed NHBE cell monolayer. These findings suggest that bronchial epithelium forms HDs and that the laminin 5-α6β4 integrin interaction may be important in stabilizing epithelial cell adhesion to the BM in the lung.
Epithelial cell adhesion to the underlying basement membrane (BM) is essential for normal lung growth and development and for preservation of barrier function and host defense (Dunsmore and Rannels 1996; Roche 1998). Epithelial cells contact the basement membrane through adhesive interactions with extracellular matrix (ECM) proteins (Ekblom et al. 1998). These adhesive interactions are mediated in many instances by cell surface integrins (Hynes 1992; Fuchs et al. 1997).
Integrins are heterodimeric transmembrane receptors that are composed of an α-regulatory and a β-signaltransducing subunit (Rabinovitz and Mercurio 1996). Integrins assemble into specialized cell-ECM connections, such as focal contacts and hemidesmosomes (HDs) (Craig and Johnson 1996; Green and Jones 1996). Focal cell contacts anchor the microfilament cytoskeleton to the cell surface in addition to their role in matrix adhesion, whereas HDs are cell-ECM connections that are associated with the keratin intermediate filament cytoskeleton (Borradori and Sonnenberg 1996; Jones et al. 1998). HDs are formed by the association of the α6β4 integrin heterodimer with laminin 5, a heterotrimeric ECM glycoprotein composed of α3-, β3-, and γ2-subunits. Each HD also possesses three structural proteins, including the bullous pemphigoid antigens I (BPAG 1 or BP230) and II (BPAG 2 or BP180) and plectin (IFAP300) (Jones et al. 1998; Schaapveld et al. 1998). Together, these proteins assemble into the classic or Type I HD. These have been described in a number of epithelia, including skin, bladder and cornea (Riddelle et al. 1991; Baker et al. 1996b; Rabinovitz and Mercurio 1996). Type II HDs lack a well-developed cytoplasmic plaque and are found in gut epithelial cells. These are believed to be complexes formed by the co-localization of laminin 5, α6β4, and plectin (Uematsu et al. 1994). Type II HDs do not possess BP180 and BP230 and therefore may be less stable than Type I HDs.
A number of integrins that facilitate adhesion and attachment have been described in normal human bronchial epithelial (NHBE) cells, but the integrins and other structural proteins that are associated with the formation of bronchial cell HDs have not been identified (Pilewski and Albelda 1993). The data we present here demonstrate that NHBE cells express the integrin heterodimer α6β4 and that the bronchial epithelium produces laminin 5, the ECM protein that co-localizes with the integrin heterodimer to form HDs (Jones et al. 1994). In addition, we have demonstrated that NHBE cells express the bullous pemphigoid antigens BP180 and BP230, and that they associate with ECM-integrin complexes to form classic or Type I HDs, which we illustrate using electron microscopy.
Materials and Methods
Cell Culture and Tissue Preparation
Normal human bronchial epithelial (NHBE) cells were obtained from Clonetics (Walkersville, MD). Cells were cultured at 37C in humidified 5% CO2 in air. NHBE cells were grown in bronchial epithelial growth medium (BEGM; Clonetics). BEGM consists of bronchial epithelial basal medium (BEBM) supplemented with recombinant human epidermal growth factor (rhEGF), insulin, hydrocortisone, transferrin, tri-iodothyronine, epinephrine, pituitary extract (PE), and retinoic acid. Cells were plated onto glass coverslips for 48 hr or until 75% confluence before fixation and immunofluorescent staining. SCC12 cells, a transformed keratinocyte cell line known to express most if not all hemidesmosomal antigens (Baker et al. 1996a), were used as controls and maintained in keratinocyte serum-free medium, KSF-M (Gibco; Gaithersburg, MD) supplemented with rhEGF and PE.
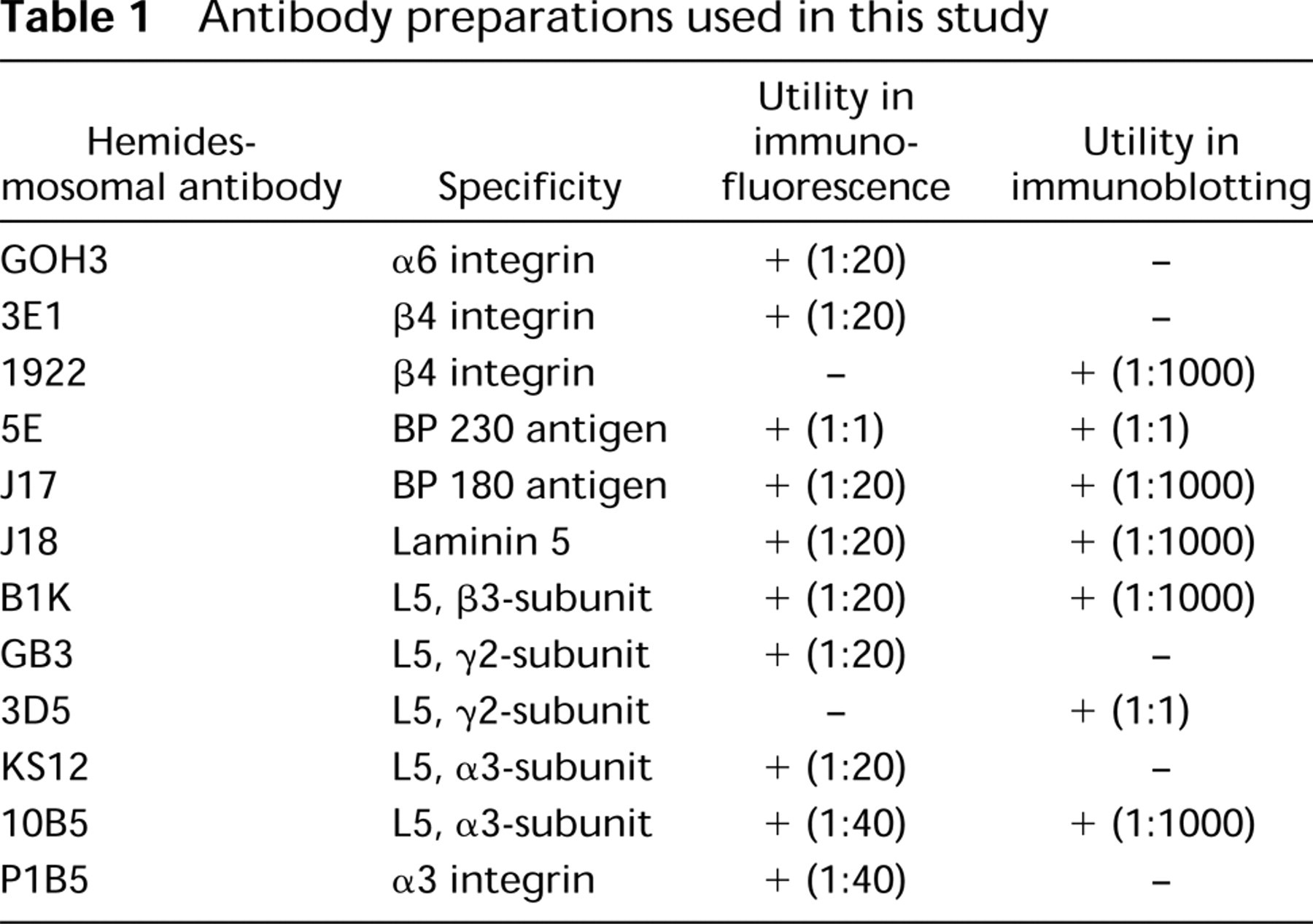
Antibody preparations used in this study
Normal bronchial tissues, surplus pathological specimens removed during staging pneumonectomy, were snap-frozen in liquid N2. Sections of frozen tissue material were prepared on a cryomicrotome and mounted on glass slides in the immunohistochemistry facility at Northwestern Memorial Hospital (Chicago, IL). Some sections were stained with hematoxylin and eosin for specimen orientation purposes, and others were prepared for immunofluorescence analyses (see below).
Antibodies
A description of antibodies is provided in Table 1. In brief, mouse monoclonal antibodies (MAbs) 3E1 and P1B5 (Gibco) and rat MAb GOH3 (Immunotech; Westbrook, ME) against cell surface integrins β4, α3, and α6, respectively, were used in these studies. Integrin β4 antibody AB1922 was purchased from Chemicon (Temecula, CA) for use in immunoblotting studies. For bullous pemphigoid antigens, we used 5E and 10C5A, MAbs against BP230 (BPAG I), whose preparation has been described previously (Hopkinson et al. 1992; Hashimoto et al. 1993). J17, a rabbit polyclonal antibody to BP180 (BPAG II), was also prepared as previously described (Hopkinson et al. 1992) (Table 1).
GB3 antibodies (Harlan Bioproducts for Science; Indianapolis, IN), which recognizes the γ-chain of laminin 5, was used for immunofluorescence microscopy. In addition, KS12 (Desmos; San Diego, CA), an α3-laminin chain-specific mouse MAb, and B1K (clone 17) (Transduction Laboratories; Lexington, KY), a β3-laminin chain-specific antibody, were utilized for immunofluorescence studies. MAb 10B5, an α3-chain-specific antibody (Goldfinger et al. 1998), and MAb 3D5, which recognizes the γ2-laminin chain of the laminin 5 molecule, were used for Western immunoblotting (Jones and Goldfinger, unpublished data). Antibody dilutions used to perform these studies are indicated in Table 1.
Secondary fluorescein-conjugated goat anti-mouse, goat anti-rat, goat anti-rabbit, and goat anti-human antibodies were purchased from Jackson Immunoresearch Laboratories (West Grove, PA). Horseradish peroxidase-conjugated goat anti-mouse, goat anti-rabbit, and goat anti-human secondary antibodies were obtained from the same supplier.
Immunofluorescence Microscopy
Cells on coverslips were rinsed in PBS. If the cells were to be probed with antibodies against the BP antigens or the matrix antibodies, the specimens were placed in acetone for 2 min at −20C and subsequently air-dried. Alternatively, those cells to be processed with integrin antibodies were fixed in 3.7% formaldehyde for 5 min at room temperature, extracted in −20C acetone, and then air-dried. Primary antibodies were overlaid on the cells plated on coverslips that were incubated for 1 hr at 37C. After a 5-min wash in PBS, appropriate secondary antibodies were overlaid on the cells for a second 1-hr incubation. After washes in PBS, the coverslips were mounted on slides in mounting medium and dried in the dark before examination (Hopkinson et al. 1992).
Cryosections of tissue samples were fixed in acetone at −20C for 5 min. After incubation in primary antibody, the slides were gently washed in PBS to prevent disruption of the tissue specimens. After a 1-hr incubation with the secondary antibody, the sections were washed again in PBS, mounted, and dried as described above.
Protein Preparations, Sodium Dodecyl Sulfate-polyacrylamide Gel Electrophoresis, and Western Immunoblotting
HD protein and integrin expression were assessed in whole-cell extracts of cultured cells, whereas laminin 5 expression was determined in ECM preparations. The latter were prepared using a modification of a technique described by Gospodarowicz (1984) (Goldfinger et al. 1998). In brief, cells were removed from their matrix by exposure to 20 mM ammonium hydroxide for 5 min. The matrix was rinsed in distilled, deionized H2O and PBS. The cell monolayers and the ECM preparations were then solubilized in a sample buffer consisting of 8 M urea, 1% sodium dodecyl sulfate (SDS) in 10 mM Tris-HCl, pH 6.8, and 15% β-mercaptoethanol. Proteins in the preparations were separated by SDS-PAGE and transferred to nitrocellulose for immunoblotting (Laemmli 1970). Blots were developed by the chemiluminescence method using the ECL Western blotting system (Amersham; Arlington Heights, IL).
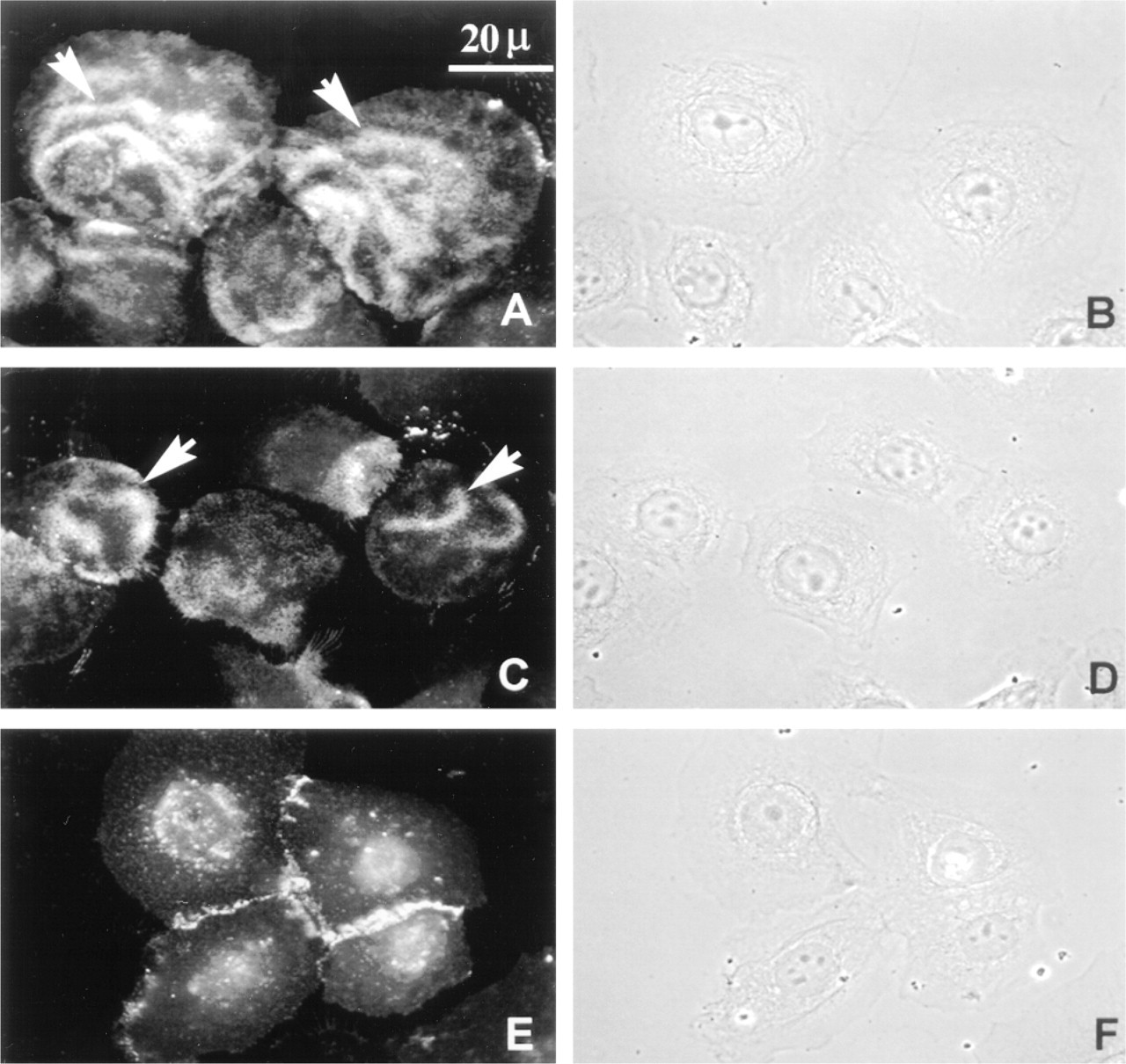
Immunofluorescent localization of the hemidesmosomal integrin heterodimer subunits α6 and β4 in NHBE cells (
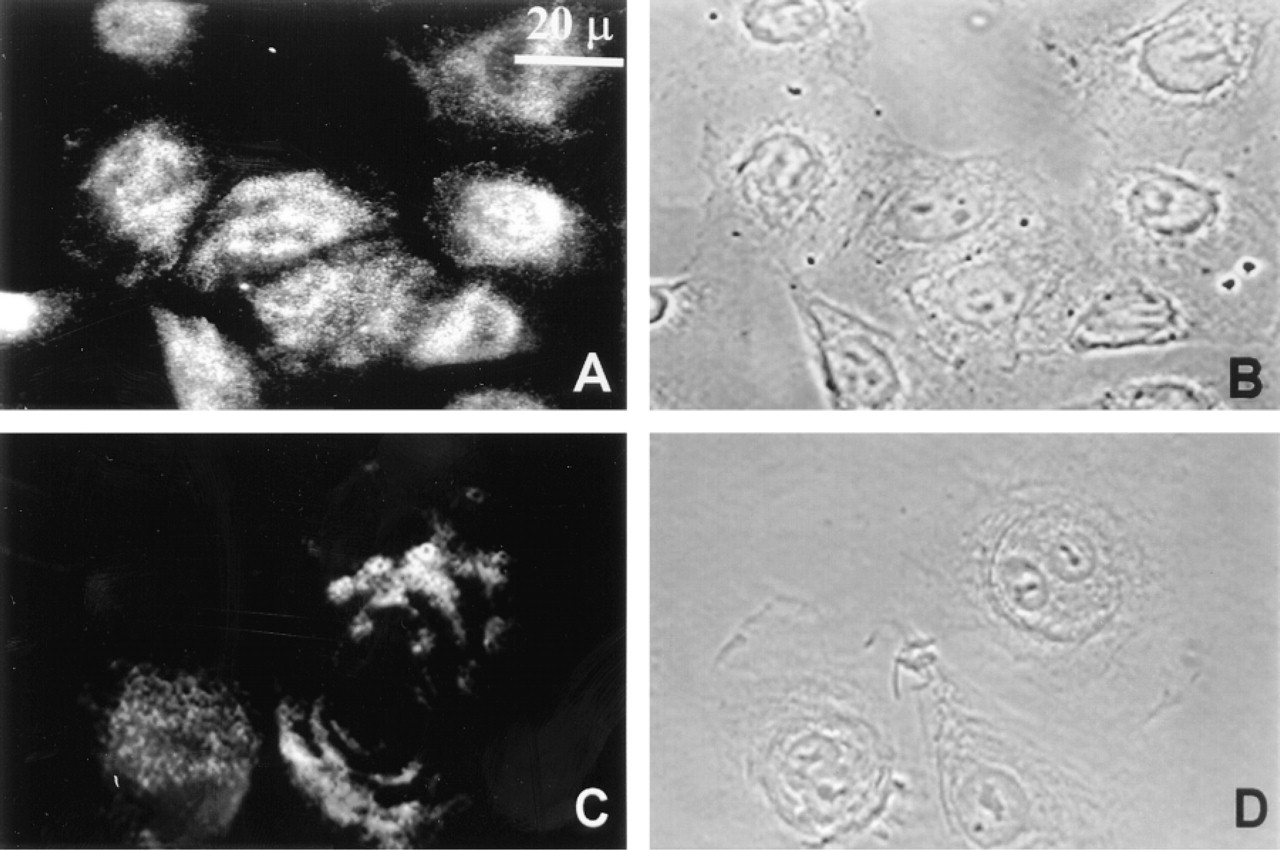
Immunofluorescent localization of the bullous pemphigoid antigens BP180 and BP230 (
Electron Microscopy
Cells maintained on tissue culture plastic until confluent were fixed for 30 min in 1% glutaraldehyde in 0.1 M sodium cacodylate buffer, pH 7.2. After fixation, cells were washed three times in 0.1 M sodium cacodylate buffer and postfixed in 1% OsO4 containing 0.8% potassium ferricyanide. Preparations were then stained with uranyl acetate, dehydrated in ethanol, and embedded in Epon-Araldite resin (Tousimis; Rockville, MD). Thin sections of the embedded material were stained with lead nitrate and sodium citrate and viewed at 60 kV in a JEOL 100CX electron microscope (JEOL USA; Peabody, MA).
Results
Immunofluorescence of HD Proteins in Human Bronchial Epithelial Cells In Vitro
To ascertain whether human bronchial epithelial cells express the HD integrins α6 and β4 and the bullous pemphigoid proteins, we performed immunofluorescence staining, examining NHBE cells grown on glass coverslips. Antibodies 3E1 and GOH3 against β4 and α6, respectively, stained NHBE cells in a “wavy” configuration with basilar localization, as determined by carefully examining the cells at different focal planes from the apical to basal surface (see arrows, Figures 1A and 1C). This pattern was similar to that observed in keratinocytes and was not seen with negative controls, using appropriate secondary antibody alone (not shown) (Baker et al. 1996b). In contrast, the antibody P1B5, which recognizes α3 integrin, localized to the cell-cell contacts (Figure 1E). For each image in which antibody localization is shown, a matched phase-contrast image is presented to demonstrate the cell morphology and provide spatial orientation (Figures 1B and 1D).
Staining for the bullous pemphigoid antigens BP180 and BP230 using antibodies J17 and 5E, respectively, also generated a similar wavy basal staining pattern as seen in keratinocytes (Figures 2A and 2C) (Hopkinson et al. 1992). Corresponding phasecontrast images are presented to illustrate cell morphology (Figures 2B and 2D).
Production of Laminin 5 in Human Bronchial Epithelial Cells In Vitro
We next examined laminin 5 production by NHBE cells. Laminin 5 is a ligand for the α6β4 integrin and is essential for the assembly of HDs in epithelial cells (Jones et al. 1994; Baker et al. 1996b; Borradori and Sonnenberg 1996). Again, we assessed the localization of laminin 5-subunits in NHBE cell cultures using a panel of antibodies specific for the three subunits of laminin 5, α3, β3, and γ2. For these studies, the MAbs KS-12 (α3), B1K (β3), and GB3 (γ2) were used, and all demonstrated significant ECM staining, as well as some intracellular staining (Figures 3A, 3C, and 3E). Phase-contrast images are presented for each fluorescence localization (Figures 3B, 3D, and 3F). It should be noted that the β3 antibody probe generated more intense cytoplasmic staining of NHBE cells compared to the other laminin 5-subunit antibodies (compare Figure 3C with Figures 3A and 3E). This may reflect differences in the ability of our three laminin 5-subunit antibodies to stain cytoplasmic pools of their respective antigens or, possibly, differences in the levels of the laminin 5-subunits in the cytosol.
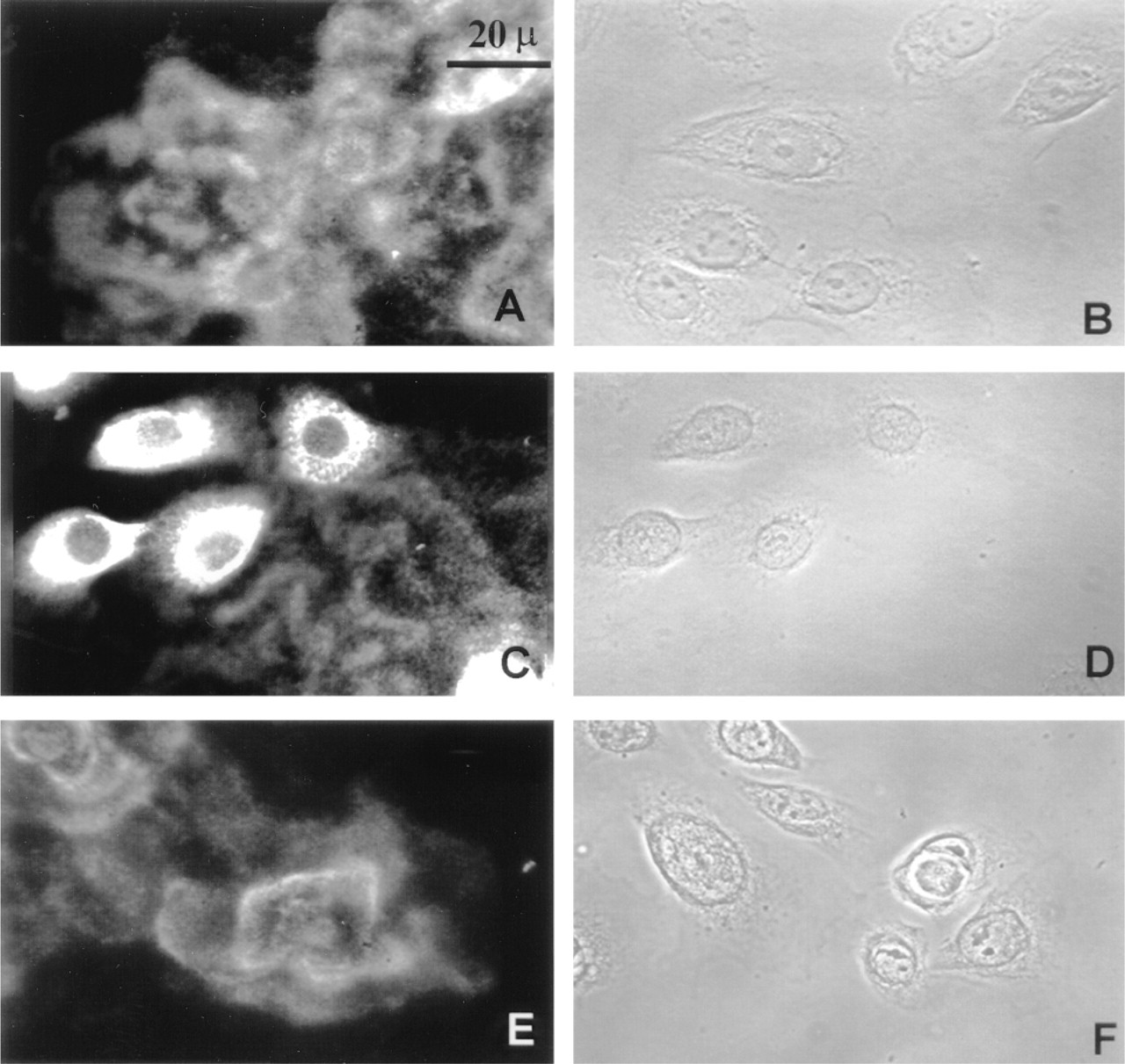
Localization of laminin 5 subunits in NHBE cells is demonstrated in NHBE cells. The immunofluorescent staining pattern of the laminin 5 subunits is demonstrated using MAbs to the specific laminin subunit. (
Immunoblotting of HD Proteins and HD ECM Elements in NHBE Cells In Vitro
To establish the nature of the HD proteins expressed by NHBE cells and to characterize the ECM produced and deposited by this cell type, we performed Western blot analyses. Using antibodies AB1922, J17, and 5E, we determined whether NHBE cells express integrin β4 and the BP antigens (BP180 and BP230) and compared the electrophoretic gel mobility of the NHBE cell antigens with those in the squamous cell carcinoma cell line SCC12 (Figure 4). As demonstrated, both the β4 integrin protein and the two BP proteins were readily expressed by NHBE cells and appeared to migrate identically to their keratinocyte counterparts.
Analysis of the ECM deposited by the NHBE cells was then performed by Western immunoblotting. Antibodies 10B5, B1K, and 3D5, specific for the α3-, β3-, and γ2-subunits, respectively, recognize proteins of 190160 kD, 145 kD, and 155 kD, respectively, in ECM preparations derived from NHBE cells (Figure 5). Species of similar molecular weight were recognized in SCC12 ECM preparations by the same set of antibodies (Figure 5). Whereas the predominant polypeptide recognized by the 10B5 antibody in the ECM of SCC12 cells was 190 kD, the same antibody showed more intense staining of the 160 kD species in NHBE ECM (Goldfinger et al. 1998) (Figure 5).
Electron Microscopy of HDs in Bronchial Epithelial Cells In Vitro
Monolayers of NHBE cells grown under routine cell culture conditions were fixed in glutaraldehyde and prepared for EM as described. Mature HDs complete with dense plates were seen (Figure 6A). At high magnification, each mature HD plaque has a tripartite architecture that is associated with the keratin intermediate filament network (Figure 6B). In Figure 6B, one immature hemidesmosome was observed. This has a less well-defined cytoplasmic plaque than the mature hemidesmosome that lies nearby.
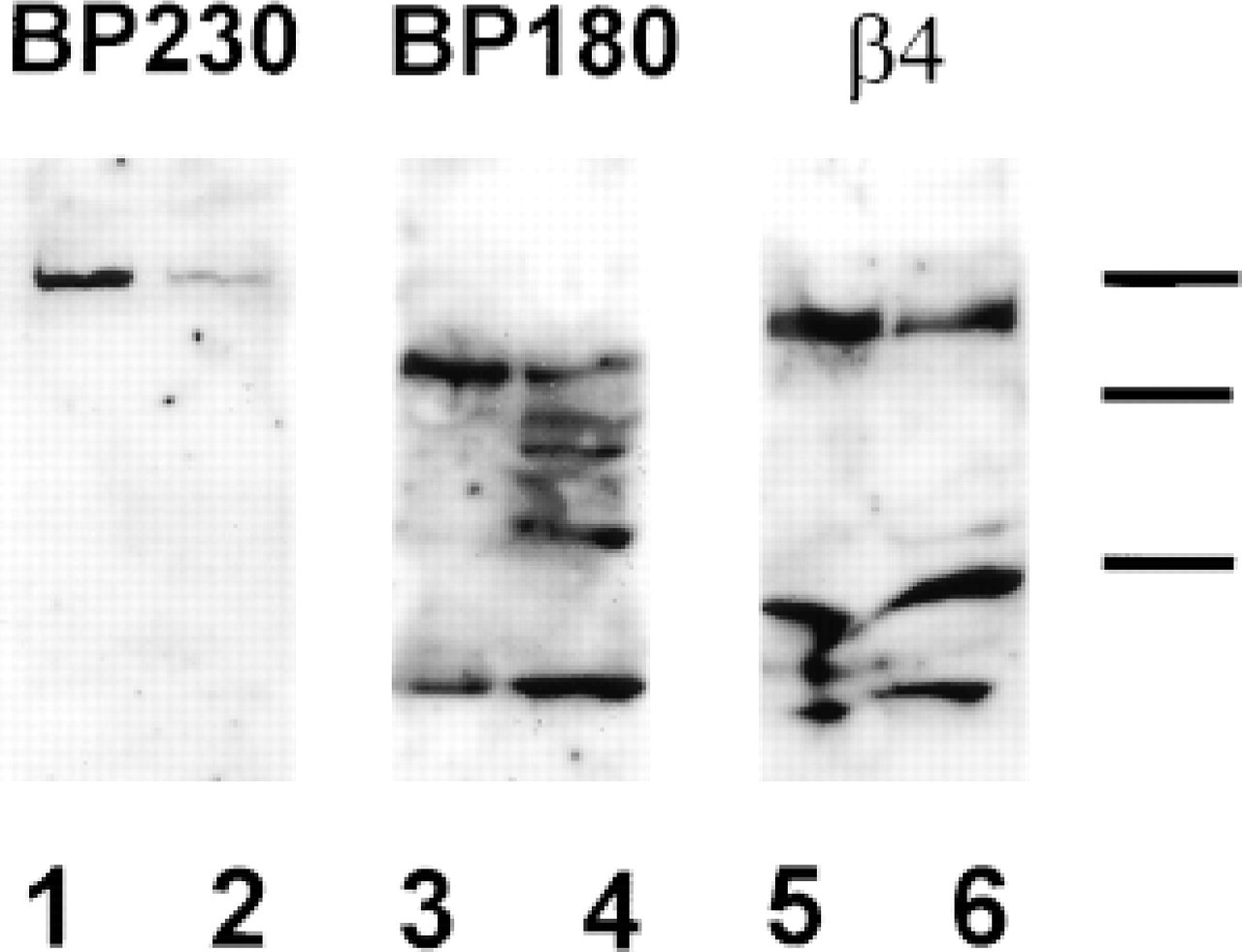
Immunoblots of bronchial epithelial whole-cell extracts. Western analysis of whole-cell extracts of NHBE cells was performed to determine the expression of specific hemidesmosomal components. NHBE cells (Lanes 1, 3, and 5) were contrasted to a keratinocyte control cell line (SCC12) known to form hemidesmosomes and to express all the hemidesmosomal elements (Lanes 2, 4, and 6). Immunoblotting was performed using antibodies against BP230 (Lanes 1 and 2), BP180 (Lanes 3 and 4), and integrin β4 (Lanes 5 and 6). Molecular weight standards of 210, 190 and 120 kD are indicated by dashes. Comparable to the SCC12 cells, antibodies to the β4 integrin recognize an ∼200-kD peptide in the NHBE cells. In addition, the BP180 (J17) and BP230 (5E) recognize polypeptides migrating at 180 and 230, respectively. The low molecular weight species recognized by J17 serum in Lanes 3 and 4 is likely to be the cytoplasmic domain of BP180 which is a product of BP180 proteolysis (Hopkinson et al. 1992). The functional significance of this fragment is unknown at present.
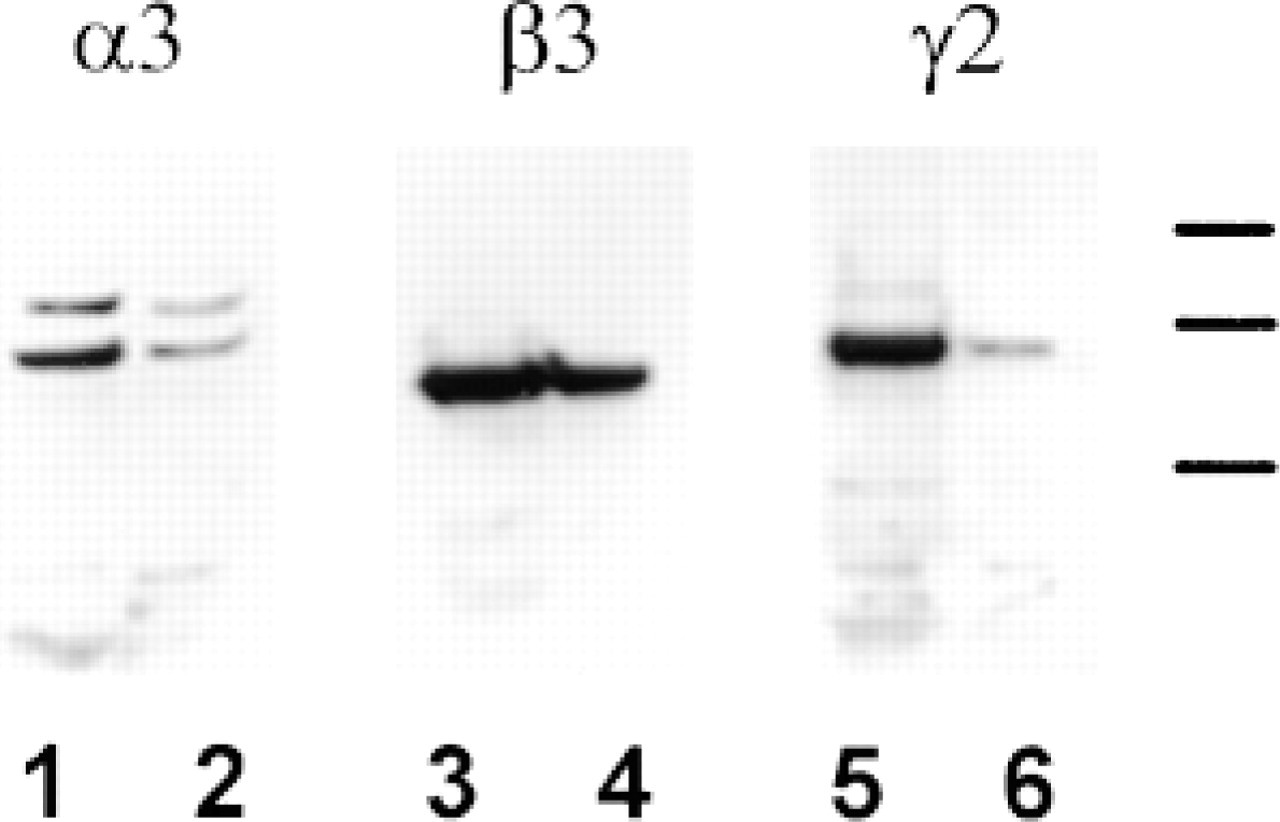
Immunoblots of ECM produced by NHBE cells. Western blot analysis was performed using antibodies against the α3-, β-3, and γ2-subunits of laminin 5. The expression of the specific laminin chains was evaluated in ECM deposited by NHBE cells (Lanes 1, 3, and 5) and compared to that produced by a keratinocyte control cell line (SCC12; Lanes 2, 4, and 6). Molecular weight standards of 210, 190, and 160 kD are indicated by dashes. It should be noted that NHBE cells express both the processed and unprocessed α3-subunit of laminin 5. Compared to SCC12 cells, the processed α3-subunit predominates in whole-cell extracts of NHBE cells.
Expression of Hemidesmosomal Integrins and Laminin 5 in Human Bronchial Biopsy Specimens
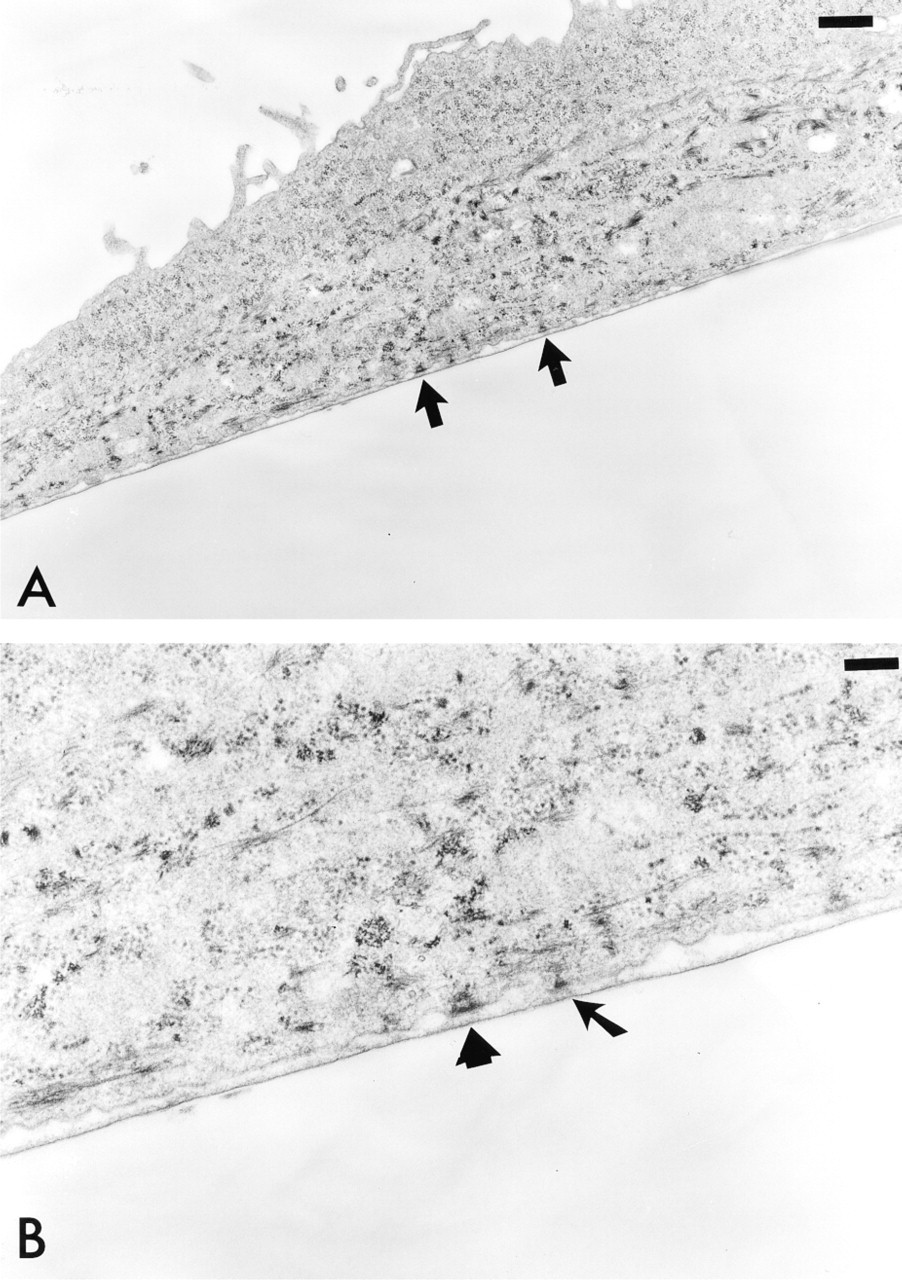
Ultrastructural analysis of NHBE cells. Cells were fixed and processed for EM. Sections were prepared of the fixed cells perpendicular to their substrate attachment. (
To relate the immunocytochemical results we obtained in cultured cells with expression of HD proteins in lung tissue, we next analyzed bronchial tissue specimens by immunofluorescence microscopy. As shown in Figure 7, bronchial biopsy specimens contained a characteristic pseudostratified columnar epithelium, and as seen in the phase-contrast images (Figures 7B, 7D, 7F, 7H, and 7J), the airway epithelium, lumen, and BM were clearly visible. In Figures 7A and 7C, antibodies GOH3 and 3E1 showed strong immunofluorescence staining at the cell-BM interface, confirming the localization of the integrins α6 and β4 to the basal surface of the epithelial cell, with no significant staining at the cell-cell contacts. Similarly, laminin 5 staining, using the GB3 antibody (Figure 7E), was confined to the BM. Like α6 and β4 integrin, the hemidesmosomal proteins BP180 and BP230 were also localized to the epithelial cell-BM interface (Figures 7G and 7I).
Discussion
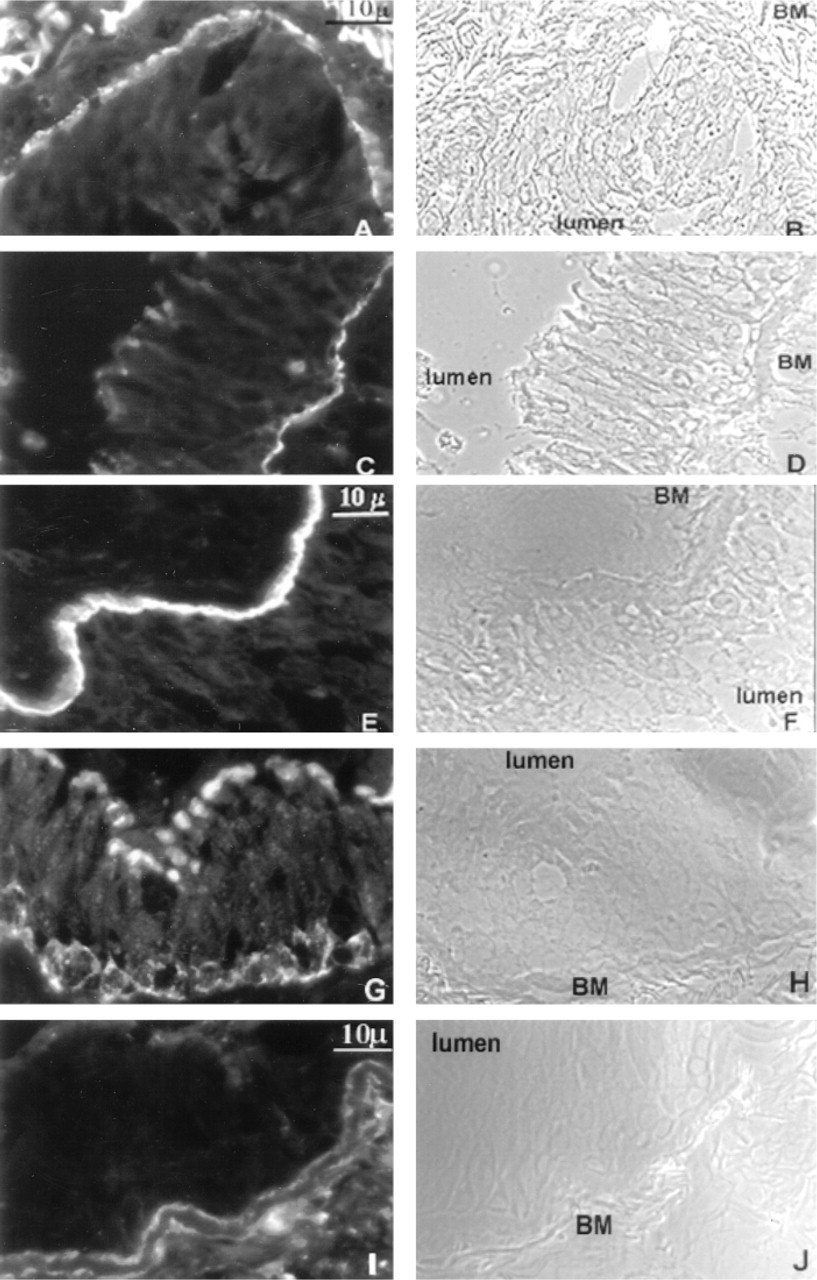
Immunofluorescence staining of the HD proteins and laminin 5 subunits is demonstrated in a cryosection of a bronchial biopsy specimen. (
In previous studies, laminin 1 production by bronchial epithelial cells has been reported in the developing airway preceding the presence of the epithelium (Virtanen et al. 1996; Schuger 1997). In addition, Koukoulis et al. (1997) have identified the laminin binding integrins α6β4 and α3β1 in the distal airways of patients undergoing biopsy for diagnosis of pulmonary carcinoma. However, the expression of laminin isoforms in the BM of airway epithelium has not been studied extensively. Using a panel of antibodies specific for the α3-, β3-, and γ2 laminin-subunits, we have shown by immunoblot analyses that primary human bronchial epithelial cells produce laminin 5 in vitro. Furthermore, utilizing immunofluorescence microscopy on human bronchial sections, we have been able to provide evidence that laminin 5 is a component of the bronchial BM.
In NHBE cells, we see two forms of the α3-subunit of laminin 5 by Western immunoblotting. The larger of the two migrates at 190 kD on SDS-PAGE, whereas the smaller shows a molecular weight of approximately 160 kD. Previous studies have shown that the α3-subunit is first secreted as a 190-kD species and then undergoes cleavage within its G-domain, resulting in the production of a 160-kD α3 isoform (Marinkovich et al. 1992; Goldfinger et al. 1998). Moreover, recent work has suggested that laminin 5 containing the processed α3-subunit is required for the nucleation of HD assembly (Goldfinger et al. 1998). Our results are consistent with these findings in that the matrix of NHBE cells contains predominantly the 160-kD α3 laminin-subunit isoform and these cells assemble bona fide HDs.
In epithelial cells, the two major integrin receptors for laminin 5 are αβ4 and αβ1 (Rabinovitz and Mercurio 1996). In certain keratinocytes and mammary epithelial cells, the heterodimer α3β1 occurs primarily at sites of cell-cell contact, whereas the integrin pair α6β4, a component of HDs, occurs basally (Borradori and Sonnenberg 1996; Stahl et al. 1997; Jones et al. 1998). Our data for bronchial epithelia demonstrate similar localization of these integrins in airway epithelial cells in vitro. Moreover, the distribution of integrins α6 and β4 on the basal surface of NHBE cells is similar to that observed for HD proteins, particularly the bullous pemphigoid antigens BP180 and BP230. Therefore, the HDs of NHBE cells contain all of the components of the classical or Type I HD seen in keratinocytes. This, taken together with the similarity between the appearance of NHBE HDs and epidermal HDs, leads us to propose that airway epithelial cells form the classical type of HD.
Demonstrating bona fide HDs in the airways has a multitude of implications. HDs are considered to be sites of stable cell-ECM anchorage that are involved in the maintenance of tissue integrity (Burgeson and Christiano 1997). For example, in dermatological diseases such as junctional epidermolysis bullosa (JEB), in which there is absence or hypoplasia of HDs, the epidermis blisters because of a separation of keratinocytes from the underlying dermis (Khavari 1998). We assume that, in the lung, one function for HDs is to ensure stable anchorage of airway epithelial cells to the BM. Loss of HDs in airway epithelial cells may explain the development of pulmonary symptoms in patients with acquired and hereditary bullous disorders and may help to explain the high morbidity/mortality associated with certain of the EB subgroups (Shaw et al. 1997; Dabelsteen 1998).
In epithelial cells, HDs have additional roles besides adhesion and morphogenesis (Burgeson and Christiano 1997; Jones et al. 1998). They are also involved in signal transduction via their integrin components (Howe et al. 1998). For example, recent investigations have linked tyrosine phosphorylation and Ras-MAP kinase activation to the ligation of laminin 5 and the epithelial cell integrin α6β4 (Mainiero et al. 1995; Gonzales et al. 1999). Furthermore, an induction of tyrosine phosphorylation has been linked to a known epithelial cell mitogen, EGF (Mainiero et al. 1996). Studies to determine whether the integrins of the HDs of airway epithelial cells play a role in regulating their response to growth factors and ECM signals during development and after lung injury are currently under way.