
Abstract
Antiseptics such as chlorhexidine digluconate (CHX) are widely used in clinical dental practice, but their potential risks, particularly regarding antimicrobial resistance (AMR), are not yet known. This study explores the genomic and transcriptomic mechanisms of CHX adaptation in 3 clinical isolates of Streptococcus spp. and their adapted counterparts. The genomic analysis revealed mutations in genes related to membrane structure, DNA repair, and metabolic processes. Mutations include those in diacylglycerol kinase that occurred in Streptococcus salivarius and the autolysin N-acetylmuramoyl-L-alanine amidase homologues in both Streptococcus mitis and Streptococcus vestibularis, which may contribute to enhanced CHX resistance. Our findings showed stress response genes constantly expressed in all 3 CHX-adapted strains, regardless of acute CHX exposure. Commonly upregulated genes were related to oxidative stress, DNA repair, and metabolic pathway changes, especially amino acid related metabolism. In addition, cell surface restructuring, multiple ABC transporter genes, as well as antimicrobial resistance–associated genes were constitutively expressed. Homologue genes that were significantly upregulated across all 3 species after mutation included recD (DNA repair), potE (amino acid transport), and groEL (stress response). In addition, we saw an increase in a gene associated with the penicillin-binding protein PBP2a in all strains. Beyond these conserved adaptations, we observed species-specific shifts under prolonged CHX exposure. In S. vestibularis, glutathione synthesis genes increased while fatty acid metabolism genes were downregulated. S. salivarius showed elevated expression of genes related to organic anion transport and RNA modification. S. mitis exhibited changes in pyrimidine metabolism, ion homeostasis, and pyruvate dehydrogenase complex genes. Uniquely, S. mitis also showed acute CHX response with upregulation of carbohydrate metabolism and phosphotransferase system genes. These findings highlight the complexity of CHX-induced adaptation, suggesting connections to genetic mutations and emphasizing the need for further research to understand and mitigate AMR risks.
Introduction
Chlorhexidine digluconate (CHX) is widely considered the gold standard antiseptic in dentistry and oral home care (Cieplik et al. 2019; Weber et al. 2023). Numerous clinical studies have well-documented its efficacy in reducing plaque accumulation and gingivitis (Figuero et al. 2019). In addition, it is used as preprocedural mouthwash as part of a series of infection prevention measures in dental practice (Bonn et al. 2023; Weber et al. 2023). Despite the widespread clinical use of CHX in the dental field, there is little awareness among dental professionals regarding the potential risks associated with this widespread use, such as detrimental changes in the oral microbial ecology and antiseptic resistance (Cieplik et al. 2019; Bescos et al. 2020; Weber et al. 2023). A number of recent studies have demonstrated that bacteria can adapt phenotypically in response to repeated exposure to sublethal concentrations of antiseptics in vitro, as evidenced by increases in minimal inhibitory concentrations (MICs) (Wand et al. 2017; Verspecht et al. 2019; Laumen et al. 2021; Schwarz et al. 2021; Auer et al. 2022). Concomitantly, the MICs of several antibiotics also increased, suggesting the development of cross-resistances in the adapted strains (Wand et al. 2017; Laumen et al. 2021). Similarly, a recent study examined the effects of CHX or cetylpyridinium chloride on microcosm biofilms with simulated mouthwashes twice a day over a 7-d period and found antiseptic-resistant phenotypes, which also exhibited resistance to several antibiotics (Mao et al. 2022). Consequently, the selective pressure exerted by antiseptic treatment may result in the emergence of antibiotic-resistant strains within the oral microbiome (Wand et al. 2017; Verspecht et al. 2019; Mao et al. 2022). A recent study employing RNA sequencing to analyze the transcriptomic stress response of a Streptococcus mutans–type strain upon subinhibitory treatment with CHX found significantly differential expression of numerous genes and pathways related to stress response, biofilm formation, and membrane transporters, suggesting a potential link between the antiseptic-driven stress response and development of cross-resistances against antibiotics (Muehler et al. 2022). However, laboratory-type strains may have undergone a loss of certain crucial pathophysiological properties that were previously exclusive to clinical isolates (Fux et al. 2005). In a previous study, we investigated the phenotypic adaptation of clinical isolates of early colonizers in dental plaque toward antiseptics and found that some Streptococcus spp. exhibited up to an 8-fold increase in MICs after 10 passages to subinhibitory concentrations of CHX, accompanied by changes in biofilm-forming ability (Auer et al. 2022). However, the actual mechanisms behind this adaptation on genome and transcriptome level are not clear yet.
Therefore, the aim of the present study was to combine whole-genome sequencing and transcriptome sequencing (RNA-Seq) to investigate the genomic and transcriptomic mechanisms underlying the phenotypic adaptations observed in 3 adapted Streptococcus species from a previous study (Auer et al. 2022) in comparison with their wild-type counterparts.
Material and Methods
Bacterial Strains
Wild-type isolates of oral Streptococcus species were taken from patients with active caries as described previously. The subjects from whom the samples were taken from have not taken antibiotics within 6 mo from sampling. Furthermore, they did not regularly use antiseptic mouthrinses (Anderson et al. 2023). Some of these strains were then described in a second study where they were passaged 10 consecutive times in the presence of subinhibitory concentrations of CHX and then frozen at −80 °C (Auer et al. 2022). These frozen cultures were subsequently used for further CHX exposure, as well as DNA and RNA extraction. The strains selected for the current study, 73/73a (Streptococcus salivarius), 78/78e (Streptococcus vestibularis), and 93/93d (Streptococcus mitis), exhibited the highest increases in MIC (Auer et al. 2022). The study described herein focuses on the genomic and transcriptomic adaptations and responses to CHX.
Preparation and Bioinformatical Analysis
The supplementary file contains information on the bacterial culture and treatment conditions as well as the preparation methods and software used for genomic and transcriptomic data analysis.
Results
Genomic Variants and Mutational Landscape in Chlorhexidine-Adapted Streptococcus Species
We compared the whole genomes of wild-type and CHX-adapted strains for S. salivarius (73A), S. vestibularis (78E), and S. mitis (93D) to identify genetic differences. Genomic variant calling revealed 13 mutations in isolate 73A, 45 in isolate 78E, and 13 in isolate 93D (Supplementary Table 4). Many of these were classified as missense or disruptive. In isolate 73A, mutations affected genes encoding a 50S ribosomal protein L21, diacylglycerol kinase, and a hypothetical protein. Isolate 78E had disruptive mutations in genes encoding N-acetylmuramoyl-L-alanine amidase, transposases, a replicon stabilization protein and a transcriptional accessory protein Tex/SPT6. In isolate 93D, missense variants affected genes encoding an ABC transporter; spermidine/putrescine import ATP-binding protein PotA; glycosyl hydrolase family 25 protein, which is a homologue of N-acetylmuramoyl-L-alanine amidase; and the alpha/beta hydrolase and tRNA 2-thiouridine(34) synthase MnmA. A detailed list of mutations is in Supplementary Table 4.
Transcriptomic Analysis of Streptococcus Strains following Prolonged Chlorhexidine Exposure
To investigate transcription differences between wild-type and long-term CHX-adapted streptococci and the additional effect of acute CHX exposure, we conducted RNA-Seq and differential gene expression analyses across 3 independent experiments comparing CHX-treated and untreated streptococci.
This study is primarily focused on the difference between wild-type and CHX-adapted strains. The response varied across species, with pathways related to cell wall organization enriched in S. mitis, biosynthesis pathways highlighted in S. salivarius, and genes associated with external stimulus response more highly expressed in S. vestibularis (Supplementary Tables 5–7).
Transcriptomic Adaptations of S. vestibularis to Chlorhexidine Exposure
Upon acute chlorhexidine exposure, no genes were differentially expressed in the adapted S. vestibularis strain compared with conditions without acute CHX exposure. However, many stress response genes were already upregulated in the adapted strain compared with the wild-type strain even without additional CHX treatment. Principal component analysis (PCA; Fig. 1A) revealed 66% of the variance in RNA expression in PC1 was due to differences between wild-type and adapted strains, independent of chlorhexidine presence. Chlorhexidine-adapted replicates clustered similarly across conditions. The volcano plot (Fig. 1B) shows gene regulation differences between the wild-type and adapted cells without acute exposure. Of 1,397 significantly differentially expressed genes, the 10 with the lowest P value are labeled. Notably, the top 10 upregulated genes in adapted cells were much less upregulated in wild-type even upon CHX contact (Figs. 1C, D).

Differential gene expression analysis of Streptococcus vestibularis strains before and after adaptation to chlorhexidine exposure (isolate 78). The genotypes of S. vestibularis strains mutated after long-term chlorhexidine stress are represented by blue triangles and wild-type (WT) cells by gray circles. Samples with direct exposure to 0.125 mg/mL chlorhexidine before the measurement are represented by dark blue/dark gray and without exposure in light blue/light gray. (
Gene Set Enrichment Analysis (GSEA) identified significant enrichment comparing wild-type and adapted strains without chlorhexidine exposure in 3 nonredundant gene sets (padj < 0.05; Fig. 1E, Supplementary Table 8). One was linked to the glutathione metabolic process (GO:0006749). Additional stress-related genes include recF (DNA repair), purB (purine biosynthesis), and several ABC transporter system components such as yfiB. Other upregulated genes include those involved in anaerobic metabolism (pflA) and cell surface processes (pksG) including b-lactam resistance–associated pbp2a. Conversely, genes such as fab related to fatty acid metabolism and transmembrane transport (GO:0006631, GO:0055085) were less expressed in the adapted strain (Fig. 1E).
Transcriptomic Adaptations of S. salivarius to Chlorhexidine Exposure
In S. salivarius, no genes were differentially expressed upon acute chlorhexidine exposure in adapted strains; instead, stress response genes were already significantly upregulated without treatment. PCA (Fig. 2A) revealed that 80% of the variance was due to differences between the wild-type and adapted replicates. In total, 829 genes were significantly regulated (padj < 0.05; Fig. 2B).

Differential gene expression analysis of Streptococcus salivarius strains before and after adaptation to chlorhexidine exposure (isolate 73). The genotype of S. salivarius strains mutated after long-term chlorhexidine stress are represented by red triangles and wild-type (WT) cells by gray circles. Samples with direct exposure to 0.125 mg/mL chlorhexidine before the measurement are represented by dark red/dark gray and without exposure in light red/light gray. (
GSEA of wild-type and adapted strains without chlorhexidine exposure identified 18 enriched nonredundant sets from 2,729 Gene Ontology (GO) terms (Appendix Table 9). The most enriched terms (Fig. 2E) were associated with RNA modification (GO:0009451) and the transport of organic anions and acids (GO:0015711, GO:0015849). TopA, a type I DNA topoisomerase, was notably upregulated, along with genes in cell metabolism (the malic enzyme sfcA, xanthine/uracil permease nCS2, transaminase glms), cell wall restructuring (endopeptidase, peptidoglycan associated murA2 and flgJ, and the cell surface protein sdrC), and multidrug resistance mdlB ABC transporter (Figs. 2C, D). Interestingly, signal transduction genes (GO:0035556) regulating environmental responses were expressed less in adapted cells.
Transcriptomic Adaptations of S. mitis to Chlorhexidine Exposure
In S. mitis, unlike the other 2 streptococcal species tested herein, PCA analysis (Fig. 3A) revealed that the wild-type strain showed a remarkably similar expression pattern irrespective of whether the cells were exposed to chlorhexidine or not. Adapted replicates showed clear separation, explaining 14% of the variance. Most variance (66% in PC1) was due to differences between wild-type and adapted strains. A total of 1,123 genes were differentially expressed in the adapted strain compared with the wild-type strain (padj < 0.05; Fig. 3B).

Differential gene expression analysis of Streptococcus mitis strains before and after adaptation to chlorhexidine exposure (isolate 93). The genotype of S. mitis strains mutated after long-term chlorhexidine stress are represented by green triangles and wild-type (WT) cells by gray circles. Samples with direct exposure to 0.125 mg/mL chlorhexidine before the measurement are represented by dark green/dark gray and without exposure in light green/light gray. (
GSEA comparing wild-type and adapted strains without chlorhexidine exposure identified 5 significantly enriched nonredundant GO terms (padj < 0.05; Supplementary Table 10), with the most enriched shown in Figure 3E. They include pyrimidine metabolism pathways, such as UMP biosynthesis (GO:0072528, GO:0006222), involving upregulated genes such as pyrC, pyrK, and pyrDB in adapted strains. Amino acid–associated pathways (GO:1901605) were also enriched, with genes involved in the biosynthesis of polyamine such as nspC or methionine such as O-acetylhomoserine sulfhydrylase. In addition, genes related to ion and cation homeostasis (GO:0098771) were enriched, including a Nramp family member and mgtA. Among the most upregulated genes were galE, part of galactose metabolism, and those encoding pyruvate dehydrogenase complex enzymes (lpdA, pdhA, aceF; Figs. 3C, D).
Gene Expression Changes in Streptococcus mitis following Acute Chlorhexidine Exposure
We examined the differences in gene expression in CHX-adapted S. mitis cells upon acute exposure to CHX. The expression of a total of 439 genes was significantly altered (Fig. 4A). GSEA revealed 37 nonredundant gene sets enriched, with 11 especially expressed under CHX exposure, related to carbohydrate transport and metabolism (GO:0008643) and the polysaccharide biosynthetic process (GO:0000271). Conversely 26 gene sets, many of them containing genes whose expression was upregulated compared with the wild-type strain in the absence of acute exposure to chlorhexidine, were mostly downregulated when the CHX-adapted strain was subject to acute chlorhexidine treatment. These gene sets included pathways for pyrimidine, UMP, organic acid, and anion transport as well as ion transport/homeostasis (Fig. 4B and Supplementary Table 11).

Analysis of gene expression in chlorhexidine-adapted strains of Streptococcus mitis with or without exposure to 0.125 mg/mL chlorhexidine. (
Cross-Strain Analysis of Chlorhexidine Response
Commonalities among all 3 oral streptococcal species tested were identified through a comparative analysis of UniRef50 clusters and clusters of orthologous groups (COG) from significantly upregulated genes (adjusted P value <0.05 and log2FC >0.5). By intersecting UniRef50 clusters, we found that most genes were upregulated in each of the adapted strains compared with their corresponding wild type, independently of acute CHX exposure (Fig. 5A). Only 3 genes, recD, potE, and groEL, were significantly upregulated in all adapted subtypes, linked to DNA damage repair, amino acid transport, and stress response. Clustering by COG terms showed that CHX-adapted cells consistently enriched genes involved in amino acid transport/metabolism (E) and translation/ribosomal structure (J) across all species. S. mitis, however, displayed a shift toward carbohydrate metabolism (G) upon CHX exposure (Fig. 5B). In Figure 5D, we focused on the differences between CHX-adapted strains and wild-type strains without acute exposure to CHX. In total, 10 orthologous genes were upregulated in all species, including ABC-type amino acid transporter such as hisJ or glnQ, genes associated with peptidoglycan crosslinking such as pbp2a, and also ABC-F type ribosomal protection proteins and again groEL involved in protein stability (Appendix Table 12). Using the funcscan nf-core pipeline (including amrfinderplus, rgi, and abricate), a total of 5 antibiotic resistance genes were identified in the genomes of the 3 species. We focused on the difference in expression between wild-type and adapted strains without acute exposure to CHX to discover constitutively higher expressed resistance genes (Fig. 5C). These included genes encoding for efflux pumps associated with macrolide resistance observed for all 3 species as well as a fluoroquinolone resistance gene observed for S. mitis. A tetracycline resistance gene was also noted in S. mitis, but its expression was not found to differ after prolonged CHX adaption.
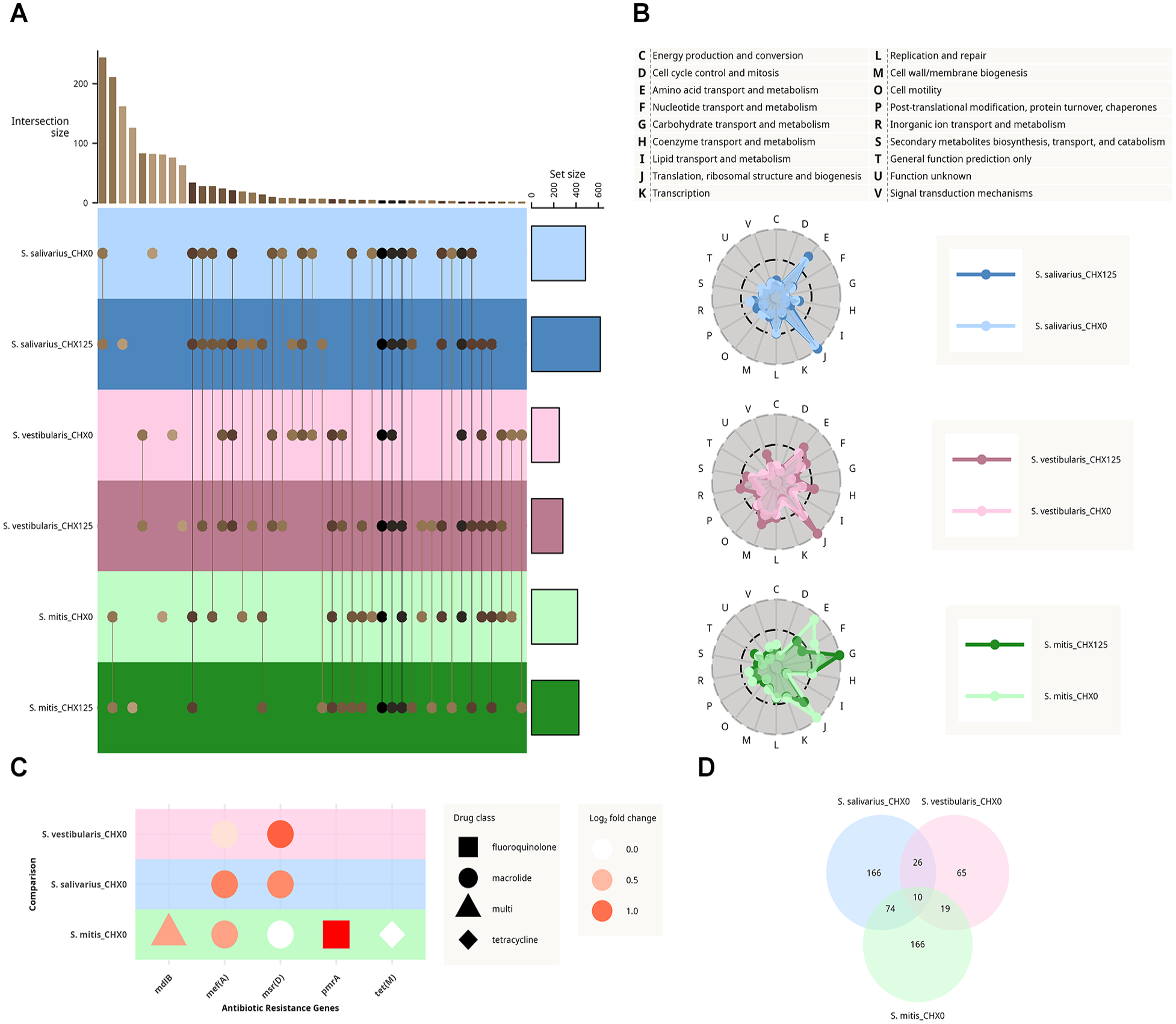
Analysis of the expression pattern in upregulated genes in chlorhexidine-adapted strains compared with wild-type strains from Streptococcus salivarius (blue), Streptococcus vestibularis (red), and Streptococcus mitis (green). The strains were either treated with 0.125 mg/mL chlorhexidine (darker color) or without it (lighter color). Significantly upregulated genes were selected based on the adjusted P value (<0.05) and log2-fold change (>0.5). (
Overall, the upregulation of genes involved in cellular compound biogenesis, cell surface restructuring, DNA/protein damage mitigation, and ABC transporters was observed for each of the 3 CHX-adapted strains compared with their wild-type counterparts.
Discussion
The oral gold standard antiseptic CHX has been linked to resistant bacterial strains (Kampf 2016; Cieplik et al. 2019). Our study focused on the genomic and transcriptomic mechanisms of CHX adaptation in Streptococcus spp., building on previous research that demonstrated increased MICs in certain CHX-adapted clinical isolates (Auer et al. 2022). We selected the 3 Streptococcus species that commonly occur in the oral microbiome based on their pronounced CHX resistance as reported in an earlier study on the same strains (Auer et al. 2022). They are commonly known as commensal bacteria and play a vital role in biofilm formation of dental plaque (Anderson et al. 2023). By working with these clinical isolates rather than laboratory strains, we ensure that our findings more accurately mirror real-world conditions to gain clinically relevant insights into CHX adaptation.
CHX exposure has been shown to affect cell homeostasis leading to oxidative stress and ultimately damage to DNA and proteins (Rzycki et al. 2021). We prepared our study according to Muehler et al. (2022). Acknowledging that our sampling time points may have limited the detection of immediate transcriptional changes during acute CHX exposure, our primary focus remained on the longer-term adaptive responses, including constitutively expressed genes, that persist following prolonged exposure. This may provide at least 1 reason why most genes associated with stress response are upregulated in CHX-adapted strains, regardless of subsequent CHX exposure.
An adaptation observed in all cells was the upregulation of DNA and protein damage repair and prevention mechanisms. All adapted strains showed increased expression of genes from the RecBCD pathway, especially recD. Protein damage prevention included groEL upregulation across all strains with mutations after CHX exposure, consistent with findings in S. mutans (Muehler et al. 2022) and Caulobacter crescentus (Susin et al. 2006).
We saw an upregulation of amino acid synthesis and metabolism-related genes in all species. These included polyamine metabolism and antioxidants such as methionine or glutathione in S. mitis and S. vestibularis. Polyamine deficiency increased ROS and oxidative stress in Streptococcus pneumoniae (Nakamya et al. 2021), and glutathione and methionine acted as antioxidants in various organisms (Potter et al. 2012; Campbell et al. 2016). In S. salivarius, branched amino acid transport and synthesis genes were prominent. They have been shown to respond to acid stress (Santiago et al. 2012). In general, amino acids such as histidine, glutamate, and alpha-ketoglutarate have been associated with hydrogen peroxide–induced oxidative stress survival (Kajfasz et al. 2017).
In a previous study by Auer et al. (2022) that examined the same strains as those used in the current investigation, resistance to the macrolide erythromycin was observed in both the wild-type and the mutants. In our study, we confirmed the presence of macrolide resistance genes in these genomes. However, although we detected increased expression of these genes in the CHX-adapted strains compared with their wild-type counterparts, this elevated expression did not align with the increased susceptibility to macrolides following long-term CHX exposure that Auer et al. (2022) described.
Nonetheless, Auer et al. (2022) also noted a slight decrease in susceptibility to b-lactam antibiotics in the CHX-adapted strains. They observed that for S. mitis, prolonged CHX exposure shifted its response to ampicillin and amoxicillin from fully susceptible to intermediate resistance. In the current study, we observed mutations in an ABC transporter in S. mitis and in the N-acetylmuramoyl-L-alanine amidase gene described later in detail in both S. mitis and S. vestibularis. A previous study found that the deletion of an ABC transporter led to increased resistance to oxidative stress, biofilm formation, and resistance to b-lactam antibiotics in S. mutans (Liu et al. 2017). In our study, we also found the pbp2a gene that was upregulated in all adapted cells compared with the wild-type isolate. It has been shown to increase the resistance against specific b-lactam antibiotics such as ampicillin in Staphylococcus aureus (Fergestad et al. 2020). In combination, these results underscore the importance of exercising caution with CHX treatments, as they may contribute to the increase in susceptibility to some antibiotics but can also lead to cross-resistance after long-term exposure to CHX.
The transcriptome in S. mitis uniquely showed pronounced changes upon CHX contact. CHX is known to reduce phosphotransferase system (PTS) activity in Streptococcus strains, particularly in S. mitis (Marsh et al. 1983). Upon CHX exposure, we observed that PTS-related genes were highly upregulated. In addition, S. mitis showed a significant increase of carbohydrate transport genes essential for sugar uptake and metabolism, which could potentially lead to a formation of acids relevant for caries development.
In the genomic variant analyses of the 3 CHX-adapted Streptococcus spp. assessed herein, mutations were identified in genes related to membrane structure, DNA repair, and metabolic processes.
Mutations in a diacylglycerol kinase homolog in S. mutans had previously been identified after prolonged exposure to CHX with the mutant strains showing reduced susceptibility to CHX (Kaspar et al. 2019). Other researchers observed a reduced capability to react to environmental changes and reduced production of bactericides in S. mutans (Yamashita et al. 1993; Chen et al. 1998). In our current study, we saw a downregulation of expression of signal transduction genes in the adapted strain of S. salivarius, which was found upon genomic sequencing to bear a mutation in the diacylglycerol kinase.
In S. vestibularis, mutations were found in the transcriptional accessory protein Tex/SPT6. The knockout of the tex gene was shown to decrease the ability to adapt to the environment in S. pneumoniae (He et al. 2006). A tex homolog was upregulated in Burkholderia pseudomallei under oxidative stress conditions (Jitprasutwit et al. 2014).
In CHX-adapted S. mitis, we identified mutations in the polyamine transporter potA. The expression of this mutated gene was significantly upregulated in mutated strains. The potABCD system is pivotal for the oxidative stress response in prokaryotes (Bontemps-Gallo et al. 2018; Nakamya et al. 2021).
Both S. mitis and S. vestibularis showed a mutation in the same gene cluster (UniRef50_A0A4V6IXY4) encoding an N-acetylmuramoyl-L-alanine amidase of the glycosyl hydrolase family 25, crucial for cell separation and autolysis. Similar enzymes have been linked to autolysis upon contact with antibiotics (Heidrich et al. 2001) and increased resistance (Diaz Caballero et al. 2018). Auer et al. (2022) reported increased biofilm-forming capacity in both strains. Other autolysins such as atlA or lytC have been shown to be crucial for biofilm formation in streptococci (Ahn and Burne 2006), hinting at a possible connection between the mutation observed in this study and the increased biofilm formation in these CHX-adapted strains reported earlier (Auer et al. 2022). Autolysins also play a role in horizontal gene transfer, promoting increased eDNA levels in biofilms (Moscoso and Claverys 2004). Streptococci in general and especially S. mitis have been known to exchange genetic material with other species including pathogenic species such as S. pneumoniae (Roberts and Kreth 2014; Kalizang’oma et al. 2021). Further research is needed to understand how the adaption to CHX could contribute to horizontal gene transfer and the spread of resistance traits.
Across all 3 species evaluated, conserved upregulation of genes involved in amino acid transport/metabolism, DNA repair, and protein stability underscores a shared adaptive framework within this subset of the Streptococcus genus. Most mutations observed in this study are related to survivability, cell surface composition, and oxidative stress resistance. One notable mutation occurred in 2 species independently in the cell surface–associated autolysin N-acetylmuramoyl-L-alanine amidase in S. mitis and S. vestibularis. At the same time, we observed species-specific shifts such as the reaction to acute CHX exposure in S. mitis and only partial overlaps in upregulated genes in all 3 species. Understanding the mechanisms in greater detail is key to learning more about common concepts and addressing CHX-related antimicrobial resistance in oral streptococci as well as other organisms. Further research is needed to explore the observation of both genetic and transcriptomic changes linked to adaptation to chronic antiseptic usage and to identify potential targets of therapeutic intervention to stem the growing problem of antimicrobial resistance.
Author Contributions
B. Daller, contributed to conception, design, data analysis and interpretation, drafted and critically the revised manuscript; D.L. Auer, W. Buchalla, S. Bartsch, contributed to acquisition, critically the revised manuscript; A. Gessner, contributed to conception, design, critically the revised manuscript; N.S. Jakubovics, A. Al-Ahmad, contributed to conception, critically the revised manuscript; A. Hiergeist, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically the revised manuscript; F. Cieplik, contributed to conception, design, data acquisition and interpretation, drafted and critically the revised manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345251320912 – Supplemental material for Genomic and Transcriptomic Adaptation to Chlorhexidine in Streptococcus spp.
Supplemental material, sj-docx-1-jdr-10.1177_00220345251320912 for Genomic and Transcriptomic Adaptation to Chlorhexidine in Streptococcus spp. by B. Daller, D.L. Auer, W. Buchalla, S. Bartsch, A. Gessner, N.S. Jakubovics, A. Al-Ahmad, A. Hiergeist and F. Cieplik in Journal of Dental Research
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Deutsche Forschungsgemeinschaft (grants CI 263/3-1 and AL 1179/4-1) and by the Bavarian Ministry of Science and the Arts in the framework of the Bavarian Research Network “New Strategies against Multi-Resistant Pathogens by Means of Digital Networking-bayresq.net,” Förderkennzeichen: Kap. 1528 TG 83.
Data and Code Availability Statement
Genomic data have been deposited in the Sequence Read Archive (SRA) and is accessible under project accession number PRJNA1158744. In addition, raw RNA-seq files and TPM (transcripts per million) values of annotated genes are available through GEO under accession number GSE277379.
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.